

Почвоведение, 2021, № 7, стр. 815-826
Оценка температурной чувствительности эмиссии СО2 с поверхности торфяных почв севера Западной Сибири методом трансплантации почвенных монолитов
Г. В. Матышак a, М. О. Тархов a, *, И. М. Рыжова a, О. Ю. Гончарова a, А. Р. Сефилян a, С. В. Чуванов a, Д. Г. Петров b
a МГУ им. М.В. Ломоносова
119991 Москва, Ленинские горы, 1, Россия
b Институт географии РАН
119017 Москва, Старомонетный пер., 29, Россия
* E-mail: tarkhov.mo@gmail.com
Поступила в редакцию 28.08.2020
После доработки 04.12.2020
Принята к публикации 28.12.2020
Аннотация
Торфяные почвы криолитиозоны играют важную роль в глобальном цикле углерода. В условиях прогнозируемых климатических изменений торфяные почвы могут функционировать как значительный источник выбросов парниковых газов в атмосферу. В четырехлетнем полевом эксперименте методом трансплантации (переноса почвенных монолитов торфяного горизонта высотой 20 см, диаметром 10 см) оценена температурная чувствительность эмиссии СО2 с поверхности торфяных почв севера Западной Сибири (Надымский район, зона распространения многолетнемерзлых пород) в условиях существенно более высоких температур функционирования. Увеличение температуры на 7°С обусловило значимый положительный отклик эмиссии СО2 (оцененный методом закрытых камер) с поверхности трансплантированных образцов по сравнению с контрольными – от 30 до 70%. Температурная зависимость эмиссии СО2 с поверхности трансплантированных образцов почв была наиболее выражена в условиях максимальной контрастности термического режима в первые 2 года наблюдений (R2 = 0.8), в последующие годы теснота связи между эмиссией СО2 и температурой становилась слабее. Напротив, температурная чувствительность эмиссии СО2 с поверхности трансплантированных образцов почв для большинства периодов наблюдений характеризовалась высокими значениями коэффициента Q10 (3–6), что указывает на сохранение повышенной скорости минерализации органического вещества торфяных почв криолитозоны в течение продолжительного периода (четырех лет). Полученные результаты могут быть полезны для уточнения прогнозных данных углеродного баланса исследуемой территории, учитывающих вклад органогенных почв криолитозоны.
ВВЕДЕНИЕ
Почвы криолитозоны – важный экосистемный маркер прогнозируемых климатических изменений, которые могут оказать значительное влияние на углеродный баланс экосистем высоких широт [36]. Запасы органического углерода в почвах Земли в метровой толще оцениваются в размере 1500 Пг С, существенно превышая его запасы в атмосфере (800 Пг С) и в наземной фитомассе (500 Пг С) [17]. Почвы криолитозоны, в случае прогнозируемого потепления климата, будут играть значительную роль в регулировании углеродного баланса, так как на их долю приходится до 1/3 от величины общемирового запаса углерода в почвах Земли [16, 22, 36], преимущественно содержащегося в многолетнемерзлых горизонтах почв при значимом вкладе торфяных почв криолитозоны [44]. На сегодняшний момент получено недостаточное количество полевых экспериментальных данных, с помощью которых можно было бы с достаточной степенью точности прогнозировать влияние меняющихся климатических параметров на интенсивность минерализации органического вещества почв криолитозоны и их температурную чувствительность.
В современных исследованиях используются различные полевые методы для изучения последствий нагревания почв. В обзорной работе [12] обсуждаются 4 основных метода полевого нагревания, условно подразделяемых на активные и пассивные. Активные методы включают прогревание с использованием нагревающих кабелей (Heat-resistance cables) и прогревание с использованием инфракрасных подвесных ламп (Overhead IR lamps). К пассивным методам относятся пассивное ночное прогревание с использованием ИК-отражающих материалов (Passive nighttime warming) и пассивное прогревание поверхности почв с использованием трапециевидных конструкций с открытым верхом (Open-top chamber). Активно применяют и другие методы: прогревание поверхности почв с использованием парника (Greenhouse) [21], зимнее снегозадержание (Snow Fence) [29, 43], а также менее распространенный метод трансплантации почвенных монолитов, примененный в данном исследовании [34].
По сравнению с прочими методами полевого прогревания трансплантация является менее популярным, но не менее эффективным вариантом количественной оценки скорости изменения эмиссии СО2 с поверхности почв (температурной чувствительности) в полевых условиях при повышении температуры. Трансплантация заключается в переносе образцов почв ненарушенного сложения между биомами, отличающимися по значениям параметров функционирования: годовой температурой воздуха, температурой почвы, влажностью почвы и др. [34]. Основные плюсы метода – прогревание почв в естественных условиях, без внешних воздействий; возможность прямых измерений эмиссии СО2 в образцах ненарушенного строения; доступность и низкая стоимость оборудования. Трансплантация часто используется для биомов, в которых температура поверхности почв и среднегодовая температура воздуха отличается в среднем на 2–4.5°С или более существенно [38, 40]. Подобный контраст температур позволяет оценить первичный отклик эмиссии СО2 с поверхности почв на нехарактерные для них температуры, а также адаптивную способность почв к контрастным изменениям термического режима.
Последние исследования в криолитозоне демонстрируют однозначный положительный отклик эмиссии СО2 с поверхности почв на экспериментальное прогревание. Так, двухлетний эксперимент с применением методов зимнего снегозадержания и пассивного летнего прогревания поверхности почв с использованием трапециевидных конструкций с открытым верхом обусловил значимый положительный (+35%) отклик эмиссии СО2 с поверхности почв арктической тундры [31]. Положительный отклик эмиссии СО2 также получен по результатам долговременного пассивного прогревания поверхности почв в невегетационный период на Шпицбергене [32]. С использованием метода трансплантации по результатам трехлетнего эксперимента установлено увеличение интенсивности дыхания прогретых почв тундровых областей Канады на 27–60% по сравнению с контрольными участками [38]. Тем не менее, полученных полевых данных по-прежнему недостаточно для достоверных прогнозов отклика скорости минерализации органического вещества почв криолитозоны в условиях климатических изменений.
Цель исследования – количественная оценка отклика эмиссии СО2 с поверхности торфяных почв криолитозоны при увеличении температуры в полевых условиях методом трансплантации.
Задачи исследования включали: 1) оценку температурной чувствительности эмиссии СО2 с поверхности торфяных почв криолитозоны в полевых условиях; 2) апробацию метода трансплантации монолитов торфяного горизонта для изучения скорости отклика эмиссии СО2 с поверхности торфяных почв криолитозоны при увеличении температуры.
ОБЪЕКТЫ И МЕТОДЫ
Район исследования и почвы. Район исследования – север Западной Сибири (Надымский район, Тюменская область, ЯНАО, 65°20′ N, 72°55′ E), граница распространения северо-таежных и лесотундровых территорий в краевой части третьей озерно-аллювиальной равнины р. Надым, междуречье р. Хейгияха – Левая Хетта. Для района исследований в связи с высокой заозеренностью и заболоченностью наиболее характерны гидроморфные и полугидроморфные ландшафты, представленные как современными болотными экосистемами на переувлажненных территориях (термокарстовые депрессии, ложбины стоков, мочажины), так и специфическими криогенными ландшафтами – бугристыми (плоско- и крупнобугристыми) торфяниками различного возраста, формирующимися при активном влиянии многолетнемерзлых пород (ММП), залегающих в пределах 1–2 м [2]. Растительный покров преимущественно представлен кустарничково-багульниково-лишайниковыми и кустарничково-багульниково-сфагновыми сообществами. Почвообразующие породы в большинстве случаев представлены озерно-аллювиальными песчаными отложениями [6].
Исследования проводили на торфяно-криоземах (Turbic Histic Cryosol (Dystric)) (WRB 2014) – типичных почвах бугристых торфяников, наиболее характерных экосистем исследуемой территории [4]. Торфяно-криоземы функционируют в условиях слабоотрицательных среднегодовых температур профиля и активно идущих криогенных процессов. Близость расположения кровли ММП, залегающих в пределах 1 м, обусловливает высокую неоднородность свойств торфяно-криоземов, выражающуюся в чередовании торфяных горизонтов разного типа, вихреватых границах, сильном варьировании мощностей и инверсии горизонтов в результате морозобойного растрескивания, пучения и криотурбаций. Мощность органопрофиля, как правило, изменяется в пределах 0.5 м, и включает 2–3 торфяных горизонта различного ботанического состава и степени разложения, характеризующих современные условия развития торфяника. Более подробное описание торфяно-криоземов есть в наших предыдущих публикациях [7, 10].
Методы исследования. Организация эксперимента, метод трансплантации. Температурная зависимость и температурная чувствительность эмиссии СО2 с поверхности торфяных почв криолитозоны изучены в полевых условиях методом трансплантации в августе 2016–2019 гг. Под температурной зависимостью понимается зависимость отклика эмиссии СО2 с поверхности почв (R) от температуры (T), тогда как температурная чувствительность означает скорость изменения отклика эмиссии СО2 с поверхности почв по отношению к изменению температуры (dR/dT) [33].
В работе применен вариант трансплантации в рамках одной и той же подзоны (северная тайга). Для эксперимента были определены “холодный” и “теплый” участки бугристого торфяника. Участки были выбраны на основе анализа значений глубины сезонного протаивания почв, среднемесячных температур почв в слое 0–10 см и объемной влажности почв в слое 0–20 см в пик вегетационного периода (август) (табл. 1).
Таблица 1.
Гидротермические параметры торфяных почв на контрольных участках в пик вегетационного сезона (среднее за август 2016–2019 гг. ± SE, здесь и далее жирным шрифтом выделены значимые различия)
| Контроль | Почва | Глубина сезонного протаивания, см | Температура почв в слое 0–10 см, °С | Объемная влажность почв в слое 0–20 см, % |
|---|---|---|---|---|
| Холодный участок | Торфяно-криозем типичный | 40 ± 3 (n = 10) |
4.3 ± 0.9 (n = 71) |
38 ± 2 (n = 71) |
| Теплый участок | Торфяная олиготрофная деструктивная мерзлотная | 80 ± 3 (n = 10) |
11.0 ± 0.6 (n = 70) |
34 ± 2 (n = 70) |
В качестве холодного участка выбран типичный участок бугристого торфяника с морошково-багульниково-сфагново-лишайниковым растительным покровом и типичным торфяно-криоземом (ТО–Т1–Т2–СR). В качестве теплого участка выбран деградирующий участок бугристого торфяника с угнетенным морошково–багульниково–сфагново–лишайниковым растительным покровом и торфяной олиготрофной деструктивной мерзлотной почвой (ТОmd–T1md–T2md–T3). Средние значения параметров функционирования изученных почв и общие свойства образцов почв участков представлены в табл. 1 и 2 для горизонтов Т1 (в соответствии с типом трансплантированного горизонта).
Таблица 2.
Физические и химические свойства образцов торфяных почв (среднее ±95%-й доверительный интервал среднего, n = 3)
| Контроль | Почва | Гори-зонт | Степень разложения | Влажность, % от массы | ${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$ | Сырая зола | Собщ | Nобщ |
|---|---|---|---|---|---|---|---|---|
| % | ||||||||
| Холодный участок | Торфяно-криозем типичный | Т1 | 4 | 853 | 4.3 ± 0.3 | 8.1 ± 4.7 | 43.6 ± 3.3 | 1.0 ± 0.6 |
| Теплый участок | Торфяная олиготрофная деструктивная мерзлотная | Т1 | 8 | 313 | 3.8 ± 0.5 | 6.0 ± 2.2 | 49.2 ± 3.1 | 2.4 ± 0.4 |
Монолиты торфяно-криозема с наиболее типичным горизонтом Т1 трансплантировали из холодного в теплый участок бугристого торфяника в полипропиленовых трубах высотой 20 см и диаметром 10 см (мезокосмы) (n = 5). Расстояние между участками по прямой составляло 840 м. Пустые трубы аккуратно врезали в почву холодного участка и, заполненные почвенной массой, в тот же день вынимали обратно для трансплантации в теплый участок. В теплом участке заранее готовили пространство, идентичное размеру мезокосмов, в него опускали мезокосмы с трансплантированной почвенной массой из холодного участка. Расстояние между повторностями мезокосмов составляло 15 см. На холодном и теплом участках также устанавливали контрольные мезокосмы: пустые трубы аккуратно врезали в почву участков на расстоянии 40 см от мезокосмов с трансплантированной почвой.
Далее образцы торфяно-криозема на холодном контрольном участке бугристого торфяника будут наименоваться как образцы холодного контрольного участка; контрольные образцы торфяной олиготрофной деструктивной мерзлотной почвы на теплом участке бугристого торфяника – образцы теплого контрольного участка; образцы торфяно-криозема, трансплантированные из холодного в теплый участок бугристого торфяника – трансплантированные образцы.
Определение параметров функционирования почв. В контрольных и трансплантированных мезокосмах изучали основные параметры функционирования почв: эмиссию СО2, температурный режим, объемную влажность. Эмиссию СО2 в мезокосмах ежедневно измеряли методом закрытых камер в пятикратной повторности [3, 9]. В качестве закрытых камер использовали стальные камеры объемом 850 мл с герметичным клапаном для отбора проб воздуха с помощью шприца (10 мл). Подробно примененная методика описана ранее [1].
Для усиления герметичности камеры использовали пластиковые основания диаметром 10 см и высотой 5 см с желобом для установки камеры. Основания предварительно устанавливали в мезокосм на глубину 3 см. В желоб оснований приливали воду для уменьшения диффузии и возможных газообразных потерь. Концентрацию СО2 определяли на портативном инфракрасном газоанализаторе RMT DX6210 после 10 мин экспозиции (точность 0.002%). Измерения проводили через 2–3 дня после осуществления трансплантации – данная задержка необходима для нивелирования механических воздействий от трансплантации и для стабилизации выделения СО2 трансплантированными образцами [38]. Эмиссию СО2 рассчитывали по формуле [9]:
где 3.18 – коэффициент, учитывающий числовые значения входящих в формулу констант (газовая постоянная и молярная масса) и соотношения размерностей (Па/кПа, час/мин, м/см, мг/г, %/ppm), ΔС – изменение концентрации газа в камере (ppm), Р – барометрическое (атмосферное) давление (кПа), h – высота камеры от поверхности почвы (см), T, °С – температура по шкале Цельсия, Δt – время экспозиции камеры в минутах. Результаты выражали в мг CO2/(м2 ч).Температурный режим изучали с использованием термозондов Hanna Instruments (HI 98509, точность ±0.2°С) и логгеров Thermochron iButtonTM (точность ±1°С). Термозондами в августе 2016–2019 гг. ежедневно в трехкратной повторности проводили измерения температуры почв в мезокосмах в слое 0–10 см в градусах Цельсия. Логгеры использовали для оценки годовой (август 2017–июль 2018) динамики температур в мезокосмах на глубине 10 см в градусах Цельсия в двукратной повторности.
Объемную влажность почв (W) определяли потенциометрически с использованием полевого влагомера FieldScout TDR 100 (Spectrum) (точность ±3%) в слое 0–20 см в процентах в трехкратной повторности.
Глубину сезонного протаивания (мощность сезонно-талого слоя ММП) почв в см определяли методом зондирования щупом по ГОСТ 26262-2014.
Определение физических и химических свойств почв. Степень разложения образцов торфяных почв определяли в полевых условиях по методике фон Поста [25]. Зольность (содержание сырой золы) и ${\text{p}}{{{\text{H}}}_{{{{{\text{H}}}_{{\text{2}}}}{\text{O}}}}}$ (соотношение торф : раствор 1 : 25) оценивали по руководству Воробьевой [11]. Массовую влажность определяли термогравиметрически на приборе OHAUS MB 35. Общее содержание углерода (Собщ) и азота (Nобщ) определяли на элементном CNHS-анализаторе Element Analyzer Vario EL III фирмы Elementar. Все исследования проводили в трехкратной повторности для каждого из образцов.
Количественная оценка температурной чувствительности эмиссии СО2 с поверхности почв. Коэффициент температурной чувствительности Q10, отражающий скорость изменения отклика эмиссии СО2 с поверхности почв при увеличении температуры на 10°С, рассчитывали двумя способами: 1) по формуле [20, 37]:
(2)
${{Q}_{{10}}} = ~{{\frac{{R2}}{{R1}}}^{{{{10} \mathord{\left/ {\vphantom {{10} {\left( {T2 - T1} \right)}}} \right. \kern-0em} {\left( {T2 - T1} \right)}}}}},$2) с использованием уравнения экспоненциальной зависимости эмиссии СО2 от температуры почв по формуле (3) [23, 39]
где R – величины эмиссии СО2 для контрольных мезокосмов холодного участка и мезокосмов, трансплантированных в теплый участок (мг СО2/(м2 ч)); Т – величины среднесуточных температур в слое 0–10 см для соответствующих мезокосмов (°С); a > 0 и b > 0 – параметры.Важно отметить, что экспоненциальная модель не отражает зависимость Q10 от температуры, несмотря на общеизвестный факт об уменьшении значений данного коэффициента с увеличением температуры [15, 18, 24, 28].
Статистическая обработка данных. Для анализа полученных данных использованы описательная статистика, сравнение средних по параметрическому критерию Стьюдента (t-критерий), корреляционный анализ с включением определения рангового коэффициента корреляции Спирмена и регрессионный анализ. Выбранный уровень значимости p = 0.05. Для статистической обработки данных использовали программные пакеты Statistica 10 и Microsoft Excel 2016. Планки погрешностей на столбчатых диаграммах и графиках означают стандартную ошибку.
РЕЗУЛЬТАТЫ
Условия функционирования торфяных почв на контрольных участках. Для исследуемых торфяных почв контрольных участков отмечены значимые различия в значениях температур функционирования (табл. 1).
Так, температура в слое 0–10 см на холодном контрольном участке в среднем в 2.5 раза ниже по сравнению с теплым контрольным участком. Сопоставимая разница отмечена и для глубины протаивания: ее средние значения в пик вегетационного сезона для холодного участка в среднем в 2 раза ниже. Напротив, объемная влажность в слое 0–20 см значимо не различается между участками. Таким образом, изученные торфяные почвы существенно различаются в значениях температур функционирования, что позволяет считать данные участки подходящими объектами для задач данного исследования.
Физические и химические свойства образцов торфяных почв на контрольных участках. Величина ${\text{р}}{{{\text{Н}}}_{{{{{\text{Н}}}_{{\text{2}}}}{\text{О}}}}}{\text{,}}$ как и содержание сырой золы, значимо не различаются между участками. Для содержания Собщ также отмечено отсутствие значимых различий, тогда как содержание Nобщ значимо больше для образцов теплого участка. Наибольшие различия между участками отмечены по показателям степени разложения и массовой влажности. Для образцов холодного контрольного участка значение степени разложения в 2 раза меньше, чем для образцов теплого участка. Напротив, массовая влажность образцов холодного участка в 2.7 раз превышает значение образцов теплого участка. Таким образом, за исключением степени разложения, массовой влажности и содержания Nобщ, образцы торфяных почв контрольных участков характеризуются схожими значениями общих химических свойств.
Параметры функционирования контрольных и трансплантированных образцов почв. Температурный режим. Среднемесячная температура почв на холодном контрольном участке в пик вегетационного сезона варьировала от 2.2 ± 0.1°С до 6.2 ± ± 0.2°С, на теплом контрольном участке от 9.4 ± ± 0.4°С до 12.4 ± 0.3°С, в трансплантированных образцах от 8.9 ± 0.2°С до 13.0 ± 0.2°С (рис. 1).
Рис. 1.
Среднемесячная температура почв в слое 0–10 см на участке “контроль” и в образцах трансплантированных почв (пик вегетационного сезона, август, среднее ± SE, n = 5).
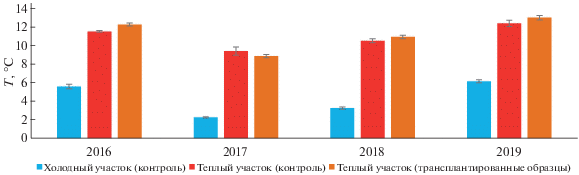
В среднем за 4 года наблюдений в трансплантированных образцах почв установлено трехкратное превышение температуры в слое 0–10 см по сравнению с холодным контрольным участком. Максимальное различие в температурах (в 4 раза) зафиксировано в 2017 г., минимальное (в 2.1 раз) в 2019 г., различия средних статистически значимы (p < 0.05). Отмеченные существенные различия, вероятно, связаны с различной глубиной протаивания на холодном и теплом контрольном участках, в среднем отличающейся в 2 раза. При этом трансплантированные образцы почв и контрольные образцы теплого участка значимо не различались между собой по значению среднемесячной температуры в изученных периодах, что указывает на сходство их термических режимов.
Среднегодовая температура почв на холодном контрольном участке и участке “трансплантация” составила 0.2 и 1.0°С соответственно (рис. 2).
Рис. 2.
Годовая динамика среднемесячных температур почв на глубине 10 см на участке “контроль” и в образцах трансплантированных почв (среднее ± SE, n = 2).
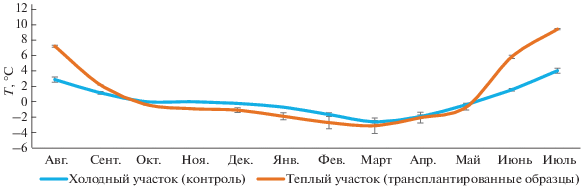
Среднемесячная температура на холодном контрольном участке в течение года изменялась от –2.6 ± 0.2 до 4 ± 0.3°С, в трансплантированных образцах – от –3.1 ± 1.0 до 9.3 ± 0.1°С. Значения среднемесячных температур образцов трансплантированных почв превышали значения холодного контрольного участка только в теплые месяцы (август–сентябрь 2017 г., июнь–июль 2018 г.) – в среднем в 3 раза. В холодный период различия температур статистически значимы только для интервала октябрь 2017 г.–январь 2018 г. (p < 0.05), в остальные месяцы различия не значимы.
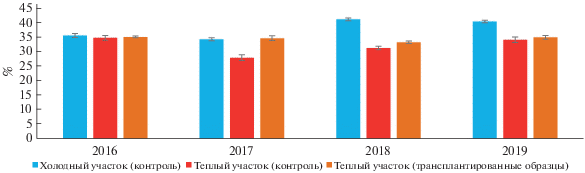
Объемная влажность. Для большей части изученных периодов выявлены статистически значимые различия в объемной влажности почв (рис. 3). Этот показатель на холодном контрольном участке варьировал от 34 ± 1 до 41 ± 0.5%, на теплом контрольном участке от 28 ± 1 до 35 ± 1%, в трансплантированных образцах от 33 ± 0.4 до 35 ± 1%. В августе 2016 г. сравниваемые участки значимо не различались по значениям объемной влажности (p < 0.05). Статистически значимые различия для холодного контрольного участка и трансплантированных образцов почв выявлены в августе 2017–2019 гг. – в этот период условия функционирования почв нельзя считать идентичными по влажности, что может быть обусловлено межгодовой вариабельностью и различной интенсивностью циклов промерзания–оттаивания в сезонно-талом слое многолетнемерзлых пород.
Рис. 3.
Среднемесячная объемная влажность почв в слое 0–20 см на участке “контроль” и в образцах трансплантированных почв (пик вегетационного сезона, август, среднее ± SE, n = 5).
Значения эмиссии СО2 с поверхности почв на холодном контрольном участке в среднем за период наблюдений составили 107 ± 23 мг CO2/(м2 ч), на теплом контрольном участке – 181 ± 24 мг CO2/(м2 ч), с поверхности трансплантированных образцов почв – 217 ± 31 мг CO2/(м2 ч) (рис. 4).
Рис. 4.
Среднемесячные значения эмиссии СО2 с поверхности почв на участке “контроль” и трансплантированных почв (пик вегетационного сезона, август, среднее ± SE, n = 5).
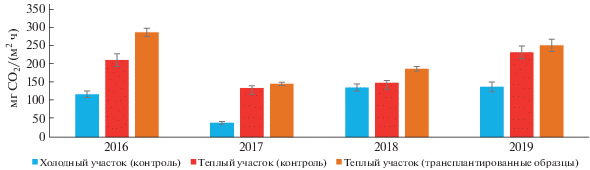
На холодном контрольном участке значения эмиссии СО2 варьировали от 39 ± 3 до 137 ± ± 13 мг CO2/(м2 ч), на теплом контрольном участке от 134 ± 7 до 231 ± 17 мг CO2/(м2 ч), с поверхности трансплантированных образцов почв от 146 ± 4 до 286 ± 11 мг CO2/(м2 ч). В среднем за 4 года наблюдений трансплантация образцов торфяно-криозема обусловила 2.3-кратное увеличение эмиссии СО2 по сравнению со значениями холодного контрольного участка.
Оценка корреляционной связи между значениями эмиссии СО2 и температурой почв. Для оценки корреляции между значениями эмиссии СО2 с поверхности торфяно-криозема и температурой почв в слое 0–10 см для всей совокупности данных рассчитан ранговый коэффициент корреляции Спирмена. Его значения составили в 2016 г. 0.891, в 2017 г. 0.871, в 2018 г. 0.504, в 2019 г 0.686 при значении объединенной выборки за период 2016–2019 гг. 0.752.
Для каждого отдельного периода и в целом за весь период наблюдений (2016–2019 гг.) отмечены статистически значимые корреляции между температурой торфяно-криозема в слое 0–10 см и значениями эмиссии СО2 с поверхности контрольных (холодный участок) и трансплантированных почв. Наибольшие значения коэффициента корреляции отмечены в первые два года наблюдений (2016–2017 гг., r = 0.9), что соотносится с данными температурного режима и эмиссии СО2 с поверхности почв: в этот период различия между контрольными (холодный участок) и трансплантированными почвами по исследуемым показателям максимальны. Дальнейшее уменьшение r до 0.5/0.7 может быть обусловлено совокупным влиянием других факторов среды (в частности, влажности и глубины протаивания), осложняющим проявление температурной зависимости эмиссии СО2 с поверхности почв.
Оценка зависимости эмиссии СО2 с поверхности почв от температуры с использованием экспоненциальной модели. Для количественного описания температурной зависимости используются различные математические функции – среди них наибольшее распространение получила экспоненциальная функция [15, 39], примененная в данном исследовании (рис. 5).
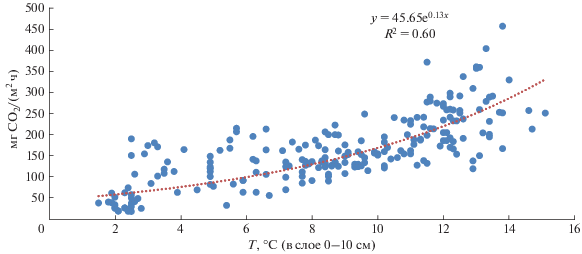
Наилучшим образом экспоненциальная модель описывает результаты наблюдений за период 2016–2017 гг.: значение коэффициента детерминации (R2) составляет 0.8, то есть экспоненциальная модель объясняет до 80% изменчивости значений эмиссии СО2 с поверхности торфяно-криоземов на холодном контрольном участке и участке трансплантация в зависимости от температуры. В целом для объединенного периода наблюдений (2016–2019 гг.) качество экспоненциальной модели ухудшается, значение коэффициента детерминации не превышает 0.6. Тем не менее, достаточный уровень аппроксимации данных позволяет использовать экспоненциальную модель для расчетов коэффициента температурной чувствительности Q10.
Оценка температурной чувствительности эмиссии СО2 с поверхности почв с использованием коэффициента Q10. В целом за период наблюдений (2016–2019 гг.) получены высокие значения коэффициента температурной чувствительности Q10.
Значения коэффициента Q10 в различные годы исследований варьировали от 1.8 ± 0.1 до 6.3 ± 0.3. Максимальное значение Q10 отмечено в 2017 г. на второй год наблюдений (6.3 ± 0.3), что соотносится с максимальной контрастностью термического режима (рис. 1) и значений эмиссии СО2 с поверхности участков (рис. 4), а также с наибольшим значением коэффициента корреляции между эмиссией СО2 и температурой почв в слое 0–10 см. Минимальное значение Q10 зафиксировано в 2018 г. на третий год наблюдений (1.8 ± 0.1).
Для объединенного периода наблюдений (2016–2019 гг.) отмечено высокое значение коэффициента Q10 (3.5/3.7), что свидетельствует о высокой температурной чувствительности эмиссии СО2 с поверхности торфяно-криозема в условиях четырехлетнего полевого трансплантационного эксперимента в рамках наблюдаемого среднего интервала температур (4–11°С) и объемной влажности (30–40%) в пик вегетационного сезона.
ОБСУЖДЕНИЕ
Отклик эмиссии СО2 с поверхности торфяных почв криолитозоны на повышение температуры (температурная зависимость эмиссии СО2 в результате трансплантации). Максимальный отклик значений эмиссии СО2 отмечен в стартовый период наблюдений (2016–2017 гг.), что может объясняться наиболее выраженной в данный период контрастностью термического режима в слое 0–10 см изученных почв (рис. 1), высокой корреляцией эмиссии СО2 с температурой почв в слое 0–10 см, а также с отмеченной в литературе повышенной интенсивностью минерализации легкодоступного органического вещества, наиболее характерной для стартовых периодов трансплантации [34, 38]. В последующий период (2018–2019 гг.) разница в эмиссии СО2 с поверхности трансплантированных образцов почв и образцов холодного контрольного участка характеризовалась меньшей контрастностью, несмотря на значимое различие средних значений – это свидетельствует о постепенной адаптации трансплантированных образцов к изменению гидротермических условий. Некоторому сближению значений эмиссии СО2 с поверхности трансплантированных образцов почв и образцов холодного контрольного участка в период 2018–2019 гг. также может способствовать межгодовая вариабельность значений среднемесячных температур в слое 0–10 см, объемной влажности в слое 0–20 см и глубины сезонного протаивания. Так, в данный период для холодного контрольного участка отмечены двукратный разброс значений среднемесячных температур (3–6°С в слое 0–10 см) и максимальные величины объемной влажности (40–41% в слое 0–20 см), что в сочетании с увеличением глубины сезонного протаивания может обусловливать рост значений эмиссии СО2 с поверхности почв контрольного участка. Важно отметить отсутствие значимой разницы в значениях эмиссии СО2 с поверхности трансплантированных образцов почв и образцов теплого контрольного участка – тенденция характерна для всего изученного периода (2016–2019 гг.), что указывает на возможность быстрой адаптации пересаженных монолитов почв к новым гидротермическим условиям.
Выявленный положительный отклик эмиссии СО2 с поверхности трансплантированных образцов почв соотносится с результатами, полученными в схожих экспериментах. Так, группой шведских авторов по результатам двухлетнего эксперимента показан 42%-ный рост эмиссии СО2 с поверхности органогенных образцов почв тундры, пересаженных в более теплые участки горных лесных экосистем [34]. Тремблэй с соавт. [38] на основании результатов трехлетнего эксперимента установили 27–60%-ное увеличение интенсивности дыхания органогенных горизонтов тундровых почв, перемещенных в более теплый биоклиматический пояс (по сравнению с контрольными участками).
В некоторых публикациях отмечено, что трансплантация образцов почв в более благоприятные гидротермические условия не привела к всплеску значений эмиссии СО2. Например, в работе финских исследователей пересадка органогенных горизонтов лесных почв с северной границы бореальной зоны в южную не привела к увеличению эмиссии СО2 и значений коэффициента Q10 [40]. Близкая закономерность отмечена в работе Уолдропа с соавт., в которой трансплантация образцов луговых почв из холодных участков в теплые не вызвала положительного отклика микробного дыхания [42]. Таким образом, перенос образцов почв из холодных участков в теплые не всегда сопровождается увеличением отклика эмиссии СО2, что в ряде случаев может объясняться большей зависимостью их реакции от типа субстрата и других параметров окружающей среды, чем от температуры.
Эмиссия СО2 активно используется в российских и зарубежных исследованиях как универсальный маркерный показатель отклика дыхательной активности почв на изменение гидротермических условий [5, 41]. Данные по эмиссии СО2 не следует рассматривать и интерпретировать исключительно с биологических позиций, так как данный показатель, помимо биологической составляющей, позволяет выявить особенности физических и физико-химических процессов, протекающих в органогенных и органо-минеральных горизонтах [8, 9].
Температурная чувствительность эмиссии СО2 с поверхности торфяно-криозема в результате трансплантации. В большинстве периодов наблюдений значения коэффициента Q10 находились в диапазоне 3–6, что свидетельствует о высоком отклике пересаженных образцов. Полученный диапазон значений Q10 подтверждается литературными данными, показывающими его возможную вариабельность для почв торфяников и тундровых областей [14, 18]. Исключением являются данные за 2018 г. – в этот период температурная чувствительность принимала наиболее низкие значения за все 4 года наблюдений (Q10 = 1.6/1.8). Это может быть обусловлено повышенными величинами эмиссии СО2 с поверхности контрольных образцов холодного участка, а также связано со значимой корреляцией между эмиссией СО2 и прочими гидротермическими параметрами (в частности с объемной влажностью в слое 0–20 см, r = 0.3, р < < 0.05) на этом же участке. В остальные периоды наблюдений (2016–2017 и 2019 гг.) для трансплантированных образцов отмечена высокая температурная чувствительность эмиссии СО2, что позволяет говорить об ускорении минерализации органического вещества в торфяных почвах криолитозоны в случае значимого увеличения температуры (в 2–4 раза). Несмотря на уменьшение корреляционной связи между эмиссией СО2 и температурой с увеличением длительности эксперимента, температурная чувствительность характеризовалась иной закономерностью и, за исключением 2018 г., оставалась стабильно высокой (Q10 = 3–6). Это позволяет говорить о нелинейных механизмах отклика торфяных почв криолитозоны и о необходимости его дальнейшего детального изучения в полевых и лабораторных условиях.
Напротив, в ряде схожих исследований температурная чувствительность эмиссии СО2 с поверхности трансплантированных образцов почв значимо не превышала значения контрольных образцов. В работах скандинавских авторов трансплантация органогенных образцов почв в более теплые условия не привела к достоверным отличиям в значениях коэффициента Q10 [34, 40]. В исследовании Тремблэй с соавт. изменение значений Q10 в результате пересадки образцов почв зависело не от температуры, а от особенностей субстрата и типа фитоценоза на участках отбора образцов [38]. Приоритетное влияние субстрата на микробиологическую активность пересаженных образцов почв также отмечено в ряде других публикаций [13, 19]. В некоторых работах по трансплантации показано значимое влияние прочих сезонных факторов окружающей среды (влажности) на биологическую активность почв – отмечается, что эффект от воздействия сезонных факторов может превышать эффект от непосредственного переноса [26]. Важным аспектом также является возможный запуск микробных сукцессий, вызванных трансплантацией, и их влияние на биологическую активность пересаженных образцов почв [35]. Пролонгированый эффект от воздействия трансплантации (наблюдения проводились в течение трех и более лет) при переносе образцов почв в более теплые климатические условия в ряде случаев может приводить к уменьшению содержания общей микробной биомассы [19, 38, 40], а также лабильных и стабильных фракций органического вещества в пересаженных образцах [27, 30].
В современных манипуляционных исследованиях по моделированию отклика почв на климатические изменения методу трансплантации уделяется все большее внимание [45]. Его основными плюсами, помимо простой установки, являются прогревание почв в нативных условиях и возможность прямых измерений эмиссии СО2 в пересаженных образцах. Также отмечается, что метод в большей степени применим для оценки первичного отклика образцов почв на резкие изменения гидротермического режима – с увеличением продолжительности эксперимента эффект от переноса может быть замещен влиянием сезонных и иных факторов окружающей среды [13, 19, 26, 35]. В нашем исследовании с использованием метода трансплантации получен большой объем данных, иллюстрирующих значимые различия между параметрами функционирования почв изученных участков. Следовательно, данный метод может быть использован для количественной оценки температурной чувствительности эмиссии СО2 с поверхности торфяных почв криолитозоны в исследуемом регионе.
ЗАКЛЮЧЕНИЕ
Температурная чувствительность эмиссии СО2 с поверхности торфяно-криоземов севера Западной Сибири, оцененная по результатам четырехлетнего полевого эксперимента методом трансплантации, характеризовалась высокими значениями коэффициента температурной чувствительности Q10, который варьировал от 2 до 6 и в среднем за 4 года наблюдений составил 3.5 ± 0.2. Полученные значения коэффициента Q10 свидетельствуют о повышенной температурной чувствительности эмиссии СО2 с поверхности трансплантированных образцов почв в условиях существенно более высоких температур функционирования – подобный отклик торфяно-криозема, с учетом его сохранения в течение продолжительного периода, необходимо учитывать при моделировании углеродного баланса исследуемой территории. Таким образом, метод трансплантации может быть использован для количественной оценки температурной чувствительности эмиссии СО2 с поверхности торфяных почв криолитозоны в исследуемом регионе. Для более детального прогнозирования отклика торфяных почв криолитозоны на климатические изменения необходимы дальнейшие исследования взаимосвязи гидротермических параметров и температурной чувствительности эмиссии СО2 с поверхности почв, а также изучение состава органического вещества почв и его изменчивости на предмет определения потенциальных климатических маркеров.
Список литературы
Бобрик А.А., Гончарова О.Ю., Матышак Г.В., Рыжова И.М., Макаров М.И. Влияние геокриологических условий и свойств почв на пространственное варьирование эмиссии СО2 почвами плоскобугристых болот островной криолитозоны Западной Сибири // Почвоведение. 2016. № 12. С. 1445–1456. https://doi.org/10.7868/S0032180X1610004X
Васильевская В.Д., Иванов В.В., Богатырев Л.Г. Почвы севера Западной Сибири. М.: Изд-во Моск. ун-та, 1986. 227 с.
Гончарова О.Ю., Матышак Г.В., Бобрик А.А., Москаленко Н.Г. Продуцирование диоксида углерода почвами северной тайги Западной Сибири (Надымский стационар) // Криосфера Земли. 2014. Т. 18(2). С. 66–71.
Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
Курганова И.Н., Лопес де Гереню В.О., Галлардо Ланчо Х.Ф., Ем К.Т. Оценка скорости минерализации органического вещества почв в лесных экосистемах внутриконтинентального умеренного, средиземноморского и тропического муссонного климата // Почвоведение. 2012. № 1. С. 82–94.
Ландшафты криолитозоны Западно-Сибирской нефтегазоносной провинции / Под ред. Е.С. Мельникова. Новосибирск: Наука, 1983. 165 с.
Матышак Г.В., Богатырев Л.Г., Гончарова О.Ю., Бобрик А.А. Особенности развития почв гидроморфных экосистем северной тайги Западной Сибири в условиях криогенеза // Почвоведение. 2017. № 10. С. 1155–1164.
Наумов А.В. Дыхание почвы: составляющие, экологические функции, географические закономерности. Новосибирск: Изд-во СО РАН, 2009. 208 с.
Смагин А.В. Газовая фаза почв. М.: Изд-во Моск. ун-та, 2005. 301 с.
Тархов М.О., Матышак Г.В., Рыжова И.М., Гончарова О.Ю., Бобрик А.А., Петров Д.Г., Петржик Н.М. Температурная чувствительность дыхания почв бугристых торфяников севера Западной Сибири // Почвоведение. 2019. № 8. С. 946–955.
Теория и практика химического анализа почв / Под ред. Л.А. Воробьевой. М.: Геос, 2006. 400 с.
Aronson E.L., McNulty S.G. Appropriate experimental ecosystem warming methods by ecosystem, objective, and practicality // Agricultural and Forest Meteorology. 2009. V. 149. P. 1791–1799.
Bani A., Borruso L., Fornasier F., Pioli S., Wellstein C., Brusetti L. Microbial Decomposer Dynamics: Diversity and Functionality Investigated through a Transplantation Experiment in Boreal Forests // Microbial Ecology. 2018. V. 76(4). P. 1030–1040. https://doi.org/10.1007/s00248-018-1181-5
Chen H., Tian H.-Q. Does a General Temperature-Dependent Q10 Model of Soil Respiration Exist at Biome and Global Scale? // J. Integrative Plant Biology. 2005. V. 47. P. 1288–1302.
Davidson E.A., Janssens I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change // Nature. 2006. V. 440. P. 165–173.
Euskirchen E.S., Edgar C.W., Turetsky M.R., Waldrop M.P., Harden J.W. Differential response of carbon fluxes to climate in three peatland ecosystems that vary in the presence and stability of permafrost // JGR Biogeosciences. 2014. V. 119. P. 1576–1595.
FAO 2017. Soil organic carbon – the hidden potential. Food and Agriculture Organization of the United Nations Rome, Italy.
Hamdi S., Moyano F., Sall S., Bernoux M., Chevallier T. Synthesis analysis of the temperature sensitivity of soil respiration from laboratory studies in relation to incubation methods and soil conditions // Soil Biology & Biochemistry. 2013. V. 58. P. 115–126.
Hedenec P., Jilkova V., Lin Q. et al. Microbial communities in local and transplanted soils along a latitudinal gradient // Catena. 2019. V. 173. P. 456–464.
Hilasvuori E., Akujarvi A., Fritze H. et al. Temperature sensitivity of decomposition in a peat profile // Soil Biol. Biochem. 2013. V. 67. P. 47–54.
Hobbie S.E., Stuart Chapin F. The Response of Tundra Plant Biomass, Aboveground Production, Nitrogen, and CO2 Flux to Experimental Warming // Ecology. 1998. V. 79. P. 1526–1544.
Hugelius G., Strauss J., Zubrzycki S. et al. Estimated stocks of circumpolar permafrost carbon with quantified uncertainty ranges and identified data gaps // Biogeosciences. 2014. V. 11. P. 6573–6593.
Kirschbaum M.U.F. The temperature dependence of organic matter decomposition – still a topic of debate // Soil Biol. Biochem. 2006. V. 38. P. 2510–2518.
Kirschbaum M.U.F. The temperature dependence of soil organic matter decomposition and the effect of global warming on soil organic C storage // Soil Biol. Biochem. 1995. V. 27. P. 753–760.
Landva A.O., Pheeney P.E. Peat fabric and structure // Can. Geotech J. 1980. V. 17. P. 416–435.
Lazzaro A., Gauer A., Zeyer J. Field-Scale Transplantation Experiment To Investigate Structures of Soil Bacterial Communities at Pioneering Sites // Applied and Environmental Microbiology. 2011. V. 77. P. 8241–8248.
Link S.O., Smith J.L., Halvorson J.J., Bolton H. A reciprocal transplant experiment within a climatic gradient in a semiarid shrub-steppe ecosystem: effects on bunchgrass growth and reproduction, soil carbon and soil nitrogen // Global Change Biology. 2003. V. 9. P. 1097–1105.
Lloyd J., Taylor J.A. On the temperature dependence of soil respiration // Functional Ecology. 1994. V. 8. P. 315–323.
Natali S.M., Schuur E.A.G., Webb E.E., Hick Pries C.E., Crummer K.G. Permafrost degradation stimulates carbon loss from experimentally warmed tundra // Ecology. 2014. V. 95. P. 602–608.
Puissant J., Mills R.T.E. Robroek B.J.M. et al. Climate change effects on the stability and chemistry of soil organic carbon pools in a subalpine grassland // Biogeochemistry. 2017. V. 132. P. 123–139.
Ravn N.R., Elberling B., Michelsen A. Arctic soil carbon turnover controlled by experimental snow addition, summer warming and shrub removal // Soil Biol. Biochem. 2020. V. 142. https://doi.org/10.1016/j.soilbio.2019.107698
Semenchuk P.R., Krab E.J., Hedenstrom M., Phillips C.A., Ancin-Murguzur F.J., Cooper E.J. Soil organic carbon depletion and degradation in surface soil after long-term non-growing season warming in High Arctic Svalbard // Science of the Total Environment. 2019. P. 158–167.
Sierra C.A. Temperature sensitivity of organic matter decomposition in the Arrhenius equation: Some theoretical considerations // Biogeochemistry. 2012. V. 108. P. 1–15.
Sjogersten S., Wookey P.A. Climatic and resource quality controls on soil respiration across a forest–tundra ecotone in Swedish Lapland // Soil Biol. Biochem. 2002. V. 34. P. 1633–1646.
Sun B., Wang F., Jiang Y., Li Y., Dong Z., Li Z., Zhang X. A long-term field experiment of soil transplantation demonstrating the role of contemporary geographic separation in shaping soil microbial community structure // Ecology and Evolution. 2014. V. 4(7). P. 1073–1087.
Tarnocai C., Canadell J.G., Schuur E.A.G., Kuhry P., Mazhitova G., Zimov S. Soil organic carbon pools in the northern circumpolar permafrost region // Global Biogeochemical Cycles. 2009. V. 23. № 2. GB2023.https://doi.org/10.1029/2008GB003327
Thiessen S., Gleixner G., Wutzler T., Reichstein M. Both priming and temperature sensitivity of soil organic matter decomposition depend on microbial biomass – An incubation study // Soil Biol. Biochem. 2013. V. 57. P. 739–748.
Tremblay S.L., D’Orangeville L., Lambert M.-C., Houle D. Transplanting boreal soils to a warmer region increases soil heterotrophic respiration as well as its temperature sensitivity // Soil Biology & Biochemistry. 2018. V. 116. P. 203–212.
Tuomi M., Vanhala P., Karhu K., Fritze H., Liski J. Heterotrophic soil respiration – comparison of different models describing its temperature dependence // Ecological Modelling. 2008. V. 211. P. 182–190.
Vanhala P., Karhu K., Tuomi M., Bjorklof K., Fritze H., Hyvarinen H., Liski J. Transplantation of organic surface horizons of boreal soils into warmer regions alters microbiology but not the temperature sensitivity of decomposition // Global Change Biology. 2011. V. 17. P. 538–550.
Voigt C., Treat C.C. Ecosystem carbon response of an Arctic peatland to simulated permafrost thaw // Global Change Biology. 2019. V. 25. P. 1746–1764.
Waldrop M.P., Firestone M.K. Response of Microbial Community Composition and Function to Soil Climate Change // Microbial Ecology. 2006. V. 52. P. 716–724.
Xue K. et al. Tundra soil carbon is vulnerable to rapid microbial decomposition under climate warming // Nature Climate Change. 2016. V. 6. P. 595–600.
Yu Z.C. Northern peatland carbon stocks and dynamics: a review // Biogeosciences. 2012. V. 9. P. 4071–4085.
Zhao M., Sun B., Wu L. et al. Dissimilar responses of fungal and bacterial communities to soil transplantation simulating abrupt climate changes // Molecular Ecology. 2019. V. 28. P. 1842–1856.
Дополнительные материалы отсутствуют.