

Сенсорные системы, 2021, T. 35, № 1, стр. 38-42
Перспективы использования данных о вибрационной коммуникации для разработки безопасных методов контроля численности насекомых
Л. С. Шестаков 1, 2, *, А. Эль Хашаш 2
1 Федеральное государственное бюджетное учреждение науки
Институт проблем передачи информации им. А.А. Харкевича РАН
127051 Москва, Большой каретный пер., 19, Россия
2 Российский университет дружбы народов
117198 Москва, ул. Миклухо-Маклая, 8, Россия
* E-mail: zicrona@yandex.ru
Поступила в редакцию 23.09.2020
После доработки 02.11.2020
Принята к публикации 06.11.2020
Аннотация
Накопленные данные о вибрационной коммуникации насекомых показывают, что использование естественных и модифицированных сигналов вредителей может препятствовать их успешной коммуникации на растении, тем самым снижая их численность. В данной работе мы обсуждаем эффективность применения стимулов на примере широко распространенного и повреждающего большой спектр растений вида Pentatoma rufipes (L.).
ВВЕДЕНИЕ
В настоящее время традиционные методы мониторинга и контроля численности вредителей теряют свою эффективность. Так, к инсектицидам насекомые вырабатывают устойчивость и их приходится применять в более высоких концентрациях, что сказывается на качестве и безопасности продукции. Существует мнение, что продукция, произведенная с применением большого количества пестицидов, может служить одной из причин возросшего количества онкологических заболеваний и аллергических реакций.
В связи с этим во многих странах Европейского Союза, Азии, в США нарастает тенденция перехода к органическому земледелию и новым методам мониторинга и контроля численности вредителей. Одним из таких методов является использование вибрационной коммуникации насекомых для их обнаружения или управления поведением (de Groot et al., 2010, Avosani et al., 2020).
В связи с перемещением между странами больших партий сельскохозяйственной продукции, семенного и посадочного материала, возросшим числом транспортных перевозок, которые могут служить путем проникновения опасных карантинных вредителей (таких как гватемальская картофельная моль, коричнево-мраморный клоп, и другие карантинные виды), существующие методы досмотра и диагностики не всегда являются достаточными. В случаях с посадочным материалом, необработанной древесиной и многими другими факторами, вредители могут быть скрыты в твердых субстратах и их визуальное обнаружение невозможно. Для таких субстратов могут быть эффективны акустические методы мониторинга. При этом всегда есть опасность, что инвазивные виды могут “обрушить” всю местную экосистему и нанести непредсказуемый экономический ущерб. Так, проникший на юг Италии опасный инвазивный вредитель Halyomorpha halys (Stål), повреждающий широкий спектр растительных культур, уже в 2017–2018 гг. был обнаружен на севере страны (Malek et al., 2019).
При этом многие виды, обычные для средней полосы России, в странах Европы являются вредителями, которые могут наносить серьезный ущерб (Panizzi et al., 2000). Так, широко распространенный в России клоп Pentatoma rufipes (L.) в северной Европе может значительно вредить фруктовым насаждениям (Powell, 2020). Развитию сельского хозяйства в новых перспективных регионах могут препятствовать в том числе и высокие риски перехода таких видов с дикорастущих растений на новые сельскохозяйственные сорта.
Этот комплекс причин определяет необходимость поиска и развития новых методов контроля численности вредителей (Panizzi et al., 2000). Акустический метод контроля по степени воздействия на окружающую среду является наиболее безопасным. Более того, появляется все больше данных о его перспективности и способности повышать эффективность таких традиционных методов, как клеевые и феромонные ловушки (Suckling et al., 2019).
В предыдущем исследовании на Pentatoma rufipes (L.) мы показали, что дизруптивные сигналы самок, воспроизводимые на кормовое растение, могут быть эффективным стимулом, препятствующим копуляции (Шестаков, 2020). В данной работе мы проверили эффективность других типов сигналов, как привлекающих стимулов.
МЕТОДИКА
Регистрация и анализ сигналов
Для регистрации сигналов применяли лазерный виброметр PDV 100 (Polytec, Germany). Портативный лазерный виброметр PDV 100 разработан, в частности, для регистрации вибраций живых объектов. Он имеет следующие преимущества по сравнению с контактными датчиками: позволяет минимизировать влияние исследователя на объект, возможность получения высокого соотношения сигнал/шум (высокая помехозащищенность); позволяет измерять механические колебания сложной формы на значительных расстояниях от объекта измерения; минимизация влияния параметров внешней среды на параметры сигнала в отличие от пьезоэлектрических датчиков; компактность, автономность и малый вес для работы в полевых условиях, возможность работы с программным обеспечением стороннего производителя для обработки сигналов.
Пьезокристаллический адаптер ГЗК-661 идеален в условиях, когда использование лазерного виброметра затруднено, например, для записи сигналов в густом переплетении растений или при плохих погодных условиях в полевых условиях. Основными плюсами пьезоэлемента являются простота и скорость крепления датчика, т.е. он необходим прежде всего для записи первичного сигнала в полевых условиях. Лазерный виброметр необходим для калибровки тестовых стимулов, что не позволяет сделать пьезоэлектрический адаптер.
Для регистрации (проверки наличия) акустической компоненты сигнала запись параллельно велась на микрофон (4191, 1/2 inch; Bruel & € Kjær, Nærum, Denmark), расположенный на расстоянии 1–3 см от насекомых.
Кроме того, регистрацию вибрационных сигналов в лаборатории проводили на мембране низкочастотного микрофона, как на нерезонансном субстрате. Данный метод, несмотря на простоту, до сих пор применяется при записи сигналов, используемых для создания модельных стимулов, чтобы минимизировать влияние живого природного субстрата на исходную структуру сигнала (Cokl, 2008).
Частота дискретизации сигнала во всех случаях 44.1 кГц. Такая частота более чем достаточна, так как доминантная частота вибрационных сигналов в основном находится в районе 200 Гц.
Измерение амплитудно-временных параметров сигналов проводили в программе CoolEdit (США, Syntrillium).
Предъявление стимулов
Для генерации стимулов применяли вибростенд 4810 (Bruel & Kjaer). Оборудование дает возможность воспроизводить сигнал с заданной частотой и амплитудой в частотном диапазоне от 0 до 18 кГц, что полностью покрывает частотный диапазон вибраций насекомых. В качестве стимулов служили ранее зарегистрированные естественные коммуникационные сигналы. Предполагается, что использование вибрационных стимулов более перспективно, чем акустических, так как они подаются непосредственно на растение и не оказывают раздражающего эффекта на другие объекты, находящиеся в окружающем пространстве. Новые данные говорят о том, что на практике вибрационные стимулы должны быть эффективны на широком спектре растений, за исключением разве что травянистых культур.
В качестве стимулов использовали фрагменты призывного (рис. 1, б) и конкурентного сигналов (д) длительностью 5 с.
Рис. 1.
Осциллограммы (а, б, г, д) и сонограммы (в, е) конкурентного (а–в) и призывного (г–е) сигналов самца Pentatoma rufipes. Развертка осциллограмм: a, г – 30 с; б, д – 5 с. Сонограммы даны для осциллограмм – б, д.

В эксперименте использовали самок Pentatoma rufipes (L.), выведенных из нимф и не имевших опыта копуляции.
Для фото и видеотипирования поведения во время экспериментов использовалось следующее оборудование: видеокамера Sony HDR-CX405 и системная камера Sony Alpha Nex-3N + макрообъектив Sony SEL 30M35. Для содержания культуры насекомых использовались террариумы Hagen ExoTerra 30 × 30 × 45 см.
РЕЗУЛЬТАТЫ
Самцы P. rufipes L. издают два основных типа вибрационных сигналов – конкурентные (а–в) и призывные (г–е).
Конкурентные сигналы самцов представлены серией непродолжительных посылок. Они издаются при непосредственном контакте самцов. При эмиссии сигнала насекомое приподнимается над субстратом и покачивается в сторону соперника, насекомые могут сцепляться передними лапами и пытаться сбросить друг друга с субстрата. После эмиссии сигнала на растении всегда остается один самец, или самец и самка.
Призывные сигналы издаются как одиночно, так и в смешанных по полу группах и не сопровождаются агонистическим поведением. Подробно характеристики вибрационных сигналов описаны в предыдущей работе (Shestakov, 2015).
В качестве стимулов мы использовали и призывные (N = 16), и конкурентные (N = 15) сигналы. Это связано с тем, что теоретически сигналы могут быть полифункциональными и конкурентный сигнал для самца (для самки) может, наоборот, нести информацию о потенциальном партнере.
Оценивали два основных поведенческих ответа самок на стимул – латентный период от начала применения стимула до старта активного перемещения самки и процент успешных подходов к источнику стимула. Контроль – отсутствие стимула (N = 15).
Латентные периоды при стимуляции и без значительно отличались. В большинстве случаев при предъявлении стимула самки переходили из состояния покоя/питания в состояние активного перемещения по растению. В контроле насекомые в основном сидели на растении, либо перемещались на незначительные расстояния, меняя точку питания. При этом процент положительных ответов на стимулы был значительно выше, чем в контроле (U – критерий Манна–Уитни; *p < 0.05). Латентный период до начала активности в отсутствие стимула, наоборот, был значительно больше (U – критерий Манна–Уитни; p < 0.05) (рис. 2).
Рис. 2.
Латентные периоды до начала движения и процент успешных подходов к источнику стимула у самок Pentatoma rufipes.
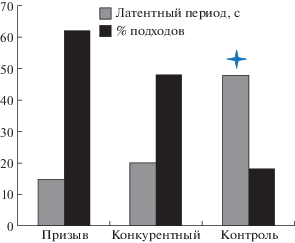
Достоверных различий в количестве подходов к источнику стимула при трансляции призывного и конкурентного сигнала не выявлено. Однако заметна тенденция увеличения количества подходов при проигрывании призывного сигнала (62 и 48% соответственно).
ОБСУЖДЕНИЕ
Применение вибрационных стимулов эффективно меняет поведение насекомых на растении (Djemai et al., 2001; de Groot et al., 2010). В нашей предыдущей работе показано, что искусственное предъявление дизруптивных сигналов самки эффективно препятствует копуляции уже сложившихся пар, чем снижает численность насекомых на растении (Шестаков, 2020). Эти данные хорошо согласуются с результатами, полученными на других группах насекомых. Так, на листоблошках показано, что трансляция дизруптивных сигналов самцов снижает численность насекомых на растении винограда (Nieri, Mazzoni, 2018; Avosani, 2020). Этот эффект связан с невозможностью успешной коммуникации и нарушением поиска и распознавания самцов и самок из-за помех, создаваемых транслируемым стимулом. Предполагается, что растение перестает быть комфортным субстратом для насекомых в виду невозможности успешной коммуникации на нем.
Применение привлекающих стимулов также может быть весьма эффективным. Реакции P. rufipes на конкурентный и призывный сигналы достоверно отличались от контроля, и процент успешных подходов к источнику стимула был достаточно высок. Незначительная разница в реакции самок на стимулы, созданные на основе конкурентного и призывного сигналов самца, по нашему мнению, может быть обусловлена несколькими факторами: сходством частотного диапазона и возможной полифункциональностью сигнала агрессии. Многие животные могут использовать один и тот же тип сигнала для решения разных задач. В данном случае сигнал агрессии используется при взаимодействии между самцами, но теоретически он может давать находящимся рядом самкам информацию о находящихся рядом самцах и их индивидуальных характеристиках.
Оба типа сигналов эффективно привлекали самок к источнику стимула по сравнению с контролем. Таким образом, вибрационные стимулы могут использоваться как дополнительный фактор привлечения насекомых в различные типы ловушек: клеевые, феромонные. Некоторые данные говорят о том, что совместное использование феромонных и вибрационных стимулов делает клеевые ловушки достоверно более привлекательными.
Кроме того, сигналы жертвы могут привлекать на растение потенциальных хищников и паразитов (Virant-Doberlet et al., 2019). Трансляция искусственных сигналов вредителей могла бы заранее привлекать на растение поедающих их хищников и тем самым предотвращать массовые вспышки численности.
Мы считаем, что в условиях перехода к безопасному органическому земледелию акустические методы контроля могли бы стать хорошей альтернативой пестицидам и дополнительным инструментом, наряду с феромонными, механическими ловушками и бактериальными препаратами.
Исследование выполнено при поддержке РФФИ (грант № 20-04-00553 А).
Список литературы
Шестаков Л.С. Дизруптивные сигналы – эффективный механизм прерывания нежелательной копуляции у Pentatoma rufipes. Сенсорные системы. 2020. Т. 34 (1). С. 12–14. https://doi.org/10.31857/S0235009220010102
Avosani S., Sullivan T.E, Ciolli M., Mazzoni V., David Maxwell Suckling. Can Vibrational Playbacks Disrupt Mating or Influence Other Relevant Behaviours in Bactericera cockerelli (Triozidae: Hemiptera)? Insects. 2020. V. 11. P. 299. https://doi.org/10.3390/insects11050299
Cokl A. Stink bug interaction with host plants during communication. J. Ins. Physiol. 2008. V. 54. P. 1113–1124.
Djemai I., Casas J., Magal C. Matching host reactions to parasitoid wasp vibrations. Proc. R. Soc. B. 2001. V. 268. P. 2403–2408. https://doi.org/10.1098/rsbp.2001.1811
de Groot M., Cokl A., Virant-Doberlet M. Effects of heterospecific and conspecific vibrational signal overlap and signal-to-noise ratio on male responsiveness in Nezara viridula (L.). J. Exp. Biol. 2010. V. 213. P. 3213–3222.
Nieri R., Mazzoni V. Open-field vibrational mating disruption: the effect on leafhopper pests and their predators. IOBC-WPRS Working Group “Integrated Protection in Viticulture”. 2018. V.139. P. 31–34.
Malek R., Zapponi L., Eriksson A., Ciolli M., Mazzoni, Anfora G., Tattoni C. Monitoring 2.0: Update on the Halyomorpha halysInvasion of Trentino. ISPRS Int. J. Geo-Inf. 2019. V. 8. P. 564. https://doi.org/10.3390/ijgi8120564
Powell G. The biology and control of an emerging shield bug pest, Pentatoma rufipes (L.) (Hemiptera: Pentatomidae) . Agricultural and Forest Entomology. 2020. https://doi.org/10.1111/afe.12408
Panizzi A.R., McPherson J.E., James D.G., Javahery M., McPherson R.M. Economic importance of stink bugs (Pentatomidae). Eds Schaefer C.W., Panizzi A.R. Heteroptera of Economic Importance. CRC Press, Boca Raton, Florida. 2000. P. 421–474.
Shestakov L.S. A comparative analysis of vibrational signals in 16 sympatric bug species (Pentatomidae, Heteroptera). Entomological Review. 2015. V. 95. № 3. P. 310–325.
Suckling D.M., Mazzoni V., Roselli G., Levy M., Ioriatti C., Stringer L., Zeni V., Deromedi M., Anfora G. Trapping Brown Marmorated Stink Bugs: “The Nazgȗl” Lure and Kill Nets. Insects. 2019. V.10. P. 433. https://doi.org/10.3390/insects10120433
Virant-Doberlet M., Kuhelj A., Polajnar J., Šturm R. Predator-Prey Interactions and Eavesdropping in Vibrational Communication Networks. Front. Ecol. Evol. 2019. V. 7. P. 1–15.
Дополнительные материалы отсутствуют.
Инструменты
Сенсорные системы