

Растительные ресурсы, 2023, T. 59, № 1, стр. 54-64
Компонентный состав кормовых растений и лишайников оленьих пастбищ Надымского района (Ямало-Ненецкий автономный округ)
Е. А. Болдырева *
Тюменский научный центр СО РАН
г. Тюмень, Россия
* E-mail: leta-92@list.ru
Поступила в редакцию 18.09.2022
После доработки 12.12.2022
Принята к публикации 09.02.2023
- EDN: XXOKCI
- DOI: 10.31857/S0033994623010041
Аннотация
В связи с планируемым расширением оленеводства на территории Надымского района Ямало-Ненецкого автономного округа (Западная Сибирь) был изучен элементный состав доминантов растительного покрова тундр и редколесий: листьев карликовой березы Betula nana L.; побегов кустарничков: голубики обыкновенной Vaccinium uliginosum L., багульника болотного Ledum palustre L., мирта болотного Chamaedaphne calyculata (L.) Moench; талломов кустистого лишайника кладонии звездчатой Cladina stellaris (Opiz.) Brodo., побегов сфагновых мхов Sphagnum sp. L. С помощью метода рентгенофлуоресцентного анализа проведена оценка содержания Ca, K, P, Si, Mg, Na, S, Zn, Cu, Ni, Co, Fe, Mn, Cr, Ti и Al в этих растениях. Выявлены биогеохимические особенности исследованных видов, используемых в качестве кормовой базы оленей. Содержание эссенциальных макроэлементов низкое, в особенности в талломах лишайника, где концентрация Ca, K, P более чем на один порядок меньше среднемировых значений. Содержание большинства микроэлементов (Cu, Ni, Co, Cr, Mn) в высших сосудистых растениях и сфагновых мхах, напротив, превышает среднемировые величины. Для Cladina stellaris характерен дефицит микроэлементов. Сравнение наших результатов с данными из других районов тундровой зоны показало сходный характер накопления элементов. Так, листья карликовой березы отличаются накоплением Mg и Mn, содержание Al, Fe, Si повышено в сфагновых мхах, Mn накапливается в побегах кустарничков, для кладонии звездчатой характерен дефицит большинства элементов. Для профилактики заболеваний оленей в зимний период, когда основу рациона составляют лишайники, необходимо увеличение доли “зеленых” кормов.
Оленеводство, являющееся основной отраслью животноводства Ямало-Ненецкого автономного округа (ЯНАО), полностью базируется на естественной кормовой базе. Растительный покров ЯНАО подвергается высокой пастбищной нагрузке [1]. Признаки перевыпаса были отмечены еще в начале XX в. [2]. С тех пор пастбищные нагрузки многократно возросли, а растительность глубоко трансформирована и неуклонно утрачивает ресурсный потенциал [3–9]. Особую опасность для пастбищ представляет разработка многочисленных месторождений нефти и газа. Вблизи мест разведки, добычи и транспортировки полезных ископаемых происходит техногенное разрушение покрова, снижается качество кормовых угодий, сокращается объем кормовых запасов и ухудшается их доступность [10]. Поэтому возможности для развития северного оленеводства ограничиваются количеством и качеством пастбищ. Согласно данным Департамента агропромышленного комплекса ЯНАО [11], за последние 5 лет численность поголовья оленей снизилась на 160 тыс. голов, поскольку оленеемкость пастбищ в 2 раза ниже количества животных.
Обеспеченность пищей северных оленей зависит не только от ее доступности в окружающей среде, но и от ее качества [12]. Для животных важен макро- и микроэлементный состав кормовых видов. Нехватка каких-либо элементов приводит к тому, что происходит снижение массы тел животных, возникают различные заболевания и рождается нежизнеспособное потомство [13].
