

Растительные ресурсы, 2020, T. 56, № 2, стр. 99-111
Жизненное состояние, скорость роста и надземная фитомасса Pinus sylvestris (Pinaceae) в средневозрастных северотаежных лесах
П. Н. Катютин 1, *, В. В. Горшков 1
1 Ботанический институт им. В.Л. Комарова РАН
г. Санкт-Петербург, Россия
* E-mail: PauRussia@binran.ru
Поступила в редакцию 26.01.2020
После доработки 28.02.2020
Принята к публикации 18.03.2020
Аннотация
Целью работы являлась оценка различий биологического и ресурсного потенциала деревьев сосны обыкновенной разных категорий жизненного состояния и верификация коэффициентов, используемых для расчета индексов жизненного состояния древостоев. Исследование выполнено в 80-летних северотаежных зеленомошно-лишайниковых сосновых лесах на территории Кольского полуострова. Проанализирована структура надземной фитомассы и показатели роста деревьев сосны обыкновенной (Pinus sylvestris L.), отсящихся к четырем виталитетным категориям (здоровые, умеренно ослабленные, сильно ослабленные и усыхающие). Установлены достоверные (1.5–4.5-кратные) различия особей разных категорий по абсолютной и относительной (на единицу высоты и диаметра) массе охвоенных ветвей и массе хвои. Охарактеризованы различия в динамике радиального прироста деревьев разного уровня жизненного состояния. Дана оценка доли деревьев разных виталитетных категорий в общем запасе стволовой древесины, ветвей и хвои средневозрастных древостоев. Предложены верифицированные по массе хвои коэффициенты уравнения для расчета индекса жизненного состояния древостоев.
Классификация деревьев по социальному положению и роли в древесном ярусе является одной из наиболее давних, но не утративших своей актуальности проблем лесоведения. Активная разработка принципов диагностики жизненного состояния деревьев и древостоев лесообразующих видов получила дополнительный импульс во второй половине прошлого–начале нынешнего века в связи с оценкой влияния промышленного загрязнения на состояние лесов [1–11] и сохраняет свое значение до настоящего времени [12–17].
В настоящее время согласно действующему законодательству на территории Российской Федерации принята шкала, которая включает 11 категорий состояния деревьев: I – здоровые (без признаков ослабления); II – ослабленные; III – сильно ослабленные; IV – усыхающие; V – свежий сухостой; VI – старый сухостой; VII – свежий ветровал; VIII – свежий бурелом; IX – старый ветровал; X – старый бурелом; XI – аварийные деревья [18]. В практике мониторинга состояния лесов, подвергающихся антропогенному воздействию, выделяют, как правило, от 3 до 6 виталитетных категорий деревьев [1, 5, 6, 11, 19–25]. Эти классификации в своей основе применимы и к оценке состояния древостоев фоновых территорий [26–31], где виталитетная дифференциация обусловлена процессами конкуренции. Все предложенные в отечественной лесной науке классификации жизненного состояния деревьев в значительной степени опирались на принципы оценочной шкалы, разработанной и обоснованной в начале 70-х годов прошлого века [18, 32, 33]. Она отличалась, с одной стороны, относительной простотой и незначительными трудозатратами, с другой – достаточно высокой скоростью получения результата и удовлетворительной информативностью.
Диагностика состояния деревьев осуществляется путем глазомерной качественной или полуколичественной оценки одного или нескольких признаков. Основными критериями выделения виталитетных категорий особей выступают характеристики крон и ассимиляционного аппарата. Оцениваются, прежде всего, такие параметры, как степень дефолиации и дехромации, относительная плотность (ажурность) кроны по отношению к эталонным значениям. Определение эталона является особой проблемой, подходы к решению которой неодинаковы у разных исследователей.
На основе соотношения особей разных категорий состояния были разработаны интегральные индексы состояния древостоев [21, 34] или лесных сообществ в целом [10]. При оценке состояния древостоев каждой из виталитетной категорий присваивается условный коэффициент, который должен с той или иной степенью приближения отражать ее жизненный и продукционный потенциал, определяемый, прежде всего, объемом способных к нормальному функционированию ассимиляционных органов. Предложенные в настоящее время коэффициенты массы хвои особей разных категорий состояния определены преимущественно на основе экспертных оценок или по принципу равномерного уменьшения от высших категорий к низшим.
Целью настоящей работы является оценка объективности выделения виталитетных категорий у древесных растений и верификация коэффициентов, используемых для расчета индексов жизненного состояния древостоев на основе определения надземной фитомассы и показателей роста особей сосны обыкновенной (Pinus sylvestris L.), имеющих разный уровень жизненного состояния.
МАТЕРИАЛЫ И МЕТОДЫ
Исследование выполнено в подзоне северной тайги на территории Кольского полуострова, в районе среднего течения реки Лива (67°30′–68°10′ с.ш., 33°57′–34°21′ в.д.) в 80-летних зеленомошно-лишайниковых сосновых лесах, сформировавшихся после катастрофических пожаров конца 20-х годов прошлого века.
Изученные сообщества располагаются на первой надпойменной террасе р. Ливы, сформированной песчаными водно-ледниковыми отложениями. Почвообразующие породы отличаются высокой водопроницаемостью и бедностью элементами минерального питания. На них формируются подзолы иллювиально-железистые ненасыщенные, мелкоподзолистые, иллювиально-малогумусовые, песчаные, средне скелетные [35]. Почвы отличаются коротким профилем (до 60 см) и накоплением оксалаторастворимых соединений Al и Fe в иллювиальном горизонте при низком содержании в нем (не более 1.5%) фульватного гумуса [36, 37].
В исследованных сообществах основу древесного яруса (70–100% по запасу) составляют особи Pinus sylvestris L. послепожарного происхождения. Таксационные характеристики деревьев разных категорий состояния представлены в табл. 1.
Таблица 1.
