

Почвоведение, 2022, № 6, стр. 673-686
Подстилки городских насаждений как индикатор интенсивности биологического круговорота в условиях мегаполиса
О. В. Семенюк a, *, В. М. Телеснина a, Л. Г. Богатырев a, Ф. И. Земсков a
a МГУ им. М.В. Ломоносова
119991 Москва, Ленинские горы, 1, Россия
* E-mail: olgatour@rambler.ru
Поступила в редакцию 13.10.2021
После доработки 06.01.2022
Принята к публикации 10.01.2022
- EDN: DNGIYM
- DOI: 10.31857/S0032180X22060119
Аннотация
Оценена интенсивность биологического круговорота городских парковых насаждений на основе оценки следующих показателей структуры и функционирования подстилок: общие запасы подстилки, запасы и доля детрита в горизонте L, запасы и доля ЛРК в горизонте L, отношение запасов горизонта L к сумме запасов нижележащих горизонтов, зольность горизонта L, запасы золы ЛРК и доля золы этих компонентов в общем запасе золы горизонта L. Исследования проводили на территории Битцевского парка под насаждениями ели, березы и липы. Для городских насаждений выявлены существенные отличия от естественных в составе живого напочвенного покрова, в котором появляются нетипичные для лесов луговые и сорно-рудеральные виды, указывающие на антропогенное воздействие. Городские парковые насаждения отличаются от природных экосистем повышенной интенсивностью биологического круговорота, на что указывают общие запасы и доля детрита, легкоразлагаемых компонентов, запасы и доля золы легкоразлагаемых компонентов в горизонте L. В городских условиях подстилки хвойных насаждений характеризуются упрощенным строением, малой мощностью (не более 4–5 см) и повышенной зольностью отдельных компонентов (более 10%), которая отражает значительное участие механической миграции твердых частиц различного происхождения. По большинству характеристик подстилки лиственных городских насаждений не так существенно отличаются от аналогичных естественных, как подстилки хвойных насаждений, поэтому именно подстилки ельников могут служить индикатором антропогенного воздействия на городские экосистемы. В целом показатели биологического круговорота, связанные с особенностями лесной подстилки, указывают на повышенную по сравнению с аналогичными естественными фитоценозами интенсивность биогенной миграции. Это необходимо учитывать при разработке прогноза развития озелененных территорий городов.
ВВЕДЕНИЕ
В системе познания сущности почвообразования биологическому круговороту принадлежит ведущая роль, что отмечено в классических работах В.Р. Вильямса, общепризнанной концепции и методологии, сформулированных в работах Н.И. Базилевич с соавт. [21] и современных работах А.А. Титляновой [1, 33]. Согласно В.И. Вернадскому, биологический круговорот является одним из важнейших показателей устойчивости биосферы и на современном уровне рассматривается в качестве важнейшего критерия устойчивого развития биосферы [24]. Эти положения касаются не только естественных фитоценозов, но и рукотворных, чья роль в обеспечении оптимальных условий жизнедеятельности в условиях современных мегаполисов приобретает все большее значение. Процессы, происходящие в лесных насаждениях в пределах городов, требуют особого внимания, в том числе необходим поиск индикаторов, отражающих закономерности их функционирования.
В городах древесные насаждения имеют огромное экологическое значение [50], выполняя широкий набор экологических функций, в том числе формирование геохимического барьера в виде подстилки, которая задерживает поллютанты [35]. Рекреационная функция насаждений в условиях современных мегаполисов едва ли может быть переоценена. Следует отметить, что функционирование фитоценозов в мегаполисах отличается от естественных условий по многим факторам, что отражено в многочисленных работах, посвященных городским фитоценозам и почвам. Повышенное содержание тяжелых металлов в городских почвах [38, 43, 55, 57] требует оценки рисков для здоровья населения [51]. Исследованиями последних лет установлены особенности микробного пула [39] в почвах под лесными насаждениями в городах, специфике преобразования органического вещества и особенностей его миграции [54]. Подчеркивается необходимость учета содержания органического вещества под непроницаемыми поверхностями [37], поступления аэрозолей и пыли [41]. Обращается внимание на засорение городских биогеоценозов антропогенным мусором [52]. Подчеркивается роль мезофауны в разложении подстилки, особенно при поступлении в урбоэкосистемы азота техногенной и/или антропогенной природы [48], отмечено активное участие макрофауны и значимость видового разнообразия растений в функционировании лесных экосистем в условиях города [47]. Интересны работы по установлению роли мелких корневых систем в повышении интенсивности разложения подстилки [56]. В рамках стехиометрии показана динамика изменения соотношения С : N : P в листьях и корнях растений, подстилке и почве [42]. На примере исследования 27 парков Финляндии установлено, что под вечнозелеными растениями интенсивность накопления углерода может быть больше по сравнению с лиственными древесными породами [49], но эта способность меньше, чем в аналогичных естественных лесных сообществах. На основании близких величин содержания углерода в парковых почвах и естественных лесах выдвинуто предположении, что уборка листвы не оказывает серьезного влияния на накопление углерода в почве.
Представленный материал убедительно показывает возрастающий интерес к изучению компонентов городских экосистем и их функционированию, исследованию особенностей биологического круговорота на урбанизированных территориях. В отечественной научной литературе применяют показатели, характеризующие состояние подстилки. Фундаментальным показателем является соотношение запасов подстилки и опада, характеризующее интенсивность круговорота [21]. Состояние подстилки используют при оценке и группировке лесных земель, а также в целях бонитировки лесных почв [34]. Морфогенез подстилок лежит в основе их классификации [3, 26], а типология подстилок хорошо коррелирует с критериями, отражающими скорость круговорота веществ в биогеоценозах [9], что убедительно показано для насаждений Ботанического сада МГУ им. М.В. Ломоносова. Расчет соотношений запасов подстилки, определенных в летний и осенний периоды [29], позволил оценить величину реализации органического вещества, сосредоточенного в подстилках, в годичном цикле. В теоретическом отношении методическая основа исследования подстилок естественных и городских насаждений однотипна – от типологической принадлежности к структурной организации. При близком характере функционирования городских и естественных фитоценозов, который определяется в первую очередь видовой принадлежностью насаждений, поиск специфики преобразования наземного детрита является актуальным в научном и практическом отношении. Следует признать, что до сих пор нет точных индикаторов и показателей, позволяющих дифференцировать естественные и городские фитоценозы в рамках биологического круговорота. Это обусловлено относительно слабой изученностью фитоценозов в пределах городских ландшафтов и позволяет заключить, что изучение биологического круговорота в условиях мегаполиса актуально.
Цель работы – выявление интенсивности биологического круговорота урбанизированных территорий на основе оценки параметров структурной организации подстилок городских парковых насаждений.
ОБЪЕКТЫ И МЕТОДЫ
Природно-исторический парк “Битцевский лес” – особо охраняемая природная территория площадью 2208 га, образованная в 1994 г. в пределах г. Москвы. Климат на территории парка влажный, умеренно континентальный, имеет четко выраженную сезонность. Средняя многолетняя температура составляет +5.4°С, среднегодовое количество осадков – 650 мм, а годовая испаряемость – 434 мм. Территория парка находится на юго-восточной, приподнятой части Теплостанской возвышенности, которая представляет собой отдельный природный район, сформированный на доледниковом останце, являющемся частью Москворецко-Окской равнины [8]. Характер рельефа холмистый, с густой овражно-балочной сетью, которая представлена глубокими эрозионными долинами, балками и оврагами [25]. Доминирующей почвообразующей породой водораздельных пространств являются некарбонатные средние и тяжелые покровные суглинки, московская и днепровская морены и флювиогляциальные отложения на них. Территория природно-исторического парка “Битцевский лес” принадлежит к подзоне хвойно-широколиственных лесов. Лесные экосистемы (еловые, дубовые, липовые и березовые насаждения) занимают до 63% площади парка [36]. Средний возраст лесных пород на территории составляет 84 г. Почвенный покров неоднородный, с преобладанием дерново-подзолистых преимущественно среднесуглинистых почв разной степени оподзоленности [14].
Объекты исследования представлены тремя типичными насаждениями природно-исторического парка “Битцевский лес”, каждый из которых занимает около 400 м2. Хвойное и липовое насаждения находятся в 70 м от ближайшей проезжей части (ул. Голубинская) и в 15 м друг от друга. Такое пространственное расположение не исключает возможность обмена опадом. Березняк отдален от двух других объектов на расстояние 3.5 км, от автомобильной дороги – на 315 м (ул. Академика Капицы). Насаждения расположены на хорошо дренированных участках пологих склонов водораздельного пространства.
Березняк марьянниково-злаковый – 70 лет. Древостой сформирован березой повислой (Betula pendula). В обильном травостое (проективное покрытие до 90%) доминируют ежа сборная (Dactylis glomerata), марьянник дубравный (Melampyrum nemorosum) и полевица гигантская (Agrostis gigantea). Почва – дерново-подзолистая [13] (Albic Retisol).
Липняк лютиково-живучковый – 70 лет. Древесный ярус представлен липой сердцелистной (Tilia cordata). Травяной ярус преимущественно состоит из живучки ползучей (Ajuga reptans) и лютика кашубского (Ranunculus cassubicus). Почва – дерново-подзолистая (Albic Retisol).
Ельник крапивно-живучковый – 100 лет. В древостое главенствующую роль играет ель обыкновенная (Picea abies) с участием липы сердцелистной (Tilia cordata) – 7Е3Л. В составе травянистого яруса, проективное покрытие которого составляет менее 30%, преобладает живучка ползучая (Ajuga reptans) и крапива двудомная (Urtica dioica). Почва – дерново-подзолистая (Albic Retisol).
Во всех насаждениях практически нет подлеска (кустарников) и очень мало подроста, то есть ярусная структура упрощенная, что типично для городских насаждений.
Все полевые работы проводили в выбранных для изучения фитоценозах на пробных площадках 10 × 10 м. Для изучения растительности выполняли геоботанические описания. Виды травяного яруса, которому уделяли особое внимание как наиболее чувствительному к локальным экологическим условиям, относили к эколого-ценотическим свитам или группам свит по А.А. Ниценко [19]. Отбор подстилок проводили на площади 50 × 50 см в 9-кратной повторности в лиственных насаждениях. В еловых насаждениях образцы органогенного горизонта отбирали в 5-кратной повторности с учетом положения в тессере: ствол–крона–окно. Для определения классификационной принадлежности подстилок использовали классификацию Л.Г. Богатырева [3].
Подстилки отбирали по горизонтам. Верхний горизонт L разбирали на фракции: листья, хвою, ветки, кору, шишки, ветошь (опад трав). Мелкие компоненты горизонта L, происхождение которых установить невозможно, обозначали как детрит. Совокупность фракций ветоши и листьев называли легкоразлагаемыми компонентами (ЛРК), содержание и запасы которых очень важный показатель, так как именно эти компоненты в первую очередь разлагаются микроорганизмами и активно трансформируются в результате деятельности почвенных беспозвоночных животных [40]. Запасы всех фракций и подстилки в целом рассчитывали на абсолютно сухую массу. Зольность компонентов подстилки определяли при 450°С. На основе полученных результатов рассчитывали показатели функционирования подстилок: 1) общие запасы подстилки; 2) тип строения (деструктивная, ферментативная или гумифицированная); 3) отношение запасов горизонта L к сумме запасов остальных горизонтов (при наличии); 4) запасы и доля ЛРК в горизонте L; 5) запасы и доля детрита в горизонте L; 6) средневзвешенная зольность горизонта L; 7) запасы золы ЛРК и ее доля в общем запасе золы горизонта L.
Для выявления особенностей биологического круговорота в городских условиях на основе опубликованных ранее данных [32, 53] рассчитывали показатели круговорота для аналогичных естественных насаждений ели, липы и березы, расположенных в Подмосковье на территории УОПЭЦ “Чашниково” на дерново-подзолистых почвах.
Для обработки полученных данных использовали программы Excel и Statistica.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Особенности живого напочвенного покрова. Городской лес, каким является изучаемый фрагмент Битцевского парка, – это образование, промежуточное между лесом и парком [18], что отражается на живом напочвенном покрове. Изученные фитоценозы соответствуют третьей (березняк) и третьей–четвертой (липняк и ельник) стадиям дигрессии. Об этом свидетельствуют наличие в живом напочвенном покрове луговых и сорно-рудеральных видов и их соотношение с терминальными видами [23]. При этом фитоценозы существенно различаются по составу живого напочвенного покрова (рис. 1).
Рис. 1.
Соотношение видов разных эколого-ценотических групп в травяном ярусе. 1 – виды сорно-рудеральных свит, 2 – виды луговой нитрофильной свиты, 3 – виды прочих луговых свит, 4 – виды мелколиственной опушечно-полянной свиты, 5 – виды осиновой свиты, 6 – виды неморальных свит, 7 – виды еловых свит, 8 – эвритопные виды.
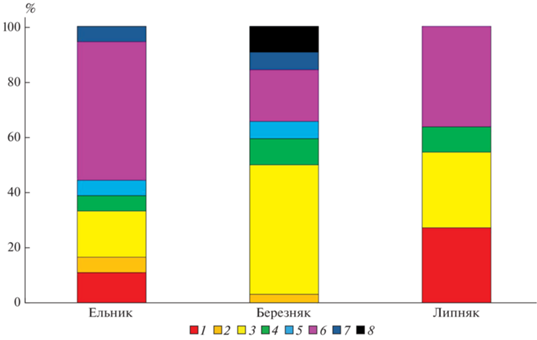
Максимальное число видов выявлено в березовом насаждении (23), минимальное – в липовом (11). Помимо возможной рекреационной нагрузки, разнообразие видов живого напочвенного покрова в ельнике и липняке детерминировано степенью освещенности, которая в этих насаждениях меньше, чем в подобных естественных насаждениях, так как световой режим начинает существенно изменяться с третьей стадии дигрессии [6].
Во всех насаждениях выявлено много неморальных видов – спутников широколиственных пород, что типично для парков Москвы [11, 20]. В травяном ярусе березовых насаждений высока доля видов луговых – 47% (полевица гигантская (Agrostis gigantea)) и неморальных – 19% (марьянник дубравный (Melampyrum nemorosum)) видов. Также значительно участие опушечных (зверобой пятнистый (Hypericum maculatum)) и эвритопных видов (лютик едкий (Ranunculus acris)). Подобное сочетание свит характерно для мелколиственных насаждений, расположенных в подзоне хвойно-широколиственных лесов. Сорные виды не обнаружены, что, вероятно, обусловлено удаленностью участка от пешеходных маршрутов и автомобильных дорог, а также конкуренцией со стороны луговых видов, особенно злаков.
Липовое насаждение характеризуется низким разнообразием видов живого напочвенного покрова, а также минимальным числом эколого-ценотических свит. Виды живого напочвенного покрова относятся к неморальной группе – 36% (лютик кашубский (Ranunculus cassubicus) и живучка ползучая (Ajuga reptans)). Присутствует луговая (вербейник монетчатый (Lysimachia nummularia)) и сорно-рудеральная группы (крапива двудомная и недотрога мелкоцветковая (Impatiens parviflora)). Следует отметить, что вербейник монетчатый, доля покрытия которого особенно велика в липняке, хорошо переносит вытаптывание [5]. Большая доля синантропных растений может быть объяснена близостью расположения к крупным парковым пешеходным тропам и проезжей части (70 м).
В ельнике доля неморальных видов (живучка ползучая и гравилат городской (Geum urbanum)) максимальна – 50%, отмечено преобладание луговой растительности (вербейник монетчатый, лютик ползучий (Ranunculus repens), щавель конский (Rumex confertus)) – 17%. Наличие светолюбивых видов может быть связано с отсутствием подлеска и малым количеством подроста, которые не дают сильного затенения, свойственного естественным хвойным насаждениям. В небольшом количестве встречаются растения еловой свиты – золотарник обыкновенный (Solidago virgaurea).
Повышенная доля неморальных видов в живом напочвенном покрове может быть обусловлена особенностями городского микроклимата, который отличается от микроклимата естественных фитоценозов. Сорно-рудеральные виды могут быть результатом как непосредственно рекреации, так и переноса семян из городских ландшафтов селитебной зоны. Упрощенная вертикальная структура парковых фитоценозов также создает дополнительное открытое пространство [28], что способствует распространению не свойственных естественным лесам видов, характеризующихся повышенной зольностью. Последнее характеризует особенности биологического круговорота.
За счет упрощения вертикальной структуры фитоценоза, биомасса подроста и подлеска или снижена, или отсутствует, что уменьшает запас органического вещества и зольных элементов в экосистеме, то есть емкость биологического круговорота.
Свойства лесных подстилок. Хвойные насаждения. Изучение подстилок на территории Битцевского парка показало, что встречаются два их типа: деструктивные и ферментативные (табл. 1). При этом в приствольных пространствах преобладают ферментативные, в окнах – деструктивные, а под кронами – деструктивные и ферментативные примерно в равном соотношении. Ферментативные горизонты имеют промежуточный характер между L и F в насаждениях, подверженных рекреации, где гумифицированные и ферментативные горизонты подстилок маломощны. Последнее может быть связано как с высокой скоростью разложения органического вещества, так и механическим разрушением подстилок при их вытаптывании [2, 7, 15].
Таблица 1.
Распределение подстилок различных типов в пределах тессер еловых насаждений, количество
| Тип подстилки | Тессеры | ||
|---|---|---|---|
| стволы | кроны | окна | |
| Деструктивная | 1 | 3 | 4 |
| Ферментативная | 4 | 2 | 1 |
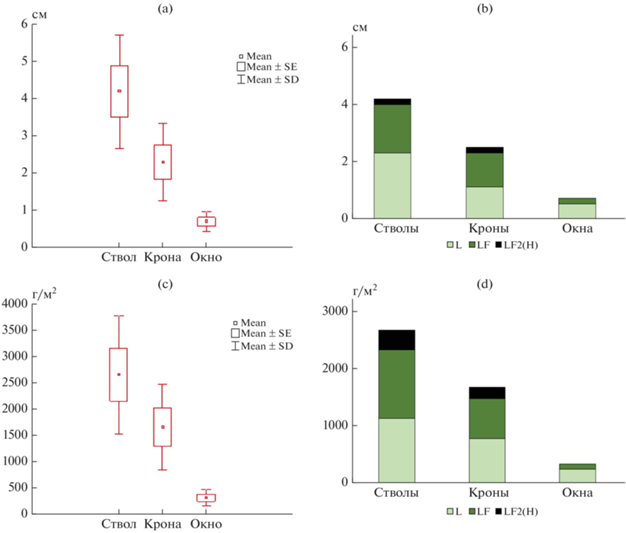
Данные морфологические особенности не типичны для естественных еловых насаждений – в последних чаще всего встречаются не только ферментативные, но и гумифицированные подстилки, особенно в приствольных пространствах [53]. Подстилки характеризуются низкой мощностью, которая в пределах тессеры уменьшается от ствола к межкроновому пространству, составляя в последнем менее 1 см (рис. 2), что не типично для естественных еловых фитоценозов. Значимо отличается от других компонентов тессеры только мощность лесных подстилок в окнах. Горизонты LF2, переходные к гумифицированным, очень малы (менее 1 см) и развиты только под кронами и в приствольных пространствах. Сходным образом меняются запасы подстилок – в приствольных пространствах этот показатель превышает соответствующее значение в окнах почти в 10 раз. В естественных ельниках такие различия в пределах тессеры составляют 15–20 раз [4, 53]. Можно считать, что условия города несколько нивелируют варьирование мощности и запасов подстилок в пределах фитоценоза. Уменьшение запасов и мощности подстилок по сравнению с естественными лесами связано в том числе с рекреационным воздействием [45].
Рис. 2.
Мощность (a, b) и запасы (c, d) подстилок хвойных насаждений: a, c – общие; b, d – по горизонтам.
В составе горизонта L всех подстилок ельника преобладают ветки и хвоя (рис. 3). Повышенный опад веток может говорить о неблагоприятных условиях роста деревьев из-за повышенного антропогенного воздействия [45] – хвойные насаждения менее устойчивы к атмосферному загрязнению, нежели лиственные [12]. Сравнительно небольшую долю по сравнению с подстилками естественных ельников [31] занимает детрит – не более 5–6%. Возможно, это связано с лучшими условиями разложения органического вещества по сравнению с естественными фитоценозами, или измельчением детрита при рекреационной нагрузке, что ускоряет его минерализацию. В горизонте L лесных подстилок, приуроченных к окнам в пределах тессеры, отмечено минимальное относительное содержание детрита. Практически отсутствует фракция ветоши (<2 г/м2), что может быть обусловлено небольшим проективным покрытием травяного яруса. Запасы и доля листьев в горизонте L увеличиваются от стволов к окнам. Запасы легкоразлагаемых фракций подстилки практически не меняются от стволов к окнам, а их доля увеличивается в этом направлении в 4–5 раз. Аналогичная картина выявлена при изучении естественных ельников тайги и подзоны хвойно-широколиственных лесов [31, 53]. Видимо, это связано с изменением состава опада и условий его разложения в направлении от приствольных элементов тессеры к окнам.
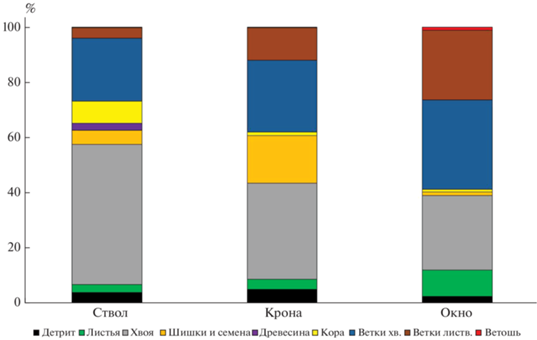
Средневзвешенная зольность горизонтов L подстилок составляет 8–12%, при этом наиболее высокозольные компоненты: ветошь (26–27%) и детрит (16–22%), что согласуется с полученными ранее данными по зольности подстилок в городских ландшафтах [27]. По-видимому, имеет место существенное пылевое загрязнение растений и поступление частиц пыли вместе с опадом в подстилку [10]. В ходе рекреации появляется возможность дополнительного поступления минеральных частиц. Возможно, именно это является причиной относительно высоких запасов постилки при низкой мощности. Зольность нижних горизонтов подстилки, сильно перемешанных с минеральными горизонтами, достигает 30% (табл. 2). Общие запасы зольных элементов, накапливаемые в подстилках и их отдельных горизонтах, уменьшаются в 4 раза от приствольных пространств к окнам, что детерминировано подобной закономерностью пространственного распределения запасов подстилок. При этом доля зольных элементов в горизонте L по отношению к общей сумме золы в подстилке составляет 20–30% в пристволье и под кроной, тогда как в окне – более 50%. В горизонтах L наибольшая доля зольных элементов аккумулируется за счет хвои и ветвей.
Таблица 2.
Свойства компонентов подстилок хвойных насаждений, среднее ± стандартная ошибка
| Горизонт или его компонент | Стволы | Кроны | Окно |
|---|---|---|---|
| Зольность, % | |||
| Листья | 15.7 ± 1.4 | 13.6 ± 1.5 | 16.1 ± 0.6 |
| Ветошь | 26.4 ± 0.7 | – | 27.0 ± 3.6 |
| Хвоя | 11.96 ± 0.1 | 12.9 ± 1.3 | 17.58 ± 1.3 |
| Детрит | 17.5 ± 2.8 | 16.0 ± 1.8 | 22.0 ± 4.0 |
| Ветки хвойных | 6.13 ± 0.6 | 5.81 ± 0.5 | 7.5 ± 0.1 |
| Ветки лиственных | 5.5 ± 0.5 | 5.3 ± 0.5 | 8.7 ± 0.6 |
| Кора | 7.7 ± 1.1 | 7.9 ± 0.4 | 7.0 ± 1.7 |
| Древесина | 5.5 ± 1.4 | – | – |
| Шишки | 4.9 ± 0.5 | 3.5 ± 0.8 | – |
| Семена | 13.7 ± 1.6 | – | 21.7 ± 4.0 |
| L (среднезвешенное значение) | 9.86 ± 0.6 | 8.4 ± 0.5 | 12.24 ± 1.5 |
| F | 25.6 ± 4.4 | 17.1 ± 3.3 | 25.5 |
| H | 39.1 | 31.1 | – |
| Запасы золы, г/м2 | |||
| Листья | 4.2 ± 2.0 | 4.14 | 3.63 |
| Ветошь | 0.43 ± 0.3 | – | 1.09 |
| Хвоя | 71.3 ± 16 | 30.17 | 10.84 |
| Детрит | 6.6 ± 2.0 | 5.97 | 2.41 |
| Ветки хвойных | 15.4 ± 1.3 | 10.89 | 5.33 |
| Ветки лиственных | 2.8 ± 2.8 | 5.94 | 7.85 |
| Кора | 8.2 ± 1.4 | 1.34 | 0.45 |
| Древесина | 1.4 ± 0.7 | – | – |
| Шишки | 2.6 ± 0.6 | 3.24 | – |
| Семена | 0.8 ± 0.2 | 1.9 | 0.75 |
| L (сумма) | 114.1 ± 18 | 61.4 ± 11 | 27.5 ± 5.5 |
| LF | 162.1 ± 32 | 121.0 ± 41 | 23.6 ± 23.6* |
| LF2(Н) | 134.6 ± 134.6* | 61.0 ± 61* | – |
| Сумма для подстилки (средняя) | 410.8 | 243.4 | 51.1 |
Итак, подстилки городских еловых насаждений в условиях активной рекреации отличаются упрощенным строением, малой мощностью при повышенных запасах и зольности, а также выраженной пространственной дифференциацией в пределах тессер. Как правило, ферментативные подстилки тяготеют к приствольным повышениям, в пределах которых обнаруживаются более мощные запасы мортмассы по сравнению с другими участками тессер.
Лиственные насаждения. Подстилки обоих изученных лиственных насаждений были определены как деструктивные примитивные – состоящие из одного горизонта (табл. 3). Мощность подстилок очень низкая, не превышает 1 см, что меньше, чем в условиях ботанического сада факультета МГУ им. М.В. Ломоносова, для таких же насаждений [30]. В случае Битцевского парка это вызвано влиянием рекреационной нагрузки. При этом запасы подстилки (рис. 4) в целом соответствуют данным, полученным для других парковых насаждений из липы и березы [32]. Запасы подстилок в липняке значимо больше запасов в березовых насаждениях, что связано не только с тем, что липовые насаждения крупнолистные, а березовые – мелколистные, но и с другими факторами. В условиях более открытого пространства, чем в естественных фитоценозах или фитоценозах ботанического сада МГУ, происходит интенсивный латеральный перенос как опада, так и компонентов подстилки, причем перенос березовых листьев из-за их маленького размера происходит активнее. Поэтому в городских насаждениях подстилки формируются из опада как деревьев-эдификаторов, так и поступающего с ветровым переносом. Во фракционном составе подстилок обоих лиственных насаждений преобладают ветки и листья (рис. 5), причем листьев больше в подстилке березняка. Ветошь почти отсутствует в подстилке липняка, поскольку в условиях повышенного затенения слабо развит травяной ярус. Подстилка березняка, напротив, содержит до 10% ветоши. Подстилки липняка по причине латерального переноса опада посредством ветра из соседнего фитоценоза содержат хвою и шишки, что часто наблюдается в парковых насаждениях [4, 29, 30]. Доля детрита мала, особенно в насаждениях березы, и встречается он не во всех изученных подстилках.
Таблица 3.
Морфологическое строение подстилок лиственных насаждений
| Фитоценоз | Горизонт | Мощность, см | Классификационное положение |
|---|---|---|---|
| Березняк марьянниково-злаковый | L | 0.7 ± 0.1 | Деструктивные среднесопряженные примитивные очень маломощные лиственные |
| Липняк лютиково-живучковый | L | 0.8 ± 0.2 |
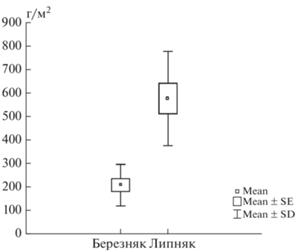
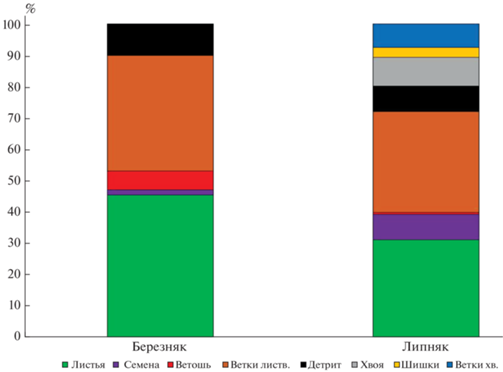
Наиболее зольными компонентами подстилок являются листья (12.6 и 14.9% для березняка и липняка) и ветошь (соответственно 16.5 и 27.0%). В целом зольность горизонта L составляет 9.4% для березняка, 11.4% – для липняка, что мало отличается от горизонтов L подстилок в изученных хвойных насаждениях (табл. 4). Запасы золы в подстилке в целом, а также во фракции листьев и детрита больше в липовых насаждениях, чем в березовых, как за счет общего запаса подстилок, так и за счет величины зольности. При этом общая сумма зольных элементов, накапливаемых горизонтом L подстилки березняка, гораздо меньше, чем аналогичные показатели, полученные для еловых насаждений. Сумма зольных элементов в подстилке липняка превышает сумму зольных элементов в горизонтах L подстилок ельника – кроме приствольных повышений.
Таблица 4.
Свойства компонентов подстилок лиственных насаждений, среднее ± стандартная ошибка
| Горизонт или его компонент | Лиственные насаждения | |
|---|---|---|
| березняк, 70 лет | липняк, 70 лет | |
| Зольность, % | ||
| Листья | 12.6 ± 0.8 | 14.9 ± 1.6 |
| Ветошь | 16.5 ± 1.0 | 27.0* |
| Хвоя | – | 21.2 ± 1,6 |
| Детрит | 14.4* | 20.1* |
| Ветки хвойных | – | 8.3 ± 0.3 |
| Ветки лиственных | 4.1 ± 0.5 | 7.6 ± 0.5 |
| Кора | – | 14.5* |
| Шишки | – | 6.4 ± 0.8 |
| Семена | 5.9* | 9.2 ± 0.6 |
| L (среднезвешенное значение) | 9.4 ± 0.8 | 11.4 ± 1.7 |
| Запасы золы, г/м2 | ||
| Листья | 13.0 ± 2.6 | 32.8 ± 12.2 |
| Ветошь | 2.2 ± 0.6 | 0.33 ± 0.1 |
| Хвоя | – | 10.5 ± 3.6 |
| Детрит | 2.5 ± 1.8 | 7.9 ± 4.8 |
| Ветки хвойных | – | 2.9 ± 1.1 |
| Ветки лиственных | 3.6 ± 0.7 | 14.2 ± 1.8 |
| Кора | – | 0.6 ± 0.2 |
| Шишки | – | 0.8 ± 0.6 |
| Семена | 0.1 ± 0.04 | 3.3 ± 1.9 |
| L (сумма) | 23.2 ± 2.3 | 73.87 ± 14.6 |
Показатели функционирования подстилок. Для характеристики биологического круговорота определяли показатели подстилок, являющиеся индикаторами интенсивности биологического круговорота (табл. 5). Они включают в себя целый ряд характеристик: от типологии подстилок до детрита и легкоразлагаемых компонентов и их запасов.
Таблица 5.
Показатели, отражающие специфику функционирования изученных лесных подстилок
| Показатель | Ельники | Березняки | Липняки | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| городские | естественные | городс-кие | естест-венные | городс-кие | естест-венные | |||||
| ствол | крона | окно | ствол | крона | окно | |||||
| Запасы подстилки, г/м2 | 2600 | 1600 | 350 | 4428 | 3526 | 2056 | 200 | 397 | 590 | 248.5 |
| Соотношение типов подстилок Д : Ф : Г* |
20 : 80 : 0 | 60 : 40 : 0 | 80 : 20 : 0 | 60 : 40 : 0 | 65 : 20 : 15 | 40 : 60 : 0 | 100 : 0 : 0 | 45 : 55 : 0 | 100 : 0 : 0 | 100 : 0 : 0 |
| Запасы детрита, г/м2 | 43.9 | 36.9 | 5.0 | 70.6 | 163.2 | 93.3 | 8.1 | 20.6 | 39.4 | 0 |
| Детрит, % | 3.8 | 4.9 | 2.3 | 10.4 | 20.0 | 20.0 | 4.0 | 6.3 | 8.0 | 0 |
| Запасы ЛРК**, г/м2 | 34.6 | 27.8 | 23.4 | 77.2 | 148.9 | 179.6 | 114.0 | 169.1 | 194.0 | 38.1 |
| ЛРК, % | 2.9 | 3.7 | 10.7 | 11.4 | 18.3 | 38.6 | 56.0 | 51.4 | 33.0 | 15.3 |
| L/(F + H)*** | 0.79 | 0.50 | 0.60 | 0.18 | 0.30 | 0.30 | – | 4.8 | – | – |
| Зола ЛРК, г/м2 | 4.6 | 4.1 | 4.7 | 4.9 | 15.6 | 16.4 | 15.2 | 29.8 | 45.0 | 19.8 |
| Доля золы ЛРК в L, % | 4.0 | 6.7 | 17.1 | 25.2 | 42.9 | 97.0 | 65.0 | 66.5 | 44.0 | 22.1 |
| Зольность L, % | 9.8 | 8.4 | 12.2 | 7.3 | 10.8 | 10.7 | 9.4 | 14.5 | 11.4 | 7.3 |
Такие показатели подстилок, как соотношение горизонтов, запасы, содержание детрита и ЛРК, связаны с интенсивностью круговорота в экосистеме [4, 21, 29, 34], особенно когда их рассматривают не отдельно, а в комплексе друг с другом. Низкие запасы подстилок и их упрощенное строение связаны с высокой скоростью разложения органического вещества подстилки, как и низкие запасы и доля детрита. Повышение скорости разложения подстилок в целом увеличивает скорость оборота органического вещества и зольных элементов, то есть интенсивность биологического круговорота в экосистеме увеличивается.
