

Радиационная биология. Радиоэкология, 2020, T. 60, № 3, стр. 229-238
НЕКОТОРЫЕ АСПЕКТЫ ТРАНСЛЯЦИИ РАДИАЦИОННО-ИНДУЦИРОВАННЫХ ИЗМЕНЕНИЙ ПРИ “ЭФФЕКТЕ СВИДЕТЕЛЯ”
В. В. Петушкова 1, *, И. И. Пелевина 1, И. Н. Когарко 1, Е. А. Нейфах 1, Б. С. Когарко 1, О. В. Ктиторова 1
1 Федеральный исследовательский центр химической физики им. Н.Н. Семёнова РАН
Москва, Россия
* E-mail: pele@chph.ras.ru
Поступила в редакцию 16.09.2019
После доработки 16.09.2019
Принята к публикации 18.09.2019
Аннотация
В работе приведены первые результаты исследований и наблюдения в рамках комплексного проекта по оценке радиационно-индуцированных “немишенных эффектов”. Дистанционный “эффект свидетеля” – наименее изученный с точки зрения проявлений и механизмов феномен, относящийся к “немишенным эффектам”. Международная лабораторная практика в области данного явления начиналась с демонстрации “эффекта свидетеля” на рыбах и иных аквакультурах. В нашем эксперименте у необлученных мышей, находившихся на протяжении 3 мес. в контакте с облученными (3 Гр) особями, выявлены признаки патологий, сходных с радиационно-индуцированными. Отмечались изменение поведенческих реакций, статистически значимая тенденция к уменьшению веса селезенки (r = –0.416; p = 0.048), повышению площади алопеции (r = –0.631; p = 0.001) и иные аномалии. Обсуждение возможных механизмов данного явления связывается авторами как с теорией радиотоксинов А.М. Кузина (1966), так и с трактовкой гипотез, исследуемых К. Мазерсил (Carmel E. Mothersill, 2010–2019). Высказывается предположение о важной роли производных липоперекисей в формировании изучаемого эффекта. Полученные данные свидетельствуют о необходимости проведения дальнейших исследований лучевого “эффекта свидетеля” с использованием молекулярно-клеточных методических подходов.
Исследование направлено на изучение новых аспектов явления, известного в радиобиологии как “немишенные эффекты”, которые заключаются в способности передачи изменений от облученных клеток (т.е. клеток-мишеней для радиационного воздействия) к необлученным. В последние годы очередное открытие в области “немишенных эффектов” выявило “дистанционно индуцируемые эффекты свидетеля”, именуемые в зарубежных исследованиях “эффектами свидетеля на уровне организма” (radiation-induced inter-organism bystander effects). Было показано, что после воздействия радиации изменения и повреждения могут передаваться не только от клетки к клетке, но и от целого облученного организма к необлученному, включая высшие организмы и, предположительно, человека [1].
В настоящее время за рубежом проводятся систематические исследования в области “немишенных эффектов”. Большой вклад в развитие направления принадлежит В.Ф. Моргану [2] и Д.Б. Литтлу [3]. На базе канадского университета Макмастер проводятся регулярные экспериментальные работы по изучению “эффекта свидетеля” in vitro и in vivo [1]. Именно с исследовательской группой К. Мазерсил (университет г. Макмастера) связаны первые успехи в области изучения дистанционно-индуцируемых радиационных “эффектов свидетеля”, передающихся между организмами. После проведения К. Мазерсил цикла исследований (2010–2019) на рыбах – радужной форели и Danio Rerio (Zebrafish), результаты которых окончательно подтвердили гипотезу о возможности передачи радиационно-индуцированных изменений от облученного организма к необлученному, было доказано, что с дистанционными “эффектами свидетеля” необходимо считаться при оценке ущерба и рисков облучения в малых дозах [4, 5]. Первоначально выявленные на одноклеточных эти явления вслед за тем были показаны на различных беспозвоночных и позвоночных животных [4–8].
Разработки в области “немишенных эффектов” в последние годы ведутся в различных странах мира: Канада, США, Великобритания, Германия, КНР, Япония, Франция, Финляндия, Индия, Южная Корея, Иран. Возросший интерес к различным проявлениям “немишенных эффектов” связан с необходимостью оценки рисков и последствий техногенных катастроф в тех районах, где речь идет о длительном облучении биологических объектов в малых дозах. После аварии на атомной станции Фукусима к исследованиям активно подключилась Япония и, сравнительно недавно, Южная Корея. [9, 10]. В недавние годы имела место практика международных экспедиций, проводимых с целью получения и обработки материалов для изучения “немишенных эффектов” в регионах, подвергшихся радиационному загрязнению, в том числе – Чернобыльской зоны [11].
Говоря о российских исследованиях, вплотную подошедших к изучению дистанционно-индуцируемых радиационных “немишенных эффектов”, передающихся между организмами, необходимо прежде всего упомянуть гипотезу о “радиотоксичности” чл.-корр. АН СССР А.М. Кузина [12, 13], которая вносила существенные дополнения в классические представления Н.В. Тимофеева-Ресовского о воздействии радиации на биологические объекты: теорию мишени и принцип попадания [14]. В последующие годы идеи А.М. Кузина широкого развития в нашей стране не получили, его труды практически неизвестны за рубежом.
Систематических и комплексных исследований в области “немишенных эффектов” в России по-прежнему мало, среди недавних необходимо отметить работы исследовательских групп И.И. Пелевиной [15–17] (ИХФ им. Семёнова РАН, Москва) и Б.П. Суринова [18] (МРНЦ им. А.Ф. Цыба – филиал ФГБУ “НМИРЦ” Минздрава России, Обнинск). Следует сказать о различиях понятийного аппарата, используемого в западных и отечественных исследованиях. Определение “дистанционный эффект свидетеля” у нас включает эффекты, передающиеся не только между организмами, но и на уровне одного организма, возникающие в отдаленных по расстоянию клетках, органах и тканях, не являющихся прилежащими к облученным [13, 15].
В недавних исследованиях группы И.И. Пелевиной была создана цитогенетическая модель оценки рассматриваемого феномена, основанная на изучении аберраций хромосом в клетках периферической крови человека. Продемонстрирована схема генотоксического действия лимфоцитов больных злокачественными новообразованиями на клетки крови здоровых индивидуумов в среде (in vitro), имитирующей естественную, а тем самым показана возможность дистанционной передачи подобных изменений. Та же модель оказалась применимой к дистанционно-индуцируемым радиационным “немишенным эффектам”. В дальнейшем, на примере абскопальных эффектов (от слова abscopal – не в месте облучения) Пелевиной и соавт. были показаны in vivo дистанционно-индуцированные “немишенные эффекты”, возникающие в пределах одного организма на лимфоцитах прошедших радиотерапию больных [16, 17]. В связи с актуальностью темы лучевого “эффекта свидетеля”, И.И. Пелевиной был запланирован комплекс исследований на животных с целью изучения явлений передачи радиационных изменений от облученного организма к необлученному.
В данной работе представлены первые результаты исследований, выполненных на мелких млекопитающих, демонстрирующие изменения, возникающие у необлученных мышей, находящихся в контакте с облученными животными.
МАТЕРИАЛЫ И МЕТОДИКА
Исследование выполнено на 25 беспородных мыша-самках в возрасте от 3 до 4 мес. Все животные получены из питомника “Столбовая”, до опыта содержались группами по 10 особей в условиях вивария на сертифицированном корме, сбалансированном пищевом рационе при обычном водном режиме и естественном освещении (осенне-зимнего периода). Применялась подстилка из натурального сертифицированного гипоаллергенного материала (древесная стружка). До начала эксперимента мышей выдерживали не менее 2 нед. в стандартных металлических сетчатых клетках с целью адаптации к условиям лабораторного вивария. В опыте использовали здоровых животных, не подвергавшихся ранее другим экспериментальным воздействиям. Группы формировали методом сплошной выборки. При проведении экспериментальных процедур соблюдались соответствующие международные правила работы с животными [19].
Для облучения десять мышей были помечены с помощью красителя и рассажены по пять особей в специальные пластиковые боксы с индивидуальными секциями. Мыши были подвергнуты радиационному воздействию в дозе 3 Гр на установке: γ-терапевтический аппарат РОКУС-АМ (АО “Равенство”, Санкт-Петербург, Россия, зав. № 71); источник γ-излучения: радионуклид 60Со; энергия γ-излучения: 1.25 МэВ; радиационный выход: мощность экспозиционной дозы (МЭД) на расстоянии 75 см от источника на 24.09.2018 г. в центре поля 10 × 10 см равна 64.7 Р/мин. Облучение производилось с одного направления – сверху вниз, при этом тело каждой мыши было облучено полностью со стороны спины, с достижением эффекта равномерности.
Контейнер с мышами помещали на лечебном столе γ-аппарата таким образом, чтобы радиационное поле 22 × 22 см охватывало с запасом весь контейнер. При этом расстояние от источника до объекта (поверхность спины мыши) устанавливали 75 см. Расчет дозы (3 Гр) проводили на глубину 1 см (98% от дозы в максимуме). При этом максимум дозы (100%) находился на глубине 0.5 см (3.06 Гр) и, если взять толщину объекта (мыши) 2.0 см, то на выходе из тела мыши доза составила 93.1%, т.е. 2.8 Гр.
После облучения мыши (n = 10) были возвращены в виварий и рассажены в клетки для свободного контакта с необлученными особями (n = = 10) по следующей схеме: использовали две стандартные металлические клетки, в каждой из которых находилось по пять облученных мышей и по пять необлученных (мыши-“свидетели”). Облученные и необлученные мыши могли беспрепятственно контактировать друг с другом. В остальном условия содержания соответствовали тем, которые были до радиационного воздействия. Таким образом, 20 мышей (беспородные мыши-самки), участвовавшие в эксперименте, содержались в двух самостоятельных клетках при равных условиях, что исключало случайности (рис. 1, 2). Эксперимент продолжался 3 мес. За мышами велось постоянное наблюдение.
Рис. 1.
Схема рассадки мышей по клеткам № 1–2. Этап 1: пять мышей подвергаются облучению в дозе 3 Гр, в качестве “мышей-свидетелей” используются пять здоровых мышей из той же группы без облучения.
Fig. 1. Mice irradiation scheme for the сages № 1–2. In the first stage, five mice were exposed to γ-radiation at a dose of 3 Gy.

Рис. 2.
Схема рассадки мышей по клеткам № 1–2. Этап 2: пять облученных мышей (3 Гр) содержатся совместно с необлученными мышами (мышами-“свидетелями”) на протяжении 3 мес.
Fig. 2. Mice irradiation scheme for the сages № 1–2. In the second stage, five irradiated (3 Gy) mice were kept together with non-irradiated companions for three months.

Третью контрольную группу составили пять мышей-самок в возрасте от 3 до 4 мес., содержавшихся отдельной группой из пяти особей в условиях вивария на том же стандартном сбалансированном пищевом рационе и водном режиме при естественном световом режиме в металлической клетке с сертифицированной подстилкой из древесных стружек. Сроки содержания контрольной группы в условиях вивария соответствовали датировкам по основной группе.
На протяжении всего эксперимента вели контроль над изменением поведения животных. По истечении срока 3 мес. мышей забивали. У мышей трех исследованных групп оценивали вес тела, вес селезенки, состояние волосяного покрова, некоторые признаки изменений внутренних органов и поведенческих реакций. Статистическую обработку результатов проводили с помощью программного пакета STATISTICA-10. Значимость различий между группами оценивали с помощью непараметрического U-критерия Манна–Уитни и точного критерия Фишера. Проводили также регрессионный анализ.
РЕЗУЛЬТАТЫ
Изменение поведенческих реакций
После определенного периода адаптации, примерно через 2 нед. после начала эксперимента, были зафиксированы перемены в поведении грызунов, которые формировали беспородные мыши-самки. Отмечались изменения в поведении как облученных, так и необлученных мышей-“свидетелей” по сравнению с нормой (контрольные особи) и тем состоянием, которое было отмечено у животных до начала эксперимента. Особей с данными породными характеристиками и происхождением (питомник Столбовая) отличает достаточно стабильная нервная система, для них характерно уравновешенное и спокойное поведение. Подопытные животные, как облученные, так и мыши-“свидетели”, на протяжении всего периода наблюдения (около 3 мес.) отчетливо проявляли повышенную активность и нервозность по сравнению с контрольными мышами и тем, что должно быть в обычных условиях.
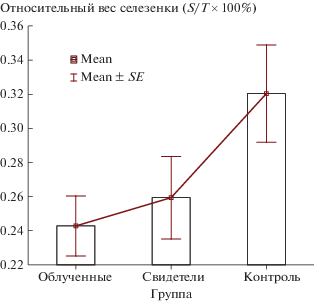
Изменения в относительном весе селезенки
Результаты оценки относительных весов селезенки в трех группах мышей представлены в табл. 1 и на рис. 3. Из расчета удалены два индивидуальных значения, существенно выходящих за пределы 95%-ного доверительного интервала. В группе облученных мышей рассматриваемый показатель был существенно ниже такового у животных контрольной группы (р = 0.046 по тесту Манна–Уитни). Совершенно очевидно, что между облученными мышами и “свидетелями” нет существенных различий в индексе массы селезенки (р = 0.659), при этом наблюдается явная тенденция к уменьшению рассматриваемого показателя у “мышей-свидетелей” по сравнению с контролем, хотя значимых различий не выявлено: р = 0.182 (табл. 1). Наиболее наглядно полученные результаты представлены на рис. 3. Условное ранжирование полученных данных позволило провести регрессионный анализ зависимости рассматриваемого показателя от воздействия радиационного фактора (необлученные мыши – 0, мыши-“свидетели” – 1, облученные – 2). Рассматриваемая зависимость значимо описывалась линейным уравнением вида Y = 0.309 ± 0.024 – 0.036 ± 0.017x (r = –0.416; p = = 0.048).
Таблица 1.
Относительный вес селезенки животных (S/T × 100)* для трех исследованных групп беспородных мышей-самок Table 1. Relative values of spleen weights (S/T × 100)* for the three groups of animals studied: irradiated mice (3 Gy), bystander mice (0 Gy) and the control group (0 Gy)
| Группы животных | ||||||||
|---|---|---|---|---|---|---|---|---|
| облученные 3 Гр (n = 10) irradiated | свидетели 0 Гр (n = 10) bystander | контроль 0 Гр (n = 5) control | ||||||
| номер животного | индивидуальные значения веса сел. S | среднее значение (M ± SE) | номер животного | индивидуальные значения веса сел. S | среднее значение (M ± SE) | номер животного | индивидуальные значения веса сел. S | среднее значение (M ± SE) |
| O1 | 0.2667 | 0.243 ± 0.018 | M1 | 0.2308 | 0.2594 ± 0.024 | К1 | 0.2905 | |
| O2 | 0.2656 | M2 | 0.3698 | K2 | 0.2684 | |||
| O3 | 0.1686 | M3 | 0.3515 | K3 | 0.3324 | |||
| O4 | 0.1881 | M4 | 0.2852 | K4 | 0.2842 | |||
| O5 | 0.2985 | M5 | 0.2111 | K5 | 0.4265 | 0.3204 ± 0.029 | ||
| O6 | 0.2085 | M6 | 0.1508 | |||||
| O7 | 0.28 | M7 | 0.1863 | |||||
| O8 | 2.57** | M8 | 1.347** | |||||
| О9 | 0.1968 | M9 | 0.2788 | |||||
| O10 | 0.3125 | M10 | 0.2701 | |||||
Примечание. Выявлено статистически значимое различие по относительному весу селезенки между группами “облученные мыши” и “контрольные мыши” (р = 0.046 по тесту Манна–Уитни). Совершенно очевидно, что между облученными мышами и “свидетелями” нет существенных различий в индексе массы селезенки (р = 0.658), при этом наблюдается явная тенденция к уменьшению рассматриваемого показателя у “мышей-свидетелей” по сравнению с контролем, хотя значимых различий не выявлено (р = 0.182). Условное ранжирование полученных данных позволило провести регрессионный анализ зависимости рассматриваемого показателя от воздействия радиационного фактора (необлученные мыши – 0, мыши-“свидетели” – 1, облученные – 2). Рассматриваемая зависимость значимо описывалась линейным уравнением вида Y = 0.309 ± 0.024 – 0.036 ± 0.017X (r = –0.416; p = 0.048). * S/T, S – вес селезенки (spleen weight), T – вес тела (body weight). ** Патологические значения весов, удаленные из расчета.
Рис. 3.
Относительный вес селезенки у мышей исследованных групп (S/T × 100%, S – вес селезенки, T – вес тела). Средние значения (M ± SE): облученные – 0.243 ± 0.018; мыши-“свидетели” – 0.2594 ± 0.024; контрольная группа – 0.3204 ± 0.029.
Fig. 3. Relative weights of spleen for the three groups of mice studied. (S/T × 100%, S – spleen weight, T – body weight).
Изменение состояния волосяного покрова, алопеция
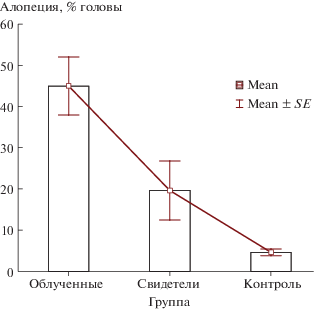
Через 1 мес. после начала эксперимента у подопытных мышей, как облученных, так и необлученных, появились отчетливые признаки алопеции. Этот процесс начинался с зоны, примыкающей к носу, и постепенно распространялся по направлению к верхней части головы. Алопеция проявлялась в полной опоясывающей потере шерсти на охваченных процессом участках головы и носила отчетливо патологический характер. У отдельных особей появлялись дополнительные очаги на верхней части головы и/или на теле, имевшие почти правильную геометрическую форму. На лишенных шерсти участках имела место эритема. Содержание мышей в двух не связанных между собой клетках при рассадке по схеме пять облученных мышей и пять мышей-“свидетелей” в каждой клетке на протяжении нескольких месяцев исключает случайные причины возникновения указанного признака.
Результаты оценки волосяного покрова в трех группах мышей представлены в табл. 2 и на рис. 4. Уровень алопеции у облученных мышей существенно превышал таковой у контрольных животных (р = 0.0026 по тесту Манна–Уитни). Между облученными животными и “свидетелями” также выявлены существенные различия по рассматриваемому показателю (р = 0.021), при этом наблюдается явная тенденция к увеличению уровня алопеции у “мышей-свидетелей” по сравнению с контролем, хотя значимых различий не выявлено (р = 0.178). Наиболее наглядно полученные результаты представлены на рис. 4. Условное ранжирование полученных данных позволило провести регрессионный анализ зависимости рассматриваемого показателя от воздействия радиационного фактора (необлученные мыши – 0, мыши-свидетели – 1, облученные – 2). Рассматриваемая зависимость значимо описывалась линейным уравнением вида Y = 1.663 ± 7.590 + + 20.914 ± 5.367X (r = –0.631; p = 0.001).
Таблица 2.
Алопеция у мышей трех групп: облученные, “свидетели” и контроль. Процент головы, пострадавший от потери шерсти Table 2. Alopecia. The scale of hair loss for the three groups of mice studied: irradiated (3 Gy), bystander (0 Gy), control group (0 Gy). Percentage of head affected by alopecia
| Группы животных | ||||||||
|---|---|---|---|---|---|---|---|---|
| облученные 3 Гр (n = 10) | свидетели 0 Гр (n = 10) | контроль 0 Гр (n = 5) | ||||||
| номер животного | % головы, пострадавший от алопеции | среднее значение (M ± SE) | номер животного | % головы, пострадавший от алопеции | среднее значение (M ± SE) | номер животного | % головы, пострадавший от алопеции | среднее значение (M ± SE) |
| O1 | 33.3 | 44.970 ± 7.051 | М1 | 33.3 | 19.620 ± 7.176 | К1 | 3.3 | 4.620 ± 0.808 |
| O2 | 33.3 | М2 | 16.6 | К2 | 6.6 | |||
| O3 | 83.3 | М3 | 16.6 | К3 | 6.6 | |||
| O4 | 33.3 | М4 | 16.6 | К4 | 3.3 | |||
| O5 | 50 | М5 | 16.6 | К5 | 3.3 | |||
| O6 | 16.6 | М6 | 50 | |||||
| O7 | 33.3 | М7 | 66.6 | |||||
| O8 | 83.3 | М8 | 33.3 | |||||
| O9 | 33.3 | М9 | 16.6 | |||||
| O10 | 50 | М10 | 6. 6 | |||||
Примечание. Выявлено статистически значимое различие по уровню алопеции между группами “облученные мыши” и “контрольные мыши” (р = 0.002 по тесту Манна–Уитни). Между облученными мышами и “свидетелями” также определены существенные различия по рассматриваемому показателю (р = 0.021), при этом наблюдается явная тенденция к увеличению рассматриваемого показателя у “мышей-свидетелей” по сравнению с контролем, хотя значимых различий не выявлено (р = 0.178).
Рис. 4.
Показатели алопеции для трех групп животных: облученные (3 Гр), “свидетели” (0 Гр), контрольная группа (0 Гр). Средние значения (M ± SE): облученные – 44.970 ± 7.051; мыши-“свидетели” – 19.620 ± 7.176; контрольная группа – 4.620 ± 0.808.
Fig. 4. Alopecia scale for the three groups of animals studied: irradiated (3 Gy), bystander (0 Gy) and the control group (0 Gy). Percentage of head affected by hair loss.
Другие патологии
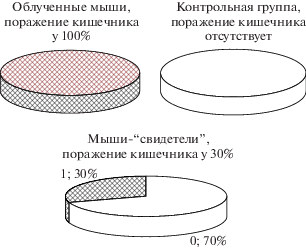
Помимо снижения относительной массы селезенки и патологической алопеции, выявленной у облученных мышей и мышей-“свидетелей” данной экспериментальной группы, в обеих категориях исследованных животных отмечались изменения кишечника. Представляется необходимым упомянуть об этих признаках, так как кишечник наряду с селезенкой относится к наиболее радиочувствительным органам [12, 13]. Выявлены статистически значимые различия между облученной и контрольной группами животных по показателю, характеризующему изменения кишечника (р = 3.3 × 10–4 по точному критерию Фишера): налет на кишечнике сложной этиологии обнаружен у 10 из 10 облученных животных (100%) и отсутствует у мышей контрольной группы (рис. 5). По данному критерию отмечались и существенные различия между облученными животными и мышами-свидетелями (р = 0.003). Хотя в группе мышей-“свидетелей” выявлены особи с рассматриваемыми изменениями (три из 10, или 30%), при таких объемах выборок отличия от контроля не обнаружено (р = 0.51).
Рис. 5.
Наличие (отсутствие) признаков поражения кишечника (налет сложной этиологии) в изучаемых группах мышей: облученные (3 Гр), свидетели (0 Гр) и контроль (0 Гр).
Fig. 5. The presence of signs of bowel damage (plaque of complex etiology) in the groups of mice studied: irradiated (3 Gy), bystander (0 Gy) and control (0 Gy).
ОБСУЖДЕНИЕ
Таким образом, продемонстрирована реальность индукции “эффекта свидетеля” в необлученном организме в результате его контакта с организмом, подвергшимся радиационному воздействию. Полученные результаты приводят к формулировке нескольких вопросов, ответ на которые приблизит к пониманию механизмов радиационно-индуцированных “байстендерных” эффектов, проявившихся в опыте. “Возможны ли условия, при которых признаки радиационно-индуцированных изменений организма возникнут без облучения?” “Каковы “транспортные” механизмы передачи радиационно-индуцированных изменений от облученных организмов к необлученным?” Для объяснения радиационно-индуцированного “эффекта свидетеля” сегодня предлагается три механизма: 1) непосредственный контакт и передача повреждающих факторов; 2) выделение токсических соединений, которые могут вызывать отдаленные по расстоянию эффекты облученных клеток; 3) действие биофотонов [1].
В настоящее время проблематике “немишенных эффектов” посвящен огромный массив публикаций; сам термин возник поздно, на рубеже XX–XXI веков в результате деятельности общеевропейского проекта “Радинстаб”. По свидетельству К. Мазерсилл и др., к этому моменту следовали периодические повторные открытия радиационного “эффекта свидетеля”, но “тема в определенные периоды появлялась лишь для того, чтобы исчезнуть вновь” [6]. Практически неизвестным за рубежом является тот факт, что важные механизмы действия современных “немишенных эффектов” с точки зрения клеточных и биохимических процессов были показаны в 1960-е годы в экспериментах А.М. Кузина и нашли отражение в его трудах, посвященных действию радиотоксинов (РТ).
Характеризуя этапы развития “типичной формы” лучевой болезни человека, возникающей при дозах тотального облучения 1–10 Гр, А.М. Кузин отмечал, что уже в момент радиационного воздействия во всех тканях экспонированного организма возникают свободные радикалы воды и органических соединений. В присутствии кислорода они дают такие токсические вещества, как перекись водорода, гидроперекиси и пероксиды ненасыщенных жирных кислот, семихиноидные и хиноидные формы полифенолов, токсические адукаты перекиси водорода с гистидином и другие первично образующиеся РТ ([13], с. 122–124).
Действие радиотоксинов относится к вторичным изменениям, возникающим в результате облучения клеток. Многочисленные эксперименты показали способность первичных радиотоксинов задерживать деление клеток, вызывать их гибель, а также, что важно для обсуждения результатов нашего эксперимента, – имитировать лучевое поражение у животных ([12], с. 88). Радиотоксины, образующиеся в облученных растениях, легко экстрагируются, при введении подобных выделенных экстрактов необлученным животным происходит падение веса тела с последующим угнетением роста, резкое уменьшение количества лейкоцитов в первые часы после инъекции, а также изменение веса некоторых внутренних органов ([12], с. 88).
Далее рассмотрим дистанционные эффекты радиации с точки зрения теории радиотоксинов. Как отмечали А.М. Кузин и Л.М. Крюкова, облучение листа растения Vicia faba при полной экранировке всего остального растения приводит к угнетению митотического деления в точках роста, причем удаление облученного листа в ближайшие 4 ч после облучения приводило к снятию отмеченного эффекта ([12], с. 19). Установлено, что наряду с угнетением митотического деления происходит нарушение хромосомного аппарата ([12], с. 19). Таким образом, на вопрос о причастности первичных радиотоксинов к абскопальным дистанционным эффектам радиации, упомянутым во вступлении, дается положительный ответ.
В 1997–1998 гг. в работах Б.П. Суринова было показано, что совместное пребывание в клетке облученных и необлученных мышей и крыс приводит к таким проявлениям “эффекта свидетеля” у необлученных животных, как снижение уровня лейкоцитов и ядерных нейтрофилов периферической крови, а также снижение иммунной реактивности. Аналогичные эффекты вызывала подстилка, перенесенная под клетки с необлученными животными из-под клеток с облученными грызунами. Была высказана гипотеза о том, что сигнал лучевого “эффекта свидетеля” (“байстендерный” сигнал) между животными передается за счет летучих соединений, зависящих от пола, – феромонов [20–25].
Авторы трактуют результаты проведенного эксперимента с позиций теории радиотоксинов. Липоперекиси, образующиеся в результате облучения, являются высокотоксичными летучими соединениями. Продукты липоперекисного окисления, появление летучих метаболитов могут стать причиной возникновения молекулярного “байстендерного” сигнала [26, 27].
Необходимо также сказать о серии новейших экспериментов, проводимых группой канадских исследователей университета г. Макмастера во главе с К. Мазерсилл [28–30]. В качестве объяснения механизмов “эффекта свидетеля” тестируется гипотеза о роли биофотонов ультрафиолетового спектра в передаче “байстендерного” сигнала между организмами, и некоторые положения данной теории уже находят свое подтверждение. Предполагается, что использование указанных принципов квантовой биофизики выведет обсуждение механизма феномена на принципиально новый уровень [28–30].
Как было показано в предшествующих экспериментах Е.А. Нейфаха [31–33], процесс перекисного окисления в организме приводит к хемилюминесценции, появлению ультраслабого потока биофотонов сине-зеленого спектра, что вызвано взаимодействием радикалов. Не исключается роль данных фотонов, наряду с действием метаболитов липоперекисей, в процессе формирования и передачи “байстендерного” сигнала между клетками и организмами при облучении.
ЗАКЛЮЧЕНИЕ
Наши первые результаты в рамках запланированных исследований, являющихся важной задачей современной радиобиологии, продемонстрировали реальность индукции “эффекта свидетеля” в необлученном организме в результате его контакта с организмом, подвергшимся радиационному воздействию. Показано, что совместное содержание облученных (3 Гр) и необлученных мышей в течение 3 мес. вызвало у необлученных животных изменения селезенки, кишечника, кожи, подобные радиационно-индуцированным. Патологии, выявленные у облученных мышей, фиксировались у необлученных животных-“свидетелей” в менее выраженной форме.
Феномен “немишенных эффектов” и его механизмы по-прежнему недостаточно изучены. Ежегодно публикуются новые данные о проявлениях “немишенных эффектов”, появляются также принципиально новые подходы к изучению этого явления в связи с прогрессом лабораторной техники и методик. Сделанные в ходе исследования выводы являются предварительными, в дальнейшем предполагается провести тестирование различных гипотез, относящихся к механизмам передачи “байстендерного” сигнала на уровне организма. Актуальность задачи исследования лучевого дистанционного эффекта свидетеля связана с широкой применимостью результатов. “Дистанционно индуцированные эффекты свидетеля”, являющиеся темой настоящего исследования, создают угрозу передачи радиационных повреждений в масштабах популяции, включая человеческую, что требует особого изучения и внимания. Результаты оценки “немишенных эффектов” весьма востребованы в различных областях: в теоретической радиобиологии как одно из основополагающих знаний о воздействии радиации на клетки и организмы, в практической медицине для радиотерапии, а также в радиоэкологии при оценке долгосрочных последствий техногенных катастроф и изменений природной среды. В XXI веке в связи с развитием атомной энергетики государство и общество столкнулись с такими угрозами, как радиационные риски для населения и окружающей среды. Знание о проявлениях “немишенных эффектов” – важнейших звеньях в цепи последствий радиационного воздействия на биологические объекты – поможет выработке подходов к защите населения от негативных факторов, связанных с повышенным радиационным фоном.
Список литературы
Mothersill C., Rusin A., Seymour C. Low doses and non-targeted effects in environmental radiation protection; where are we now and where should we go? // Environ. Res. 2017. V. 159. P. 484–490.
Morgan W.F. Non-targeted and delayed effects of exposure to ionizing radiation: I. Radiation-induced genomic instability and bystander effects in vitro // Radiat. Res. 2003. V. 159. № 5. P. 567–580.
Литтл Д.Б. Немишенные эффекты ионизирующих излучений: выводы применительно к низкодозовым воздействиям // Радиац. биология. Радиоэкология. 2007. Т. 47. № 3. С. 262–272. [Littl D.B. Nemishennye effecty ioniziruyushih izlucheniy: vyvody primenitel’no k nizkodozovym vozdeystviyam // Radiats. Biol. Radioecol. 2007. V. 47. № 3. P. 262–272. (In Russian)]
Mothersill C., Smith R.W., Saroya R. et al. Irradiation of rainbow trout at early life stages results in legacy effects in adults // Int. J. Radiat. Biol. 2010. V. 86. № 10. P. 817–828.
Mothersill C., Smith R., Wang J. et al. Biological Entanglement–Like Effect after Communication of Fish Prior to X-Ray Exposure // Dose-Response: Int. J. January. 2018. P. 1–17.
Mothersill C., Rusin A., Fernandez-Palomo C., Seymour C. History of bystander effects research 1905 – present; what is in a name // Int. J. Radiat. Biol. 2018. V. 94. № 8. P. 696–707.
Smith R.W., Moccia R.D. The Radiation Induced Bystander Effect: Is there Relevance for Aquaculture? // Ann. Aquatic. Res. 2016. V. 3. № 3. P. 1026.
Reis P., Lourenço J., Carvalho F.P. et al. RIBE at an inter-organismic level: A study on genotoxic effects in Daphnia magna exposed to waterborne uranium and a uranium mine effluent // Aquat. Toxicol. 2018. V. 198. P. 206–214.
Ariyoshi K., Miura T., Kasai K. et al. Radiation-induced bystander effect in large Japanese field mouse (Apodemus speciosus) embryonic cells // Radiat. Environ. Biophys. 2018. V. 57. Issue 3. P. 223–231.
Lee M.H., Lee K.M., Kim E.H. Neighbor effect: penumbra-dose exposed neighbor cells contribute to the enhanced survival of high-dose targeted cells // Int. J. Radiat. Biol. 2017. V. 93. № 11. P. 1227–1238.
Galván I., Bonisoli-Alquati A., Jenkinson S. et al. Chronic exposure to low-dose radiation at Chernobyl favours adaptation to oxidative stress in birds // Br. Ecol. Soc. Function. Ecol. 2014. V. 28. P. 1387–1403.
Радиотоксины, их природа и роль в биологическом действии радиации высокой энергии / Ред. А.М. Кузин. М.: Атомиздат, 1966 г. [Radiotoxins, their nature and role in the biological effects of high energy radiation / Ed. A.M. Kuzin. M.: Atomizdat, 1966. (In Russian)]
Кузин А.М., Копылов В.А. Радиотоксины. М.: Наука, 1983. [Kuzin A., Kopylov V. Radiotoxins. M.: Nauka, 1983. (In Russian)]
Тимофеев-Ресовский Н.В., Савич А.В., Шальнов М.И. Введение в молекулярную радиобиологию. М.: Медицина, 1981. [Timofeev-Ressovsky N., Savich A., Shalnov M. Introduction to molecular radiobiology. M.: Medicina, 1981. (In Russian)]
Пелевина И.И., Петушкова В.В., Бирюков В.А. и др. Роль “немишенных эффектов” в реакции клеток человека на радиационное воздействие // Радиац. биология. Радиоэкология. 2019. Т. 59. № 3. С. 261–273. [Pelevina I.I., Petushkova V.V., Biryukov V.А. et al. The “Radiation-Induced Non-Targeted Effects” and their Role in Human Cell Response to Low Radiation Forcing // Radiation biology. Radioecology. 2019. V. 59. № 3. P. 261–273. (In Russian)]
Пелевина И.И., Серебряный А.М., Петушкова В.В. и др. Аберрации хромосом лимфоцитов при “немишенных эффектах” как маркер радиационно-индуцированных реакций организма // Сб. тр. конф. “Современные вопросы радиационной генетики”, Дубна: ОИЯИ, 2019. [Pelevina I.I., Serebryanyi A.M., Petushkova V.V. et al. Aberrations of Human Blood Lymphocytes as a Marker of Radiation-Induced Reactions // “Contemporary issues of radiation genetics”, Selected works of the Conference, Dubna, 2019. (In Russian)]
Пелевина И.И., Серебряный А.М., Петушкова В.В. и др. Аберрации хромосом лимфоцитов при “немишенных эффектах” как маркер радиационно-индуцированных реакций организма // Сб. тр. конф. ИХФ РАН / Ред. О.Т. Касаикина, И.С. Ионова. М.: РУДН, 2019. [Pelevina I.I., Serebryanyi A.M., Petushkova V.V. et al. Aberrations of Human Blood Lymphocytes as a Marker of Radiation-Induced Reactions // N.N. Semenov Federal Research Center for Chemical Physics, Russian Academy of Sciences, Selected works of the annual Conference. M.: RUDN University, 2019. (In Russian)]
Суринов Б.П., Духова Н.Н., Исаева В.Г. Свойства летучих соединений, выделяемых облученными ионизирующей радиацией животных и дистанционный “эффект свидетеля” (bystander effect) // Радиация и риск (Бюлл. Национального радиационно-эпидемиологического регистра). 2015. Т. 24. № 3. С. 105–114. [Surinov B.P., Dukhova N.N., Isaeva V.G. Properties of volatile compounds emitted by irradiated animals and radiation-induced inter-organism bystander effects // Radiation and Risk (Bulletin of the National Radiation and Epidemiological Registry). 2015. V. 24. № 3. P. 105–114. (In Russian)]
Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010.
Суринов Б.П., Исаева В.Г., Духова Н.Н. Коммуникативное умножение вторичных нарушений показателей крови и иммунитета в группах интактных мышей, опосредованное летучими выделениями облученных особей // Радиац. биология. Радиоэкология. 2004. Т. 44 № 4 С. 387–391. [Surinov B.P., Isaeva V.G., Dukhova N.N. Communicative multiplication of secondary disorders in blood formula and immunity in groups of intact mice caused by volatile compounds excreted by irradiated animals // Radiats. Biol. Radioecol. 2004. V.44. № 4. P. 387–391. (In Russian)]
Surinov B.P., Isaeva V.G., Dukhova N.N. Postirradiation volatile secretions of mice: syngeneic and allogeneic immune and behavioral effects // Bull. Exp. Biol. Med. 2004. V. 138. P. 384–386.
Суринов Б.П., Исаева В.Г., Токарев О.И. Аллопатическая активность летучих соединений облученных животных // Радиац. биология. Радиоэкология. 2001. Т. 41. С. 645–649. [Surinov B.P., Isaeva V.G., Tokarev O.I. Allelopathic activity of volatile secretions in irradiated animals // Radiats. Biol. Radioecol. 2001. V. 41. P. 645–649. (In Russian)]
Суринов Б.П., Карпова Н.А., Исаева В.Г. Пострадиационная коммуникативная индукция нарушений крови и иммунитета // Патол. физиология и эксперим. терапия. 1998. № 3. С. 7. [Surinov B.P., Karpova N.A., Isaeva V.G. Postradiacionnaya communicativnaya indukcia narusheniy krovi i immuniteta // Patol. Fiziol. Eksperim. Ter. 1998. № 3. P. 7. (In Russian)]
Суринов Б.П., Карпова Н.А., Исаева В.Г., Кулиш Ю.С. Пост стрессорные состояния и коммуникативные нарушения иммунитета и крови // Патологическая физиология и экспериментальная терапия. 2000. № 4. С. 9–11. [Surinov B.P., Karpova N.A., Isaeva V.G., Kulish I.S. 2000. Post-stress state and communicative alterations of immunity and blood // Patol. Fiziol. Eksperim. Ter. 2000. № 4. P. 9–11. (In Russian)]
Суринов Б.П., Исаева В.Г., Карпова Н.А. Контактная передача пострадиационного иммунодефицитного состояния // Иммунология. 1997. № 6. С. 18. [Surinov V.P., Isaeva V.G., Karpova N.A. Natural excretions of mice in the postradiation period and contact induction of immunodeficiency // Immunologiya. 1997. № 6. P. 18. (In Russian)]
Нейфах Е.А. Эмбриологический метод исследования перекисного окисления липидов // Симп. Свободно-радикальное перекисное окисление липидов в норме и при патологиях. М., 1976. С. 120. [Neyfakh E.A. The embryological method of investigation for lipoperoxidation // Symp. Free-radical lipid peroxidation in norm and pathology. M., 1976. P. 120. (In Russian)]
Нейфах Е.А. Высокочувствительный метод исследования токсичности перекисного окисления // Вопр. мед. химии. 1977. Т. 23. С. 131. [Neyfakh E.A. The high-sensitive method for lipoperoxide toxicity investigation // Voprosy Med. Chem. 1977. V. 23. P. 131. (In Russian)]
Mothersill C., Le M., Rusin A., Seymour C. Biophotons in radiobiology: inhibitors, communicators and reactors // Radiat. Prot. Dosim. 2019. V. 183. P. 136–141.
Le M., McNeill F.E., Seymour C.B. et al. Modulation of oxidative phosphorylation (OXPHOS) by radiation-induced biophotons // Environ. Res. 2018. V. 163 P. 80–87.
Le M., Fernandez-Palomo C., McNeill F.E. et al. Exosomes are released by bystander cells exposed to radiation-induced biophoton signals: Reconciling the mechanisms mediating the bystander effect // PLoSOne. 2017. V. 12. № 3. e 0173685.
Нейфах Е.А., Эммануэль Н.М. Возможности хемилюминесцентного метода исследования окисления незаменимых жирных кислот // Симп. Свободно-радикальные процессы в биосистемах. М.: Наука, 1964. P. 36. [Neyfakh E.A., Emmanuel N.M. Chemiluminescence method’s possibilities for investigation of essential fatty acid oxidation. Symp. Free radical processes in biosystems. M.: Nauka, 1964. P. 36. (In Russian)]
Нейфах Е.А. Исследование механизмов сверхслабой хемилюминесценции // Симп. Сверхслабая люминесценция в биологии. М.: МГУ, 1969. С. 44. [Neyfakh E.A. Investigation of mechanisms for ultra-weak chemiluminescence // Symp. Ultra-weak luminescence in biology. M.: MSU, 1969. P. 44. (In Russian)]
Нейфах Е.А. Свободно-радикальный механизм сверхслабой хемилюминесценции, сопряженной с перекисным окислением ненасыщенных жирных кислот // Биофизика. 1971. Т. 16. № 3. С. 560. [Neyfakh E.A. Free-radical mechanism of ultra-weak chemiluminescence coupled with UFA peroxidation // Biofizika. 1971. V. 16. № 3. P. 560. (In Russian)]
Дополнительные материалы отсутствуют.
Инструменты
Радиационная биология. Радиоэкология