Параметры деревьев разных категорий состояния в изученных древостоях сосны обыкновенной Table 1. Parameters of the trees of different vitality classes in the studied Scots pine stands
| Категория состояния Vitality class |
N | A0, лет A0, years |
A1.3, лет A1.3, years |
D0, см D0, cm |
D1.3, см D1.3, cm |
H, м H, m |
|---|---|---|---|---|---|---|
| ПП 1 Sample plot 1 |
||||||
| I | 33 | 66–80 | 44–70 | 11.3–29.4 | 9.6–22.5 | 8–15.4 |
| II | 30 | 71–80 | 54–69 | 8.1–24.6 | 5.8–20 | 5.1–15.4 |
| III | 27 | 65–77 | 43–68 | 6.2–26.1 | 4–21.4 | 4.5–14.5 |
| IV | 11 | 66–73 | 50–63 | 6.2–12.1 | 4.1–10 | 4.7–11.2 |
| V | 5 | 73 | – | 8.7–13.5 | 7.5–9 | 5.8–8 |
| ПП 2 Sample plot 2 |
||||||
| I | 10 | 65–80 | 60 | 21.5–33.3 | 11.8–26.1 | 10–13 |
| II | 37 | 68–80 | 56–70 | 13.6–39.4 | 11.4–29.4 | 8.7–16 |
| III | 44 | 62–71 | 49–60 | 11–26.7 | 4.2–22.9 | 8.5–15.5 |
| IV | 20 | 61–68 | 47–55 | 6–16.3 | 4–13.6 | 5.2–11 |
| V | 10 | – | – | 8.1–15.9 | 5.2–16.5 | 4.2–10.3 |
| В среднем Average |
||||||
| I | 43 | 73 ± 4 | 61 ± 9 | 21 ± 5 | 18 ± 4 | 12 ± 2 |
| II | 67 | 75 ± 7 | 63 ± 6 | 20 ± 5 | 16 ± 4 | 12 ± 2 |
| III | 71 | 71 ± 7 | 59 ± 7 | 16 ± 5 | 12 ± 4 | 11 ± 2 |
| IV | 31 | 67 ± 3 | 58 ± 5 | 10 ± 3 | 8 ± 3 | 8 ± 2 |
| V | 15 | – | – | 11 ± 2 | 8 ± 3 | 7 ± 2 |
Примечание. I – неугнетенные (здоровые) деревья; II – умеренно угнетенные (умеренно ослабленные) деревья; III – сильно угнетенные (сильно ослабленные) деревья; IV – усыхающие деревья; V – сухие деревья. N – количество деревьев; A0, A1.3 – возраст, установленный у основания ствола и на высоте 1.3 м; D0, D1.3 – диаметр ствола у основания и на высоте 1.3 м; H – высота. Note. I – unoppressed (healthy) trees; II – moderately oppressed (moderately weakened) trees; III – strongly oppressed (strongly weakened) trees; IV – dying trees; V – dry trees. N – number of trees; A0, A1.3 – tree age determined at the tree base and at breast height (1.3 m); D0, D1.3 – trunk diameter at the base and at breast height (1.3 m); H – height.
Категории жизненного состояния деревьев сосны обыкновенной определялись с учетом имеющихся рекомендаций [5, 8, 18, 21, 27, 33, 38]. Основным критерием служил показатель плотности кроны, который оценивался по отношению к плотности кроны эталонной особи (сформировавшейся в условиях минимального конкурентного воздействия) соответствующего возраста, принимаемой за 1. Выделялось 5 категорий состояния: I – неугнетенные (здоровые) особи – относительная плотность кроны (CD) > 75–100% (в среднем 87.5%), II – умеренно угнетенные (умеренно ослабленные) особи – CD > 50–75% (в среднем 62.5%), III – сильно угнетенные (сильно ослабленные) особи – CD > 25–50% (в среднем 37.5%), IV – усыхающие особи – CD > 0–25% (в среднем 12.5%), V – сухие особи. Если принять относительную плотность кроны здоровых особей в составе древостоев (87.5%) за 1.0, то коэффициенты плотности кроны умеренно ослабленных, сильно ослабленных и усыхающих особей составят соответственно 0.71, 0.43 и 0.14. Эти величины и были предложены В.Т. Ярмишко с соавторами [39] для расчета величины индекса жизненного состояния древостоев.
В травяно-кустарничковом ярусе преобладают Vaccinium vitis-idea L., Vaccinium myrtillus L., Empetrum hermaphroditum Hagerup. В мохово-лишайниковом ярусе доминируют лишайники рода Cladonia (Cladonia stellaris (Opiz.) Brodo, Cladonia rangiferina (L.) Nyl., Cladonia mitis (Sandst.) Hustich). Покрытие мхов (Pleurozium schreberi (Brid) Mitt. и Dicranum polysetum Sw.) составляет около 15%.
На первом этапе исследования были отобраны 70–80-летние модельные деревья сосны (19 деревьев в лесном сообществе и 2 модельных дерева на вырубке-гари) с диаметром ствола на высоте 1.3 м 14–18 см. Количество модельных деревьев, относящихся к категориям здоровых, умеренно ослабленных, сильно ослабленных и усыхающих составило соответственно 4, 5, 6 и 4. Группы особей, относящихся к разным категориям состояния, по величине среднего диаметра достоверно не различались (K–W = 2.64; P = 0.45). Среднее значение диаметра в пределах всей выборки модельных особей составило 15.9 ± 0.3 см.
После спиливания у всех моделей измеряли диаметры ствола через каждые два метра, начиная от основания, общую высоту и протяженность кроны. Затем методом полного взвешивания отдельных фракций на электронных весах с точностью 0.01–0.001 г определяли сырую массу ствола в коре, массу живых неохвоенных ветвей, массу ветвей, несущих хвою, массу хвои и массу сухих ветвей [40]. Для дальнейшего определения воздушно-сухой массы отбирались образцы каждой фракции. Всего было отобрано 300 образцов.
Для изучения динамики радиального прироста у моделей отбирали спилы древесины у основания ствола, на высоте 1.3 м. После подготовки (шлифовки) спилов измерение радиального прироста проводилось по общепринятым методикам [41, 42] с использованием микроскопа МБС-10 и полуавтоматической установки LINTAB-6 с точностью 0.01 мм.
Коэффициент синхронности Kс прироста деревьев разного жизненного состояния определялся на основе уравнения вида [41]:
где п+ – количество совпавших по направлению годичных отрезков кривой; п – длительность временного интервала, лет.При анализе скорости роста использовались средние значения, полученные путем сглаживания величин радиального прироста за 5 лет.
Обработка данных проводилась с использованием методов описательной статистики, корреляционного и регрессионного анализа.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Надземная фитомасса модельных деревьев
Особи, имеющие одинаковый диаметр и относящиеся к разным категориям состояния, по величине общей надземной фитомассы, массы ствола и скелетной части кроны достоверно не различаются (табл. 2). Общая воздушно-сухая надземная фитомасса модельных деревьев сосны обыкновенной составляет в среднем 76 кг; у отдельных особей она имеет двукратные различия, варьируя от 52 до 106 кг. Масса ствола и неохвоенных ветвей варьирует соответственно от 33 до 79 кг и от 3.4 до 15.5 кг, составляя в среднем 54 ± 3 и 9.5 ± 0.7 кг. В процентном выражении на массу ствола приходится в среднем 73%, на массу неохвоенных ветвей – 13% от общей надземной фитомассы (табл. 3).
Таблица 2.
Воздушно-сухая фитомасса (кг) модельных деревьев сосны обыкновенной разных категорий состояния Table 2. Air-dry biomass (kg) of model Scots pine trees of different vitality classes
| Категория состояния Vitality class |
N | Ствол в коре Trunk with bark |
Неохвоенные живые ветви Live branches |
Сухие ветви Dry branches |
Охвоенные ветви Branches with needles |
Всего Total |
||
|---|---|---|---|---|---|---|---|---|
| всего total |
хвоя needles |
ветви branches |
||||||
| В составе древостоя In the forest stand |
||||||||
| I | 4 | $\frac{{61 \pm 8{\text{a*}}}}{{37{\kern 1pt} --{\kern 1pt} 74}}$ | $\frac{{12 \pm 1{\text{a}}}}{{9.5{\kern 1pt} --{\kern 1pt} 14.5}}$ | $\frac{{3.6 \pm 0.8{\text{a}}}}{{2.8{\kern 1pt} --{\kern 1pt} 6.1}}$ | $\frac{{9.3 \pm 0.6{\text{a}}}}{{8.1{\kern 1pt} --{\kern 1pt} 10.4}}$ | $\frac{{6.6 \pm 0.4{\text{a}}}}{{5.7{\kern 1pt} --{\kern 1pt} 7.6}}$ | $\frac{{2.7 \pm 0.3{\text{a}}}}{{2.0{\kern 1pt} --{\kern 1pt} 3.4}}$ | $\frac{{86 \pm 10{\text{a}}}}{{57{\kern 1pt} --{\kern 1pt} 104}}$ |
| II | 5 | $\frac{{58 \pm 9{\text{a}}}}{{33{\kern 1pt} --{\kern 1pt} 79}}$ | $\frac{{9.6 \pm 0.8{\text{a}}}}{{8.2{\kern 1pt} --{\kern 1pt} 12.8}}$ | $\frac{{4.7 \pm 0.9{\text{ab}}}}{{3.1{\kern 1pt} --{\kern 1pt} 8.3}}$ | $\frac{{6.2 \pm 0.1{\text{b}}}}{{6.0{\kern 1pt} --{\kern 1pt} 6.8}}$ | $\frac{{4.4 \pm 0.2{\text{b}}}}{{3.9{\kern 1pt} --{\kern 1pt} 4.9}}$ | $\frac{{1.8 \pm 0.1{\text{b}}}}{{1.5{\kern 1pt} --{\kern 1pt} 2.2}}$ | $\frac{{78 \pm 10{\text{a}}}}{{52{\kern 1pt} --{\kern 1pt} 106}}$ |
| III | 6 | $\frac{{54 \pm 3{\text{a}}}}{{42{\kern 1pt} --{\kern 1pt} 64}}$ | $\frac{{8.4 \pm 0.6{\text{a}}}}{{6.9{\kern 1pt} --{\kern 1pt} 10.9}}$ | $\frac{{5.1 \pm 0.8{\text{ab}}}}{{3.2{\kern 1pt} --{\kern 1pt} 8.7}}$ | $\frac{{4.8 \pm 0.3{\text{c}}}}{{3.5{\kern 1pt} --{\kern 1pt} 5.4}}$ | $\frac{{3.5 \pm 0.2{\text{c}}}}{{2.7{\kern 1pt} --{\kern 1pt} 4.3}}$ | $\frac{{1.4 \pm 0.1{\text{b}}}}{{0.8{\kern 1pt} --{\kern 1pt} 1.8}}$ | $\frac{{72 \pm 3{\text{a}}}}{{58{\kern 1pt} --{\kern 1pt} 83}}$ |
| IV | 4 | $\frac{{50 \pm 5{\text{a}}}}{{41{\kern 1pt} --{\kern 1pt} 62}}$ | $\frac{{9 \pm 2{\text{a}}}}{{3.4{\kern 1pt} --{\kern 1pt} 15.5}}$ | $\frac{{8 \pm 2{\text{b}}}}{{3.4{\kern 1pt} --{\kern 1pt} 13.4}}$ | $\frac{{2.1 \pm 0.2{\text{d}}}}{{1.5{\kern 1pt} --{\kern 1pt} 2.5}}$ | $\frac{{1.5 \pm 0.2{\text{d}}}}{{1.0{\kern 1pt} --{\kern 1pt} 1.8}}$ | $\frac{{0.6 \pm 0.1{\text{с}}}}{{0.5{\kern 1pt} --{\kern 1pt} 0.8}}$ | $\frac{{69 \pm 6{\text{a}}}}{{58{\kern 1pt} --{\kern 1pt} 83}}$ |
| На вырубке At felling |
||||||||
| I | 2 | $\frac{{41{\text{ }} \pm 16{\text{a}}}}{{25{\kern 1pt} --{\kern 1pt} 58}}$ | $\frac{{12 \pm 4{\text{a}}}}{{8{\kern 1pt} --{\kern 1pt} 16}}$ | $\frac{{0.4 \pm 0.2{\text{с}}}}{{0.2{\kern 1pt} --{\kern 1pt} 0.6}}$ | $\frac{{15 \pm 2{\text{e}}}}{{12{\kern 1pt} --{\kern 1pt} 17}}$ | $\frac{{10 \pm 2{\text{e}}}}{{8{\kern 1pt} --{\kern 1pt} 12}}$ | $\frac{{4.7 \pm 0.2{\text{e}}}}{{4.2{\kern 1pt} --{\kern 1pt} 5.2}}$ | $\frac{{68 \pm 24{\text{a}}}}{{45{\kern 1pt} --{\kern 1pt} 92}}$ |
Примечание. I – неугнетенные (здоровые) деревья; II – умеренно угнетенные (умеренно ослабленные) деревья; III – сильно угнетенные (сильно ослабленные) деревья; IV – усыхающие деревья; V – сухие деревья. Над чертой – среднее значение фитомассы с ошибкой, под чертой – диапазон значений. * Значения, отмеченные разными буквенными индексами, достоверно различаются при уровне значимости P < 0.05. Note. I – unoppressed (healthy) trees; II – moderately oppressed (moderately weakened) trees; III – strongly oppressed (strongly weakened) trees; IV – dying trees; V – dry trees. Above the line – the average value of phytomass and an error, below the line – the range of values. * Values marked with different letter indices differ at a significance level of P < 0.05.
Таблица 3.
Соотношение (%) фракций фитомассы модельных деревьев сосны обыкновенной разных категорий состояния Table 3. The ratio (%) of biomass fractions of model Scots pine trees of different vitality classes
| Категория состояния Vitality class |
N | Ствол в коре Trunk with bark |
Неохвоеные живые ветви Live branches |
Сухие ветви Dry branches |
Охвоенные ветви Branches with needles |
Всего Total |
||
|---|---|---|---|---|---|---|---|---|
| всего total |
хвоя needles |
ветви branches |
||||||
| В составе древостоя In the forest stand |
||||||||
| I | 4 | $\frac{{70 \pm 2{\text{a*}}}}{{64{\kern 1pt} --{\kern 1pt} 73}}$ | $\frac{{14 \pm 1{\text{a}}}}{{11{\kern 1pt} --{\kern 1pt} 17}}$ | $\frac{{4 \pm 1{\text{a}}}}{{3{\kern 1pt} --{\kern 1pt} 6}}$ | $\frac{{11 \pm 1{\text{a}}}}{{9{\kern 1pt} --{\kern 1pt} 14}}$ | $\frac{{8.0 \pm 0.8{\text{a}}}}{{6.5--10}}$ | $\frac{{3.2 \pm 0.4{\text{a}}}}{{2{\kern 1pt} --{\kern 1pt} 4}}$ | 100 |
| II | 5 | $\frac{{73 \pm 3{\text{a}}}}{{63{\kern 1pt} --{\kern 1pt} 79}}$ | $\frac{{13 \pm 1{\text{a}}}}{{10{\kern 1pt} --{\kern 1pt} 16}}$ | $\frac{{6 \pm 1{\text{a}}}}{{3{\kern 1pt} --{\kern 1pt} 9}}$ | $\frac{{8 \pm 1{\text{b}}}}{{6{\kern 1pt} --{\kern 1pt} 12}}$ | $\frac{{6.0 \pm 0.9{\text{b}}}}{{4{\kern 1pt} --{\kern 1pt} 9}}$ | $\frac{{2.5 \pm 0.3{\text{ab}}}}{{1{\kern 1pt} --{\kern 1pt} 3}}$ | 100 |
| III | 6 | $\frac{{74 \pm 1{\text{a}}}}{{71{\kern 1pt} --{\kern 1pt} 78}}$ | $\frac{{12 \pm 1{\text{a}}}}{{9{\kern 1pt} --{\kern 1pt} 13}}$ | $\frac{{7 \pm 1{\text{a}}}}{{4{\kern 1pt} --{\kern 1pt} 12}}$ | $\frac{{7 \pm 0.4{\text{b}}}}{{5{\kern 1pt} --{\kern 1pt} 8}}$ | $\frac{{4.8 \pm 0.2{\text{b}}}}{{4{\kern 1pt} --{\kern 1pt} 5.5}}$ | $\frac{{2.0 \pm 0.2{\text{b}}}}{{1{\kern 1pt} --{\kern 1pt} 3}}$ | 100 |
| IV | 4 | $\frac{{73 \pm 1{\text{a}}}}{{70{\kern 1pt} --{\kern 1pt} 76}}$ | $\frac{{13 \pm 3{\text{a}}}}{{5{\kern 1pt} --{\kern 1pt} 19}}$ | $\frac{{12 \pm 4{\text{b}}}}{{4{\kern 1pt} --{\kern 1pt} 21}}$ | $\frac{{3.1 \pm 0.4{\text{c}}}}{{2{\kern 1pt} --{\kern 1pt} 4}}$ | $\frac{{2.2 \pm 0.3{\text{c}}}}{{1.4{\kern 1pt} --{\kern 1pt} 2.8}}$ | $\frac{{1.0 \pm 0.1{\text{c}}}}{{0.7{\kern 1pt} --{\kern 1pt} 1.4}}$ | 100 |
| На вырубке At felling |
||||||||
| I | 2 | $\frac{{59 \pm 4{\text{b}}}}{{55{\kern 1pt} --{\kern 1pt} 63}}$ | $\frac{{17.5 \pm 0.4{\text{b}}}}{{17{\kern 1pt} --{\kern 1pt} 18}}$ | $\frac{{0.5 \pm 0.1{\text{a}}}}{{0.4{\kern 1pt} --{\kern 1pt} 0.6}}$ | $\frac{{23 \pm 4{\text{d}}}}{{19{\kern 1pt} --{\kern 1pt} 28}}$ | $\frac{{16 \pm 3{\text{d}}}}{{13{\kern 1pt} --{\kern 1pt} 18}}$ | $\frac{{8 \pm 2{\text{d}}}}{{6{\kern 1pt} --{\kern 1pt} 9}}$ | 100 |
Примечание. I – неугнетенные (здоровые) деревья; II – умеренно угнетенные (умеренно ослабленные) деревья; III – сильно угнетенные (сильно ослабленные) деревья; IV – усыхающие деревья; V – сухие деревья. Над чертой – среднее значение с ошибкой, под чертой – диапазон значений. * Значения, отмеченные разными буквенными индексами, достоверно различаются при уровне значимости P < 0.05. Note. I – unoppressed (healthy) trees; II – moderately oppressed (moderately weakened) trees; III – strongly oppressed (strongly weakened) trees; IV – dying trees; V – dry trees. Above the line – the average value and an error, below the line – the range of values. * Values marked with different letter indices differ at a significance level of P < 0.05.
Масса сухих ветвей, охвоенных ветвей и масса хвои имеют значимую связь с уровнем жизненного состояния особей сосны (соответственно R = 0.46, P < 0.05; R = – 0.89, P < 0.001 и R = – 0.95, P < 0.001). Достоверными, примерно двукратными различиями массы сухих ветвей характеризуются здоровые (3.6 ± 0.8 кг) и усыхающие (8.0 ± 2 кг) особи (табл. 2). У здоровых деревьев масса сухих ветвей варьирует от ~3 до 6 кг, у усыхающих – от 3 до 13 кг. Как показывают полученные данные, одинаковые по диаметру особи всех категорий состояния могут иметь одинаковую и, в ряде случаев, довольно низкую (~3 кг), массу сухих ветвей (табл. 2). Доля сухих ветвей в общей фитомассе у здоровых, умеренно и сильно ослабленных деревьев достоверно не различается и составляет в среднем ~5.5%, у усыхающих – она является в 2 раза более высокой (табл. 3).
Существенное различие деревьев разных категорий состояния выявлено по массе охвоенных ветвей и хвои. Воздушно-сухая масса охвоенных ветвей имеет значимые различия у здоровых, умеренно ослабленных и усыхающих особей. У здоровых деревьев сосны обыкновенной она максимальна и составляет в среднем ~3 кг, у умеренно и сильно ослабленных особей она достоверно не различается и является более низкой (от 1.5 до ~2 кг), минимальное значение характерно для усыхающих деревьев – в среднем ~0.5 кг (табл. 2). Доля охвоенных ветвей в общей надземной фитомассе снижается от ~3% у здоровых деревьев до 1% – у усыхающих (табл. 3).
Масса хвои достоверно различается у особей всех категорий состояния. В ряду от здоровых до усыхающих особей ее средняя величина изменяется от ~6.5 до 1.5 кг (табл. 2). В общей надземной фитомассе доля хвои уменьшается от 8% у здоровых особей сосны до ~2% – у усыхающих. Средние значения массы охвоенных ветвей и массы хвои у здоровых деревьев соответственно в 1.5, 2 и 4.5 раза превышают соответствующие значения у умеренно ослабленных, сильно ослабленных и усыхающих особей. В соответствии с полученными данными, эти параметры можно рассматривать как индикационные в отношении различий между особями разного уровня жизненного состояния.
Установленное процентное соотношение основных фракций фитомассы здоровых и умеренно ослабленных деревьев сосны в целом соответствует оценкам, полученным ранее в молодых и средневозрастных сосновых древостоях на Кольском полуострове и сопряженных районах, находящихся вне зоны промышленного загрязнения [8, 43–49]. Структура фитомассы сильно ослабленных и усыхающих особей в настоящем исследовании сопоставима со структурой, характерной для деревьев сосны, сформировавшихся в зоне с высоким уровнем промышленного загрязнения [8, 50]. В последнем случае регистрируется увеличение доли сухих ветвей (до 3-крат) и снижение доли хвои (1.5–2-кратное) в общей надземной фитомассе по сравнению со здоровыми деревьями в древостоях фоновых территорий.
Здоровые деревья сосны обыкновенной, произрастающие на вырубке-гари и имеющие близкие к остальным модельным особям показатели диаметра ствола, отличаются от здоровых деревьев в составе древостоя достоверно более высокими значениями массы охвоенных ветвей и хвои (табл. 2). Различия по массе ствола и общей надземной фитомассе (1.3–1.5-кратные) в данной выборке не удалось доказать статистически. Кроме того, здоровые деревья на вырубке по сравнению со здоровыми деревьями в древостое имеют достоверно более низкую долю в общей фитомассе стволовой древесины (59% против 70%) и более высокую долю охвоенных ветвей (8% против 3%) и хвои (16% против 8%).
Анализ относительных величин массы хвои позволил установить, что показатель массы хвои на единицу высоты значимо различается (K–W = 18.48, P < 0.001) у особей всех категорий состояния. У здоровых деревьев в составе древостоев на каждый метр высоты приходится 0.54 кг воздушно-сухой массы хвои; в условиях свободного развития на вырубке этот показатель в 2 раза выше. У умеренно ослабленных, сильно ослабленных и усыхающих особей масса хвои на единицу высоты ствола составляет соответственно 0.33, 0.26 и 0.13 кг, то есть 60, 48 и 24% от величины у здоровых особей в древостое.
При анализе относительной массы хвои, приходящейся на единицу диаметра на высоте 1.3 м, было выявлено значимое различие этих величин у особей всех категорий состояния за исключением умеренно и сильно ослабленных. На 1 см диаметра ствола на высоте 1.3 м у здоровых деревьев составе древостоя приходится соответственно 0.4 кг хвои, у умеренно ослабленных, сильно ослабленных и усыхающих особей соответственно 0.27, 0.23 и 0.09 кг, что составляет 67, 57 и 22% от указанной величины. Здоровые деревья на вырубке по величине массы хвои на единицу диаметра ствола превышают здоровые особи в составе древостоев в 1.5 раза.
Радиальный прирост
Анализ роста модельных деревьев сосны обыкновенной, имеющих одинаковую величину диаметра ствола на высоте 1.3 м (18 см), но разную категорию состояния (рис. 1), позволяет выделить два основных периода, различающихся по характеру динамики радиального прироста. На протяжении первого периода, который продолжался примерно 40 лет (с конца 30-х до конца 70-х годов прошлого века), средняя величина прироста особей не имела однозначной связи с уровнем их жизненного состояния, установленным в 80-летнем возрасте. Она варьировала от 0.8–0.9 до 1.2 мм/год и была наиболее высокой у умеренно ослабленной и отмирающей особей, а наиболее низкой – у сильно ослабленной и здоровой. Однако здоровое дерево в отличие от деревьев всех остальных категорий состояния на протяжении рассматриваемого периода имело достоверную тенденцию линейного увеличения прироста (F = 8.98, P < 0.05, R = 0.75). Его величина возросла от 0.5–0.7 мм/год до 1.7 мм/год (рис. 1).
Рис. 1.
Динамика радиального прироста основания ствола модельных деревьев Pinus sylvestris (диаметр ствола на высоте груди 18 см). По горизонтали – годы; по вертикали – радиальный прирост, мм в год. Fig. 1. Dynamics of Pinus sylvestris model trees radial growth at the trunk base (DBH 18 cm). X-axis – years; Y-axis – radial increment, mm per year.
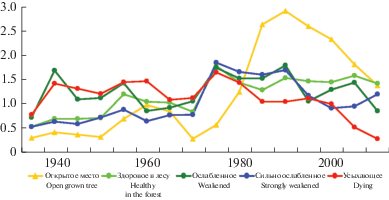
В течение второго периода динамики, продолжавшегося с конца 70-х годов прошлого века до конца исследованного периода (2010 г.), прирост здорового дерева оставался стабильным и в среднем составлял 1.47 ± 0.04 мм/год. Прирост особей остальных категорий состояния достоверно снизился (F = 7.81–44.46, P < 0.05, R = –0.75...–0.94: у умеренно и сильно ослабленных особей – в 1.5–2 раза, у усыхающей – в 5 раз. Ход кривых и сопоставление прироста особей разных категорий состояния на протяжении второго периода динамики, свидетельствует о том, что скорость роста и соответственно статус особей только двух крайних категорий состояния можно считать достаточно определенным. Что касается умеренно и сильно ослабленных особей, то выраженная вариабельность их прироста позволяет предполагать, что их статус может меняться с течением времени как сторону повышения, так и в сторону понижения.
Согласно величине коэффициента синхронности, динамика прироста здоровой и умеренно ослабленной особей сосны отличается более довольно высокой степенью согласованности с усредненной кривой (Kс = 84 и 77%, P < 0.001) и высокой взаимной сопряженностью (Kс = 70%, P < 0.001). Кривая прироста сильно ослабленного дерева имеет более низкую степень соответствия усредненной кривой (Kс = 67%, P < 0.01), а кривая усыхающего дерева с ней не согласуется (Kс = 57%, P > 0.05). Прирост сильно ослабленного и усыхающего деревьев не синхронизирован с приростом здорового и умеренно ослабленного и не имеет достоверной взаимной согласованности (Kс = = 44–60%, P > 0.05).
Модельное дерево сосны, которое до начала 80-х годов прошлого века произрастало под пологом и конкурентным воздействием пережившего пожар древостоя, на протяжении периода с конца 30-х до конца 70-х годов, также как большинство особей, развивавшихся на открытой гари, не имело направленной тенденции изменения прироста. Его средняя величина (0.51 ± 0.08 мм/год) была достоверно наиболее низкой среди других модельных деревьев. После рубки допожарного древостоя в начале 80-х годов произошло существенное (более, чем 2-кратное) увеличение прироста дерева, оказавшегося на открытом пространстве, за которым последовало его снижение до величины, зарегистрированной у здорового дерева в составе средневозрастного древостоя (рис. 1). В течение рассматриваемого периода модельное дерево, произрастающее на открытом месте, отличалось низким уровнем согласованности (Kс не более 65%) в динамике роста с деревьями, произрастающими в составе древостоя.
Представляет интерес сопоставление параметров прироста отдельных модельных деревьев сосны с параметрами прироста групп особей аналогичных категорий состояния, представленных в составе средневозрастных древостоев сосны (рис. 2).
Рис. 2.
Динамика радиального прироста основания ствола деревьев Pinus sylvestris разного жизненного состояния в средневозрастных зеленомошно-лишайниковых сосновых лесах. По горизонтали – годы; по вертикали – радиальный прирост мм/год. Fig. 2. The dynamics of Pinus sylvestris radial growth at the trunk base in trees of different vitality classes in middle-aged Scots pine – lichen – green-moss forest. X-axis – years; Y-axis – radial increment, mm per year.
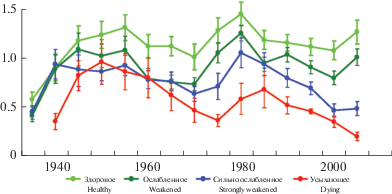
За весь исследованный период зарегистрировано два выраженных периода роста значений прироста (с конца 30-х до конца 50-х годов и с начала 70-х до начала 80-х годов прошлого века) и два периода спада (с конца 50-х годов до начала 70-х и с начала 80-х до начала 2000-х). Однонаправленное изменение (снижение) прироста за весь исследованный период (F = 7.01, P < 0.05, R = – 0.61) выявлено только у категории усыхающих особей.
Все виталитетные группы деревьев имеют достоверные различия по величине среднего прироста за весь исследованный период (K–W = 31.53, P < 0.001). Прирост умеренно ослабленных, сильно ослабленных и усыхающих деревьев составляет соответственно 80, 67 и 50% от прироста здоровых, составляющего 1.14 ± 0.05 мм/год. В тоже время следует отметить, что если лидирующая роль здоровых деревьев, достоверно проявляющаяся с начала 50-х годов, оставалась устойчивой на протяжении всего исследованного периода, то позиции особей остальных категорий состояния достаточно четко определились лишь с начала 70-х годов, т.е. на 20 лет позже. В последние 20 лет жизни особей (с начала 80-х до середины 2000-х годов) радиальный прирост здоровых и умеренно ослабленных деревьев оставался стабильным, прирост сильно ослабленных и усыхающих особей достоверно снижался (F = 93.58 и 30.51, P < 0.001 и < 0.01, R = = – 0.97 и – 0.94).
Кривая прироста здоровых деревьев характеризуется очень высокой степенью синхронности с усредненной кривой (Kс = 95%, P < 0.001); у умеренно и сильно ослабленных деревьев степень синхронности является более низкой (Kс = 83–84%, P < 0.001). Кроме того, прирост деревьев этих трех виталитетных категорий отличается довольно отчетливой взаимной согласованностью (Kс = 78–84%, P < 0.001). Прирост усыхающих особей не синхронизирован с приростом особей других категорий состояния (Kс = 47–57%, P > 0.05). Этот результат с целом согласуется с оценками степени синхронности прироста модельных деревьев разных категорий состояния и подтверждает высказанное еще во второй половине прошлого века мнение [51], что прирост деревьев низкого уровня жизненного состояния, регулируется, прежде всего, конкурентным воздействием, а не климатическими факторами.
Фитомасса древостоев средневозрастных сосновых лесов
На основе полученных результатов, характеризующих структуру фитомассы модельных деревьев сосны и данных о соотношении числа особей разных категорий состояния в составе древостоев, дана оценка запаса основных фракций надземной фитомассы сосны в изученных средневозрастных сосновых лесах. Величина общего запаса фитомассы и соотношение отдельных фракций в его составе (табл. 4) находятся в соответствии с данными других исследователей, полученными в средневозрастных северотаежных лесах [43, 48, 52, 53].
Таблица 4.
Соотношение фракций фитомассы древостоя в средневозрастном сосняке лишайниково-зеленомошном Table 4. The ratio of biomass fractions of the stand in the middle-aged Scots pine–lichen–green-moss forest
| Категория состояния Vitality class |
Доля участия, % Proportion, % |
Ствол в коре Stem with bark |
Неохвоенные живые ветви Live branches |
Охвоенные ветви Branches with needles |
Всего Total |
||
|---|---|---|---|---|---|---|---|
| всего total |
хвоя needles |
ветви branches |
|||||
| I | 33 | $\frac{{9.92}}{{30}}$ | $\frac{{1.96}}{{34}}$ | $\frac{{1.28}}{{35}}$ | $\frac{{0.91}}{{36}}$ | $\frac{{0.37}}{{35}}$ | $\frac{{13.16}}{{31}}$ |
| II | 30 | $\frac{{11.22}}{{34}}$ | $\frac{{1.87}}{{32}}$ | $\frac{{1.34}}{{37}}$ | $\frac{{0.94}}{{37}}$ | $\frac{{0.40}}{{37}}$ | $\frac{{14.43}}{{34}}$ |
| III | 27 | $\frac{{9.73}}{{29}}$ | $\frac{{1.52}}{{26}}$ | $\frac{{0.89}}{{25}}$ | $\frac{{0.63}}{{24}}$ | $\frac{{0.26}}{{25}}$ | $\frac{{12.13}}{{28}}$ |
| IV | 10 | $\frac{{2.46}}{7}$ | $\frac{{0.44}}{8}$ | $\frac{{0.10}}{3}$ | $\frac{{0.07}}{3}$ | $\frac{{0.03}}{3}$ | $\frac{{3.00}}{7}$ |
| ВСЕГО: TOTAL: |
100 | $\frac{{33.33}}{{100}}$ | $\frac{{5.78}}{{100}}$ | $\frac{{3.62}}{{100}}$ | $\frac{{2.55}}{{100}}$ | $\frac{{1.07}}{{100}}$ | $\frac{{42.72}}{{100}}$ |
Примечание. I – неугнетенные (здоровые) деревья; II – умеренно угнетенные (умеренно ослабленные) деревья; III – сильно угнетенные (сильно ослабленные) деревья; IV – усыхающие деревья; V – сухие деревья. Над чертой – т/га, под чертой – %. Note. I – unoppressed (healthy) trees; II – moderately oppressed (moderately weakened) trees; III – strongly oppressed (strongly weakened) trees; IV – dying trees; V – dry trees. Above the line – t/ha, below the line – %.
Значительный интерес представляет оценка вклада деревьев разных категорий состояния в общую фитомассу древостоев средневозрастных сосновых лесов. Установлено, что здоровые (неугнетенные), умеренно ослабленные и сильно ослабленные особи имеют близкую долю (28–34%) в общей фитомассе древостоя и общем запасе стволовой древесины (табл. 4). В общей фитомассе живых неохвоенных ветвей, охвоенных ветвей и хвои доля здоровых (34–36%) и умеренно ослабленных (32–37%) деревьев одинакова, а доля сильно ослабленных (24–26%) является несколько более низкой. Очень низкой долей в общем запасе всех фракций фитомассы (3–14%) отличаются усыхающие особи.
Оценка степени охвоенности крон деревьев разных категорий состояния
На основе данных об относительной массе хвои у модельных деревьев с одинаковыми параметрами среднего диаметра ствола (16 ± 0.3 см) и средней высоты (12.6 ± 0.4 м) был произведен расчет массы хвои деревьев разных категорий состояния, диаметр которых соответствует среднему диаметру в изученных сосновых древостоях. Значения диаметра здоровых, умеренно ослабленных, сильно ослабленных и усыхающих деревьев составили соответственно 18, 16, 12 и 8 см, высота – 12, 12, 11 и 8 м (табл. 1). Величина массы хвои, рассчитанная на основе данных о массе хвои на 1 см диаметра и реальных средних значений диаметров деревьев разных категорий состояния в изученных древостоях составила 6.6, 4.4, 2.8 и 0.7 кг. Величина массы хвои, рассчитанная на основе данных о массе хвои на 1 м высоты и средних значений высот особей разных категорий состояния, составила: 6.6, 4.4, 2.9, 1.0 кг.
В последующем анализе эти величины были использованы для верификации коэффициентов массы хвои, предложенных разными авторами для оценки различий степени охвоения крон (объема функционирующего ассимиляционного аппарата) у деревьев разных категорий жизненного состояния (табл. 5).
Таблица 5.
Коэффициенты массы хвои (плотности кроны) у деревьев разных категорий жизненного состояния Table 5. Needle mass ratio (crown density) in trees of different vitality classes
| Источник Literature source |
Категории состояния Vitality class |
||||
|---|---|---|---|---|---|
| здоровое
(вырубка) healthy (felling) |
здоровое healthy |
умеренно ослабленное moderately weakened |
сильно ослабленное strongly weakened |
усыхающее dying |
|
| Алексеев, 1989 [21] Alekseev, 1989 [21] |
– | 100% (1.0) |
70% (0.70) |
40% (0.40) |
5% (0.05) |
| Manual…, 1994 [23] | – | >90% (1.0) |
75–89% (0.86) |
40–74% (0.60) |
1–39% (0.21) |
| Бебия, 2000 [26] Bebiya, 2000 [26] |
100% (1.0) |
70% (0.70) |
40% (0.40) |
<20% (<0.20) |
|
| Методы…, 2002 [38] Metods…, 2002 [38] |
– | 1.0 | 0.70 | 0.40 | 0.10 |
| Мозолевская, Шарапа, 2003 [24] Mozolevskaya, Sharapa, 2003 [24] |
– | 1.0 | 0.80 | 0.60 | 0.20 |
| Ярмишко и др., 2005 [39] Yarmishko etc, 2005 [39] |
– | 1.0 | 0.71 | 0.43 | 0.14 |
| Данное исследование (по массе хвои) This research (by needle mass) |
– | 1.0 | 0.66 | 0.43 | 0.13 |
| Данное исследование (по массе хвои) This research (by needle mass) |
1.0 | 0.67 | 0.44 | 0.28 | 0.09 |
Приведенные данные показывают, что коэффициенты, предложенные на основе экспертных оценок [5, 23, 24] или равномерных шкал [27, 38] у умеренно ослабленных особей имеют несколько завышенные значения по сравнению с наиболее объективной оценкой по массе хвои. Оценка сильно ослабленных – иногда завышена [23, 24], но чаще корректно отражает различия в массе хвои, а оценка особей, имеющих наиболее низкий уровень жизненного состояния (усыхающих) в разных источниках оказалась либо заниженной [5], либо завышенной [23, 24]. Наиболее близкое соответствие наблюдается с величинами коэффициентов, представленных в работе В.Т. Ярмишко с соавторами [39].
ЗАКЛЮЧЕНИЕ
Результаты исследования, выполненного в средневозрастных северотаежных сосновых лесах, не испытывающих антропогенного воздействия, позволили установить, что 80-летние деревья сосны обыкновенной с одинаковыми размерными параметрами, но имеющие разный уровень жизненного состояния, достоверно различаются по массе охвоенных ветвей и массе хвои. У здоровых деревьев средние значения этих показателей в 1.5, 2 и 4.5 раза превышают значения у умеренно ослабленных, сильно ослабленных и усыхающих особей. Аналогичное соотношение имеют величины относительной массы хвои на единицу высоты и диаметра особей. Здоровые деревья сосны, выросшие на открытом месте, по сравнению со здоровыми особями в составе древостоев отличаются в 2–2.5 раза более высокими показателями массы хвои и охвоенных ветвей. Согласно полученным данным показатели абсолютной и относительной массы хвои имеют достаточно высокую информативность для дифференциации особей разных категорий состояния.
Различия в величине радиального прироста 80-летних модельных особей сосны обыкновенной, имеющих одинаковые размерные параметры и разный уровень жизненного состояния, достоверно проявляются с 60-летнего возраста. В составе древостоев дифференциация разных виталитетных групп деревьев по величине радиального прироста начинается в возрасте 45–50 лет.
Оценка вклада деревьев разных категорий состояния в общую фитомассу древостоев средневозрастных сосновых лесов показала, что здоровые, умеренно ослабленные и сильно ослабленные особи имеют близкую долю (~30%) в общем запасе стволовой древесины. Масса ветвей и хвои здоровых и умеренно ослабленных особей в 1.3–1.5 раза выше, чем сильно ослабленных.
Сравнительный анализ коэффициентов массы хвои деревьев разных категорий состояния, предлагаемых в разных классификациях для интегральной оценки состояния древостоев, свидетельствует, что для умеренно ослабленных деревьев, коэффициенты, как правило, завышаются на 5–30%, а для усыхающих – завышаются или занижаются. Выполненное исследование показало, что если за эталон принять массу хвои здорового дерева (1.0), сформировавшегося в составе древостоя, то коэффициенты умеренно ослабленного, сильно ослабленного и усыхающего деревьев будут равны соответственно, 0.66, 0.43 и 0.13. В том случае, если в качестве эталона (1.0) рассматривать здоровое дерево, сформировавшееся на открытом пространстве, соответствующие коэффициенты составят: 0.67, 0.44, 0.28 и 0.09.
Список литературы
Miller P.R., Millecan A.A. 1971. Extent of oxidant air pollution damage to some pines and other conifers in California. – Plant Disease Reporter 55: 555–559.
Smith W.H. 1981. Air pollution and forests: Interactions between air contaminants and forest ecosystems. New York etc. 379 p.
Цветков В.Ф. 1981. К методике оценки повреждений лесов промышленными выбросами. – В кн.: Природа Севера и ее охрана. Мурманск. С. 28–32.
Алексеев В.А. 1982. Особенности описания древостоев в условиях атмосферного загрязнения. – В кн.: Взаимодействие лесных экосистем и атмосферных загрязнителей. Ч. 1. Таллинн. С. 97–115.
Алексеев В.А. 1990. Некоторые вопросы диагностики и классификации поврежденных загрязнением лесных экосистем. – В кн.: Лесные экосистемы и атмосферное загрязнение. Л. С. 38–54.
Muir P., McCune B. 1987. Index construction for foliar symptoms of air pollution injury. – Plant Disease. 71: 558–565.
Ярмишко В.Т. 1990. Состояние сосновых древостоев. – В кн.: Влияние промышленного атмосферного загрязнения на сосновые леса Кольского полуострова. Л. С. 104–109.
Ярмишко В.Т. 1997. Сосна обыкновенная и атмосферное загрязнение на Европейском Севере. СПб. 210 с.
Vanninen P., Mäkelä A. 2000. Needle and stem wood production in Scots pine (Pinus sylvestris) trees of different age, size and competitive status. – Tree Physiology. 20(8): 527–533. https://doi.org/10.1093/treephys/20.8.527
Черненькова Т.В. 2002. Реакция лесной растительности на промышленное загрязнение. М. 191 с.
Цветков В.Ф., Цветков И.В. 2003. Лес в условиях аэротехногенного загрязнения. Архангельск. 354 с.
Augustaitis A, Šopauskiene D., Baužiene I. 2010. Direct and indirect effects of regional air pollution on the crown defoliation. – Baltic For. 16: 23–34. https://www.balticforestry.mi.lt/bf/PDF_Articles/2010-16[1]/BF10161%2023_24%20Augustaitis%20et%20al.pdf
Lorenz M., Clarke N., Paoletti E., Bytnerowicz A., Grulke N., Lukina N., Sase H., Staelens J. 2010. Air pollution impacts on forests in changing climate. – In: Forest and Society. Responding to Global Drivers of Change. IUFRO World Series (25). Vienna. P. 55–74. https://www.iufro.org/fileadmin/material/publications/iufro-series/ws25/055-074.pdf
Yarmishko V.T. 2015. Radial growth of Scots pine (Pinus sylvestis L.) in varied environment influenced by air pollution in the European North of Russia. – Forestry ideas (Bulgaria). 21(2 (50)): 375-383. https://forestry-ideas.info/issues/issues_Download.php?download=260
Прохоренко Н.Б., Демина Г.В., Мингазова Д.Н. 2017. Оценка жизненного состояния деревьев в урбанизированных условиях Казани. – Известия Самарского научного центра Российской академии наук. 19(2–3): 507–512. http://www.ssc.smr.ru/media/journals/izvestia/2017/2017_2_507_512.pdf
Здорнов И.А., Нагимов З.Я., Капралов А.В. 2018. Санитарное состояние придорожных защитных лесных полос в условиях Северного Казахстана. – Успехи современного естествознания. 3: 44–51.
The State of World’s Forests 2018. FAO Report. 2018. 139 p. http://www.fao.org/3/i9535en/i9535en.pdf
Правила санитарной безопасности в лесах. 2014. – Российская газета. 151(6423). https://rg.ru/2014/07/09/lesa-dok.html
Тарчевский В.В. 1964. Влияние дымогазовых выделений промышленных предприятий Урала на растительность.– В кн.: Растения и промышленная среда. Вып. 1. Свердловск. С. 5–24. http://elar.urfu.ru/bitstream/10995/30154/1/bio_1964_03.pdf
Голутвин Г.И., Кондратов В.И., Поповичев Б.Г. 1980. Динамика состояния сосняков в зоне интенсивного промышленного воздействия. – В кн. Экология и защита леса. Вып. 5. С. 1104–1106.
Алексеев В.А. 1989. Диагностика жизненного состояния деревьев и древостоев. – Лесоведение. 4: 51–57.
Черненькова Т.В. 1986. Методика комплексной оценки состояния лесных биогеоценозов в зоне влияния промышленных предприятий. – В кн.: Пограничные аспекты экологии. Свердловск. С. 116–127.
Manual on methods and criteria for harmonized sampling, assessment, monitoring and analysis of the effects of air pollution on forest. 1994. Hamburg. 177 p.
Мозолевская Е.Г., Шарапа Т.В. 2003. Некоторые результаты применения индекса состояния насаждений при мониторинге. – Лесной вестник. 2: 142–145. https://les-vest.msfu.ru/les_vest/2003/Les_vest_2_2003.pdf
Manual for visual assessment of forest crown condition. 2014. Pristina. 17 p. http://www.fao.org/3/a-i4214e.pdf
Бебия С.М. 2000. Дифференциация деревьев в лесу, их классификация и определение жизненного состояния древостоев. – Лесоведение. 4: 35–43.
Ярмишко В.Т., Горшков В.В., Ставрова Н.И. 2003. Виталитетная структура Pinus sylvestris L. в лесных сообществах с разной степенью и типом антропогенной нарушенности (Кольский полуостров). – Растительные ресурсы. 39(4): 1–19.
Торлопова Н.В., Ильчуков С.В. 2003. Жизненное состояние коренных сосняков Печоро-Илычского биосферного заповедника. – Лесоведение. 3: 34–40. http://lesovedenie.ru/index.php/forestry/article/view/48
Демидко Д.А. 2006. Виталитетная структура ненарушенных древостоев кедра сибирского в субальпийском подпоясе и на верхней границе леса в горном Алтае. – Экология. 37(5): 394–397.
Демидко Д.А. 2011. Состояние кедровых древостоев Северо-Восточного Алтая и методы его оценки. – Лесоведение. 1: 19–27. http://lesovedenie.ru/index.php/forestry/article/view/280
Ставрова Н.И., Горшков В.В., Катютин П.Н. 2016. Формирование структуры ценопопуляций лесообразующих видов в процессе послепожарного восстановления северотаежных лесов. – Труды Карельского научного центра Российской академии наук. Серия Биогеография. 3: 10–28. http://journals.krc.karelia.ru/index.php/biogeo/article/view/187/215
Санитарные правила в лесах СССР. 1970. М. 16 с.
Санитарные правила в лесах Российской Федерации. Положение о государственной лесной охране Российской Федерации. 2006. М. 48 с.
Алексеев А.С. 1997. Мониторинг лесных экосистем. СПб. 116 с.
Шишов Л.Л., Тонконогов В.Д., Лебедева И.И., Герасимова М.И. Классификация и диагностика почв России. 2004. Смоленск. 342 с.
Никонов В.В. 1987. Почвообразование на северном пределе сосновых биогеоценозов. Л. 142 с.
Переверзев В.Н. 2004. Лесные почвы Кольского полуострова. М. 232 с.
Методы изучения лесных сообществ. 2002. СПб. 240 с.
Ярмишко В.Т., Горшков В.В., Ставрова Н.И., Катютин П.Н. 2005. Многолетняя динамика виталитетной структуры древесного яруса хвойных лесов при разном уровне промышленного загрязнения на Кольском полуострове. – В кн. Проблемы экологии растительных сообществ. СПб. С. 58–70.
Молчанов А.А., Смирнов В.В. 1967. Методика изучения прироста древесных растений. М. 100 с.
Шиятов С.Г. 1986. Дендрохронология верхней границы леса на Урале. М. 136 с.
Methods of dendrochronology. Applications in environmental sciences. 1990. Dordrecht. 394 p. https://www.springer.com/gp/book/9780792305866
Манаков К.Н. 1961. Поглощение растительностью минеральных элементов и азота из почвы в лесах Кольского полуострова. – Почвоведение. 8: 34–41.
Вакуров А.Д. 1974. Определение общей фитомассы в сосняках чернично-зеленомошных. – В кн. Продуктивность органической и биологической массы леса. М. С. 11–15.
Зябченко С.С., Иванчиков А.А. 1978. Зональные особенности формирования сосняков черничных Карелии и Кольского полуострова и динамика структуры растительной массы в них. – В кн. Формирование и продуктивность сосновых насаждений Карельской АССР и Мурманской области. Петрозаводск. С. 30–75.
Albrekston A. 1980. Relation between tree biomass fractions and conventional silvicultural measurments. – Ecol Bull. 32: 315–327.
Ingestad T., Agren G. I. 1991. The influence of plant nutrition on biomass allocation. – Ecological Applications. 1(2): 168–174. https://doi.org/10.2307/1941809
Лукина Н.В. 1996. Запас фитомассы древостоев сосняков лишайниковых на северном пределе их распространения. – Лесоведение. 3: 28–37.
Усольцев В.А. 2016. Фитомасса модельных деревьев лесообразующих пород Евразии: база данных, климатически обусловленная география, таксационные нормативы. Екатеринбург. 336 с. http://elar.usfeu.ru/bitstream/123456789/5696/1/Usoltsev_2016.pdf
Лукина Н.В., Никонов В.В. 1996. Биогеохимические циклы в лесах севера в условиях аэротехногенного загрязнения. Апатиты. 213 с.
Буяк А.В., Карпов В.Г. 1983. Сравнительный анализ динамики радиального прироста ели. – В кн.: Факторы регуляции экосистем еловых лесов. Л. С. 65–78.
Vanninen P., Ylitalo H., Sievänen R., Mäkelä A. 1996. Effects of age and site quality on the distribution of biomass in Scots pine (Pinus sylvestris L.). – Trees. 10: 231–238. https://doi.org/10.1007/BF02185674
Wirth C., Schulze E., Schulze W. et al. 1999. Above-ground biomass and structure of pristine Siberian Scots pine forests as controlled by competition and fire. – Oecologia. 121: 66–80. https://doi.org/10.1007/s004420050908
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы