

Сенсорные системы, 2020, T. 34, № 1, стр. 15-18
Роль акустических и вибрационных сигналов во взаимоотношениях паразитических двукрылых сем. Tachinidae и клопов-щитников
Л. С. Шестаков 1, 2, *, А. Эль Хашаш 2
1 Федеральное государственное бюджетное учреждение науки
Институт проблем передачи информации им. А.А. Харкевича РАН
127051 Москва, Большой Каретный пер. 19, Россия
2 Российский Университет Дружбы Народов
117198 Москва, ул. Миклухо-Маклая, 8, Россия
* E-mail:
Поступила в редакцию 09.10.2019
После доработки 25.10.2019
Принята к публикации 06.11.2019
Аннотация
Мухи семейства Tachinidae паразитируют на представителях многих отрядов насекомых. При поиске жертвы они часто ориентируются на ее коммуникационные сигналы. Показано, что вибрационные сигналы полужесткокрылых вызывают изменения поискового поведения мух на растении и повышают активность. Шумы, возникающие при перемещении мухи по растению, никак не влияли на активность клопов. Зараженные особи Pentatomidae сохраняют акустическую активность, но их сигналы могут существенно менять свою амплитудно-временную структуру по сравнению с контролем.
ВВЕДЕНИЕ
Вибрационные сигналы являются необходимым компонентом внутривидовых и межвидовых взаимодействий многих видов членистоногих (Cocroft, Rodríguez, 2005). Вибрации субстрата могут использоваться не только для поиска полового партнера, но и для обнаружения добычи или избегания хищников (Castellanos, Barbosa, 2006; Casas, Magal, 2006).
Хищные членистоногие при поиске жертвы часто используют случайные колебания, возникающие при перемещении по субстрату. Так, паукообразные находят свою добычу, используя вибрации, возникающие при ее движении (Barth, 1998). Сигналы, сопровождающие механическую активность насекомых, как правило, имеют заметно меньшую амплитуду, чем вибрационные сигналы, используемые для коммуникации, а их частотный максимум расположен в более высоком частотном диапазоне, что приводит к более сильному затуханию сигнала в субстрате (Barth, 1998). Однако хищные членистоногие с успехом используют и их для обнаружения добычи. Так, клоп Podisus maculiventris надежно локализует источник вибраций, создаваемых гусеницами, питающимися на растении (Pfannenstiel et al., 1995). Использование случайных вибрационных сигналов для обнаружения жертвы широко распространено среди паразитических перепончатокрылых (Meyhöfer, Casas, 1999). Личинки муравьиного льва используют колебания песка для обнаружения жертвы и ее поимки (Fertin, Casas, 2009).
Однако хищные насекомые не ограничиваются простым подслушиванием, некоторые из них имитируют сигналы для привлечения жертвы. Например, клоп-хищнец Stenolemus, охотящийся на пауков, имитирует вибрации, генерируемые борющейся добычей, чтобы привлечь паука (Wignall, Taylor, 2011). На другом виде хищных клопов – Phymata crassipes – показано, что он может имитировать вибрационные сигналы ухаживания жертвы и вступать с ней в перекличку (Gogala et al., 1984). Предполагалось, что таким образом хищник может привлекать потенциальную жертву, однако гипотеза не была проверена. В наших исследованиях на клопах семейства Asopinae было показано, что при случайных колебаниях субстрата они замирают и сидят неподвижно, что может быть связано с механизмом обнаружения жертвы и защитой от хищников (Shestakov, 2015). Аналогичное поведение отмечено для пауков-волков, которые замирают и остаются неподвижными в ответ на сигналы потенциального хищника (Sitvarin et al., 2016). Многие насекомые в таких ситуациях так же прекращают издавать коммуникационные сигналы и сидят неподвижно (Djemai et al., 2001).
Лабораторные исследования показали, что в отсутствие других стимулов паразитоид Telenomous podisi избирательно привлекается сигналом самки своего предпочитаемого хозяина – Euschistus heros (Laumann et al., 2007; 2011). В нашей работе мы проверили, как реагируют на вибрационные сигналы жертвы мухи семейства Tachinidae, сравнив поведенческую реакцию Gymnosoma nudifrons (Diptera, Tachinidae) на сигналы, возникающие при перемещении по субстрату и коммуникационные сигналы Pentatoma rufipes.
МАТЕРИАЛ И МЕТОДИКА
Исходный живой материал P. rufipes собран в августе 2019 г. в Серпуховском районе Московской области. Зараженные тахинами особи были отобраны и помещены в индивидуальные контейнеры. Gymnosoma, как правило, откладывает яйца на спинной стороне жертвы, что облегчает идентификацию. Всего в эксперименте принимало участие 28 самок G. nudifrons. Самок и самцов до опыта содержали совместно, так как мотивация для поиска жертвы у оплодотворенной самки теоретически должна быть выше.
В качестве стимулов были выбраны неспецифический сигнал, возникающий при перемещении P. rufipes по субстрату (рис. 1), и призывный сигнал самца (рис. 2). Выбор данного сигнала в качестве стимула был обусловлен тем, что именно этот сигнал клопы чаще всего издают в скоплениях, а следовательно, он с наибольшей вероятностью может использоваться хищниками для обнаружения жертвы. Стимул предъявлялся циклически в течение 5 мин. Каждая особь G. nudifrons использовалась в эксперименте только 1 раз, т.е. каждой особи предъявляли один стимул.


Для записи сигналов в полевых условиях и в лаборатории применяли лазерный виброметр PDV 100 (Polytec, Германия), пьезоэлектрический адаптер ГЗП 661. Частота дискретизации сигнала 44.1 кГц. Для генерации стимулов использовали вибростенд 4810 (Bruel & Kjaer).
Для изучения реакции на сигналы потенциальной жертвы и исключения влияния других стимулов (химических, визуальных) мух помещали в закрытую сверху прозрачным стеклом арену с соотношением диаметра и высоты 10 : 15 см, в центре которой находилась веточка растения, на которую подавали сигнал. Параллельно вели видеотипирование и запись вибрационных сигналов. Длительность каждого предъявления – 5 мин. Затем сравнивали общую длительность разных поведенческих элементов. Статистический анализ проводили в программах Excel и Statistica.
РЕЗУЛЬТАТЫ
Вибрации, возникающие при перемещении P. rufipes по растению, имеют неупорядоченную амплитудно-временную структуру и шумовой спектр с частотным максимумом в районе 200 Гц. Выделить отдельные элементы сигнала затруднительно (рис. 1).
Призывный сигнал самца, напротив, состоит из одиночных вибрационных посылок, хорошо различимых на фоне сопутствующих шумов (рис. 2). Средняя длительность посылки – 229 ± 15 мс. Частотный максимум – 116 ± 29 Гц. Спектр сигнала линейчатый. Сигнал издается вибрацией крыльев, при этом насекомое вытягивает конечности и наклоняется вперед. После эмиссии данного сигнала самец замирает и несколько секунд сидит неподвижно, ожидая ответа самки. Подробно поведение при эмиссии сигналов описано в предыдущей работе (Shestakov, 2015).
Нами были выделены следующие основные типы двигательных актов: взлет, ходьба, прыжок на месте, замирание на месте, кручение на месте, чистка антенн, чистка крыльев, так же измеряли латентные периоды от момента подачи сигнала до начала перемещения (рис. 3).
Рис. 3.
Общая длительность различных элементов поведения (с) G. nudifrons при демонстрации стимулов.
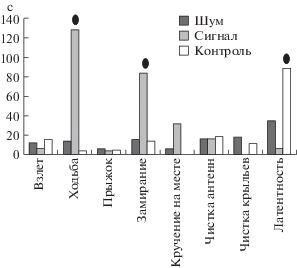
Общая длительность большинства поведенческих актов достоверно не отличалась в контроле и при демонстрации стимулов. Однако в случае предъявления призывного сигнала мухи достоверно больше перемещались по растению и замирали, прижавшись к субстрату (рис. 3). Латентные периоды от момента начала подачи сигнала так же отличались. В контроле мухи значительно дольше оставались без движения и в целом меньше перемещались по арене.
ОБСУЖДЕНИЕ
Полученные результаты свидетельствуют о том, что Tachinidae могут использовать вибрационные сигналы для обнаружения потенциальной жертвы. При этом наибольшее изменение активности наблюдалось при предъявлении именно коммуникационного сигнала в отличие от многих других беспозвоночных, которых привлекают так же и шумы, возникающие при перемещении жертвы по субстрату (Meyhöfer, Casas, 1999).
Данный факт можно объяснить особенностями экологии полужесткокрылых, для которых характерно образовывать осенние скопления перед зимовкой. Pentatoma rufipes – один из немногих видов, зимующих на стадии яйца, а не имаго, поэтому в таких скоплениях активно издает сигналы и копулирует. Призывные сигналы похожей структуры издают и другие представители Pentatomidae в осенних скоплениях (Shestakov, 2015), что наряду с химическими стимулами может служить хорошим ориентиром при поиске жертвы. Некоторые авторы полагают, что химические сигналы могут использоваться паразитоидом для поиска жертвы на расстоянии, а уже для локализации служат акустические и вибрационные сигналы (Virant-Doberl et al., 2019). В пользу этого говорит и тот факт, что длительность поискового поведения в случае с призывным сигналом была достоверно выше, чем в контроле – в среднем мухи перемещались по субстрату половину времени нахождения на арене. Так же латентный период до начала движения был существенно ниже по сравнению с контролем. В большинстве случаев насекомые начинали перемещаться по растению через 5–13 с после стимуляции. Это согласуется с данными ряда других авторов, говорящих о том, что вибрационные сигналы жертвы могут привлекать хищника (Casas, Magal, 2006; Barth, 1998).
Интересно отметить, что длительность такого элемента, как “замирание на месте”, была выше не в контроле, как можно было бы предположить, а при предъявлении призывного сигнала. Похожее поведение отмечено у перепончатокрылых и хищных клопов (Djemai et al., 2001, Casas, Magal, 2006).
Ответных вибрационных сигналов у мух зарегистрировано не было. Таким образом, вопрос могут ли они использовать вибрации не только для поиска жертвы, но и вступать с ней в перекличку, т.е. является ли процесс подслушивания одно- либо двусторонним, остается открытым и требует дальнейшего изучения.
Работа поддержана РФФИ (грант № 20-04-00553 а).
Список литературы отсутствует.
Дополнительные материалы отсутствуют.
Инструменты
Сенсорные системы