В последние годы правительство ЯНАО уделяет большое внимание развитию в Надымском р-не изгородного оленеводства, то есть переселения части оленеводов со стадами в таежную зону [14]. От состава растительности и экологической чистоты оленьих пастбищ зависит качество и питательный состав мяса северных оленей [15]. Всестороннее удовлетворение потребностей животных в различных элементах питания обеспечивает наиболее полное проявление генетически обусловленных показателей продуктивности [16]. Микроэлементный состав кормов является одним из важных показателей, определяющих их питательную ценность. Общепризнано выделение заболеваний, характеризующихся недостаточностью (гипомикроэлементозы) и избытком (гипермикроэлементозы) микроэлементов [17]. В системе биогеохимического районирования В.В. Ковальского [13] территория севера Западной Сибири отнесена к таежно-лесной биогеохимической зоне, для которой характерен недостаток многих макро- и микроэлементов: кальция, фосфора, калия, кобальта, меди, йода, молибдена, бора, цинка. Недавние геохимические исследования в Надымской р-не [18–20] подтвердили низкое содержание микроэлементов в почвах. Так, валовое содержание Ni ниже кларка в 5 раз, Mn – в 3 раза, Sr – в 2 раза, Cr – в 1.4 раз [20]. Неблагоприятная биогеохимическая ситуация может отрицательно влиять на оленеводство. Поэтому планы по его развитию в Надымском р-не требуют учета биогеохимических факторов, в частности оценки элементного состава кормовых растений.
Цель данной работы – изучить элементный состав некоторых видов растений, которые составляют кормовую базу оленьих пастбищ на территории Надымского р-на ЯНАО.
МАТЕРИАЛ И МЕТОДЫ
Отбор проб растений производили на территории Надымского р-на Ямало-Ненецкого автономного округа (рис. 1) в летний период (июль–август) 2020 г. в ходе геоботанических исследований на фоновых участках. Пищевой рацион оленя очень разнообразен: в него входит подавляющее большинство растений, произрастающих в тундре и тайге. Летом северный олень питается зеленой растительностью, зимой основной корм оленей – лишайники [21]. Для оценки элементного состава кормовой базы были отобраны доминанты растительного покрова: карликовая береза Betula nana L.; кустарнички: багульник болотный Ledum palustre L., голубика обыкновенная Vaccinium uliginosum L., мирт болотный Chamaedaphne calyculata (L.) Moench; сфагновые мхи Sphagnum sp. L. и кустистый лишайник кладония звездчатая Cladina stellaris (Opiz.) Brodo. На каждой эталонной площадке отбирали не менее 5 экземпляров каждого вида из расчета более 0.2 кг фитомассы, из которых составляли одну сборную пробу.
Рис. 1.
Район исследования. ⚫ – пункты наблюдений и отбора проб. Fig. 1. Research area. ⚫ – observation and sampling points.
В лабораторных условиях пробы высушивали и измельчали в ступке до порошкообразного состояния. Анализ валового содержания элементов выполняли в Институте физико-химических и биологических проблем почвоведения РАН. Содержание Ca, K, P, Si, Mg, Na, S, Zn, Cu, Ni, Co, Fe, Mn, Cr, Ti, Al в кормовых растениях определяли путем измерения массовой доли металлов и оксидов металлов в порошковых пробах на рентгенфлуоресцентном спектрометре. Анализ полученных образцов производили в двух параллельных пробах; за результат измерения принято среднее арифметическое значение.
При статистической обработке результатов рассчитывали среднеарифметическое значение содержания элементов (M) и среднеквадратичное отклонение (SD). Для оценки накопления элементов в растительных образцах использовали коэффициент биологического накопления Кб, который представляет собой отношение содержания элемента в золе растений к величине кларка верхней части континентальной земной коры. При сравнении со среднемировыми показателями содержания элементов в растениях использовали расчеты В.В. Добровольского [22, 23]. Потенциальную физиологическую реакцию на недостаток или избыток элемента в фитомассе определяли путем сопоставления с пороговыми границами, приведенными в работе А. Кабата-Пендиас [24].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Элементный состав исследованных растений представлен в табл. 1. Основными элементами являются (в порядке убывания) Ca, K, Si и P. Содержание практически всех макроэлементов, за исключением Al, ниже среднемировых значений. Обращает на себя внимание низкое содержание таких физиологически значимых, как для растений, так и животных, элементов как Ca, K, P, Mg и Si. Минимальное содержание этих элементов выявлено в лишайнике, а максимальное (кроме Si) – в листьях карликовой березы.
Таблица 1.
Среднее содержание химических элементов в растениях Надымского района, мг/кг абс. сух. веса Table 1. Average element contents in plants of Nadym region, mg/kg dry weight
| Элементы Elements |
Виды растений Plant species |
Среднемировое содержание 1 B. Markert [23] 2 В.В. Добровольский [22] World average content 1 B. Markert [23] 2 V.V. Dobrovolsky [22] |
Экологические градации
по Кабата-Пендиас [24] Environmental levels according to Kabata-Pendias [24] |
|||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Кустарник Dwarf birch Betula nana L. n = 7 |
Кустарнички Dwarf shrubs |
Cladina stellaris (Opiz.) Brodo n =5 |
Sphagnum sp. L. n = 6 |
дефицит deficit |
норма standard |
избыток excess |
||||
| Ledum
palustre L. n = 5 |
Vaccinium uliginosum L. n = 5 |
Chamaedaphne calyculata (L.) Moench n = 2 |
||||||||
| Ca | 4271 ± 3501 | 3560 ± 868 | 3320 ± 602 | 2900 ± 141 | 742 ± 68 | 1283 ± 376 | 10 0001 | – | – | – |
| K | 3086 ± 615 | 2800 ± 583 | 2640 ± 439 | 2000 ± 424 | 858 ± 165 | 1767 ± 596 | 5000–34 0001 | – | – | – |
| P | 1371 ± 496 | 880 ± 377 | 1180 ± 370 | 650 ± 71 | 380 ± 73 | 483 ± 172 | 120–30 0001 | – | – | – |
| Si | 1786 ± 587 | 1600 ± 725 | 1440 ± 321 | 900 ± 141 | 3008 ± 149 | 3067 ± 3065 | 200–80001 | – | – | – |
| Mg | 1500 ± 271 | 900 ± 212 | 1260 ± 167 | 1000 ± 566 | – | 650 ± 243 | 1000–90001 | – | – | – |
| Na | 443 ± 326 | 420 ± 217 | 440 ± 532 | 350 ± 495 | – | 183 ± 271 | 35–10001 | – | – | – |
| S | 829 ± 111 | 740 ± 167 | 1300 ± 300 | 600 ± 0 | 213 ± 29 | 567 ± 82 | 600–10 0001 | – | – | – |
| Al | 1145 ± 142 | 1094 ± 156 | 1171 ± 539 | 1027 ± 117 | 576 ± 267 | 1259 ± 378 | 90–5301 | – | – | – |
| Fe | 1649 ± 80 | 137 ± 119 | 59 ± 8 | 53 ± 12 | 315 ± 213 | 308 ± 243 | 5–2001 | – | – | – |
| Zn | 161 ± 59 | 59 ± 72 | 46 ± 10 | 19 ± 2 | 17 ± 2 | 16 ± 5 | 30.02 | 10–20 | 27–150 | 100–400 |
| Cu | 24 ± 2 | 23 ± 5 | 27 ± 6 | 18 ± 2 | – | 16 ± 5 | 8.02 | 2–5 | 5–30 | 20–100 |
| Ni | 16 ± 2 | 14 ± 2 | 14 ± 2 | 10 ± 2 | 1.2 ± 0.7 | 13 ± 6 | 2.02 | – | 0.1–5 | 10–100 |
| Co | 9 ± 0.60 | 10 ± 3 | 12 ± 7 | 9 ± 3 | – | 9 ± 4 | 0.52 | – | 0.02–1 | 15–50 |
| Mn | 791 ± 485 | 770 ± 208 | 397 ± 194 | 847 ± 1055 | 58 ± 23 | 266 ± 169 | 2052 | 15–25 | 20–300 | 300–500 |
| Cr | 26 ± 6 | 24 ± 6 | 20 ± 3 | 20 ± 3 | 1.0 ± 1.1 | 26 ± 17 | 1.82 | – | 0.1–0.5 | 5–30 |
| Ti | 7 ± 0.39 | 10 ± 6 | 7 ± 1 | 16 ± 14 | 8.0 ± 7.3 | 13 ± 17 | 32.52 | – | – | 50–200 |
Содержание зольных элементов в растениях тундр определяется, прежде всего, составом региональных почв и почвообразующих пород. Так, интенсивное концентрирование растениями Таймыра кальция и магния является следствием повышенного содержания этих элементов в почвообразующих породах, а высокое содержание фосфора в растениях Кольского полуострова обусловлено широким распространением апатитов [25].
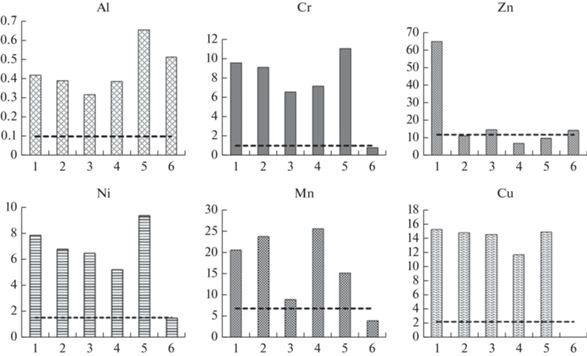
Почвы севера Западной Сибири характеризуются относительно низкими природными концентрациями микроэлементов [19, 26]. Такая же закономерность наблюдается и для макроэлементов. Среднее содержание Ca в почвах Надым-Пуровского междуречья составляет в среднем 0.31% [20], что примерно на порядок меньше кларка верхней части земной коры. Содержание К2О в разных генетических горизонтах почв варьирует от 0.42 до 2.09%, что также меньше кларка, минеральные горизонты постлитогенных почв отличаются недостатком P и S [20]. Низкое содержание Ca, P, S, K в изученных растениях связано с литогенным фактором. Наиболее низкое содержание биологически важных макроэлементов отмечено в лишайнике Cladina stellaris, который составляет основу кормового рациона оленей в зимний период. Этот факт следует учитывать в планах развития таежного оленеводства в Надымской р-не. Имеются указания на минеральное голодание оленей вследствие нехватки Ca, K, P, S [27]. Следует отметить высокое содержание Fe в листьях карликовой березы (ерника) и низкое – в побегах кустарничков. Поглощение железа зависит, прежде всего, от биологических особенностей растений, а не состава почв и пород. Также обращает на себя внимание интенсивное поглощение растениями алюминия. Коэффициенты биологического накопления этого элемента у всех исследованных видов составляют 0.31–0.65, что превышает среднемировые значения в 3.1–6.5 раз (рис. 2). Причиной этого является высокая подвижность Al в ландшафтах кислого глеевого класса. Интенсивное накопление Al типично для тундровых растений. Ранее отмечалось, что Кб этого элемента у некоторых растений (полярная ива, морошка) и лишайников местами больше 1 при средних для растительности суши значениях 0.1 и менее [27].
Рис. 2.
Значения Кб в различных группах фитобиоты Надымского района. По горизонтали – группы фитобионтов; по вертикали – значения Кб. 1 – Betula nana L.; 2 – Ledum palustre L.; 3 – Vaccinium uliginosum L.; 4 – Chamaedaphne calyculata (L.) Moench; 5 – Sphagnum sp. L.; 6 – Cladina stellaris (Opiz.) Brodo. Пунктирная линия – среднемировое значение [22]. Fig. 2. Values of the coefficient of biological accumulation in various floral and lichen biotic groups of the Nadym region. X-axis – plant and lichen biont groups; y-axis – values of the biological accumulation coefficient. 1 – Betula nana L.; 2 – Ledum palustre L.; 3 – Vaccinium uliginosum L.; 4 – Chamaedaphne calyculata (L.) Moench; 5 – Sphagnum sp. L.; 6 – Cladina stellaris (Opiz.) Brodo. The dash line is the world average [22].
Накопление в исследованных растениях микроэлементов имеет несколько иной характер, чем накопление макроэлементов, поскольку его специфика в значительной степени определяется не только влиянием литогенной основы и формами нахождения металлов в почвах, но также видовыми особенностями минерального обмена растений. Несмотря на низкое содержание в почвах севера Западной Сибири многих микроэлементов, в частности Cu, Ni, Co, Cr и Mn [19], в исследуемых видах их содержание превышает среднемировые величины (табл. 1). Только содержание Ti снижено у всех видов сосудистых растений. Таким образом, растения отличаются высокой биогеохимической активностью в отношении накопления микроэлементов.
Интенсивно накапливаются во всех высших сосудистых растениях и сфагновых мхах Ni, Cr, Cu и Mn. Кб для Ni выше среднемирового значения в 3.4–6.1 раз, Cu – в 5.2–6.8 раз, а Cr – в 6.4–10.7 раз (рис. 2).
По имеющимся данным [28], одни элементы в растениях тундровой зоны активно концентрируются (N, P, K, Ca, Mg, S, Cu, Zn, Ni, Co, Sr), другие – деконцентрируются (Al, Zr, Be, Ti, V). Наши исследования подтвердили способность тундровых растений активно накапливать Zn, Cu, Ni, Co, Fe, Mn и Cr. Вероятно, причиной этого является высокая доля подвижных форм металлов, легко усваиваемых растениями. По имеющимся данным [29], из тундрового глеезема в ацетатно-аммонийный буферный раствор переходят 13–16% Ni, Cu и Co. В почвах юга Западной Сибири доля подвижных форм многократно меньше и составляет для Ni 0.6–2.3%, Cu – 0.1–0.9%, Co – 1–3% [30]. Отмечалось, что максимальной подвижностью в тундровых почвах обладают Cu, Zn и Mn [31]. Выявленное нами интенсивное накопление этих элементов сосудистыми растениями подтверждает их высокую подвижность в почвах тундры и предтундровых редколесий.
Наблюдается заметное межвидовое варьирование содержания как микро-, так и макроэлементов. Очень высокое содержание Zn отмечено в листьях Betula nana, что превышает физиологически оптимальный уровень, составляющий, согласно А. Кабата-Пендиас [24], 27–150 мг/кг абс. сух. вещества. Способность карликовой березы накапливать Zn отмечалась ранее многими авторами [32–34]. Кроме того, этот вид отличается повышенным содержанием Ni и Fe. В побегах Chamaedaphne calyculata и сфагновых мхах Zn накапливается умеренно, величины Кб незначительно меньше среднемировых значений (рис. 2).
Среднее содержание меди у разных видов растений варьирует в пределах 16–27 мг/кг абс. сух. вещества, оно выше среднемировых значений, но по экологической градации [24], укладывается в физиологически допустимые нормы. Максимальное содержание Cu отмечено в листьях Vaccinium uliginosum. Та же закономерность была выявлена при изучении химического состава растений Большеземельской тундры, где максимум содержания Cu наблюдается в побегах этого вида [29].
Концентрации Ni варьируют от 10 мг/кг в побегах Chamaedaphne calyculata до 16 мг/кг в карликовой березе. Повышенные содержания никеля объясняются подвижностью этого элемента в кислых и влажных почвах, преобладающих в тундре и тайге. Отмечалось, что рост рН почвы вызывает снижение содержания Ni в растениях [24]. Согласно данным A.I. Syso et al. [35], повышенные концентрации цинка и никеля могут быть типичными для тундровой зоны. Максимальное содержание Cr установлено в сфагновых мхах – 24.5 мг/кг, около 8 мг/кг – в побегах Vaccinium uliginosum и Chamaedaphne calyculata.
Интенсивное накопление Mn отмечено у Chamaedaphne calyculata, минимальное – в сфагновых мхах и лишайнике Cladina stellaris (рис. 2). Повышенное содержание марганца в растениях тундр Западной Сибири отмечается в работах многих авторов [36–38]. Значения коэффициента биологического накопления Кб для Mn, варьировали в пределах от 6.4 (Cladina stellaris) до 29.2 (Chamaedaphne calyculata), в то время как среднемировое значение этого показателя для растительности суши составляет 6.86 [22].
Сфагновые мхи отличаются повышенным содержанием Si, Al, Fe. Повышенное, по сравнению с другими группами растений содержание Si и Аl было отмечено во мхах тундр Чукотки [39]. Для тундрового типа биологического круговорота веществ характерно преобладание таких элементов, как Al, Fe, Mn [27], которые отличаются высокой водомиграционной активностью. Сфагновые мхи, находящиеся в переувлажненных условиях, в наибольшей степени подвержены влиянию поверхностных вод, обогащенных этими элементами.
Для выявления особенностей содержания зольных элементов в растениях в Надымском р-не было проведено сопоставление с другими северными территориями (табл. 2). Сопоставление показало, что в Надымских тундрах содержание калия и фосфора во всех изучаемых растениях находится на среднем уровне. Повышено содержание Si, что, вероятно, вызвано преобладанием песчаных почв. Напротив, содержание Mg в растениях Надымских тундр низкое.
Таблица 2.
Содержание зольных элементов в растениях различных географических районов тундровой зоны, % на сухое вещество Table 2. Ash element content in plants in different geographical areas of the tundra zone, % dry weight
| Растения Plant species |
Элементы Elements |
Сумма элементов Sum of elements | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| P | K | Na | Mg | Al | Fe | Mn | Si | |||
| Карликовая береза Betula nana (листья) Dwarf birch (leaves) |
I | 0.20 | 0.27 | 0.02 | 0.10 | 0.02 | 0.02 | 0.20 | 0.04 | 0.87 |
| II | 0.01 | 0.34 | 0.01 | 0.38 | 0.01 | 0.01 | 0.07 | 0.02 | 0.85 | |
| III | 0.04 | 0.40 | 0.01 | 0.55 | 0.04 | 0.01 | 0.06 | 0.10 | 1.21 | |
| IV | 0.18 | 0.66 | 0.01 | 0.11 | 0.02 | 0.01 | 0.03 | 0.07 | 1.09 | |
| V | 0.11 | 0.21 | 0.01 | 0.22 | 0.07 | 0.10 | 0.16 | 0.08 | 0.96 | |
| Кустарнички Dwarf shrubs |
I | 0.12 | 0.38 | 0.02 | 0.08 | 0.03 | 0.01 | 0.09 | 0.05 | 0.78 |
| II | 0.01 | 0.36 | 0.01 | 0.25 | 0.01 | 0.01 | 0.25 | 0.03 | 0.93 | |
| III | 0.08 | 0.57 | 0.02 | 0.22 | 0.03 | 0.01 | 0.04 | 0.08 | 1.05 | |
| IV | 0.15 | 0.55 | 0.01 | 0.10 | 0.03 | 0.01 | 0.17 | 0.10 | 1.12 | |
| V | 0.09 | 0.24 | 0.04 | 0.21 | 0.12 | 0.02 | 0.04 | 0.32 | 1.08 | |
| Мхи Mosses |
I | 0.21 | 0.63 | 0.04 | 0.19 | 0.08 | 0.04 | 0.03 | 0.11 | 1.33 |
| II | 0.11 | 0.52 | сл. | 0.13 | 0.14 | 0.06 | 0.06 | 0.13 | 1.15 | |
| III | 0.06 | 0.18 | 0.05 | 0.30 | 0.16 | 0.11 | 0.02 | 0.30 | 1.18 | |
| IV | 0.06 | 0.58 | 0.05 | 0.11 | 0.13 | 0.08 | 0.05 | 0.51 | 1.57 | |
| V | 0.03 | 0.09 | 0.06 | 0.04 | 0.05 | 0.03 | 0.05 | 0.25 | 0.60 | |
| Лишайники Lichens |
I | 0.07 | 0.04 | 0.04 | 0.02 | 0.11 | 0.03 | 0.01 | 0.05 | 0.37 |
| II | 0.05 | 0.08 | – | 0.09 | 0.02 | 0.04 | 0.05 | 0.10 | 0.43 | |
| III | 0.05 | 0.35 | 0.06 | 0.06 | 0.06 | 0.02 | 0.01 | 0.07 | 0.68 | |
| IV | 0.04 | 0.08 | 0.03 | 0.03 | 0.05 | 0.03 | 0.01 | 0.18 | 0.45 | |
| V | 0.03 | 0.10 | 0.02 | 0.02 | 0.04 | 0.04 | 0.01 | 0.16 | 0.42 | |
Примечание. I – Кольская тундра [40]; II – Большеземельская тундра [41 ] ; III – тундры стационара “Агапа”, Таймыр [25]; IV – арктическое побережье Чукотки [39]; V – растительный покров Верхнеколымского нагорья [42 ] ; прочерк – нет данных. Note. I – Kola tundra [40]; II – Bolshezemelskaya tundra [41 ] ; III – tundra of Agapa observation station, Taimyr [25]; IV – Arctic coast of Chukotka [39]; V – vegetation of the Upper Kolyma Plateau [42 ] ; dash – no data.
В целом необходимо отметить, что тундровые растения разных географических районов имеют сходный характер накопления элементов. Листья Betula nana отличаются накоплением Ni, Ca, Zn, Cr, содержание Al, Fe, Si повышено во мхах, Mn накапливается в кустарничках и карликовой березе. Лишайники характеризуются низким содержанием K и Ca.
ЗАКЛЮЧЕНИЕ
Исследование элементного состава растений, составляющих основу кормовой базы оленьих пастбищ на территории Надымского р-на Ямало-Ненецкого автономного округа: листьев карликовой березы Betula nana L.; листьев и побегов кустарничков: голубики обыкновенной Vaccinium uliginosum L., багульника болотного Ledum palustre L., мирта болотного Chamaedaphne calyculata (L.) Moench; талломов кустистого лишайника кладонии звездчатой Cladina stellaris (Opiz.) Brodo, побегов сфагновых мхов Sphagnum sp. L. показало, что растения тундр отличаются низким содержанием эссенциальных макроэлементов (K и Ca), что, вероятно, обусловлено литогенным фактором – дефицитом этих элементов в преобладающих песчаных почвах. Высшие сосудистые растения и сфагновые мхи активно накапливают Cu, Ni, Co, Cr и Mn, содержание которых превышает среднемировые величины. Максимальное накопление этих элементов выявлено в листьях Betula nana. Содержание Al повышено по сравнению со среднемировым уровнем из-за высокой доли подвижных форм, легко усваиваемых растениями. Лишайник Cladina stellaris, составляющий основу рациона оленей в холодный период года, содержит крайне малое количество физиологически необходимых макроэлементов, в особенности K и Ca, а также микроэлемента Mn и Zn. Поэтому сбалансированное поступление микроэлементов в организм северного оленя требует сочетания зеленых и лишайниковых кормов.
Список литературы
Полуостров Ямал: растительный покров. 2006. Тюмень. 360 с.
Житков Б.М. 1913. Полуостров Ямал. – Записки РГО. Т. XLIX. 449 с.
Магомедова М.А. 1986. Антропогенная динамика лишайникового покрова на Тюменском севере Западной Сибири. – В сб.: Биологические проблемы Севера: Тез. докл. XI Всесоюзного симпозиума. Якутск. Вып. 2. С. 84–86.
Магомедова М.А. 1994. Лишайники предтундровых лесов Западной Сибири. – Бот. журн. 79(11): 1–11. http://arch.botjournal.ru/?t=issues&id=19941111&rid=pdf_0005103
Магомедова М.А. 1994. Мониторинг состояния растительного покрова оленьих пастбищ. – Проблемы региональной экологии. 3: 76–80.
Магомедова М.А. 1996. Лишайники как компонент северных экосистем и объект мониторинга. – Проблемы экологического мониторинга и моделирования экосистем. 16: 105–121.
Природа Ямала. Ред Л.Н. Добринский. 1995. Екатеринбург. 435 с.
Мониторинг состояния биоты в зоне воздействия промкомплекса по добыче и транспортировке газа на полуострове ЯМАЛ. Научный отчет. Екатеринбург, 1993. 328 с.
Магомедова М.А., Морозова Л.М. 2000. Восстановление оленьих пастбищ. – В сб.: Восстановление земель на Крайнем Севере. Сыктывкар. С. 126–134.
Иванов В.А. 2014. Оленеводство в арктическом субрегионе: состояние и направления развития. – Регион: Экономика и социология. 2(82): 39–51. https://www.elibrary.ru/item.asp?id=21946406
Доклад о ходе реализации и оценке эффективности реализации государственной программы Ямало-Ненецкого автономного округа “Развитие агропромышленного комплекса, рыбного хозяйства и регулирования рынков сельскохозяйственной продукции, сырья и продовольствия” за 2021 г. Салехард, 2022. 25 с. https://dapk.yanao.ru/documents/reports/165011/
Абатуров Б.Д. 2005. Кормовые ресурсы, обеспеченность пищей и жизнеспособность популяций растительноядных млекопитающих. – Зоологический журнал. 84(10): 1251–1271. https://www.elibrary.ru/item.asp?id=9174263
Ковальский В.В., Андрианова Г.А. 1970. Микроэлементы в почвах СССР. М. 182 с.
Волковицкий А.И., Терёхина А.Н. 2020. Современные проблемы Ямальского оленеводства: дискуссии и перспективы. – Этнография. 2(8): 152–169. https://doi.org/10.31250/2618-8600-2020-2(8)-152-169
Роббек Н.С., Алексеев Е.Д., Румянцева Т.Д. 2019. Содержание микроэлементов и тяжелых металлов в мясе оленей чукотской породы (харгин). – Главный зоотехник. 7: 60–65. https://www.elibrary.ru/item.asp?id=39162827
Сариев А.Х., Чербакова Н.Н., Дербенев К.В., Федина Е.В. 2019. Химический состав кормовых растений, произрастающих в зоне влияния промышленных предприятий. – Вестн. КрасГАУ. 5: 68–74. http://www.kgau.ru/vestnik/2019_5/content/10.pdf
Ермаков В.В., Тютиков С.Ф., Сафонов В.А. 2018. Биогеохимическая индикация микроэлементозов. М. 386 с.
Moskovchenko D., Shamilishvilly G., Abakumov E. 2019. Soil biogeochemical features of Nadym-Purovskiy province (Western Siberia). – Ecologia Balkanica. 11(2): 113–126.
Опекунова М.Г., Опекунов А.Ю., Кукушкин С.Ю., Ганул А.Г. 2019. Фоновое содержание химических элементов в почвах и донных осадках севера Западной Сибири. – Почвоведение. 4: 422–439. https://doi.org/10.1134/S0032180X19020114
Романенко Е.А., Московченко Д.В., Кудрявцев А.А., Шигабаева Г.Н. 2020. Подвижные формы металлов в почвах Надым-Пуровского междуречья (Западная Сибирь). – Вестник Нижневартовского университета. 2: 136–145. https://doi.org/10.36906/2311-4444/20-2/18
Бороздин Э.К. 1990. Северное оленеводство. Л. 240 с.
Добровольский В.В. 2003. Основы биогеохимии. М. 400 с.
Markert B. 1992. Presence and significance of naturally occurring chemical elements of the periodic system in the plant organism and consequences for future investigations on inorganic environmental chemistry in ecosystems. – Vegetatio. 103(1): 1–30. https://doi.org/10.1007/BF00033413
Kabata-Pendias A. 2011. Trace elements in soils and plants. Boca Raton, FL, USA. 505 p.
Василевская В.Д. 1980. Почвообразование в тундрах Средней Сибири. М. 235 с.
Московченко Д.В. 2013. Экогеохимия нефтедобывающих районов Западной Сибири. Новосибирск. 259 с.
Перельман А.И., Касимов Н.С. 1999. Геохимия ландшафта. М. 764 с.
Глазовская М.А. 1988. Геохимия природных и техногенных ландшафтов СССР. М. 328 с.
Елькина Г.Я., Денева С.В., Лаптева Е.М. 2019. Тяжелые металлы в системе почва-растение в биогеоценозах Большеземельской тундры. – Теоретическая и прикладная экология. 3: 41–47. https://doi.org/10.25750/1995-4301-2019-3-041-047
Сысо А.И. 2007. Закономерности распределения химических элементов в почвообразующих породах и почвах Западной Сибири. Новосибирск. 277 с.
Попова Л.Ф., Никитина М.В., Андреева Ю.И., Трофимова А.Н., Попов С.С. 2019. Тяжелые металлы в почвах евро-арктических территорий. – Изв. вузов. Химия и хим. технология. 62(3): 102–107.
Московченко Д.В. 1995. Биогеохимические особенности ландшафтов полуострова Ямал и их оптимизация в связи с нефтегазодобычей: Автореф. дис. … канд. геогр. наук. Санкт-Петербург. 24 с.
Тентюков М.П. 2013. Особенности распределения химических элементов в мерзлых почвах. – Криосфера Земли. 17(3): 100–107.
Башкин В.Б. 2017. Биогеохимические циклы в тундровых экосистемах импактных зон газовой индустрии. – Геохимия. 55(10): 954–966. https://doi.org/10.7868/S0016752517100028
Сысо А.И., Колпащиков Л.А., Ермолов Ю.В., Черевко А.С., Сиромля Т.И. 2014. Элементный химический состав почв и растений Западного Таймыра. – Сиб. Экол. журн. 6: 855–862. https://www.sibran.ru/upload/iblock/c12/c1247fd74f4347cd33d9c329daeac23d.pdf
Московченко Д.В., Моисеева И.Н., Хозяинова Н.В. 2012. Элементный состав растений Уренгойских тундр. – Вест. экологии, лесоведения и ландшафтоведения. 12(1): 130–136. https://www.elibrary.ru/item. asp?id=18050792
Agbalyan E.V., Zarov E.A., Filippov I.V., Pechkina Yu.A., Shinkaruk E.V., Yulbarisova K.V., Khnycheva N.A., Lapshina E.D. 2021. Biogeochemical assessment of the main plants at the Nadym district’s reindeer pastures (YamalNenets autonomous okrug). – Environmental dynamics and global climate change. 12(1): 27–41. https://doi.org/10.17816/edgcc41876
Московченко Д.В., Романенко Е.А. 2021. Микроэлементный состав кормовых растений оленьих пастбищ полуострова Ямал. – Раст. ресурсы. 57(4): 370–381. https://doi.org/10.31857/S0033994621040099
Пугачев А.А., Тихменев Е.А. 2016. Химический состав растений арктического побережья Чукотки. – Изв. Самарского науч. центра РАН. 18(5(3)): 478–483. http://www.ssc.smr.ru/media/journals/izvestia/2016/2016_5_478_483.pdf
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы