

Сенсорные системы, 2019, T. 33, № 1, стр. 70-76
Отражение активности сердца в электроэнцефалограмме кошек в периоды медленного сна
В. Д. Лаврова 1, *, И. И. Бусыгина 2, И. Н. Пигарев 1
1 Институт проблем передачи информации им. А.А. Харкевича РАН (ИППИ РАН)
127051 Москва, Большой Каретный переулок, д.19, Россия
2 Институт физиологии им. И.П. Павлова РАН
199034 Санкт-Петербург, наб. Макарова, д. 6, Россия
* E-mail: pilotfish@bk.ru
Поступила в редакцию 16.10.2018
Аннотация
В хронических экспериментах на двух кошках регистрировали интракраниальную электроэнцефалограмму (ЭЭГ) от двух пар электродов, расположенных над лобно-теменными и затылочными областями мозга. Кроме того, проводили регистрацию электрокардиограммы (ЭКГ) и движений глаз. Было показано, что в фазу медленного сна на ЭЭГ, регистрируемой над лобно-теменными участками коры, усреднялись три типа волн, синхронных с сокращениями сердца. Первый тип волн представлял собой результат прямого распространения электрического тока от сердца. Второй тип имел вид медленных волн с ритмом сердечных сокращений и скорее всего отражал механические смещения, связанные с пульсацией крови. Эти два типа волн проявлялись нерегулярно, как во сне, так и в бодрствовании, и могли быть видны в отведениях от обоих отделов коры. Третий тип волн имел вид типичного коркового вызванного ответа, появляющегося с задержкой порядка 80 мс после зубца R в ЭКГ, и регистрировался преимущественно над лобно-затылочными отделами коры и только в состоянии медленного сна. Это наблюдение поддерживает наше представление, согласно которому в период сна кора мозга вовлекается в анализ состояния висцеральной сферы организма.
ВВЕДЕНИЕ
Вторая половина двадцатого столетия и начало века двадцать первого были связаны с интенсивным изучением экстероцептивных сенсорных систем – и в первую очередь с изучением приема, передачи и обработки зрительной информации. По сравнению с количеством этих исследований очень скромную долю составляли работы, посвященные изучению интерорецепции, передачи и обработки сигналов, поступающих от внутренних органов. Это, безусловно, имело свои причины, к рассмотрению которых мы вернемся в обсуждении.
В последнее время ситуация начала постепенно меняться. Этой тенденции, в частности, способствуют появление и развитие висцеральной теории сна (Пигарев, 2013). Суть этой теории состоит в предположении, что все сенсорные зоны коры мозга, обрабатывающие в бодрствовании информацию о состоянии окружающей среды, а также проприоцептивную информацию о положении и движениях собственного тела, переключаются на обработку интероцептивной информации от внутренних органов тела во время сна. В рамках этой теории вся кора мозга во время сна становится «висцеральной», и обработка информации, поступающей от внутренних органов, направлена на диагностику их состояния и на разработку программ восстановления их работоспособности. Специфическая картина электрической активности коры мозга во время сна отражает характер информационных потоков, идущих от разных висцеральных систем организма. Эта кажущаяся на первый взгляд достаточно фантастическая гипотеза нашла подтверждение в большом количестве специально поставленных экспериментов на кошках (Pigarev, 1994), кроликах (Pigarev et al., 2004) и обезьянах (Pigarev et al., 2008).
Было показано, что во время медленного сна нейроны первичной зрительной коры действительно начинают отвечать на электрическую или магнитную стимуляцию желудка и тонкого кишечника (Pigarev, 1994; Pigarev et al., 2008). Далее было показано, что и естественная миоэлектрическая активность этих органов отражается в электрической активности зрительных зон коры в медленном сне (Pigarev et al., 2013). И, наконец, было продемонстрировано, что на статистических характеристиках импульсной активности нейронов коры мозга кошек отражается смена внутрижелудочной среды, проводимая в период медленного сна (Пигарев и др., 2014). Все эти эффекты исчезали сразу после пробуждения животных.
Столь нетривиальные результаты экспериментов, выполненных на органах желудочно-кишечного тракта, позволяли думать, что и сигналы от других висцеральных систем должны поступать в кору мозга во время сна. Настоящая работа представляет результаты первого этапа исследования отражения работы сердечно-сосудистой системы в электрической активности коры мозга кошек в цикле сон – бодрствование. На этом этапе мы исследовали изменения электроэнцефалограммы (ЭЭГ) лобно-теменных и затылочных зон коры мозга кошек, происходящие синхронно с сокращениями сердца в разные фазы цикла сон – бодрствование.
ОПИСАНИЕ МЕТОДИКИ
Эксперименты проводили на двух взрослых кошках с использованием модифицированного метода безболезненной фиксации головы (Noda et al., 1971). Элементы модификации метода даны в предшествующей статье (Pigarev et al., 2009).
ПОДГОТОВКА ЖИВОТНОГО К ЭКСПЕРИМЕНТАМ
Операции выполняли под глубоким общим наркозом. После премедикации ксилазином (0.15 мл на кг) вводили золетил в концентрации 20 мг/мл внутримышечно. Доза первой инъекции была 0.3 мл на 1 кг веса животного. Далее по ходу операции золетил вводили дозами не более 0.5 мл по мере надобности.
Для операции кошку помещали в стереотаксическую установку. Мягкие ткани удаляли с дорзальной поверхности головы. В череп на расстоянии 15 мм справа и слева от центральной оси вкручивали восемь винтиков из нержавеющей стали. Винтики охватывали проволочной рамкой. Всю конструкцию рамки заполняли самозатвердевающей стоматологической акриловой пластмассой. Тонким слоем той же пластмассы заливали и поверхность черепа внутри рамки. Такая конструкция предоставляла свободный доступ к большей части дорсальной поверхности черепа, открывая легкий доступ для погружения электродов в большинство областей головного мозга. Кроме того, во время опыта рамка служила для безболезненной фиксации головы животного в станке.
ТЕХНИКА РЕГИСТРАЦИИ ИНТРАКРАНИАЛЬНОЙ ЭЛЕКТРОЭНЦЕФАЛОГРАММЫ (ЭЭГ)
Макроэлектроды для регистрации электроэнцефалограммы погружали до поверхности твердой мозговой оболочки через конические пластиковые направляющие трубочки, установленные в просверленные отверстия в черепе и прикрепленные к рамке (Pigarev et al., 2009). Электроды делали из медицинской нержавеющей стали (Elgiloy) толщиной 0.5 мм в корпусах из самоотвердевающей стоматологической пластмассы.
Одну пару электродов располагали в лобно-теменном отделе, в правом и левом полушариях, симметрично относительно центральной оси, в точки с координатами АР +19.5 и латеральностью 7.5 мм. Аналогичную пару электродов помещали симметрично в затылочном отделе, над зрительной зоной V1 (поле 17) в точки с координатами АР (–5) и латеральностью 2 мм. Выбор мест расположения электродов в передних отделах мозга подробно мотивирован ниже, в обсуждении.
В большинстве экспериментов эти пары электродов использовали в режиме биполярного отведения ЭЭГ от двух симметричных точек правого и левого полушарий, от лобно-теменной и затылочной областей мозга. В одном эксперименте проводили отведения отдельно от правого и левого полушарий передних отделов коры. При этом референтный электрод располагали в затылочной части черепа.
РЕГИСТРАЦИЯ ЭЛЕКТРОКАРДИОГРАММЫ (ЭКГ) И ДРУГИХ ФИЗИОЛОГИЧЕСКИХ ПАРАМЕТРОВ
Эту работу проводили в длительных экспериментах на животных, задействованных ранее в изучении работы органов желудочно-кишечного тракта (Пигарев и др., 2014). Для этой цели им были имплантированы хронические фистулы желудка. Мы воспользовались этим обстоятельством и для регистрации ЭКГ помещали один электрод через фистулу непосредственно в полость желудка. Вторым электродом служил винтик, ввернутый в кость черепа в теменной области около медиального шва и используемый также для заземления животного в процессе эксперимента.
Движения глаз и век животного регистрировали инфракрасным окулометром, дающим качественную картину этих движений, достаточную для идентификации фаз сна.
Во всех каналах регистрации использовали фильтры с полосой пропускания от 0.3 до 200 Гц. Использовали усилители DL304PN компании NeuroBioLab. Выходные сигналы подавали в систему Power Lab (ADInstruments, Австралия) для записи на жесткий диск с частотой оцифровки 1 кГц.
На протяжении всего эксперимента синхронно с физиологическими параметрами проводили видеозапись поведения животного.
Все процедуры, связанные с содержанием животных и проведением экспериментов, соответствовали международным нормам по обращению с лабораторными животными «Guidelines for the Care and Use of Animals» (Zimmermann, 1987), и Хельсинкской Декларации по этическим принципам для медицинских исследований.
АНАЛИЗ ДАННЫХ
Обработку и статистическую оценку результатов проводили в режиме off-line в программе Spike2 ver.6.16 (CED, Великобритания) с использованием скриптов, написанных непосредственно для целей этой работы.
Первым этапом обработки всех записей было автоматическое выделение моментов времени, соответствующих появлению максимума зубца R на ЭКГ. Для этого при помощи алгоритма, используемого программой Spike2 для сортировки импульсов по форме, в канале записи ЭКГ проводили выделение комплексов PQRST, отражающих отдельное сердечное сокращение, с последующим маркированием моментов появления максимумов зубцов R. Маркеры, отмечающие появления максимумов зубцов R, далее использовали как триггеры для усреднения ЭЭГ и ЭКГ.
На следующем этапе проводили параллельный анализ полисомнограмм и видеозаписи для нахождения фрагментов медленного сна и бодрствования. Задача этого этапа состояла в том, чтобы выделить достаточно длительные участки медленноволнового сна и бодрствования для их последующего использования для усреднения ЭЭГ и поиска отражения работы сердца в этой активности.
Далее проводили усреднение (averaging) сигналов в канале ЭКГ отдельно на участках медленного сна и бодрствования, и в двух каналах записи электроэнцефалограммы (лобно-теменного и затылочного) с использованием маркеров моментов зубца R как триггеров для усреднения.
На заключительном этапе проводили статистическую оценку достоверности отличия усредненных волн ЭЭГ (по предположению отражающих активность сердца) от случайных отклонений.
Все проведенные усреднения мы всегда представляли как среднее, заключенное между линиями, соответствующими двум стандартным ошибкам (SEM) в ту и другую сторону (рис. 1–4). При таком представлении для большинства процессов 95% результатов заключено между этими двумя линиями. Если средние величины двух сравниваемых точек на кривой отличались между собой меньше, чем на четыре стандартных ошибки, эти точки считали неотличимыми. Таким образом, средняя величина максимума волны, по предположению претендующей на вызванный ответ, должна была отличаться от средней величины одного из ближайших минимумов более, чем на четыре стандартных ошибки (что было примерно эквивалентно р < 0.05). Если это различие было меньше (т.е. при р > 0.05), то волну исключали из дальнейшего анализа. При различиях, превышающих четыре стандартных ошибки, волна переходила на второй, более точный, этап статистической оценки с применением метода стохастического моделирования. Если на исследуемом фрагменте записи усреднение проводили по N сердечным сокращениям, то на том же участке записи программа генерировала N случайно распределенных маркеров. Теперь аналогичное усреднение проводили с использованием этих случайных маркеров как триггеров и фиксировали величину максимальных полученных отклонений. Такую процедуру проводили 500 раз для разных наборов случайно распределенных маркеров. В результате получали распределение величин случайно возможных максимальных отклонений усредненных кривых на данном участке записи для нашего числа N. По кривой этого распределения оценивали вероятность получения случайной волны, по предположению представляющей вызванный ответ в нашем конкретном эксперименте. Преимущество этого метода состоит в том, что оценка делается для каждого конкретного эксперимента и не зависит от характера распределения исходных данных. В работе был принят критерий достоверности p ≤ 0.05.
Рис. 1.
Усредняемое на канале ЭЭГ отклонение первого типа – результат прямого затекания тока от сердца.
1 – ЭКГ; 2 –ЭЭГ, регистрируемая от лобно-теменных отделов коры мозга. Число усреднений – 1186.
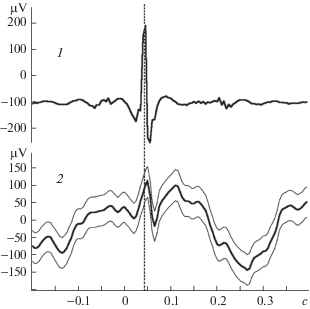
Рис. 2.
Усредняемое на каналах ЭЭГ отклонение второго типа – результат пульсовой волны.
1 – ЭКГ; 2 –ЭЭГ, регистрируемая от затылочных отделов коры мозга; 3 – ЭЭГ, регистрируемая от лобно-теменных отделов коры мозга. Число усреднений – 1310.
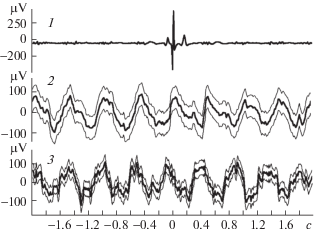
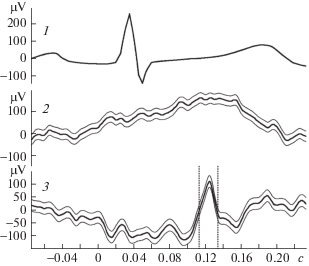
Рис. 3.
Предполагаемый вызванный ответ, отражающий активность нейронов, связанную с сердечными сокращениями (канал 3 выделен двумя вертикальными пунктирными линиями). Обозначения те же, что и на рис. 2. Число усреднений – 1997.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
В исследовании интракраниальной ЭЭГ были проанализированы результаты 12 экспериментов длительностью от 4 до 6 ч, включающие участки бодрствования и медленного сна.
В большинстве наших записей при усреднении сигналов ЭЭГ по моментам максимумов зубцов R электрокардиограммы мы получали достоверные отклонения, синхронные с сердечными сокращениями. Эти отклонения можно было разбить на три типа.
К первому типу относили короткие высокочастотные отклонения, повторяющие в значительной степени форму ЭКГ и появляющиеся одновременно с этими зубцами. Пример такого ответа приведен на рис. 1.
Обращает на себя внимание отсутствие задержки во времени между пиками в усредненной ЭКГ и усредненной ЭЭГ (на рис. 1 показано вертикальной пунктирной линией). Отсутствие латентности определенно свидетельствовало о том, что эти отклонения являлись следствием прямого распространения электрического тока от сердца. Это так называемые артефакты ЭКГ в электроэнцефалограмме. Их величина определяется как внутренним распределением электрических сопротивлений в структурах тела и мозга, так и расположением регистрирующих электродов. В наших опытах регистрирующие электроды располагались симметрично над правым и левым полушариями мозга. Такое положение давало ряд существенных преимуществ для оценки возможной локализации источника сигнала, но способствовало проявлению таких артефактов. Эти артефакты были видны на многих записях и могли проявляться как во сне, так и в бодрствовании. Отклонения этого типа, хорошо известные в электроэнцефалографии, не представляли интереса для нашего исследования.
Второй тип активности представлял собой медленные колебания, напоминающие синусоиды, идущие с частотой сердечных сокращений (рис. 2). Эти отклонения можно было наблюдать в отведениях как от лобно-теменной, так и от затылочной коры во сне и бодрствовании. В отдельных опытах они могли быть видны хорошо, а в других могли полностью отсутствовать. Мы полагаем, что эти волны связаны с пульсирующим кровотоком в зоне расположения отводящих электродов. Их природа может быть связана как с чисто механическими микросдвигами ткани в зоне кончиков электродов, так и с периодическими изменениями концентрации кислорода в результате пульсирующего тока крови в сосудах черепа или твердой мозговой оболочки. Природу этих отклонений мы также не стремились исследовать в нашей работе.
Для нас основной интерес представляли отклонения третьего типа. Это были высокочастотные отклонения, по форме напоминающие типичные корковые вызванные ответы на сенсорную стимуляцию. Они появлялись с заметным латентным периодом после момента сокращения сердца. Пример этого типа ответа приведен на рис. 3, на канале регистрации активности в лобно-теменном отделе (канал 3). Положительный компонент этого ответа на рисунке заключен между двумя вертикальными пунктирными линиями. На канале регистрации из затылочных отделов на этой же записи (канал 2) хорошо видна волна, представляющая второй тип активности.
Латентный период отклонений третьего типа, измеренный между максимумом зубца R и максимумом основной волны вызванного ответа (пиковая латентность), в разных опытах находился в пределах от 40 до 120 мс. Величина его хорошо согласуется с вероятным временем проведения нервного сигнала от сердца до исследуемой зоны коры мозга.
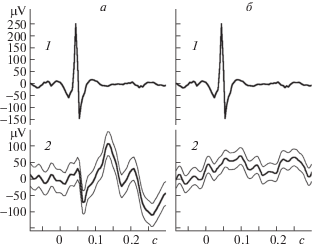
Проверяемая нами гипотеза предполагала, что функциональная связь активности сердца и активности корковых зон коры будет устанавливаться в периоды сна и должна существенно уменьшаться или полностью исчезать в периоды бодрствования. Можно было ожидать, что представляющие для нас интерес отклонения третьего типа будут появляться при усреднении исследуемых сигналов в периоды сна, и будут исчезать в периоды бодрствования. Проведение таких раздельных усреднений действительно подтвердило наше ожидание. На рис. 4 приведен пример ЭЭГ лобно-теменной зоны, усредненной по сердечным маркерам в период медленного сна (а) и в период активного бодрствования (б). Видно, что во время бодрствования явно проявляется лишь фрагмент волны второго типа, тогда как в период сна на эту волну явно накладывается высокочастотный ответ третьего типа. В записях, вошедших в данную работу, было проанализировано 29 фрагментов медленноволнового сна и 15 фрагментов бодрствования. Вызванные ответы, связанные с сердечными сокращениями, были обнаружены в 26 из этих участков сна, и ни в одном фрагменте бодрствования. Форма ответов в разных записях могла быть различной.
При сравнении усредняемых потенциалов в лобно-теменных и затылочной областях коры было выявлено, что артефакты прямого распространения тока (волны первого типа) сильнее проявлялись в затылочных отведениях, поскольку затылочные электроды находились ближе к источнику этих токов.
Волны второго типа (предположительно являющиеся следствием пульсовой активности) наблюдались одинаково хорошо как в передних, так и в задних областях. Однако в затылочных отделах их амплитуда была обычно выше приблизительно в 2–4 раза, вероятно, из-за близости венозного синуса в районе медиального шва, вблизи от расположения наших регистрирующих электродов.
Нас интересовали волны третьего типа, скорее всего отражающие активацию нейронов коры, вызванную нервными сигналами, приходящими от сердца. Они проявлялись преимущественно в отведениях между парами электродов, расположенными над лобно-теменными зонами мозга, в правом и левом полушариях. В них были отмечены 20 из полученных вызванных ответов, в то время как от электродов в затылочной коре было зарегистрировано только шесть ответов и при этом менее выраженных.
Корковые ответы в передних отделах мозга, синхронные с сокращениями сердца кошек, впервые были описаны довольно давно в работах лаборатории академика В.Н. Черниговского, в Институте физиологии им. И.П. Павлова в Ленинграде в шестидесятые годы двадцатого века (Черниговский, 1960; Мусящикова, Черниговский, 1973). Эти исследования проводили в острых опытах на кошках под глубоким наркозом. Ответы были настолько сильными, что их обнаруживали в отдельных реализациях еще до появления метода компьютерного усреднения. Более того, в исследованиях тех авторов были описаны и обратные влияния электрической стимуляции этих корковых зон, приводящие к изменению характера работы сердца. Точки коры, вызывающие эти эффекты, были названы фокусами сердечной активности. Фокусы сердечной активности представляли на топографической карте борозд и извилин мозга. В наших экспериментах мы старались помещать фронтальные регистрирующие электроды в аналогичные по анатомической структуре точки коры. В тех же исследованиях отмечалось, что менее выраженные ответы реже можно было наблюдать и за пределами первичных фокусов активности. Тогда эти зоны, по аналогии с терминологией, принятой для экстероцептивных сенсорных зон, назвали вторичными и третичными интероцептивными зонами. Эти зоны покрывали практически всю поверхность коры мозга.
В исследованиях лаборатории В.Н. Черниговского были описаны корковые фокусы активности и для других висцеральных систем. Авторы этих исследований неоднократно обращали внимание на достаточно удивительное для них явление. Корковые ответы на висцеральную стимуляцию ярко проявлялись на фоне глубокого наркоза и заметно уменьшались при снижении его глубины. Рационального объяснения этого наблюдения тогда получено не было. Более того, позже, при попытках разных лабораторий воспроизвести эти результаты на бодрствующих животных в хронических опытах без наркоза корковые ответы на висцеральную стимуляцию обнаружить не удавалось. Поскольку отрицательные результаты публиковать было не принято, информация о таких экспериментах доходила лишь по каналам личного общения. Однако это привело к тому, что корковые ответы на висцеральные раздражения и сам факт существования корковых висцеральных представительств стал считаться вероятным артефактом наркоза.
Теперь же становится понятно, что такие представительства реально существуют и проявляются у поведенчески активных животных без всякого наркоза, но только в состоянии сна. Видимо, применяемый наркоз приводил к изменениям эффективности связей между разными структурами мозга, частично сходными с таковыми в период естественного сна. В наших экспериментах корковые вызванные ответы на сердечные сокращения также проявлялись во время сна в лобно-теменных отведениях, над зонами коры, обозначенными ранее как фокусы сердечной активности. Это стало еще одним свидетельством в пользу того, что огромный экспериментальный материал, собранный в лаборатории В.Н. Черниговского, отражает не только действительно существующие физиологические явления, но и явления, чрезвычайно важные для понимания процесса сна. Полученные тогда данные могут быть использованы при планировании новых исследований вовлечения сенсорных зон коры мозга в период сна в обработку сигналов, поступающих от интерорецепторов разных висцеральных систем организма.
ЗАКЛЮЧЕНИЕ
В ходе представленного исследования было показано, что информация о работе сердечно-сосудистой системы в периоды медленного сна действительно достигает уровня лобно-теменных отделов коры головного мозга, и эта передача информации прерывается во время бодрствования. Данное наблюдение поддерживает развиваемую нами висцеральную теорию сна.
Авторы благодарят Г.О. Фёдорова за написание скриптов для программы Spike2, используемых для анализа полученных результатов.
Работа частично поддержана грантом РФФИ 16-04-00413.
Список литературы
Мусящикова С.С., Черниговский В.Н. Кортикальное и субкортикальное представительство висцеральных систем. Л.: Наука, 1973. 288 с.
Пигарев И.Н. Висцеральная теория сна. Журнал высшей нервной деятельности им. И.П. Павлова. 2013. Т. 63. № 1. С. 86–104.
Пигарев И.Н., Бибиков Н.Г., Бусыгина И.И. Изменения внутрижелудочной среды во время сна влияют на статистические характеристики нейронной активности коры мозга. Российский физиологический журнал им. И.М. Сеченова. 2014. Т. 100. № 6. С. 722–735.
Черниговский В.Н. Интерорецепторы. М.: Медгиз, 1960. 659 с.
Noda H., Freeman R.B., Gies B., Creutzfeldt O.D. Neural responses in the visual cortex of awake cats to stationary and moving targets. Experimental brain research. 1971. V. 12. P. 389–405.
Pigarev I.N. Almirall H., Marimon J., Pigareva M.L. Dynamic pattern of the viscero-cortical projections during sleep. Study in New Zealand rabbits. Journal of Sleep Research. 2004. V. 13. Suppl. 1. P. 574.
Pigarev I.N. Neurons of visual cortex respond to visceral stimulation during slow wave sleep. Neuroscience. 1994. V. 62. № 4. P. 1237–1243.
Pigarev I.N., Almirall H., Pigareva M.L. Cortical evoked responses to magnetic stimulation of macaque’s abdominal wall in sleep-wake cycle. Acta Neurobiol Exp. 2008. V. 68. P. 91–96.
Pigarev I.N., Bagaev V.A., Levichkina E.V., Fedorov G.O., Busigina I.I. Cortical visual areas process intestinal information during slow-wave sleep. Neurogastroenterology and motility. 2013. V. 25. P. 268–275.
Pigarev I.N., Saalmann Y.B., Vidyasagar T.R. A minimally invasive and reversible system for chronic recordings from multiple brain sites in macaque monkeys. Journal of neuroscience methods. 2009. V. 181. № 2. P. 151–158.
Zimmermann M. Ethical Principles for maintenance and use of animals in neuroscience research. Neuroscience Letters. 1987. V. 73. P. 1.
Дополнительные материалы отсутствуют.
Инструменты
Сенсорные системы