

Сенсорные системы, 2020, T. 34, № 1, стр. 19-24
Тонкая структура рецептивных полей ориентационно-избирательных ганглиозных клеток сетчатки рыб
А. Т. Алипер 1, *, И. Дамянович 1, А. А. Зайчикова 1, Е. М. Максимова 1, П. В. Максимов 1
1 Институт проблем передачи информации им. А. А. Харкевича РАН
127051 Москва, Большой Каретный переулок, д.19, Россия
* E-mail: outtaget@gmail.com
Поступила в редакцию 09.09.2019
После доработки 24.10.2019
Принята к публикации 06.11.2019
Аннотация
Ориентационно-избирательные ганглиозные клетки (ОИ ГК) найдены в сетчатке рыб несколько десятилетий назад, но механизмы ориентационной избирательности остаются до сих пор неизученными. ОИ ГК у рыб можно разделить на два класса, различающихся предпочтительной ориентацией, которая может быть близка к вертикальной либо горизонтальной. Остальными характеристиками эти два класса между собой не различаются. Они не избирательны к знаку контраста, т.е. имеют on-off природу. Мы записывали экстраклеточную активность окончаний аксонов сетчаточных ГК в tectum opticum живой обездвиженной рыбы. Объектом являлся серебряный карась. Параметры стимулов и экспериментальной серии задавались при помощи специально разработанного программного обеспечения. В данном исследовании мы использовали метод “шахматной доски“ со стимуляцией одной точкой и двухточечной стимуляцией. ОИ ГК – детекторы горизонтального и вертикального края – способны давать ответ на одиночные вспышки точек, что позволяет обмерять их возбуждающее рецептивное поле. Ответ на такую стимуляцию заметно слабее в сравнении с таковым на предпочтительный стимул – соответственно ориентированную линию или границу. Однако, когда стимуляция идет одновременно двумя точками, которые выступают аппроксимацией отрезка предпочтительной ориентации, ОИ ГК отвечают длящимся импульсным разрядом. Мы также наблюдали торможение, когда точки были ориентированы ортогонально предпочтительной ориентации. Таким образом, две точки выступают достаточной аппроксимацией предпочтительного или ортогонального направления, что позволяет исследовать локальные свойства рецептивных полей ОИ ГК.
ВВЕДЕНИЕ
Ганглиозные клетки (ГК) осуществляют заключительный этап обработки зрительной информации в сетчатке позвоночных. Вся зрительная сцена закодирована множеством ГК, чьи рецептивные поля (РП) распределены по поверхности сетчатки. Обычно РП ГК функционально разделены на центральную и периферическую части. Предъявление адекватного стимула в центральной части РП вызывает ответ клетки на такую стимуляцию. Если тот же стимул предъявлять только периферической части РП, никакого ответа не будет. Возбуждаемая центральная часть РП (ВРП) соответствует области, охватываемой дендритным древом клетки, которое собирает зрительный сигнал с относительно обширной области. На сетчатке кролика (Yang, Masland, 1992; 1994, Devries, Baylor, 1997) было показано, что ВРП и области, покрываемые дендритными древами ГК, имеют примерно одинаковые размеры.
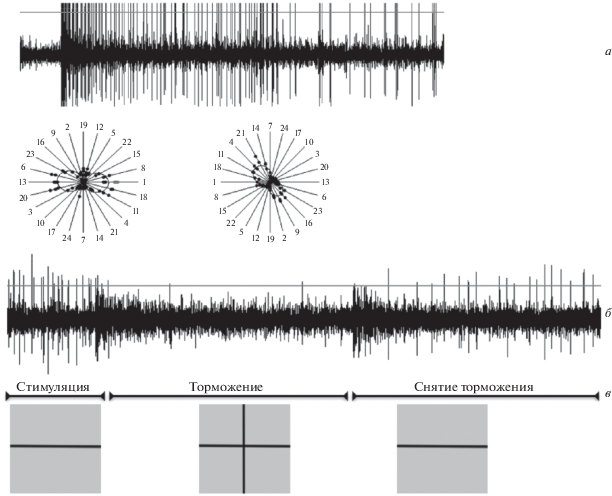
Информация о различных свойствах видимых объектов, их размер, направление движения, форма, цвет и другое, обрабатываются разными типами специализированных ГК – детекторами. Детекторы в том числе описаны в сетчатке рыб (Cronly-Dillon, 1964; Jacobson, Gaze, 1964; Зенкин, Пигарев, 1969; Liege, Galand, 1971; Максимова и др., 1971; Wartzok, Marks, 1973; Kawasaki, Aoki, 1983; Billota, Abramov, 1989). Информация, обработанная специализированными ГК, передается в первичные зрительные центры мозга рыб, в основном в одну из структур среднего мозга – tectum opticum (TO). При вертикальном продвижении электрода через слои TO можно наблюдать ответы от аксональных окончаний различных типов ГК. В поверхностных слоях преимущественно отводятся ответы дирекционально-избирательных ГК (ДИ ГК), несколько глубже наблюдаются окончания разнообразных элементов, таких как ориентационно-избирательные ГК (ОИ ГК), детекторы пятен, цветооппонентные клетки, еще глубже проецируются детекторы контраста с длящимся ответом (спонтанно-активные ГК) (Максимов, 2009). Особый интерес в данный момент прикован к ОИ ГК рыб, поскольку механизм формирования ориентационной избирательности до сих пор не изучен. У рыб представлены два класса ОИ ГК – с предпочтением к горизонтальным и вертикальным стимулами соответственно. Они могут отвечать длящимся ответом на стимуляцию стационарной линией предпочтительной ориентации, который может быть заторможен одновременным введением в РП стимула ортогональной ориентации (рис. 1). На записях импульсной активности линиями показан порог отсечки регистрируемых импульсов.
Рис. 1.
Свойства ОИ ГК рыб.
Ответ ОИ ГК на стимуляцию стационарной линией предпочтительной ориентации (а). Диаграммы направленности детектора вертикального (слева) и горизонтального (справа) краев (б). Торможение ответа на стационарную линию предпочтительной ориентации ортогональным стимулом (в).
Цель данной работы – исследование рецептивных полей ОИ ГК сетчатки рыб, выявление зон торможения и потенциации возбуждения в центральной и периферической зонах РП.
МЕТОДИКА
Объектами для проведения всех опытов служили серебряные караси Carassius auratus gibelio размером от 10 до 15 см и весом от 35 до 100 г из подмосковных прудовых хозяйств. Животные содержались в лабораторных аквариумах объемом 60 л с аэрацией и фильтрацией воды в течение нескольких месяцев при комнатной температуре и естественном световом режиме.
Для доступа к ТО у рыбы с одной стороны черепа, контралатерально относительно глаза, на который подается стимуляция, удаляли теменно-затылочную кость, а также располагающиеся под ней жировую ткань и участок мягкой оболочки мозга. На время опыта животное обездвиживали введением d-тубокурарина внутримышечно (доза составляла 0.3 мг на 100 г массы рыбы). Обездвиженную рыбу закрепляли в естественном положении в аквариуме из плексигласа, с принудительным протоком воды через жабры. В установке в течение опыта циркулировало около 10 л постоянно аэрирующейся и фильтрующейся воды. Циркуляция воды обеспечивалась термостатирующим насосом. К жаберному аппарату рыбы вода поступала под давлением 50 см водяного столба. Уровень воды в аквариуме поддерживали таким образом, чтобы глаза рыбы находились полностью под водой, но вода не заливалась на открытый мозг.
Предъявление программно-сгенерированных стимулов осуществлялось через прозрачную стенку аквариума на экране монитора LG Flatron 775FT, установленного на подвижном столике, что позволяло перемещать его в нужное место поля зрения рыбы. В основном исследовали латеральные поля зрения в довольно широкой области: свыше 60° по горизонтали и около 40° по вертикали. Расстояние от монитора до глаза рыбы в ходе эксперимента составляло 30–40 см. В качестве стимулов в экспериментах, описанных в данной работе, использовались только градации серого. Относительные спектры излучения экрана были измерены при помощи модульной спектрометрической системы MCS 500 Modules от производителя Carl Zeiss. Максимальная яркость экрана (при R = G = B = 255), измеренная фотометром ТКА-04/3, составляла 38 кд/м2. В энергетических единицах это соответствует эффективной энергетической яркости для фотопического зрения человека, равной 56 мВт ст–1 м–2. Согласно нашим данным (Максимов, 2005), фотопическая спектральная чувствительность ганглиозных клеток сетчатки карася определяется в основном его длинноволновыми колбочками и смещена на 75 нм в длинноволновую сторону. Соответствующая функция спектральной чувствительности была рассчитана по формуле Говардовского (Govardovskii et al., 2000) для зрительного пигмента системы витамина А2 с λmax = 622.5 нм с учетом поглощения в передних средах глаза (Douglas, 1989). Рассчитанная таким образом эффективная энергетическая яркость того же белого экрана для ганглиозных клеток сетчатки карася равнялась 65 мВт ст–1 м–2.
Ответы ганглиозных клеток регистрировали экстраклеточно от окончаний их аксонов в верхних слоях тектума заключенными в стеклянную оболочку металлическими платинированными микроэлектродами (Gaesteland et al., 1959) с диаметром платиновой шляпки 3–5 мкм с сопротивлением 200–500 кОм на частоте 1 кГц. Микроэлектрод под визуальным контролем микроманипулятором Sutter MP-285 подводили к интересующей области поверхности тектума (с учетом ретинотопической проекции, ориентируясь по картине кровеносных сосудов) и осторожно погружали, добиваясь стабильного одиночного отведения, о чем судили по величине импульсов и по соотношению сигнала и шума. Импульсы с выхода усилителя переменного тока с полосой пропускания от 100 Гц до 3.5 кГц (Виноградов, 1986) прослушивали при помощи громкоговорителя, наблюдали на экране осцилографа и вводили в память регистрирующего компьютера, оснащенного АЦП с частотой дискретизации 25 кГц.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЯ
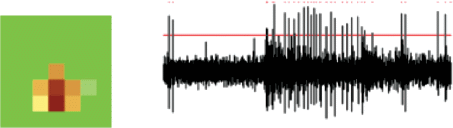
В рамках данного исследования были рассмотрены РП 37 ОИ ГК. Несмотря на то что предпочтительной для ОИ ГК является стимуляция линиями соответствующей ориентации, эти клетки могут давать также некоторый ответ на иные стимулы, например, на черные или белые пятна небольшого размера. Пример такого ответа дан на рис. 2. Это позволяет оценить размеры, форму их ВРП вспышками пятен, а также установить положение центра ВРП. Вспышки пятен предъявлялись в ячейках квадратной сетки в квазислучайном порядке (метод шахматной доски), после чего программа производила подсчет количества импульсов, возникающих в ответ на стимуляцию. В каждой ячейке сетки стимул предъявляли трижды. Серии стимуляции импульсной активности всегда начинали предъявлением стимула в центральной ячейке. В конце серии проводили повторную стимуляцию в центральном положении для контроля уровня ответа клетки. Результаты опыта, проведенного на детекторе горизонтального края, приведены на рис. 2. Область стимуляции поделена на 49 маленьких квадратов (пятен) размером чуть больше 1°. Клеточные ответы во всей области стимуляции представлены в географической палитре топографической карты. По результатам данной процедуры, область стимуляции смещалась таким образом, чтобы ее центр совпадал с предполагаемым (на основании измерений) центром ВРП исследуемой клетки. Форма ВРП исследованных клеток имела скорее однородный характер, без тенденции к большей протяженности в том или ином направлении.
Рис. 2.
Метод картирования ВРП вспышками пятен.
Слева приведен пример картированного РП детектора горизонтального края. В каждой точке светлое пятно вспыхивало 3 раза, в центральном положении проводилась контрольная стимуляция после окончания серии. Количество импульсов в ответе клетки в каждой точке стимуляции отражено в географической палитре. Справа приведен ответ клетки на стимуляцию вспышкой пятна.
Для исследования тонкой структуры РП изучаемых ОИ ГК мы использовали метод двухточечной стимуляции, которая позволяет проследить взаимодействие внутри разных зон РП. Схема эксперимента аналогична обмеру РП методом “шахматной доски“, но в данном случае в центральной ячейке стимул вспыхивает каждый раз, а параллельно с ним в квазислучайном порядке предъявляется еще один в другой ячейке сетки. Примеры результатов такого эксперимента представлены на рис. 3, а. Каждый такой эксперимент давал на выходе одну и ту же картину – ортогонально ориентированные вытянутые “восьмерки” зон активации и торможения, далеко выходящие за пределы измеренных ранее ВРП. При увеличении расстояния между ячейками можно наблюдать ограниченность таких зон (рис. 3, б).
Рис. 3.
Результаты экспериментов с двухточечной стимуляцией ОИ ГК.
Детекторы горизонтального (слева) и вертикального (справа) краев (а). Эксперимент, выполненный с увеличенным расстоянием между ячейками сетки (б). Расстояние между ячейками сетки составляет 0.55°.

Данные, приведенные в этом исследовании, противоречат тому, что показывают другие работы по изучению ориентационной избирательности в сетчатке рыб. Актуальная модель ориентационной избирательности, предложенная британскими исследователями (Antinucci et al., 2018), предполагает, что форма дендритного древа ОИ ГК вытянута в направлении предпочтительной ориентации, а торможение реализуется посредством tenm3+ амакриновых клеток, чье дендритное древо вытянуто ортогонально предпочтительной ориентации. Согласно нашим же измерениям, дендритное древо ОИ ГК, вероятно, имеет округлую форму и не вытянуто в каком-либо направлении. Похожая схема реализована у детекторов вертикального края сетчатки мыши, когда дендритное древо самой ГК имеет округлую форму, а дендрит тормозной амакриновой клетки вытянут ортогонально предпочтительной ориентации (Nath, Schwartz, 2016). Необходимо, впрочем, учитывать, что в данном случае аналогия условна, потому как в отличие от ориентационно-избирательных клеток сетчатки рыб, такие клетки у млекопитающих имеют не on-off природу, а демонстрируют избирательность, так же и к знаку контраста. Кроме того, “распознание“ точки как образующей вместе с центральной аппроксимацию отрезка предпочтительной ориентации происходит в том числе далеко за границами предполагаемого ВРП. Таким образом, есть вероятность, что кроме тормозных tenm3+ амакриновых клеток в формировании механизма ориентационной избирательности принимает участие еще один тип амакриновых клеток, усиливающий ответ и собственно формирующий ориентационное предпочтение. Можно предполагать, что одни и те же амакриновые клетки могут образовывать тормозные синапсы на одних ОИ ГК и возбуждающие на ОИ ГК с предпочтением ортогональной ориентации. Однако также ориентационная избирательность может формироваться и разными типами амакриновых клеток.
Список литературы
Виноградов Ю.А. Электронные приборы в электро-физиологических, морфологических и этологических исследованиях. Препринт № 13. Владивосток: ДВНЦ АН СССР. 1986. 23 с.
Зенкин Г.М., Пигарев И.Н. Детекторные свойства ганглиозных клеток сетчатки щуки. Биофизика. 1969. Т. 14. № 4. С. 722–730.
Максимов В.В., Максимова Е.М., Максимов П.В. Классификация дирекционально-избирательных элементов, регистрируемых в тектуме карася. Сенсорные системы. 2005. Т. 19. № 4. С. 342–356.
Максимов В.В., Максимова Е.М., Максимов П.В. Классификация ориентационно-избирательных элементов, регистрируемых в тектуме карася. Сенсорные системы. 2009. Т. 23. № 1. С. 13–23.
Максимова Е.М., Орлов О.Ю., Диментман А.М. Исследование зрительной системы нескольких видов морских рыб. Вопросы ихтиологии. 1971. Т. 11. № 5. С. 893–899.
Antinucci P., Hindges R. Orientation-Selective Retinal Circuits in Vertebrates. Front Neural Circuits. 2018.
Billota J., Abramov J. Orientation and direction tuning of goldfish ganglion cells. Visual Neurosci. 1989. P. 3–13.
Cronly-Dillon J.R. Units sensitive to direction of movement in goldfish tectum. Nature. 1964. V. 203. P. 214–215.
Dervies S.H., Baylor D.A. Mosaic arrangement of ganglion cell receptive fields in rabbit retina. J Neurophysiol. 1997. V. 78. P. 2048–2060.
Douglas R.N., McGuidan C.M. The spectral transmission of freshwater ocular media an interspecific comparison and a guide to potential ultraviolet sensitivity. Vision Res. 1989. V. 29 (7). P. 871–879.
Gaestesland R.C., Howland B., Lettvin J.Y. Pitts W.H. Comments on microelectrodes. Proc IRE. 1959. V. 47. P. 1852–1856.
Govardovskii V.I., Fyhrguist. N., Reuter T., Kuzmin D.G., Donner K. In search of the visual pigment template. Vis.Neurosci. 2000. V. 17 (4). P. 509–528
Jacobson M., Gaze R.M. Types of visual response from single units in the optic tectum and optic nerve of the goldfish. J. Exp Physiol. 1964. V. 49. P. 199–209.
Kawasaki M., Aoki K. Visual responses recorded from the optic tectum of the Japanese dace. Tribolodon hakonensis, J. Comp Physiol. 1983. V. 152. P. 147–153.
Liege B., Galand G. Types of single-unit visual responses in the trout’s optic tectum. Ed. A. Gudikov. Visual Information Processing and Control of Motor Activity. Bulgarian Academy of Sciences, Sofia, 1971. P. 63–65.
Nath A., Schwartz G.W. Cardinal Orientation Selectivity Is Represented by Two Distinct Ganglion Cell Types in Mouse Retina. J. Neurosci. 2016. V. 16. P. 3208–3221.
Wartzok D., Marks W.B. Directionally selective visual units recorded in optic tectum of the goldfish. J. Neurophysiol. 1973. V. 36. P. 588–604.
Yang G., Masland R.H. Direct visualization of the dendritic and receptive fields of direc-tionally selective retinal ganglion cells. Science. 1992. V. 258. P. 1949–1952.
Yang G., Masland R.H. Receptive fields and dendritic structure of directionally selective retinal ganglion cells. J. Neurosci. 1994. V. 14. P. 5267–5280.
Дополнительные материалы отсутствуют.
Инструменты
Сенсорные системы