В целом показатели функционирования подстилок на территории города свидетельствуют о сравнительно высокой интенсивности круговорота в условиях городских парков. Причины заключаются как в микроклиматическом влиянии города на все процессы в экосистеме, в том числе на разложение органического вещества, так и в антропогенном воздействии, что не может не отражаться на активности микроорганизмов в почве и лесной подстилке [44, 46]. Измельчение компонентов подстилок в ходе рекреации также способствует увеличению биологической активности в подстилках [16, 17, 22]. Зольные элементы, поступающие при разложении ветоши видов растений с повышенной зольностью, а также за счет привноса при пылевом загрязнении листвы, включаются в круговорот, оказывая влияние на активность микробиоты и скорость разложения органического вещества. Подстилки лиственных насаждений существенно отличаются от подстилок хвойных такими показателями, как запасы и доля ЛРК, запасы золы в этих компонентах, доля золы ЛРК в общей золе подстилки, а также средневзвешенная зольность горизонта L. Различия свидетельствуют о высокой интенсивности круговорота в лиственных насаждениях по сравнению с хвойными, что наблюдается и в естественных лесных экосистемах. При этом доля детрита в горизонте L практически не отличается для изученных насаждений. Видимо, она очень мала во всех насаждениях в условиях города, особенно при рекреационном воздействии. Несмотря на то, что доля измельченных фракций при рекреации увеличивается [45], образовавшийся детрит, особенно в деструктивных подстилках смешивается при рекреационном воздействии с минеральными горизонтами и уже не входит в состав собственно подстилки, а измельченные фракции активно разлагаются. Общие запасы детрита, видимо, в данном случае детерминированы общими запасами подстилки, поскольку доля детрита в разных насаждениях различается мало.
Строение и запасы подстилок также свидетельствуют об интенсивном разложении органического вещества в лиственных насаждениях города. При этом в пределах елового биогеоценоза запасы подстилок варьируют в большей степени, нежели между биогеоценозами, различающимися по составу деревьев-эдификаторов. В пределах ельника, даже в условиях города, где естественные процессы круговорота снивелированы антропогенным воздействием (атмосферное пылевое загрязнение, рекреация), интенсивность разложения органического вещества, по крайней мере на уровне подстилки, возрастает от приствольных пространств к окнам. Это находит отражение почти во всех изученных структурно-функциональных показателях подстилки – уменьшении мощности и запасов, переходе от ферментативных к деструктивным подстилкам, увеличении доли ЛРК и зольности горизонта L.
Для сравнения привели аналогичные показатели подстилок, рассчитанные ранее для липовых, еловых и березовых насаждений на территории окрестностей УОПЭЦ “Чашниково”, где они не подвержены рекреационному воздействию и существенному атмосферному загрязнению со стороны автотрасс [32, 53]. Развиты эти насаждения также на дерново-подзолистых суглинистых почвах. Березовые и еловые насаждения – естественные, липовые – посадки без ухода (естественных липовых фитоценозов на территории распространения дерново-подзолистых почв практически нет). Наибольшие отличия в функционировании лесных подстилок в городских насаждениях по сравнению с подстилками естественных лесных сообществ выявлены для еловых фитоценозов. Их запасы различаются в 2 раза во всех компонентах тессер. В естественном ельнике встречаются гумифицированные подстилки, больше ферментативных типов лесных подстилок. Содержание детрита в горизонте L подстилок в лесных сообществах города на порядок меньше аналогичного показателя для естественных насаждений. В то же время запасы и доля ЛРК больше в естественных насаждениях, что может быть связано с развитым подлеском и напочвенном покровом и бóльшими объемами поступающего опада.
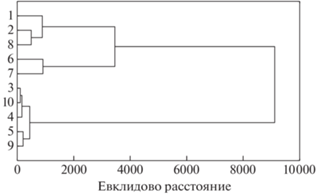
Результаты кластерного анализа (рис. 6) с учетом основных показателей лесных подстилок (общие запасы подстилки, запасы и доля детрита в горизонте L, запасы и доля ЛРК в горизонте L, запасы золы ЛРК, доля золы ЛРК в общих запасах золы горизонта L, зольность горизонта L, доля ферментативных и гумифицированных подстилок среди изученных в фитоценозе или компоненте тессеры) показывают, что выделяются две группы: подстилки хвойных и лиственных насаждений. Подстилки хвойных городских и природных насаждений объединены в одну группу, в пределах которой наблюдаются некоторые различия между подстилками городских и природных фитоценозов. Исключением являются подстилки в межкроновых пространствах городских насаждений, представленные деструктивным типом, как и подстилки лиственных пород. Для подстилок лиственных насаждений разделения по расположению в городских условиях или Подмосковье, а также по принадлежности к дереву-эдификатору не отмечено.
Рис. 6.
Результаты кластерного анализа общих характеристик подстилок изучаемых насаждений. Ельник в Битцевском парке: 1 – ствол, 2 – крона, 3 – окно; 4 – березняк в Битцевском парке, 5 – липняк в Битцевском парке. Ельник в Чашниково: 6 – ствол, 7 – крона, 8 – окно; 9 – березняк в Чашниково, 10 – липняк в Чашниково.
ВЫВОДЫ
1. Особенностями городских древесных насаждений являются упрощенная вертикальная структура и наличие в живом напочвенном покрове, как результат комплексного антропогенного воздействия, луговых и сорно-рудеральных видов, не свойственных естественным фитоценозам аналогичного состава. Упрощение вертикальной структуры фитоценозов ведет к увеличению открытых пространств, что в свою очередь способствует увеличению межбиогеоценозного взаимодействия, которое проявляется в виде активного обмена растительным опадом в системе соседствующих биогеоценозов.
2. Установлены две группы подстилок, распространенных в хвойных и лиственных насаждениях. Для последних обнаруживается максимальная скорость оборота органического вещества и зольных элементов, установленная на основе показателей, отражающих специфику их функционирования – общие запасы подстилки, запасы и доля детрита в горизонте L, запасы и доля ЛРК в горизонте L, отношение запасов горизонта L к сумме запасов нижележащих горизонтов, зольность горизонта L, запасы золы ЛРК и доля золы этих компонентов в общем запасе золы горизонта L – что характерно и для природных территорий.
3. Подстилки хвойных насаждений отличаются повышенным депонированием углерода и зольных элементов, что выражается в их мощности, сложном строении и запасах. В городских условиях основные характеристики подстилок хвойных экосистем, в отличие от лиственных, закономерно и значимо изменяются, реагируя на урбанизацию, что позволяет рассматривать их в качестве индикаторов состояния экосистем мегаполиса.
4. В городских условиях повышенная зольность подстилок как хвойных, так и лиственных насаждений отражает значительное участие механической миграции твердых частиц различного происхождения – минеральные частицы поступают в подстилку как в процессе рекреации, так и с опадом, который поглощает пылевые частицы из атмосферы.
5. Городские парковые насаждения отличаются относительно повышенной интенсивностью биологического круговорота. Емкость круговорота, частично сниженная за счет упрощения структуры фитоценоза, связанного с сокращением ярусов подроста и подлеска, компенсируется включением в круговорот веществ пылевых частиц антропогенного происхождения. Последние могут вовлекаться в процессы почвообразования.
6. На основе анализа показателей биологического круговорота, связанных с лесной подстилкой, установлено, что в условиях города интенсивность биогенной миграции больше, чем в природных экосистемах, что необходимо учитывать при разработке прогноза развития озелененных территорий и планирования охранных мероприятий природной компоненты города.
Список литературы
Базилевич Н.И., Титлянова А.А. Биотический круговорот на пяти континентах: азот и зольные элементы в природных наземных экосистемах. Новосибирск: Изд-во СО РАН, 2008. 376 с.
Бганцова В.А., Бганцов В.Н., Соколов Л.A. Влияние рекреационного лесопользования на почву // Природные аспекты рекреационного использования леса. М.: Наука, 1987. С. 71–85.
Богатырев Л.Г. О классификации лесных подстилок // Почвоведение. 1990. № 3. С. 118–127.
Богатырев Л.Г., Телеснина В.М., Семенюк О.В., Бенедиктова А.И. Динамика морфологии и химических свойств лесной подстилки в ходе естественного постагрогенноголесовосстановления и ее отражение напочвенным покровом // Вестник Моск. ун-та. Сер. 17, почвоведение. 2020. № 3. С. 3–11.
Влияние рекреации на лесные экосистемы и их компоненты. М., 2004. 288 с.
Горбунов А.С., Цветков П.А. Естественное возобновление в рекреационных сосняках зеленой зоны г. Красноярска // Хвойные бореальной зоны. 2009. № 2. С. 244–248.
Дробышев Ю.И. Устойчивость рекреационных лесных фитоценозов в связи с их структурными особенностями. Автореф. дис. … канд. биол. наук. М., 2000. 17 с.
Жидков А.Н. Трансформации ландшафта и почв Битцевского лесопарка // Вестник МГУЛ – Лесной вестник. 2016. № 5. С. 117–121.
Земсков Ф.И. Детритогенез в условиях лесных биогеоценозов урбанизированных территорий. Автореферат дис. … канд. биол. наук. М., 2021. 25 с.
Кавеленова Л.М., Здетоветский А.Г., Огневенко А.Я. К специфике содержания зольных веществ в листьях древесных растений в городской среде в условиях лесостепи (на примере Самары) // Химия растительного сырья. 2001. № 3. С. 85–90.
Каплан Б.М., Полякова Г.А. Сныть обыкновенная в парковых насаждениях // Лесное хозяйство. 1998. № 4. С. 31–32.
Киселева В.В., Ломов В.Д., Обыденников В.И., Титов А.П. История развития и современное состояние сосняков Алексеевской Рощи // Лесоведение. 2010. № 3. С. 42–52.
Классификация и диагностика почв России. Смоленск: Ойкумена, 2004.
Кузнецов В.А. Почвы и растительность парково-рекреационных ландшафтов Москвы. Автореф. дис. … канд. биол. наук. М., 2015. С. 1–26.
Лазарева И.П., Морозова Р.М. Особенности рекреационной деградации почв в Карелии. Науч. Докл. АН СССР. Карел. фил. ин-та леса. Петрозаводск, 1987. 24 с.
Лысиков А.Б., Судницына Т.Н. // Влияние рекреации на почву лиственных насаждений Серебряноборского опытного лесничества // Лесоведение. 2008. № 3. С. 47–56.
Марфенина O.E., Жевелева Е.М., Офицерова О.В., Розина М.С., Зарифова З.А. Влияние нормированных рекреационных нагрузок на свойства бурых лесных почв // Вестник Моск. ун-та. Сер. 17, почвоведение. 1984. № 3. С. 52–58.
Мозолевская Е.Г. Факторы нарушения устойчивости лесов Москвы и ближнего Подмосковья и их значение // Влияние рекреации на лесные экосистемы и их компоненты. РАН. ОБН. Институт лесоведения. М., 2004. С. 4–37.
Ниценко А.А. Об изучении экологической структуры растительного покрова // Ботанический журн. 1969. Т. 54. № 7. С. 1002–1013.
Полякова Г.А. Флора и растительность старых парков Подмосковья М.: Наука, 1992. 224 с.
Родин Л.Е., Базилевич Н.И. Динамика органического вещества и биологический круговорот азота и зольных элементов в основных типах растительности земного шара. М.–Л.: Наука, 1965. 265 с.
Рысин Л.П. Рекреационное лесопользование; научные и практические аспекты // Лесобиологические исследования на северо-западе таежной зоны России: итоги и перспективы. Мат-лы науч. конф. Петрозаводск: КарНЦ РАН, 2007. С. 83–94.
Рысин Л.П., Абатуров А.В., Савельева Л.И., Меланхолин П.Н., Полякова Г.А., Рысин С.Л. Динамика и устойчивость рекреационных лесов. М.: Т-во научных изданий КМК, 2006. 165 с.
Савенко В.С. Геохимический аспекты устойчивого развития. М.: ГЕОС, 2003. 179 с.
Самсонова С.Ю. Рельеф в управлении особо охраняемыми природными территориями г. Москвы. Автореф. дис. … канд. геогр. наук. М., 2013. 24 с.
Сапожников А.П. Лесная подстилка – номенклатура, классификация, индексация // Почвоведение. 1984. № 5. С. 96–105.
Семенюк О.В., Богатырев Л.Г., Ваганова М.А. Характеристика подстилок парковых насаждений исторических ландшафтов на примере музея-усадьбы “Архангельское” // Бюл. Москов. о-ва испытателей природы. Отд. Биол. 2017. Т. 122. Вып. 5. С. 37–49.
Семенюк О.В., Ваганова М.А. Характеристика растительного покрова как важнейшая составляющая комплексных почвенно-экологических исследований исторических парков // Бюл. Моск. о-ва испытателей природы. 2016. Т. 121. Вып. 4. С. 32–42.
Семенюк О.В., Телеснина В.М., Богатырев Л.Г., Бенедиктова А.И. О возможности использования особенностей структурно-функциональной организации подстилок для оценки интенсивности круговорота в городских насаждениях (на примере Москвы) // Почвоведение. 2021. № 5. P. 592–605. https://doi.org/10.31857/S0032180X21050178
Семенюк О.В., Телеснина В.М., Богатырев Л.Г., Бенедиктова А.И. Оценка влияния ухода за городскими насаждениями на свойства подстилок в условиях мегаполиса // Вестник Моск. ун-та. Сер. 17, почвоведение. 2021. № 2. С. 3–10.
Телеснина В.М., Семенюк О.В., Богатырев Л.Г. Свойства лесных подстилок во взаимосвязи с напочвенным покровом в лесных экосистемах подмосковья (на примере УОПЭЦ “Чашниково”) // Вестник Моск. ун-та. Сер. 17, почвоведение. 2017. № 4 С. 11–20.
Телеснина В.М., Семенюк О.В., Богатырев Л.Г., Бенедиктова А.И. Особенности напочвенного покрова и лесных подстилок в искусственных липовых насаждениях в зависимости от характера ухода // Вестник Моск. ун-та. Сер. 17, почвоведение. 2018. № 2. С. 3–11.
Титлянова А.А., Шибарева С.В. Подстилки в лесных и травяных экосистемах. Новосибирск: Изд-во Сиб. отд. РАН, 2012. 134 с.
Чертов О.Г. Экология лесных земель. Л.: Наука, 1981. 192 с.
Экология города. М.: Научный мир, 2004. 624 с.
Ясенево: История и современность, 1997. 144 с.
Bae J., Ryu Y. High soil organic carbon stocks under impervious surfaces contributed by urban deep cultural layers. // Landscape and Urban Planning. 2020. V. 204. P. 938–953. https://doi.org/10.1016/j.landurbplan.2020.103953
Bavec Š., Gosar M., Biester H., Grčman H. Geochemical investigation of mercury and other elements in urban soil of Idrija (Slovenia) // J. Geochem. Explorat. 2015. V. 154. P. 213–223. https://doi.org/10.1016/j.gexplo.2014.10.011
Beroigui M., Naylo A., Walczak M. et al. Physicochemical and microbial properties of urban park soils of the cities of Marrakech, Morocco and Toruń, Poland: Human health risk assessment of fecal coliforms and trace elements // Catena. 2020. V. 194. P. 673–677.
Curiel J., Yuste D.D., Flores-Rentería D., García-Angulo D., Heres A.M., García-Angul C., Petritan M., Petritan I.C. Cascading effect associated with climate-change-induced conifer mortality in mountain temperate forests result in hot-spots of soil CO2 emissions // Soil Biol. Biochem. 2019. V. 133. P. 50–59. https://doi.org/10.1016/j.soilbio.2019.02.017
Dehbandi R., Aftabi A. Geochemical provenance of soils in Kerman urban areas, Iran: Implications for the influx of aeolian dust // Aeolian Res. 2016. V. 21. P. 109–123. https://doi.org/10.1016/j.aeolia.2016.04.004
Huang R., Lan T., Song X., Juan Li J. Soil labile organic carbon impacts C:N:P stoichiometry in urban park green spaces depending on vegetation types and time after planting // Appl. Soil Ecol. 2021. V. 163. P. 926–958. https://doi.org/10.1016/j.apsoil.2021.103926
Kaushal S.S., Wood K.L., Galella J.G. et al. Making ‘chemical cocktails’ – Evolution of urban geochemical processes across the periodic table of elements // Appl. Geochem. 2020. V. 119. № 3. https://doi.org/10.1016/j.apgeochem.2020.104632
Killham K. Soil Ecology. Cambridge University Press, Cambridge. 1994. 242 p.
Kuznetsov V.A., Ryzhova I.M., Stoma G.V. Transformation of forest ecosystems in Moscow megapolis under recreation impacts // Eurasian Soil Science. 2019. V. 52. № 5. P. 584–592. https://doi.org/10.1134/S1064229319050065
Liddle M. Recreation Ecology: The Ecological Impact of Outdoor Recreation and Ecotourism. London: Chapman Hall, 1997. 639 p.
Liu Q. Yin R., Scheu S. Nitrogen addition and plant functional type independently modify soil mesofauna effects on litter decomposition // Soil Biol. Biochem. 2021. V. 157. P. 76–81. https://doi.org/10.1016/j.soilbio.2021.108340
Liu R., Wang D. C:N:P stoichiometric characteristics and seasonal dynamics of leaf-root-litter-soil in plantations on the loess plateau // Ecol. Indic. 2021. V. 127(1). P. 107772. https://doi.org/10.1016/j.ecolind.2021.107772
Lu Ch., Kotze D.J., Setälä H.M. Evergreen trees stimulate carbon accumulation in urban soils via high root production and slow litter decomposition // Sci. Total Environ. 2021. V. 774. P. 129–145. https://doi.org/10.1016/j.scitotenv.2021.145129
Melliger R.L., Rusterholz H.P., Baur B. Ecosystem functioning in cities: Combined effects of urbanisation and forest size on early-stage leaf litter decomposition of European beech (Fagussylvatica L.) // Urban Forestry Urban Greening. 2017. V. 28. P. 88–96. https://doi.org/10.1016/j.ufug.2017.10.009
Pecina V., Brtnický M., Baltazár T. et al. Human health and ecological risk assessment of trace elements in urban soils of 101 cities in China: A meta-analysis // Chemosphere. 2021. V. 267. P. 129–215. https://doi.org/10.1016/j.chemosphere.2020.129215
Seco Pon J.P., Becherucci M.E. Spatial and temporal variations of urban litter in Mar del Plata, the major coastal city of Argentina // Waste Management. 2012. V. 32. № 2. P. 343–348.
Semenyuk O.V., Telesnina V.M., Bogatyrev L.G., Benediktova A.I., Kuznetsova Ya.D. Assesment of intra-biogeocenotic variability of forest litters and dwarfshrub-herbaceous vegetation in spruce sand // Eurasian Soil Science. 2020. V. 53. № 1. P. 31–43. https://doi.org/10.1134/S1064229320010135
Tang J., Wang W., Feng J. et al. Urban green infrastructure features in-fluence the type and chemical composition of soil dissolved organic matter // Sci. Total Environ. 2021. V. 764. P. 144–240. https://doi.org/10.1016/j.scitotenv.2020.144240
Trifonova T.A., Zabelina O.N. Changes in the biological activity of heavy-metal- and oil-polluted soils in urban recreation territories // Eurasian Soil Science. 2017. № 4. P. 483–490. https://doi.org/10.1134/S1064229317040147
Wang W., Hu K., Tao J. Mechanical fragmentation of leaf litter by fine root growth contributes greatly to the early decomposition of leaf litter // Global Ecology and Conservation. 2021. V. 26. P. 456–464. https://doi.org/10.1016/j.gecco.2021.e01456
Xu D., Gao B., Song S., Peng W. Release risk assessment of trace metals in urban soils using in-situ DGT and DIFS model // Sci. Total Environ. 2019. V. 694. P. 624–653. https://doi.org/10.1016/j.scitotenv.2019.133624
Дополнительные материалы отсутствуют.