

Растительные ресурсы, 2020, T. 56, № 2, стр. 129-137
Особенности строения и роста корневых систем Pinus sylvestris (Pinaceae) в молодых сосновых лесах Кольского полуострова
В. Т. Ярмишко 1, *, О. В. Игнатьева 2
1 Ботанический институт им. В.Л. Комарова РАН
г. Санкт-Петербург, Россия
2 Санкт-Петербургский государственный лесотехнический университет им. С.М. Кирова
г. Санкт-Петербург, Россия
* E-mail: vasiliyarmishko@yandex.ru
Поступила в редакцию 26.02.2020
После доработки 07.03.2020
Принята к публикации 18.03.2020
Аннотация
Исследованы рост и развитие корневых систем сосны обыкновенной в молодых лишайниковых и лишайниково-зеленомошных сосновых лесах на территории Кольского полуострова. Установлено, что основная масса корней сосны сосредоточена в почве на глубине до 20 см. Сосна обыкновенная формирует характерные морфологические особенности корневых систем уже к 10–12 годам. С возрастом происходит лишь увеличение морфометрических параметров корней и их массы. Установлена линейная связь таксационных показателей деревьев с массой корневых систем в 40-летних древостоях. На сухих песчаных и супесчаных подзолах, где отсутствуют уплотненные слои почвы, радиальный прирост древесины скелетных корней наблюдается практически с одинаковой интенсивностью по всей окружности. На почвах с уплотненными прослойками стержневой корень редуцируется, а рост боковых скелетных корней сосны вниз от анатомической оси происходит значительно медленнее, чем вверх. По мере достижения уплотненных горизонтов почвы и увеличения массы надземной части дерева формирование годичных слоев древесины корней по направлению вниз от анатомической оси прекращается.
Лесной покров на территории Кольского полуострова в настоящее время представляет собой огромную сукцессионную систему, подавляющее большинство процессов в которой инициировано хозяйственной деятельностью человека. При этом как деградацию, так и восстановление лесных экосистем в большинстве случаев невозможно представить, как однонаправленный линейный процесс.
Леса Кольского п-ва, сформированные сосной обыкновенной, произрастают преимущественно на песчаных и супесчаных четвертичных отложениях, в условиях морского и переходного к континентальному климата. Эти леса достаточно устойчивы к суровым природным условиям, но в то же время чрезвычайно чувствительны к воздействию таких факторов, как пожары, рубки, атмосферное загрязнение, которые нередко приводят к их деградации [1–4].
Начавшееся в советское время развитие индустрии на Кольском п-ове потребовало огромного количества древесных ресурсов. Широкомасштабные лесозаготовки были начаты в 30–40-х гг. Объемы рубок определялись потребностью в древесине и техническими возможностями лесозаготовительных предприятий. Вопреки распространенному мнению, именно лесная промышленность нанесла наибольший ущерб природным экосистемам Кольского п-ва. Площадь лесов, нарушенных рубками, в частности наиболее разрушительными сплошными рубками с применением тяжелой тракторной техники, во много раз превышает площадь лесов, пострадавших от воздействия выбросов металлургических комбинатов [5, 6]. В период с 50-х по 80-е гг. были вырублены практически все наиболее доступные для заготовки и продуктивные лесные массивы.
В литературе можно найти большой объем информации об особенностях лесных экосистем Кольского Севера и их состоянии. Это и сведения общего характера [2, 6–8], и подробные данные о воздействии сплошных рубок и пожаров на состояние лесного фонда региона [1, 9, 10], и многочисленные работы, в которых анализируется состояние лесных сообществ, испытывающих воздействие промышленного загрязнения [11–14]. К сожалению, в этих исследованиях содержится крайне мало сведений о подземной сфере лесных сообществ, состоянии корневых систем растений.
Известно, что особенности строения и состояние подземных органов связаны с эколого-биологическими особенностями растений и во многом объясняют их приуроченность к определенным условиям местообитаний, способность конкурировать с другими видами [15–20]. Некоторые авторы отмечают, что сосна обыкновенная может приспосабливаться к разным экологическим условиям, формируя как глубоко проникающую, так и поверхностную корневую систему [6, 17, 21–24]. Пластичность является одной из особенностей корневой системы сосны обыкновенной. Произрастая в широком диапазоне почвенно-гидрологических условий, сосна достаточно активно приспосабливается к их особенностям, изменяя в определенных пределах свои морфологические характеристики [17, 25]. Для формирования корневой системы большое значение имеет мощность корнеобитаемого слоя почвы, которая может ограничиваться избыточным увлажнением и близким залеганием плотных пород или ортштейновых горизонтов.
Целью данной работы является анализ особенностей строения и роста корневых систем Pinus sylvestris L. в разных эдафических условиях при естественном восстановлении нарушенных сосновых лесов на территории Кольского п-ва.
МАТЕРИАЛ И МЕТОДЫ
Исследования проводились на постоянных пробных площадях (ППП), заложенных в семи основных районах Кольского п-ова (табл. 1) на высоте 190–220 м над ур. моря. Основная часть ППП расположена в молодых и средневозрастных лишайниковых и лишайниково-зеленомошных сосновых лесах I–III классов возраста (20–60 лет), представляющих собой различные стадии вторичных сукцессий после рубок и пожаров.
Таблица 1.
Краткая характеристика модельных деревьев сосны обыкновенной разного возраста в исследованных районах Кольского п-ова Table 1. Brief description of Scots pine model trees of different age in the studied areas of the Kola Peninsula
| № п/п No. of SP |
Район исследований Study area |
№ Моделей Number of model tree |
Характеристика исследуемых модельных деревьев Characteristics of the studied model trees | Масса надземной части
модели, кг АСВ Weight of the above-ground part of the model, kg ODW |
Масса корней модели, кг АСВ* The root mass of model, kg ODW* |
||
|---|---|---|---|---|---|---|---|
| возраст, лет age, years |
высота, м height, m | диаметр, см diameter, cm |
|||||
| 1 | Мавринский Mavrinsky |
1 | 20 | 3.7 | 4.5 | 4.11 | 0.68 |
| 2 | 20 | 3.5 | 4.3 | 3.24 | 0.59 | ||
| 3 | 18 | 3.6 | 3.7 | 2.31 | 0.39 | ||
| 4 | 23 | 3.0 | 3.4 | 0.38 | 2.31 | ||
| 5 | 22 | 4.1 | 5.1 | 4.47 | 0.91 | ||
| 2 | Чунозерский Chunozersky |
1 | 43 | 7.6 | 8.0 | 14.8 | 2.82 |
| 2 | 40 | 6.6 | 7.5 | 13.0 | 2.28 | ||
| 3 | 42 | 6.9 | 9.2 | 16.7 | 2.77 | ||
| 4 | 38 | 4.2 | 4.0 | 3.2 | 0.5 | ||
| 5 | 38 | 4.3 | 5.5 | 5.65 | 0.98 | ||
| 3 | Уполокшский Upolokshsky |
1 | 20 | 2.2 | 1.4 | 0.51 | 0.09 |
| 2 | 20 | 2.2 | 1.8 | 0.82 | 0.16 | ||
| 3 | 18 | 1.5 | 0.6 | 0.4 | 0.06 | ||
| 4 | 21 | 3.0 | 3.2 | 2.0 | 0.45 | ||
| 5 | 21 | 4.2 | 4.3 | 3.2 | 0.51 | ||
| 4 | Островской Ostrovsky |
1 | 18 | 1.7 | 1.0 | 0.3 | 0.03 |
| 2 | 18 | 2.4 | 2.3 | 1.24 | 0.14 | ||
| 3 | 17 | 2.0 | 1.2 | 0.57 | 0.06 | ||
| 4 | 20 | 2.4 | 1.9 | 1.17 | 0.13 | ||
| 5 | 22 | 3.7 | 5.2 | 4.88 | 0.83 | ||
| 5 | Ливский Livsky |
1 | 36 | 3.5 | 3.6 | 2.34 | 0.42 |
| 2 | 37 | 4.2 | 4.6 | 3.18 | 0.5 | ||
| 3 | 32 | 2.7 | 3.6 | 1.92 | 0.29 | ||
| 4 | 35 | 3.2 | 3.7 | 2.14 | 0.44 | ||
| 5 | 35 | 3.3 | 3.6 | 2.3 | 0.40 | ||
| 6 | Ловозерский Lovozersky |
1 | 36 | 3.2 | 3.8 | 2.32 | 0.36 |
| 2 | 38 | 4.3 | 5.5 | 5.65 | 0.98 | ||
| 3 | 40 | 4.8 | 5.2 | 6.0 | 1.2 | ||
| 4 | 40 | 4.3 | 5.5 | 5.7 | 1.0 | ||
| 5 | 38 | 6.1 | 6.3 | 6.3 | 1.4 | ||
| 7 | Ковдорский Kovdorsky |
1 | 145 | 14.9 | 16.0 | 66.6 | 27.5 |
| 2 | 240 | 18.5 | 34.5 | 329.6 | 41.8 | ||
| 3 | 90 | 11.0 | 16.6 | 74.0 | 29.0 | ||
| 4 | 160 | 15.1 | 24.5 | 194.6 | 39.4 | ||
| 5 | 146 | 14.2 | 24.3 | 170.2 | 49.1 | ||
Лишайниковые и лишайниково-зеленомошные сосновые леса распространены преимущественно на возвышенных элементах рельефа. Почвообразующими породами являются песчаные и супесчаные моренные и водно-ледниковые отложения, характеризующиеся сухостью и бедностью элементами питания. В условиях северной тайги на них формируются подзолы иллювиально-железистые ненасыщенные, мелкоподзолистые, иллювиально-малогумусовые, средне- и сильно скелетные [26]. Почвы отличаются коротким профилем (до 60 см), накоплением соединений Al и Fe в иллювиальном горизонте, низким содержанием (не более 2%) гумусовых веществ [27].
На каждой пробной площади были выбраны 5–10 модельных деревьев для детальных морфометрических исследований надземных и подземных органов, определения их фитомассы. Изучение корневых систем начинали с поверхностной раскопки, обнажая корни в первую очередь около ствола на глубину 20–25 см. Затем последовательно вскрывали всю корневую систему модельного дерева. На следующем этапе определяли радиус распространения корней в горизонтальной плоскости, производили их зарисовку, определяли количество корней разных порядков, интенсивность их ветвления и состояние, измеряли диаметр и длину основных корней [28]. После детального описания и осторожного удаления почвенных частиц корни взвешивали и отбирали образцы разной крупности для определения абсолютно сухого веса в лабораторных условиях.
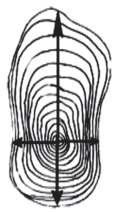
Радиальный прирост крупных скелетных корней II порядка изучали на поперечных срезах на дендрометре LINTAB-6 с точностью до 0.01 мм. Для оценки особенностей роста по диаметру корней, выполняющих не только физиологическую, но механическую функцию, проводили измерения в вертикальном и горизонтальном направлениях по 4-м радиусам: от анатомической оси вверх, вниз и по двум горизонтальным радиусам (рис. 1). В процессе измерений в годичном приросте учитывали долю ранней и поздней части древесины для оценки интенсивности их формирования.
Рис. 1.
Определение прироста скелетных корней сосны обыкновенной по четырем направлениям в вертикальной (вверх и вниз от анатомической оси) и горизонтальной плоскостях. Fig. 1. Determination of the radial increment of Scots pine skeletal roots in four directions in the vertical (up and down from the anatomical axis) and horizontal planes.
При обработке данных использовали методы описательной статистики, корреляционный и регрессионный анализ.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Основная масса боковых корней 40–60-летних деревьев сосны обыкновенной распространяется в подстилке и в верхнем минеральном слое почвы (рис. 2а). В совокупности эти горизонты обладают наилучшими водно-физическими свойствами и аэрацией, содержат основные запасы питательных веществ, а также интенсивнее прогреваются в начале вегетации.
Рис. 2.
Схема строения корневых систем модельных деревьев сосны обыкновенной на Кольском полуострове. а – в молодом возрасте (10–12 лет); b – в возрасте 20–25 лет на щебнистых почвах; c – в возрасте 35–40 лет на хорошо дренированных песчаных и супесчаных почвах. Fig. 2. Root system structure of Scots pine model trees in the Kola Peninsula. a – young aged trees (10–12 years); b – 20–25-year-old trees on gravelly soils; c – 35–40-year-old trees on well-drained sandy and sandy-loam soils.

В процессе развития корневой системы растений сосны меняется физиологическая и морфологическая роль ее отдельных компонентов. На ранних этапах органогенеза стержневой и боковые корни всходов имеют корневые волоски, выполняющие роль сосущих корней. В дальнейшем эти корни начинают выполнять, главным образом, скелетную и проводящую функции. Доля участия главного корня в общей длине корневой системы снижается. В начале формирования корневой системы боковые корни распределяются по длине главного корня относительно равномерно. Однако уже на этой стадии большая их часть разрастается в горизонтальном направлении и во многих случаях радиус корневой системы значительно превышает размеры кроны.
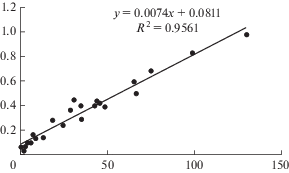
В молодых восстанавливающихся после внешних нарушений (рубок, пожаров) сосняках лишайниковых и лишайниково-зеленомошных процессы роста и развития надземных органов деревьев и формирование их корневых систем протекают синхронно. В ходе исследований установлена тесная линейная связь таксационных показателей деревьев с массой их корневых систем в 40-летних древостоях (рис. 3).
Рис. 3.
Зависимость массы корней сосны обыкновенной в 40-летних древостоях от таксационных характеристик деревьев (произведение высоты (H) на квадрат диаметра (D2)). По горизонтали – H · D2; по вертикали – масса корней, кг АСВ. Fig. 3. Dependence of the mass of Scots pine roots in the 40-year-old stands on the tree characteristics (height (H) times diameter squared (DBH2)). X-axis – H · DBH2; y-axis – mass of roots, kg ODW.
Исследованные сосновые леса произрастают на маломощных, в разной степени завалуненных почвах с выраженными уплотненными горизонтами, образованными ортштейновыми линзами или скоплением валунов. Уплотненные горизонты представляют собой определенное препятствие для проникновения корней вертикальной ориентации вглубь почвы. Корни сосны в этом случае развиваются присущими этой древесной породе темпами лишь в пределах слоя почвы, ограниченного снизу уплотненным горизонтом. Так, например, в местах скопления крупного валунного материала или при наличии близко расположенного к поверхности почвы ортштейнового горизонта главный (стержневой) корень сосны сильно редуцирован или просто отсутствует (рис. 2b). В других случаях, при достижении препятствия в почве главный корень приостанавливает свой рост или меняет направление на горизонтальное. Формируется система мощных горизонтально направленных скелетных корней, обеспечивающих механическую устойчивость надземной части дерева.
Анализ радиального роста скелетных корней сосны обыкновенной в 27-летних сообществах в р-не р. Мавра (Лапландский биосферный заповедник) показывает, что в первое 10-летие прирост скелетных корней происходит примерно одинаково по четырем основным направлениям (табл. 2). В последующий период, по мере увеличения надземной массы и достижения корнями уплотненных горизонтов почвы, их прирост по направлению вниз от анатомической оси замедляется, а затем практически прекращается. Прирост вверх от анатомической оси напротив, возрастает. В результате суммарный прирост корней за весь исследованный период по направлению вверх от анатомической оси оказывается в 3 раза выше, чем по другим направлениям (табл. 2). Следует отметить, что доля ранней части древесины годичного кольца скелетных корней практически одинакова по всем направлениям и колеблется в среднем от ~61 до 64%.
Таблица 2.
Погодичная динамика радиального прироста ранней и поздней древесины скелетных корней сосны обыкновенной во вторичных лишайниковых лесах II класса возраста на Кольском п-ове Table 2. Annual dynamics of Scots pine skeletal root early- and latewood radial increment in secondary lichen forests of age class II on the Kola Peninsula
| Годы Year |
Измерения радиального прироста скелетных корней в вертикальном и горизонтальном направлении
от сердцевины Skeletal root radial increment measured horizontally and vertically form the pith |
|||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Вертикальное – вверх Vertical – upward |
Вертикальное – вниз Vertica – downward |
Горизонтальное – запад Horizontal – West |
Горизонтальное – восток Horizontal – East |
|||||||||||||
| Прирост поздней и ранней части древесины корней, мм Root early- and latewood increment, mm |
||||||||||||||||
| поздняя latewood |
ранняя earlywood |
итого total |
% ранней древесины % of earlywood |
поздняя latewood |
ранняя earlywood |
итого total |
% ранней древесины % of earlywood |
поздняя latewood |
ранняя earlywood |
итого total |
% ранней древесины % of earlywood |
поздняя latewood |
ранняя earlywood |
итого total |
% ранней древесины % of earlywood |
|
| 2014 | 0.8 | 2.05 | 2.85 | 72 | * | * | 0.1 | 0.31 | 0.41 | 75 | 0.1 | 0.43 | 0.53 | 81 | ||
| 2013 | 0.95 | 2.04 | 2.99 | 68 | * | * | 0.11 | 0.23 | 0.34 | 58 | 0.1 | 0.37 | 0.47 | 79 | ||
| 2012 | 1.16 | 2.75 | 3.91 | 70 | * | * | 0.12 | 0.40 | 0.52 | 77 | 0.15 | 0.35 | 0.5 | 70 | ||
| 2011 | 1.0 | 2.52 | 3.52 | 72 | * | * | 0.11 | 0.30 | 0.41 | 73 | 0.12 | 0.34 | 0.46 | 74 | ||
| 2010 | 1.53 | 2.2 | 3.73 | 59 | * | * | 0.13 | 0.5 | 0.63 | 79 | 0.34 | 0.43 | 0.77 | 56 | ||
| 2009 | 0.85 | 2.8 | 3.65 | 77 | * | 0.32 | 0.32 | 100 | 0.15 | 0.25 | 0.4 | 63 | 0.3 | 0.35 | 0.65 | 54 |
| 2008 | 1.07 | 2.2 | 3.27 | 67 | 0.3 | 0.63 | 0.93 | 67 | 0.2 | 0.5 | 0.7 | 63 | 0.14 | 0.24 | 0.38 | 63 |
| 2007 | 1.32 | 2.51 | 3.83 | 66 | 0.45 | 0.62 | 1.07 | 58 | 0.14 | 0.35 | 0.49 | 71 | 0.17 | 0.35 | 0.52 | 67 |
| 2006 | 1.23 | 2.0 | 3.23 | 62 | 0.4 | 0.53 | 0.93 | 62 | 0.11 | 0.28 | 0.39 | 72 | 0.16 | 0.22 | 0.38 | 68 |
| 2005 | 0.6 | 1.7 | 2.3 | 74 | 0.52 | 0.4 | 0.92 | 57 | 0.11 | 0.11 | 0.22 | 50 | 0.14 | 0.33 | 0.47 | 70 |
| 2004 | 0.87 | 1.17 | 2.04 | 57 | 0.3 | 0.43 | 0.73 | 57 | 0.06 | 0.15 | 0.21 | 57 | 0.15 | 0.25 | 0.4 | 63 |
| 2003 | 0.9 | 1.07 | 1.97 | 54 | 0.33 | 0.32 | 0.65 | 49 | 0.09 | 0.12 | 0.21 | 57 | 0.25 | 0.57 | 0.82 | 70 |
| 2002 | 0.63 | 1.16 | 1.79 | 65 | 0.27 | 0.4 | 0.67 | 50 | 0.12 | 0.36 | 0.48 | 75 | 0.28 | 0.42 | 0.7 | 60 |
| 2001 | 0.7 | 1.1 | 1.8 | 61 | 0.32 | 0.75 | 1.07 | 70 | 0.11 | 0.38 | 0.49 | 78 | 0.18 | 0.22 | 0.4 | 55 |
| 2000 | 0.85 | 1.37 | 2.22 | 62 | 0.53 | 0.73 | 1.26 | 68 | 0.18 | 0.5 | 0.68 | 74 | 0.31 | 0.33 | 0.64 | 52 |
| 1999 | 0.55 | 1.28 | 1.83 | 70 | 0.36 | 0.72 | 1.08 | 67 | 0.22 | 0.52 | 0.74 | 70 | 0.25 | 0.43 | 0.68 | 63 |
| 1998 | 0.32 | 0.77 | 1.09 | 71 | 0.7 | 0.58 | 1.28 | 45 | 0.17 | 0.53 | 0.7 | 76 | 0.21 | 0.42 | 0.63 | 67 |
| 1997 | 0.32 | 0.94 | 1.26 | 75 | 0.4 | 0.8 | 1.2 | 67 | 0.4 | 0.54 | 0.94 | 57 | 0.32 | 0.5 | 0.82 | 63 |
| 1996 | 0.51 | 0.29 | 0.8 | 36 | 0.52 | 0.43 | 0.95 | 45 | 0.43 | 0.42 | 0.85 | 49 | 0.33 | 0.62 | 0.95 | 70 |
| 1995 | 0.13 | 0.23 | 0.36 | 50 | 0.12 | 0.33 | 0.45 | 73 | 0.1 | 0.29 | 0.39 | 74 | 0.13 | 0.3 | 0.43 | 70 |
| 1994 | 0.17 | 0.17 | 0.34 | 50 | 0.17 | 0.2 | 0.37 | 54 | 0.09 | 0.17 | 0.26 | 65 | 0.17 | 0.25 | 0.42 | 60 |
| 1993 | 0.1 | 0.12 | 0.22 | 54 | 0.15 | 0.17 | 0.32 | 53 | 0.2 | 0.3 | 0.5 | 60 | 0.15 | 0.16 | 0.31 | 52 |
| 1992 | 0.06 | 0.13 | 0.19 | 58 | 0.16 | 0.22 | 0.38 | 58 | 0.12 | 0.22 | 0.34 | 65 | 0.12 | 0.17 | 0.29 | 59 |
| 1991 | 0.18 | 0.24 | 0.42 | 57 | 0.15 | 0.2 | 0.35 | 57 | 0.18 | 0.23 | 0.41 | 56 | 0.15 | 0.27 | 0.42 | 64 |
| 1990 | 0.2 | 0.24 | 0.44 | 55 | 0.25 | 0.28 | 0.53 | 53 | 0.15 | 0.35 | 0.5 | 70 | 0.16 | 0.14 | 0.3 | 47 |
| 1989 | 0.1 | 0.34 | 0.44 | 77 | 0.2 | 0.3 | 0.5 | 60 | 0.13 | 0.19 | 0.32 | 59 | 0.13 | 0.2 | 0.33 | 61 |
| 1988 | 0.1 | 0.23 | 0.33 | 70 | 0.08 | 0.23 | 0.31 | 74 | 0.1 | 0.26 | 0.36 | 72 | 0.13 | 0.1 | 0.23 | 43 |
| 1987 | 0.1 | 0.3 | 0.4 | 75 | 0.18 | 0.42 | 0.6 | 70 | 0.1 | 0.26 | 0.36 | 72 | 0.14 | 0.29 | 0.43 | 67 |
| 51.22 | 64.1 | 16.87 | 61.5 | 13.25 | 64.4 | 14.33 | 62.7 | |||||||||
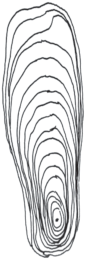
Согласно данным, представленным на рис. 4, в возрасте до 10–12 лет у сосны обыкновенной наблюдается достаточно тесная связь между приростом скелетных корней в вертикальной плоскости вверх и вниз от анатомической оси (r = 0.91; n = 10; P < 0.05). Дальнейшее существенное возрастание радиального прироста вверх от анатомической оси (в 5–9 раз) и одновременное снижение прироста в противоположном направлении приводит к тому, что на определенном этапе развития корневой системы, связь между приростом скелетных корней вверх и вниз от анатомической оси, становится отрицательной (r = –0.33; n = 21; P < 0.05).
Рис. 4.
Динамика радиального прироста скелетных корней 40-летних деревьев сосны обыкновенной на щебенисто-каменистых почвах на Кольском полуострове. По горизонтали – годы; по вертикали – прирост, мм. Сплошная линия – прирост скелетного корня вверх от анатомической оси; пунктирная линия – прирост скелетного корня вниз от анатомической оси. Fig. 4. Dynamics of the skeletal roots radial growth in 40‑year-old Scots pine trees on stony gravel soils in the Kola Peninsula. X-axis – years; y-axis – increment, mm. Solid line – skeletal root increment up from the anatomical axis; dashed line – skeletal root increment down from the anatomical axis.

На щебнисто-каменистых почвах в результате описанных выше процессов у сосны обыкновенной формируются отдельные скелетные корни, имеющие не эллипсовидную форму сечения, а сильно сплюснутую, почти досковидную (рис. 5).
Рис. 5.
Поперечный срез корня доскообразной формы у 60–80-летних деревьев сосны обыкновенной на каменистых почвах на Кольском полуострове. Fig. 5. Cross section of a board-shaped root in 60–80-year-old Scots pine trees on stony soils in the Kola Peninsula.
На достаточно глубоких, свежих, рыхлых супесчаных почвах у сосны обыкновенной в возрасте 35–40 лет развивается глубоко проникающий стержневой корень с хорошо развитыми боковыми корнями (рис. 2c). На сухих песчаных почвах с глубоким залеганием уровня грунтовых вод развивается как стержневой корень, так и поверхностные боковые корни, перехватывающие влагу даже от небольших дождей. К этому необходимо добавить, что на Кольском Севере важная роль в жизни растений вообще и в размещении корней на песчаных и супесчаных подзолах в частности, принадлежит теплообеспеченности корнеобитаемых горизонтов почвы.
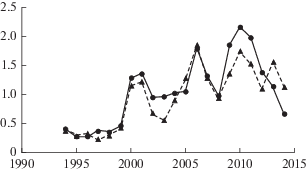
В условиях, когда практически отсутствуют уплотненные слои почвы, радиальный прирост молодых скелетных корней происходит практически с одинаковой интенсивностью во всем направлениям (рис. 6). При этом наблюдается тесная связь между приростом древесины скелетных корней в вертикальной плоскости вверх и вниз от анатомической оси (r = 0.84; n = 20; P < 0.05).
Рис. 6.
Динамика радиального прироста скелетных корней 40-летних деревьев сосны обыкновенной на песчаных и супесчаных почвах на Кольском полуострове. По горизонтали – годы; по вертикали – прирост, мм. Сплошная линия – прирост скелетного корня вверх от анатомической оси; пунктирная линия – прирост скелетного корня вниз от анатомической оси. Fig. 6. Dynamics of radial growth of skeletal roots of 40‑year-old Scots pine trees on sandy and sandy loam soils in the Kola Peninsula. X-axis – years; y-axis – increment, mm. Solid line – skeletal root increment upward from the anatomical axis; dashed line – skeletal root increment downward from the anatomical axis.
ЗАКЛЮЧЕНИЕ И ВЫВОДЫ
Исследования, проведенные на территории Кольского п-ва, показали, что на фоне разных почвенно-экологических условий у сосны обыкновенной сохраняется генетически унаследованный тип корневой системы. Вместе с тем, анализ полученных экспериментальных данных свидетельствует о высокой пластичности корневых систем сосны, что проявляется в вариабельности показателей роста и наличии ряда особенностей их строения в зависимости от условий произрастания в разных районах на исследованной территории. Установлено, что параметры корневых систем сосны обыкновенной определенным образом изменяются под влиянием почвенно-гидрологических и климатических условий, а также хозяйственной деятельности человека. Анализ полученных данных позволяет сделать следующие выводы:
1. В лишайниковых и лишайниково-зеленомошных сообществах на иллювиально-железистых подзолах Кольского Севера сосна обыкновенная образует поверхностную корневую систему. Основная масса корней сосредоточена в верхних почвенных горизонтах на глубине до 20 см.
2. Характерные морфологические особенности корневых систем сосны обыкновенной формируются уже к 10–12 годам. С возрастом принципиальных изменений в их строении не наблюдается, происходит лишь увеличение морфометрических параметров корней и их массы. Установлена линейная связь таксационных показателей деревьев с массой корневых систем на примере древостоев II класса возраста (40 лет). Подобная зависимость сохраняется и в древостоях более высоких классов возраста.
3. На рыхлых, сухих песчаных и супесчаных подзолах, характеризующихся отсутствием уплотненных горизонтов, у сосны обыкновенной формируются глубоко проникающие главный и боковые корни, а также многочисленные поверхностные корни. Радиальный прирост древесины имеет примерно одинаковую интенсивность по всей окружности скелетных корней.
4. При наличии в почвенном профиле уплотненных горизонтов, представляющих препятствие для проникновения корней вглубь почвы, строение корневой системы сосны обыкновенной изменяется. Стержневой корень редуцируется или меняет направление роста на горизонтальное. На границе уплотненных горизонтов интенсивность радиального прироста древесины скелетных корней по направлению вверх от анатомической оси возрастает и становится в 3 раза более высокой, чем по направлению вниз. В отдельных случаях формирование годичных слоев древесины по направлению вниз от анатомической оси у скелетных корней полностью прекращается.
Список литературы
Цветков В.Ф., Семенов Б.А. 1985. Сосняки Крайнего Севера. М. 116 с.
Крючков В.В. 1987. Север на грани тысячелетий. М. 268 с.
Никонов В.В. 1987. Почвообразование на северном пределе сосновых биогеоценозов. Л. 142 с.
Жиров В.К. 2001. Адаптация и возрастная изменчивость растений на Севере. Апатиты. Ч. 1–2. 355 с.
Цветков В.Ф. 1989. Формирование сосняков Кольского полуострова в связи со сплошными рубками. — В сб.: Динамическая типология лесов. М. С. 119–143.
Цветков В.Ф. 2002. Сосняки Кольской лесорастительной области и ведение хозяйства в них. Архангельск. 380 с. https://www.booksite.ru/fulltext/rusles/sosn/text.pdf
Мелехов И.С. 1961. Особенности лесов Кольского полуострова и пути их изучения. М. 150 с.
Рысин Л.П. 1975. Сосновые леса Европейской части СССР. М. 212 с.
Алексеев С.В., Молчанов А.А. 1954. Выборочные рубки в лесах Севера. М. 148 с.
Зябченко С.С. 1984. Сосновые леса Европейского Севера. Л. 247 с.
Лукина Н.В., Никонов В.В. 1993. Состояние биогеоценозов Севера в условиях техногенного загрязнения. Апатиты. 134 с.
Черненькова Т.В. 2002. Реакция лесной растительности на промышленное загрязнение М. 191 с.
Цветков В.Ф., Цветков И.В. 2003. Лес в условиях аэротехногенного загрязнения. Архангельск. 354 с.
Динамика лесных сообществ Северо-Запада России / Под ред. Ярмишко В.Т. СПб. 2009. 276 с.
Рысин Л.П., Рысина Г.П. 1987. Морфоструктура подземных органов лесных травянистых растений. М. 207 с.
Ярмишко В.Т., Цветков В.Ф. 1987. Строение, запасы и распределение в почве корневых систем растений в сообществах сосновых молодняков Кольского полуострова. – Бот. журн. 2(4): 496–505.
Ярмишко В.Т. 1997. Сосна обыкновенная и атмосферное загрязнение на Европейском Севере. СПб. 210 с.
Куприянов А.Н., Заблоцкий В.И. 2004. Формирование корней растений на гарях (юго-западная часть ленточных боров). – Ботан. иссл. Сибири и Казахстана. 10: 31–36.
Салпагарова Ф.С. 2013. Связь удельной длины корней альпийских растений Северо-Западного Кавказа с их эколого-морфологическими признаками. Автореф. дис. … канд. биол. наук. Уфа. 19 с.
Усольцев В.А. 2013. Вертикально-фракционная структура фитомассы деревьев. Исследования закономерностей. Екатеринбург. 602 с. http://elar.usfeu.ru/bitstream/123456789/2771/1/Usolsev_2013.pdf
Лащинский Н.Н. 1981. Структура и динамика сосновых лесов Нижнего Приангарья. Новосибирск. 272 с.
Бобкова К.С. 1987. Биологическая продуктивность хвойных лесов Европейского Северо-Востока. Л. 156 с.
Persson H. 1978. Root dynamics in a young Scots pine stand in central Sweden. – Oikos. 30: 508–519. https://www.jstor.org/stable/pdf/3543346.pdf
Ярмишко В.Т., Лумме И., Ярмишко М.А. 2007. Реакция тонких корней Pinus sylvestris L. на изменение среды их обитания на Карельском перешейке и юго-восточной Финляндии. – В сб. Биоразнообразие, охрана и рациональное использование растительных ресурсов Севера: Материалы XI Перфильевских чтений. Архангельск. С. 93–98.
Ярмишко В.Т., Ярмишко М.А. 2004. Радиальный прирост Pinus sylvestris L. (Pinaceae) на северном пределе распространения. – Ботан. журн. 89(7): 1092–1110. http://arch.botjournal.ru/?t=issues&id=20040707&rid=pdf_0004956
Классификация почв России. 2004. М. 234 с.
Переверзев В.Н. 2004. Лесные почвы Кольского полуострова. М. 232 с.
Методы изучения лесных сообществ. 2002. СПб. 240 с.
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы