

Российский физиологический журнал им. И.М. Сеченова, 2020, T. 106, № 7, стр. 880-889
Event-Related Changes In EEG Spectral Power Corresponding to Creative and Trivial Decisions
N. V. Shemyakina 1, *, **, Zh. V. Nagornova 1
1 Sechenov institute of evolutionary physiology & biochemistry Russian academy of sciences
St. Petersburg, Russia
* E-mail: natalia.shemyakina@iephb.ru
** E-mail: shemyakina_n@mail.ru
Поступила в редакцию 5.11.2019
После доработки 22.03.2020
Принята к публикации 4.05.2020
Аннотация
The study aimed to explore EEG correlates of a person’s creative (non-trivial) and trivial decisions finding in conditions of associative search by means of event-related synchronization/event-related desynchronization (ERS/ERD) spectral power changes analysis. Sixty subjects had to fulfill RAT (Remote associative test) alike tasks while EEG registering. There were no differences in time responses while remote association and trivial decisions search (around 3 seconds in both case). An increase in ERS was observed for the central frequencies 4 Hz at F7 site (1500–1000 ms before note of decision), 3 Hz at the Fz site (500–1000 ms before note of decision), and 3.5 Hz at T3 site (in the last 500 ms before decision). Obtained findings stressing the role of low frequency bands (theta and delta) in wide associative field search leading for creative and original decisions.
Creative activity is a complex and multifactorial mental activity that combines memory, attention, decision-making, motivation, and different types or strategies of thinking. In the case of creative activity, the transformation of existing, or the appearance of new, original associations, knowledge, and products occurs.
Creative thinking has long been investigated by psychologists in terms of the structure of intelligence and abilities of the person [1]. Various tests have been developed to determine the creative abilities level of the person and investigations conducted into the personal characteristics of more and less creative people [2–5].
At the same time, it is possible to assume that in their everyday lives each person is capable of making both creative and trivial decisions, and the brain activity in relation to qualitatively different decisions differs [6]. From the “evolutionary point of view” both trivial and creative decisions have their advantages, as the first can save the time in the critical situation, the second – can suggest a new approach. The question of distinctions in electroencephalographic (EEG) correlates of creative and trivial decisions in the same person is insufficiently investigated. Exploration of this question can matter for understanding brain mechanisms [7, 8].
According to Mednick’s theory of creative thinking [5] in the course of creative activity new associative links have to be formed. At the same time, the more remote the characteristics and elements of a problem people can connect together, the more creative and original such decisions and processes will be.
Based on this provision of associative theory, within the present research the task was focused on initiation of a verbal associative search, leading to finding creative (association of remote characteristics of concepts) and trivial decisions in the conditions of the same instruction. Today, information exists on the physiological correlates of brain maintenance of creativity [9–13]. However, there is little information on what actually occurs at the moment a person makes a creative, original decision and how processes providing creative decisions in the brain differ from processes providing trivial decisions in the conditions of the same instruction.
One of the informative parameters of a change in brain bioelectric activity, which allows the chance to obtain additional information on when distinctions occur, is the time and frequency transformation of the EEG signal in event-related synchronization/desynchronization (ERS/ERD) EEG analysis. [14, 15]. The study aimed to explore EEG correlates of a person’s creative (non-trivial) and trivial decision-making in conditions of associative search by means of ERS/ERD spectral power changes analysis.
METHODS
Participants. In total, 60 native Russian-speaking participants took part in the study (18 males, mean age 27 ± 3, two left-handed [16]), with education above the secondary school level. At the time of the study, all participants were healthy and were not following any medical treatment. Subjects gave their voluntary agreement to participate in the psychophysiological study, which was conducted in accordance with all the ethical standards of the 1964 Helsinki Declaration and its later revisions.
Tasks and procedure of the investigation. The experimental task [17] was based on the modified Remote Associates Test (RAT) [5]. The RAT is widely used in psychophysiological studies of creativity for inducing states of remote associative search and insight [10, 11]. The designed experimental task preserved the remote associative search and could lead to finding of both trivial and creative solutions.
However, differently from the classical RAT, our task consisted of words from different/remote semantic fields and many brief trials with the same instruction — to “be original”. A lot of trials were necessary to fit the stimulus-response ERP paradigm of a psychophysiological study. Participants were presented with 444 noun pairs of singular nominative form, IPM (items per million) >10. Nouns were of one, two, three, four, or five syllable length, with similar or comparable length in each pair — e.g., “cake” and “towel”. Participants were asked to come up with a non-trivial/original definition/adjective for each of the noun pairs. As an example, the remote association between the above words (“cake” and “towel”) in Russian could be “waffle”. We expected the given instruction to facilitate subjects’ associative search for remote associations between nouns in the pairs.
Participants were seated in a comfortable chair in a separate shielded room. Stimuli presentation was performed via a 17-inch LCD monitor placed 1.5 meters away from the participant’s head; the average visual angle of stimuli presentation was 4.9' × 2.1'. Nouns were presented in black bold typeset above and below the fixation cross to minimize horizontal eye movements. For the first 300 ms, participants saw a black cross on a white screen. Each noun pair was shown for 400 ms in the center of the white screen. Following the noun pair presentation, the participant was given 4500 ms to think of a non-trivial, original definition linking the two given nouns (belonging to remote semantic fields) which could also be associated and used with each of the nouns separately. If the participant came up with a definition during this creative task time, she/he had to press the button with her right forefinger and, after seeing the question mark on the screen at the end of the creative task period (5100 ms from the trial onset), tell the researcher the suggested definition via microphone. If no definition was found for the two nouns and there was no pushing of the button, the participant had to say ‘no’ after the presentation of the question on the screen. All the participants’ answers were recorded via the audio system for the following data analysis. The interval between the trials varied randomly between 1500 and 2500 ms, and trials’ presentation were also randomized across the participants to minimize order and fatigue effects. The complete investigation, together with all preparation and breaks, lasted around 70 minutes.
EEG data recording. Electroencephalogram (EEG) was recorded using the Mitsar 31 channel EEG system (Mitsar Ltd., St. Petersburg, http://www.mitsar-medical.com) by means of the WinEEG software package (V.A Ponomarev, Ju.D Kropotov, register for computer program RF № 2001610516, 08.05.2001). Silver chloride electrodes were positioned according to the 10–20 system (Fp1, Fp2, F7, F3, Fz, F4, F8, T3, C3, Cz, C4, T4, T5, P3, Pz, P4, T6, O1, O2). The input signals were referenced to the linked ears, were filtered between 0.53 Hz and 30 Hz, and were digitized at a rate of 500 Hz, notch filter 45–55 Hz. The ground electrode was located on the left hand. Resistance of the electrodes did not exceed 5 kOhm.
EEG data analysis. All the data were initially transformed by WAR (weighted averaged reference) [18] in order to diminish the influence of common reference. Eye blink artifacts were corrected by zeroing the activation curves of individual independent components corresponding to eye blinks. These components were obtained by application of independent component analysis (ICA) to the raw EEG fragments. The method has been previously described [19–21]. High and low frequency activities were marked as artifacts and were excluded from further analysis. The thresholds were set as follows: (1) 50 µV for slow waves in the 0–1 Hz band; (2) 35 µV for fast waves in the 20–35 Hz band.
For the data analysis, we synchronized participants’ EEG records on button pressing and investigated the time interval of EEGs before the button pressing to figure out distinctions in the EEG signal changes connected with the different quality of tasks performed (original, trivial). The desynchronization and synchronization of EEG connected with the events were calculated by means of Morlet wavelet transformation [22]: the frequency-time characteristics of the EEG signal were evaluated for the 5000 ms analysis period (2000 ms before pressing the button i.e., when finding the decision and 3000 ms after pressing the button) for different central frequencies in the range of 2–7.5 Hz with 0.5 Hz steps, with a five cycle width of the wavelet. To reduce data dispersion, the spectral power dynamic of EEG was smoothed by moving averages with the width of an average period equal 100 ms.
Participants’ trials with RT (response times) of less than 2000 ms were excluded from analysis to diminish the influence of the stimulus presentation on the EEG. To define how many experimental trials (short fulfilled tasks) were enough for averaging EEG data and ERS/ERD analysis, group dispersion of average EEG power in each analyzed time interval for chosen electrode was evaluated empirically with different numbers of averaged trials. When averaging more than 40 and 50 trials of each type (trivial, non-trivial answers), dispersions were practically unchanged; when averaging 30 trials, group dispersion of the average EEG power increased. Thus, for each participant, 40 was the optimum number of trials for averaging. Those subjects who had fewer than 40 artifact free trials of each type were excluded from data analysis. Values of EEG power signal dynamics for the remaining subjects (21 persons, six men, age 26.6 ± 6 years) were averaged. For reduction of the influence of ERP components on ERS/ERD, ERPs were calculated and subtracted from each fragment of EEG [23]. Distinctions in the ERS/ERD were investigated in the range of interest: frequencies of EEG (2–7.5 Hz) with a step of 0.5 Hz in three 500-millisecond intervals (1500–1000 ms; 1000–500 ms; 500–0 ms) before pressing the button. Thus we focused on two bands of interest (Δ: 2–3.8 Hz and θ: 4–7.5 Hz). These frequency bands are described as playing a role in verbal memory processes and associative search [24–27].
The obtained values of absolute power for the specified time windows for each EEG site on each person were taken under logarithm as the normalization of data. The statistically significant differences of EEG power in different conditions were evaluated by means of unpaired Student’s t-test that was chosen as provided a smaller number of positive results. The regions of the interest were assessed by the differential time-frequency maps between original and trivial findings. The power differences were observed in frontal and temporal regions of the left hemisphere. The non-accidental spectral power differences were estimated as significant differences with p < 0.01 in the regions of interest and p < 0.01 (in some cases 0.05) in the nearby ranges of frequencies, or in the nearby intervals of time. Probabilities were corrected for multiple comparisons using a false discovery rate procedure (http://sdmproject.com/utilities/?show=FDR).
RESULTS AND DISCUSSION
According to the literature data, the search for remote associations during RAT performance could be considered as a creative process leading to an original decision [28]. Remote associations [5] combine concepts on the basis of their non-obvious characteristics (their figurative/extended meaning/sense).
In the present research, subjects were asked to find an original unifying definition of the nouns shown in pairs. Subjects’ answers that focused on the determination of non-obvious/implicit signs of the concepts (making figurative sense) were considered creative and original, while those making literal sense, defining obvious external characteristics such as the size, color, or form of an object, were considered trivial.
We were interested in whether there are any physiological distinctions in subjects’ finding of qualitatively different (creative and trivial) decisions in conditions of similar instruction and performance of the same activity — searching for unifying definitions. On the basis of subjects’ answers, three types of trial were revealed: a) those connected with finding original, creative answers; b) those connected with finding trivial answers; and c) trials without answers. We did not consider the last type of trial as there was no possibility of their synchronization on pressing the button.
Explored subjects on average gave 295 ± 60 answers, of which 119 ± 37 on average were focused on the determination of non-obvious characteristics of the concepts and were considered by us to be original [28] and creative.
The average time for finding the remote defining association was about 3 ± 0.24 seconds, while for the trivial it was 3 ± 0.29 seconds: those times did not differ statistically. It should be noted that in a research, Jung-Beeman and colleagues [29] spent time finding insightful decisions (that unexpectedly came to mind) and non-insightful decisions (logically built) upon implementation of the modified RAT test that also did not reveal significant differences.
Furthermore, at separate EEG sites, for separate frequencies with 0.5 Hz step, Student’s t-test was applied for statistical assessment of EEG event-related power changes in the 2–7.5 Hz range.
When comparing average EEG power values (corresponding to original and trivial answers from subjects) in various frequency bands and various intervals of time, for the majority of EEG sites, statistically significant distinctions were not revealed or were observed chaotically.
Apparently statistically significant distinctions that appeared in the case of plurality of comparisons we considered as false alarms (statistical errors of the first type). Only at sites F7, T3, and Fz were statistically significant differences revealed (p < 0.01, and in some cases 0.05, which were in the nearby ranges of frequencies, or in the nearby intervals of time, or both). As it is rather difficult to explain these group of significant distinctions only as a result of multiple comparisons, we considered them as statistically significant effects. The greatest statistically significant differences were observed for the central frequency of 3 Hz at the Fz site, 3.5 Hz at T3, and 4 Hz at F7. These differences were significant after FDR correction (p < 0.05). Graphical representation of the dynamics of EEG power at the specified sites during the finding of original (associative) definitions and trivial answers are presented in Fig. 1.
Fig. 1.
Event-related changes of EEG power during the finding of original (associative) and trivial decision.
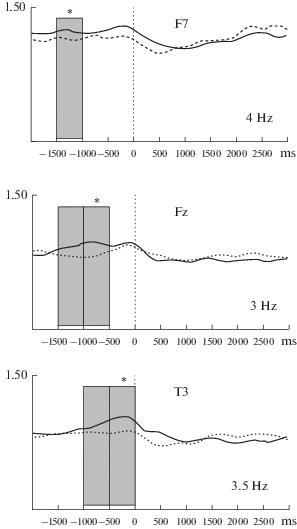
The illustration is given for a time interval 5000 ms (2000 ms before pressing the button, 3000 ms after). The continuous line – the solution of a task was followed by finding of original answers, a dashed line – the solution of a task was followed by finding of trivial answers. A vertical dashed line – pressing the button as the mark of decision finding. Ordinate axis – P (μV2), abscissa axis – time (ms).
Grey bars mark time windows of 500 ms for those site, that revealed the significant (p < 0.05) differences of event-related changes of EEG power. Star marks the time windows that revealed the maximum difference for the central frequency of wavelet. The central frequency of wavelet for different sites is presented in the in the bottom right corner.
At the F7 site, differences in EEG power during the search for original (remote) associations were observed in the time interval 1500–1000 ms before pressing the button, with the central frequency 4 Hz: differences (taking into account the properties of the wavelet) were also observed in the interval for the nearby frequencies (2.5–5.5 Hz). We do not exclude the possibility that the specified time interval could also be covered by the duration of visual ERP (800–1000 ms) and the reaction of the synchronization in delta band frequencies [30, 31] for some numbers of averaged trials with RT of 2000 ms. However, observed differences could hardly be related to the latter, as the fronto-temporal site (lateral frontal cortex) in the left hemisphere of the brain is not a typical zone for distribution of visual ERP and reactions of synchronization (traditional distribution of the visual ERP is much more pronounced in the parietal and occipital areas of the cortex). In this situation, observed differences can be connected with the specificity of the performed task, in one case, leading to the finding of an associative remote definition and, in another, to a literal definition of characteristics of concepts in conditions of similar instruction.
At Fz site, the maximum difference was observed for the central frequency 3 Hz (2.5–4.2 Hz) in the time interval of 500–1000 ms before pressing the button. At T3, the maximum distinction in power was observed for the central frequency 3.5 Hz (2.5–6 Hz) in the last 500 ms before pressing the button. It is known that spontaneous pressing of the button is connected with the emergence of readiness potential about 500 ms before pressing [32, 33]. Pressing is followed by a desynchronization in the sensorimotor cortex zones, mostly in the contralateral hemisphere at a frequency greater than 10 Hz [34, 35]. In the range of slow frequencies, emergence of EEG reaction at the central sites is possible [36, 37]. However, in our case, spontaneous pressing was present in all trials considered, and localization of observed differences (average values of absolute power and the temporal zone in the cortex in the left hemisphere of the brain were greater than at Fz site) does not allow us fully to correlate observed differences in brain reaction when pressing the button.
From this, it is possible to conclude that the finding of creative decisions was followed by a higher level of synchronization of the event-related low-frequency component of EEG in the fronto-temporal and middle temporal regions of the left hemisphere of the brain, as well as in the fronto-central region. This does not exclude the possible existence of changes in other frequency ranges, but indicates the importance of the synchronization of a low-frequency component of EEG for brain processes in associative search and related states. Moreover, in a number of studies (for a review see [38–40]) addressing the nature of EEG signals, it has been noted that synchronous low-frequency (slow wave) activity can modulate the high-frequency component of a bioelectric signal when cognitive processes occur.
The obtained results are partly in agreement with the data of our colleagues Danko et al. [41], who revealed an increase of power in the delta (1.5–3.5 Hz) EEG band in the front (bilaterally) and middle temporal areas of the left hemisphere of the brain during fulfilment of verbal creative tasks (creation of associative chains) focused on use of an insight strategy in tasks’ performance by non-actor students. For the general group of subjects, including both student actors and student non-actors, bilateral increase in power in the delta and theta (4–7 Hz) EEG frequency bands was observed in the frontal cortex zone during the creation of associative chains. The results of the present research can also be correlated to data from Razumnikova, who found an increase in EEG power in the teta-1 (4–6 Hz) frequency band in the frontal cortex areas when finding remote associations (RAT was used) in comparison with the finding of simple associations [27]. In the earlier work of Whitton [42], it was reported that four seconds before healthy subjects gave their creative answers when performing tasks with creative thinking, slow activity in the delta and theta frequencies appeared in their EEG.
At the same time, the results of the present research contradict the data [43] demonstrating a decrease in EEG power in the theta band in the earlier study when performing a task on overcoming a stereotype of long-term memory, a search for an original ending to well-known proverbs and sayings with the purpose of changing their sense. Firstly, this can be explained through features of the EEG signal processing (in the present research, we have revealed short-term synchronization of slow frequencies of EEG) and by differences in the tasks designs.
The increase in power in the delta EEG frequency band in the frontal and temporal zones of the cortex during the solving of mental tasks by healthy subjects can be considered a motivational component of the task that has to be solved [40], or can arise from attention focus and its activation in relation to internal processes, braking of irrelevant information [44], or the comparison of signals and decision-making [45]. At the same time, an increase in synchronization in the theta frequencies could be treated as an indicator of the active involvement of the corresponding cortex zones in the maintenance of mental problem-solving with different complexity levels and the demands of memory load, concentration, and emotional reactions. An increase in theta activity can characterize cortex activation level. The research of Pizagalli and colleagues revealed a positive correlation between the current density of the theta activity generator in the inferior part of the cingula gyrus BA24/32 and the metabolism of glucose [46] that characterizes the cortex activation level. After that, the phenomenon of synchronization itself is considered in the involvement of a large number of neural populations in the realization of the studied activity [47].
In our investigation, synchronization of EEG in the range of theta and delta frequencies was observed for both cases (finding of trivial and original decisions); at the same time, the search for remote associations seemed to be carried out with the higher level of activation of a number of cortex zones than for trivial associations.
According to the expected schema of the search for definitions in both cases (finding creative and trivial definitions), subjects needed to carry out a lexical analysis of words [48], to concentrate on a problem and, as the result of a search for remote associative links between concepts, to choose a decision which was internally considered to be original. As in both cases it was necessary for subjects to carry out a lexical analysis of the words and to concentrate on the problem, the quality of the found solution could presumably be affected (influenced) by the depth of the associative search – or, in other words, mental memory load during the search for associative definitions of the concepts. Also, it is impossible to exclude the condition of the instruction to “be original” from the subjects’ subjective evaluation of their own decisions that was presumably happening before pressing the button.
As the mentioned processes are tightly bound with the operation of verbal information, detected differences in the frontal and temporal cortex zones in the left hemisphere of the brain are quite explainable. Presumably, the increase in power in the delta and theta frequency bands in these zones in earlier time intervals can be connected with the processes underlying verbal associative search (memory loading) when finding the remote characteristics of the presented concepts, whereas the later stages – around 1000 ms before pressing – reflect the motivational and evaluation components connected with the search and choice evaluation [49, 50] of the original decision.
The results are indirectly comparable with those achieved in an earlier PET research [51], where an increase in the regional blood flow in the cortex in the left hemisphere of the brain was shown (BA 47) when there was a need to be original in the condition of verbal creative task performance in comparison with performance of a non-creative task.
At the same time, higher values for event-related absolute power in the fronto-central (Fz) site while finding original decisions in the range of frequencies 2.5–4.2 Hz, with the central frequency at 3 Hz, seem to be connected with the need to preserve internal attention and eliminate unnecessary information [44] for a successful search (finding an original decision) in the wide associative field of concepts. This assumption corresponds with previous results demonstrating a decrease in EEG spectral power in the beta2 frequency band in the left frontal areas in searching for the original definition of a word using another from a remote associative field [52].
CONCLUSIONS
By means of an ERS/ERD method of EEG data analysis, EEG correlates of finding verbal creative decisions were investigated, focused on a search for remote associations in comparison with making trivial decisions focused on the definition of external characteristics of the phenomena and objects. We revealed an increase in event-related synchronization in the delta and theta frequency bands in the frontal and temporal zones of the left hemisphere of the brain and in the front and central cortex areas while making creative decisions. We assume that observed differences in the early stages, before the subject made a decision, can be connected with deeper loading of the memory and the search for definitions in the wide associative field, and can reflect the choice and assessment of originality of the thought-up definition in the later stage.
Список литературы
Guilford J.P. The nature of human intelligence. N.Y. Mc-Graw Hill. 1967.
Guilford J.P. Creativity. Am. Psychol. 5: 444–454. 1950.
Guilford J.P, Wilson R.C., Christensen P.R. A factor-analytic study of creative thinking: II. Administration of tests and analysis of results. Rep. Psychol. Lab. No8: Univer. Southern Calif. 1952.
Torrance E.P. Guiding creative talent. Englewood Cliffs. N.J. US. Prentice-Hall Inc. 1962.
Mednick S.A. The associative basis of the creative process. Psychol. Rev. 69: 220–232. 1962.
Grabner R.H., Fink A., Neubauer A.C. Brain correlates of self-rated originality of ideas: evidence from event-related power and phase-locking changes in the EEG. Behav. Neurosci. 121: 224–230. 2007.
Bechtereva N.P. The Usefulness of Psychophysiology in Intellectual Life. Honorary Lecture. 14th World Congress of Psychophysiology. The Olympics the Brain. September 8–13, 2008. St. Petersburg. Russia. St. Petersburg. IHB RAS. 2008.
Bechtereva N.P. The Magic of Creativity and Psychophysiology. Considerations, thoughts, hypotheses. St. Petersburg. IHB RAS. 2006.
Sternberg R.J., O’Hara L.A. Creativity and intelligence. pp. 251–273. In: Sternberg R.J. (Ed). Handbook of Creativity. Cambridge Univer. Press. 1999.
Arden R., Chavez R.S., Grazioplene R., Jung R.E. Neuroimaging creativity: a psychometric view. Behav. Brain Res. 214: 143–156. 2010.
Dietrich A., Kanso R.A. Review of EEG, ERP, and Neuroimaging Studies of Creativity and Insight. Psychol. Bull. 136: 822–848. 2010.
Beaty R.E., Benedek M., Silvia P.J., Schacter D.L. Creative Cognition and Brain Network Dynamics. Trends Cogn. Sci. 20: 87–95. 2016.
Nagornova Zh.V., Shemiakina N.V. Differences of activations in visual and associative zones during figurative and verbal activity. Ross. Fiziol. Zh. Im. I. M. Sechenova. 100: 418–432. 2014. (In Russ).
Pfurtsheller G., Aranibar A. Event-related cortical desynchronization detected by power measurement of scalp EEG. Electroencephalogr. Clin. Neurophysiol. 42: 817–826. 1977.
Kiebel S., Tallon-Baudry C., Friston K.J. Parametric analysis of oscillatory activity as measured with EEG/MEG. Hum. Brain Mapp. 26: 170–177. 2005.
Oldfield R.G. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia. 9: 97–113. 1971.
Shemyakina N.V., Nagornova Zh.V. Comparative research of perception couples of nouns from the nearby and remote semantic fields by visual ERP approach. Materials of the conference “High Technologies, Basic and Applied Researches in Physiology and Medicine” Vol. IV. St. Petersburg. November 23–26. 2010.
Lemos M.S., Fisch B.J. The weighted average reference montage. Electroencephalogr. Clin. Neurophysiol. 79: 361–370. 1991.
Vigário R.N. Extraction of ocular artifacts from EEG using independent component analysis. Electroencephalogr. Clin. Neurophysiol. 103: 395–404. 1997.
Jung T.P., Makeig S., Humphries C., Lee T.W., McKeown M.J., Iragui V., Sejnowski T.J. Removing electroencephalographic artifacts by blind source separation. Psychophysiology. 37: 163–178. 2000.
Tereshchenko E.P., Ponomarev V.A., Kropotov Yu.D., Müller A. Comparative efficiencies of different methods for removing blink artifacts in analyzing quantitative electroencephalogram and event-related potentials. Human Physiology. 35: 241–247. 2009.
Tallon-Baudry C., Bertrand O. Oscillatory gamma activity in humans and its role in object representation. Trends Cogn. Sci. 3: 151–162. 1999.
Kalcher J., Pfurtsheller G. Discrimination between phase-locked and non-phase-locked event-related EEG activity. Electroencephalogr. Clin. Neurophysiol. 94: 381–384. 1995.
Meyer L., Grigutsch M., Schmuck N., Gaston P., Friederici A.D. Frontal-posterior theta oscillations reflect memory retrieval during sentence comprehension. Cortex. 71: 205–218. 2015.
Pavlov Y.G., Kotchoubey B. EEG correlates of working memory performance in females. BMC Neurosci.18: 26. 2017.
Yang C.Y., Huang C.K. Working-memory evaluation based on EEG signals during n-back tasks. J. Integr. Neurosci. 17(3–4): 695–707. 2018.
Razumnikova O.M. Creativity related cortex activity in the remotes associates task. Brain Res. Bull. 73: 96–102. 2007.
Wilson R.C., Guilford J.P., Christensen P.R. The measurement of individual differences in originality. Psychol. Bull. 50: 362–370. 1953.
Jung-Beeman M., Bowden E.M., Haberman J., Frymiare J.L., Arambel-Liu S., Greenblatt R., Reber P.J., Kounios J. Neural activity when people solve verbal problems with insight. PLoS Biol. 2(4): E97. 2004.
Başar E., Demiralp T., Schürmann M., Başar-Eroglu C., Ademoglu A. Oscillatory Brain Dynamics, Wavelet Analysis and Cognition. Brain Lang. 66: 146–183. 1999.
Basar E., Basar-Eroglu C. Karakas S., Schurmann M. Brain oscillations in perception and memory. Int. J. Psychophysiol. 35: 95–124. 2000.
Libet B., Gleason C.A., Wright E.W., Pearl D.K. Time of conscious intention to act in relation to onset of cerebral activity (readiness-potential). The unconscious initiation of a freely voluntary act. Brain. 106(Pt 3): 623–642. 1983.
Keller I., Heckhausen H. Readiness potentials preceding spontaneous motor acts: voluntary vs. involuntary control. Electroencephalogr. Clin. Neurophysiol. 76: 351–361. 1990.
Pfurtscheller G., Aranibar A. Evaluation of event-related desynchronization (ERD) preceding and following voluntary self-paced movement. Electroencephalogr. Clin. Neurophysiol 46: 138–146. 1979.
Leocani L., Toro C., Manganotti P., Zhuang P., Hallett M. Event-related coherence and event-related desynchronization/synchro-nization in the 10 Hz and 20 Hz EEG during self-paced movements. Electroencephalogr. Clin. Neurophysiol. 104: 199–206. 1997.
Deecke L., Grözinger B., Kornhuber H.H. Voluntary finger movement in man: cerebral potentials and theory. Biol. Cybern. 23: 99–119. 1976.
Urbano A., Babiloni C., Onorati P., Babiloni F. Dynamic functional coupling of high resolution EEG potentials related to unilateral internally triggered one-digit movements. Electroencephalogr. Clin. Neurophysiol. 106: 477–487. 1998.
Womelsdorf T., Vinck M., Leung L.S., Everling S. Selective theta-synchronization of choice-relevant information subserves goal-directed behavior. Front Hum. Neurosci 4: 210. 2010.
Nyhus E., Curran T. Functional role of gamma and theta oscillations in episodic memory. Neurosci. Biobehav. Rev. 34: 1023–1035. 2010.
Knyazev G.G. EEG delta oscillations as a correlate of basic homeostatic and motivational processes. Neurosci. Biobehav. Rev. 36: 677–695. 2012.
Danko S.G., Starchenko M.G., Bechtereva N.P. EEG Local and Spatial Synchronization during a Test on the Insight Strategy of Solving Creative Verbal Tasks. Human Physiology. 29: 502–504. 2003.
Whitton J.L., Moldofsky H., Lue F. EEG frequency patterns associated with hallucinations in schizophrenia and “creativity” in normal. Biol. Psychiatry. 13: 123–133. 1978.
Shemyakina N.V., Danko S.G., Nagornova Zh.V., Starchenko M.G., Bechtereva N.P. Changes in the power and coherence spectra of the EEG rhythmic components during solution of a verbal creative task of overcoming a stereotype. Human Physiology. 33: 524–530. 2007.
Harmony T., Fernández T., Silva J., Bernal J., Díaz-Comas L., Reyes A., Marosi E., Rodríguez M., Rodríguez M. EEG delta activity: an indicator of attention to internal processing during performance of mental tasks. Int. J. Psychophysiol. 24: 161–171. 1996.
Başar-Eroglu C., Başar E., Demiralp T., Schürmann M. P300-response: possible psychophysiological correlates in delta and theta frequency channels. Int. J. Psychophysiol. 13: 161–179. 1992.
Pizzagalli D.A., Oakes T.R., Davidson R.J. Coupling of theta activity and glucose metabolism in the human rostral anterior cingulate cortex: an EEG/PET study of normal and depressed subjects. Psychophysiology. 40: 939–949. 2003.
Klimesch W. EEG alpha rhythms and memory processes. Int. J. Psychophysiol. 26: 319–340. 1997.
Bastiaansen M.C., van der Linden M., Ter Keurs M., Dijkstra T., Hagoort P. Theta responses are involved in lexical-semantic retrieval during language processing. J. Cogn. Neurosci. 17: 530–541. 2005.
Thompson-Schill S.L., Esposito M.D., Aguirre G.K., Farah M.J. Role of left inferior prefrontal cortex in retrieval of semantic knowledge: a reevaluation. Proc. Natl. Acad. Sci. USA. 94: 14792–14797. 1997.
Kan I.P., Thompson-Schill S.L. Selection from perceptual and conceptual representations. Cogn. Affect. Behav. Neurosci. 4: 466–482. 2004.
Shemyakina N.V. PET study of verbal creativity in different evolved emotional conditions. Book of abstracts International C.I.A.N.S conference. Bratislava. Slovakia. 72. 2005.
Shemyakina N.V., Dan’ko S.G. Changes in the power and coherence of the β2 EEG band in subjects performing creative tasks using emotionally significant and emotionally neutral words. Human Physiology. 33: 20–26. 2007.
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова