

Сенсорные системы, 2021, T. 35, № 4, стр. 328-339
Взаимодействие фовеального и периферического зрения при реализации системных зрительных функций
Л. Н. Подладчикова 1, Д. Г. Шапошников 1, *, А. И. Самарин 1, Д. М. Лазуренко 1
1 Научно-исследовательский технологический центр нейротехнологий Южного федерального университета
344090 Ростов-на-Дону, пр-т Стачки, 194, Россия
* E-mail: dgshaposhnikov@sfedu.ru
Поступила в редакцию 14.12.2020
После доработки 01.07.2021
Принята к публикации 04.08.2021
Аннотация
В работе проанализированы известные факты об отражении системных функций зрения, таких как дискриминация и распознавание зрительных объектов; зрительный поиск; оценка эмоционального содержания сцены и принятие решений в фовеальном и периферическом поле зрения. Рассмотрены известные гипотезы о возможных механизмах функциональных феноменов, обнаруженных в периферическом зрении человека. Описан нейроинформационный подход к решению проблем взаимодействия фовеального и периферического зрения на основе траекторий осмотра, областей интереса и возвратных фиксаций взгляда. В компьютерных экспериментах было показано, что структура модельной траектории осмотра коррелирует с количеством возвратных фиксаций “входного окна” модели. Это позволило сделать предположение, что вероятность возвратных фиксаций можно рассматривать как количественный критерий для определения типа внимания (фокальное или пространственное) и момента его переключения.
ВВЕДЕНИЕ
В настоящее время формируется устойчивый тренд к пересмотру классических представлений о механизмах регуляции зрительного внимания, в частности, о специфике сенсорной настройки центрального (фовеального) и периферического зрений (Рожкова и др., 2019; Шелепин и др., 2009; Burnat, 2015; Cajar et al., 2016; Larson, Loscky, 2009; Laubrock et al., 2013; Loschky et al., 2017, 2019; Rosenholtz, 2016; Shaposhnikov et al., 2001; Stephenson et al., 2020; Tanrikula et al., 2020; Van Diepen, d’Ydewalle, 2003). Классическое представление о роли центрального (фовеального) и периферического механизмов зрения состоит в том, что они функционируют автономно. Так, в процессе осмотра первое, имеющее высокую остроту зрения за счет высокой плотности фоторецепторов – колбочек, обеспечивает анализ тонких деталей зрительного образа во время фиксации взгляда, второе – способствует выбору информативных областей изображений (сцен) для последующих фиксаций взгляда (Барабанщиков, 1997; Величковский, 2006; Подвигин и др., 1986; Подладчикова и др., 2015; Хьюбел, 1990). Известные сведения о свойствах элементов в области представительства периферии рецептивного поля, таких как низкая плотность колбочек, большие размеры рецептивных полей ганглиозных и корковых клеток, выраженная чувствительность к движению и к крупным объектам, преобладание нейронов Y-типа в мозговых зрительных центрах (Подвигин и др., 1986; Хьюбел, 1990), рассматриваются как основания для доминирующих представлений о роли этой части поля зрения. Такая архитектура непосредственно сказывается на особенностях зрительного восприятия, а именно, – снижении контрастной чувствительности (Pointer, Hess, 1989), остроты зрения (Anstis, 1974), способности к пространственной локализации объектов по мере увеличения эксцентриситета.
Однако в последних теоретических и экспериментальных исследованиях получены результаты, не согласующиеся с этим представлением (Burnat, 2015; Cajar et al., 2016; Hennig, Worgotter, 2003; Hughes et al., 2016; Kragic et al., 2006; Kuraguchi, Ashida, 2015; Ludwig et al., 2014; Rosenholtz, 2016; To, 2011; Torralba et al., 2006; Van Diepen, d’Ydewalle, 2003). Известно несколько обзоров о взаимодействии центрального и периферического зрений (Рожкова и др., 2019; Levi, 2008; Loschky et al., 2017; Rosenholtz, 2016; Stewart et al., 2020; Strasburger, 2011; Wiecek et al., 2012). В работе (Loschky et al.,2017) рассматриваются результаты, полученные в исследованиях, посвященных различным аспектам взаимодействия центрального и периферического зрений при предъявлении простых и семантически значимых стимулов. Подчеркивается, что периферическое и центральное зрение динамически и непрерывно взаимодействуют друг с другом и эта проблема требует дальнейшего детального изучения.
На основании анализа современных данных (Рожкова и др.,2019) указывают на значительное сходство периферического восприятия и фовеального зрения при условии пространственного масштабирования ряда параметров тестовых стимулов, в том числе при увеличении их размеров, яркости, уровня контраста с увеличением эксцентриситета и оптимизации других условий эксперимента с учетом нейрофизиологических механизмов внимания и особенностей структуры зрительных нейронных путей в корковых областях представительства периферического зрения. Кроме того, рассматривается вклад особенностей оптической системы глаза и цветового восприятия в обработку зрительной информации на периферии поля зрения. В обсуждении подчеркивается необходимость разработки комплекса новых методов для преодоления заблуждений в понимании особенностей функциональных возможностей периферического зрения.
Результатами исследований (Rosenholtz, 2016) убедительно обосновывается тот вывод, что периферическое поле зрения непосредственно участвует в решении широкого спектра зрительных задач, в том числе в реализации функций, типичных для центрального зрения. Автором обсуждаются также вопросы и перспективы для будущих исследований на пути к формированию нового понимания роли периферического зрения, прежде всего касающиеся функциональных свойств периферии. В частности, должно быть определено: какие первичные признаки изображений на периферии зрения определяют оптимальное решение частных зрительных задач; к каким физиологическим феноменам приводит потеря точности при выделении значимых признаков в периферическом зрении; каковы основы взаимодействия между зрительным вниманием и реализацией конкретных зрительных задач; какова роль информации на периферии поля зрения в реализации процессов принятия решений на верхнем уровне зрения.
В обзоре (Strausburger et al., 2011) рассматриваются исследования периферического зрения и их отношение к теориям восприятия формы, гипотезе коркового фактора магнификации, распознавание символов в периферическом зрении при низком и высоком уровне контраста, а также применение парадигмы краудинга как подхода к оценке вклада контекста и окружающих контуров. В исследованиях с распознаванием более сложных стимулов (текстуры, лица и сцены) получены результаты, указывающие на участие факторов среднего уровня и высших когнитивных факторов. Вместе с тем было показано, что простые задачи обнаружения выполняются обследуемыми одинаково успешно в любой области поля зрения при условии, что стимулы имеют подходящие размеры.
МЕТОДЫ ИЗУЧЕНИЯ НЕРЕШЕННЫХ ПРОБЛЕМ ВЗАИМОДЕЙСТВИЯ ФОВЕАЛЬНОГО И ПЕРИФЕРИЧЕСКОГО ЗРЕНИЙ
Один из методов экспериментального исследования нерешенных проблем взаимодействия центрального и периферического зрения, совмещаемый с другими методами (в частности, с фильтрацией определенных пространственных частот), состоит в избирательном блокировании той или иной части поля зрения (фовеальной, парафовеальной или периферической). В пионерских работах, в которых использовались специальные фиксирующие присоски на поверхности роговицы и избирательное физическое блокирование той или иной части поля зрения, было показано, что такие манипуляции приводят к существенному изменению длительности фиксаций взгляда и амплитуды саккад, что проявляется и в изменении пространственной структуры траектории глазных движений при осмотре одного и того же изображения (Рожкова, Ярбус, 1977; Ярбус, Рожкова, 1977; Рожкова и др., 2019).
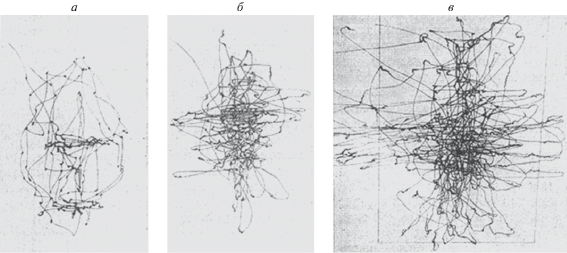
В частности, блокирование фовеального поля зрения при решении различных зрительных задач приводит к возрастанию количества глазных движений и их амплитуды (Рожкова, Ярбус, 1977; Ярбус, Рожкова, 1977; Рожкова и др., 2019). Пространственная структура траекторий глазных движений в условиях, когда при осмотре закрыта центральная (фовеальная) часть поля зрения, имеет более сложный характер (рис. 1, б,в) по сравнению со структурой траектории при открытой фовеальной области (а).
Рис. 1.
Регистрации движений глаза, полученные при рассматривании одного и того же полутонового портрета в отсутствие окклюдера (а), и при использовании окклюдеров диаметром 5°(б) и 10°(в), перекрывающих фовеальное поле зрения. (Модифицированный pис. 12 из работы Рожкова и др., 2019).
Еще один метод состоит в применении специальных контактных линз (Иомдина и др., 2020) с имплантированным окклюдером. В настоящее время разрабатываются перспективы использования таких линз для изучения дальней периферии (60–90 град.) и решаются технические проблемы, прежде всего связанные с индивидуальными особенностями оптической системы глаза.
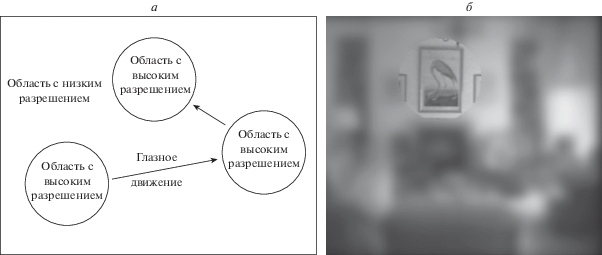
Вместе с тем разработан комплекс экспериментальных методов, обеспечивающих изменение видимой части поля зрения при помощи специализированных компьютерных программ. Наибольший интерес представляют искусственные скотомы в произвольном месте поля зрения или предъявляемого изображения, а также метод, в основе которого лежит использование дисплея с регулируемым программным множеством уровней пространственного разрешения (gaze-contingent multiresolution) (Cajar et al., 2016; Cornelissen et al., 2005; Ringer et al., 2016; Larson, Loschky, 2009; Ludwig et al., 2009; Parkhurst, Niebur, 2002). Суть последнего метода состоит в определении позиции четко видимой части поля зрения на основе координат текущей позиции фиксации взгляда человека в процессе осмотра (рис. 2).
Рис. 2.
a – схематическое представление метода gaze-contingent display с двумя уровнями разрешения; область с высоким разрешением отслеживает координаты центра взгляда человека. б – пример расположения области с высоким пространственным разрешением (модифицированный рис. 1 из работы Parkhurst, Niebur, 2002).
В экспериментальных исследованиях, проведенных при помощи этих методов, показано, что фовеальное и периферическое зрения функционируют параллельно с самых первых моментов фиксации взгляда (Cajar et al., 2016; Ludwig et al., 2014; Van Diepen, d`Ydewalle, 2003), при этом периферическое зрение играет важную роль в дискриминации зрительных объектов (To et al., 2011; Yu, Shim, 2016), оценке семантического и эмоционального содержания изображений (Kuraguchi, Ashida, 2015). Обнаружено, что фовеальное и периферическое зрения динамически взаимодействуют друг с другом, в частности, модуляция фовеального представительства оказывает влияние на решение задачи дискриминации зрительных объектов в периферической области (Yu, Shim, 2016). Примечательно, что этот феномен проявляется только при предъявлении сложных трехмерных стимулов и отсутствует при использовании простых двумерных стимулов, например решеток. Однако проблема механизмов, обеспечивающих эти феномены, в настоящее время остается полем для различных гипотез, не верифицированных однозначно в эксперименте. Известно несколько предположений о возможных механизмах. В частности, при решении сложных зрительных задач в периферическом поле зрения возможно участие полимодального признакового взаимодействия, настройки систем нижнего уровня за счет обратной связи от коры головного мозга, механизмов внимания и долговременной памяти (Burnat, 2015; Loschky et al., 2019). Эти предположения могут быть проверены в направленных экспериментах при помощи комплекса методов, включающих регистрацию движений глаз, идентификацию областей интереса и возвратных фиксаций взгляда, а также математического моделирования.
НЕЙРОИНФОРМАЦИОННЫЙ ПОДХОД К РЕШЕНИЮ ПРОБЛЕМ ВЗАИМОДЕЙСТВИЯ ЦЕНТРАЛЬНОГО И ПЕРИФЕРИЧЕСКОГО ЗРЕНИЙ
Неизученность механизмов выбора областей интереса на периферии поля зрения в реальной зрительной системе приводит к тому, что при разработке искусственных систем, основанных на активном фовеальном зрении, эта проблема решается на основе эвристических алгоритмов, построения карт выделенности (saliency maps) c учетом результатов психофизических тестов о свойствах точек фиксаций взгляда при осмотре конкретных изображений (Самарин и др., 2020).
Несмотря на устойчивый тренд к пересмотру ряда базовых представлений о роли фовеального и периферического полей зрения и о механизмах их взаимодействия, практически все исследования и разработки в области искусственного активного фовеального зрения до настоящего времени основаны на трех постулатах:
1 – центральное и периферическое поля зрения функционируют последовательно;
2 – первое – обеспечивает анализ тонких деталей зрительных сцен (изображений);
3 – второе – способствует выбору информативных областей изображений (сцен) для последующей обработки на высоком уровне разрешения (Samarin et al., 2019; 2020).
Очевидно, неполнота имеющихся экспериментальных данных, и в ряде случаев их противоречивость, а также отсутствие стандартизации экспериментальных условий затрудняют возможность обобщения и формализации результатов, полученных в различных психофизических исследованиях (Подладчикова и др., 2017; Carrasco, 2012; Foulsham, Kingstone, 2012). Как известно, условия экспериментов (от тестовой задачи до вида стимулов и способа отчета тестируемых) существенно варьируются в исследованиях, проведенных в различных лабораториях, и оказывают значительное влияние на результаты, вплоть до получения противоположных.
Тем не менее решение указанных проблем (Privitera, Stark, 2005; Samarin et al., 2015) может быть найдено в процессе создания реалистичной модели зрительного внимания, которая позволит на качественно новом уровне анализировать вклад факторов и механизмов различного типа при формировании траектории осмотра и формулировать предположения, доступные экспериментальной проверке.
В основе предлагаемого нами нейроинформационного подхода лежит, как и в наших прежних исследованиях (Самарин и др., 2020; Lomakina et al., 2016; Podladchikova et al., 2020; Samarin et al., 2015; 2019), теория активного зрения, а также формализация известных экспериментальных данных в области взаимодействия фовеального и периферического зрений в реалистичных моделях, учитывающих анализ траекторий осмотра, областей интереса и возвратных фиксаций взгляда. В настоящее время известно несколько моделей взаимодействия фовеального и периферического зрения (Foulsham, Kingstone, 2012; Laubrock et al., 2019). Эти модели рассматривают динамику амплитуды саккад и длительности фиксаций и игнорируют, в отличие от нашего оригинального подхода, структуру траекторий осмотра, весьма информативные области интереса и особенности формирования возвратных фиксаций (Подладчикова и др., 2017, Podladchikova et al., 2020).
Разработанная в рамках нового нейроинформационного подхода модель формирования траектории осмотра изображений и сцен (рис.3, а) основана на имитации пространственно-неоднородного представления входной информации от центра к периферии поля зрения (Podladchikova et al., 2021b). В качестве входной информации использовалась признаковая карта, сформированная на основе распределения первичных признаков изображения (яркость, градиент, контраст), выделенных в различных цветовых пространствах (RGB, HSV, LSV), их комбинаций (углы, протяженные перепады, области с резкими изменениями яркости и другие), семантически значимых областей (люди, лица людей, надписи, вывески) и областей интереса, сформированных на основе результатов анализа точек фиксации взгляда в исследованиях с участием добровольцев.
Рис. 3.
а – базовая структура и процедуры модели формирования траектории осмотра изображений и сцен. б – схема “входного окна” модели.

Основной процедурой в модели являлось определение координат и времени (количества циклов модели) следующей точки фиксации “входного окна” (рис. 3, б). Для каждого узла “входного окна” вычислялась сумма значений признаковой карты в пределах его “рецептивного поля” (б). За один цикл работы модели значения признаковой карты в пределах фовеальной области “входного окна” (r = 2.5°) уменьшались на величину параметра IOR, имитирующего механизм подавления возврата взгляда. Центр “входного окна” смещался в узел с максимальной суммой, нормированной на размер “рецептивного поля”.
В том случае, когда несколько узлов “входного окна” имели одинаковую сумму, следующая точка фиксации выбиралась с учетом признаков, представленных в соседних с конкурирующими узлах. Продолжительность фиксации в каждой точке зависела от количества циклов модели, в течение которых “входное окно” оставалось в одной и той же точке.
РЕЗУЛЬТАТЫ КОМПЬЮТЕРНЫХ ЭКСПЕРИМЕНТОВ И ИХ ОБСУЖДЕНИЕ
При проведении модельных экспериментов структуру траектории осмотра и количество возвратных фиксаций “входного окна” модели оценивали при условии варьирования параметра IOR. В качестве тестовых изображений использовали стоп-кадры видеоклипов из аннотированной базы данных LIRIS-ACCEDE, содержащей эмоционально значимые видеоизображения (http://liris-accede.ec-lyon.fr) (Baveye et al., 2015).
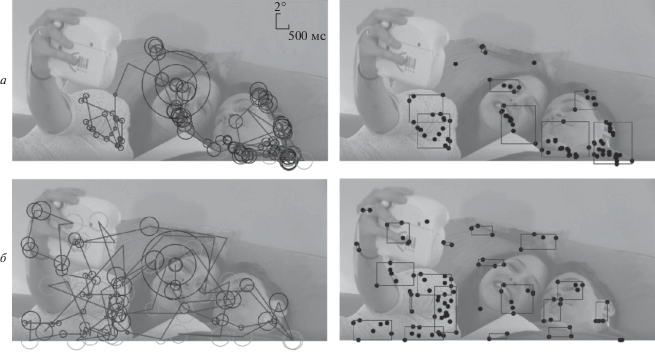
Экспериментально продемонстрировано, что при малых значениях коэффициента IOR модель зрительного восприятия, в большинстве случаев 60%, формировала траектории фокального типа (рис. 4, а). Напротив, при больших значениях коэффициента IOR преобладали модельные траектории пространственного типа (б).
Рис. 4.
Примеры модельных траекторий осмотра.
а – фокального типа (IOR =1) и б – пространственного типа (IOR = 5), сформированных при “осмотре” стоп-кадра видеоклипа № 9748 (левая колонка), и областей интереса (правая колонка). Диаметр окружностей пропорционален продолжительности фиксации окна ввода в текущих точках.
Кроме того, было показано, что вероятность формирования возвратных фиксаций достоверно (p < 0.05) коррелирует со структурой траектории осмотра изображений, а именно, для фокальных траекторий она составляла 0.125 ± 0.075, тогда как для пространственных – 0.005 ± 0.001. Результаты проведенного моделирования хорошо согласуются с базовой концепцией в рамках теоретических моделей управления глазными движениями (Reingold et al., 2012; Trukenbrod, Engbert, 2014). Согласно общим положениям теории, в условиях высокой семантической нагрузки наблюдается сопряженное повышение длительности фиксации взгляда. Исходя из полученных в ходе тестирования модели фактов, можно сделать вывод, что не только длительность фиксации, но и количество возвратов являются значимыми признаками, указывающими как на доминирующий тип зрительного восприятия, так и на прогнозируемую траекторию осмотра изображения, включая семантически значимые зоны интереса (Baddeley, Tatler, 2006). С точки зрения механизма глобального контроля глазных движений (Trukenbrod, Engbert 2014), согласно которому длительность фиксации взгляда определяется наиболее общими, в том числе физическими особенностями и характеристиками стимульного материала, поисковой задачей и индивидуальными стратегиями их решения, в разработанной нами модели получены результаты, указывающие на непосредственную роль адаптационных процессов, наблюдаемых при взаимодействии и переключении пространственного и фокального типов обработки зрительной информации.
Предположения, сформулированные в рамках новой модели, были проверены при обработке результатов ранее проведенных психофизических исследований (Подладчикова и др., 2017; Podladchikova et al., 2009; Podladchikova et al., 2018; Podladchikova et al., 2020; Samarin et al., 2015). Использовались экспериментальные данные, полученные на одной и той же выборке добровольцев (n = 12) от 19 до 28 лет в условиях свободного просмотра изображений и поиска модифицированных фрагментов трех репродукций (Ярбус, 1965) (“Не ждали” Репина И.Е., “Лес графини Мордвиновой” И.И. Шишкина, “Березовая роща” И.И. Левитана). Экспериментальная методика была утверждена комиссией по биоэтике НИИ нейрокибернетики им. А.Б. Когана ЮФУ (протокол. № 2 от 05.02.2008 г.), а сами исследования проводились в соответствии с этическими принципами и требованиями Хельсинкской декларации.
Выявлено, что вероятность формирования возвратных фиксаций взгляда при свободном просмотре изображений была достоверно выше, чем при решении поисковой задачи (p < 0.05 по критерию t-Стьюдента для независимых групп). Кроме того, результаты согласуются со сведениями, полученными ранее при помощи других методов анализа глазных движений. В частности, при свободном осмотре изображений и решении поисковых задач обнаружены существенные отличия, как по показателям амплитуды саккад и продолжительности фиксации, так и по количеству областей интереса и общей площади зон просмотра.
Обсуждая полученные результаты, можно предположить, что существует определенный консенсус, в рамках которого центральное и периферическое поля зрения успешно решают конкурирующие задачи, достигают своей цели и эффективно функционируют при наличии весьма ограниченных ресурсов, необходимых для обработки информации. В то время как фовеальное зрение позволяет обеспечить максимальную остроту и контрастную чувствительность в небольшом поле вокруг центральной оси, совпадающей с положением взгляда, периферическое зрение позволяет успешно детектировать объекты с высокой позиционной неопределенностью, ориентироваться в средах с высокой пространственной плотностью (скученностью), хотя и в условиях низкого разрешения и контрастной чувствительности (Stewart e.a., 2020).
Несмотря на такие очевидные различия в способах обработки зрительной информации в центральной и периферической областях, они могут быть успешно компенсированы в рамках механизма транссаккадической перекалибровки и реинтеграции последовательно совершенных фиксаций взгляда (Germeys et al., 2010). Другим возможным механизмом взаимодействия полей зрения при формировании стратегий осмотра изображений, в целом, и возвратных фиксаций, в частности, может являться экстраполяция в рамках обратной афферентации информации из центральной в периферическую область сетчатки (De Weerd, 2006; Gloriani, Schütz, 2019). Показано, что временная блокировка ретинотопических областей центрального поля зрения в головном мозге при помощи метода транскраниальной магнитной стимуляции приводила к достоверному снижению способности категоризации зрительных стимулов в периферическом поле (Chambers et al., 2013). Характер таких транссаккадических переносов был также широко изучен в рамках исследований зрительной рабочей памяти (Irwin et al., 1990) и механизмов избирательного внимания (Higgins, Rayner, 2014).
ЗАКЛЮЧЕНИЕ
В настоящем исследовании проанализированы известные факты об отражении системных функций зрения, таких как дискриминация и распознавание зрительных объектов, оценка семантического и эмоционального содержания изображений, решение поисковых задач, в фовеальном и периферическом поле зрения. Описан, разработанный авторами, новый нейроинформационный подход к исследованию взаимодействия фовеального и периферического зрения, включающий формализацию известных экспериментальных данных в области взаимодействия фовеального и периферического зрения в реалистичных моделях, анализ траекторий осмотра, областей интереса и возвратных фиксаций взгляда. Результаты компьютерного моделирования показали, что структура модельной траектории осмотра изображений коррелирует с вероятностью возвратных фиксаций “входного окна”. Модельное предположение о возможности количественной характеристики структуры траектории на основе возвратных фиксаций подтверждается результатами психофизических тестов, полученных нами ранее (Подладчикова и др., 2017; Podladchikova et al., 2009; Podladchikova et al., 2018; Podladchikova et al., 2020; Samarin et al., 2015), что позволяет рассматривать вероятность возвратных фиксаций как количественный критерий типа внимания (фокальное или пространственное) и моментов его переключения. На следующих этапах работы планируется применение нейроинформационного подхода и разработанной модели для оценки влияния структуры фовеального и периферического полей зрения и локальных признаков изображения на механизмы переключения типов внимания (фокальное и пространственное) в задачах межперсонного взаимодействия (Podladchikova et al., 2021a).
Список литературы
Барабанщиков В.А. Окуломоторные структуры восприятия. М.: Институт психологии РАН, 1997. 384 с.
Величковский Б.М. Когнитивная наука: основы психологии познания: в 2 т. М.: Академия, 2006. 405 с.
Иомдина Е.И., Селина О.М., Рожкова Г.И., Белокопытов А.В., Ершов Е.И. Контактная линза с имплантированным окклюдером как средство для оценки дальнего периферического зрения в естественных условиях. Сенсорные системы. 2020. Т. 34. № 2. С. 100–106. https://doi.org/10.31857/S0235009220020043
Подвигин Н.Ф., Макаров Ф.Н., Шелепин Ю.Е. Элементы структурно-функциональной организации зрительно-глазодвигательной системы. СПб.: Наука, 1986. 251 с.
Подладчикова Л.Н., Колтунова Т.И., Самарин А.И., Петрушан М.В., Шапошников Д.Г., Ломакина О.В. Современные представления о механизмах зрительного внимания. Ростов-на-Дону: Изд-во ЮФУ, 2017. 168 с.
Рожкова Г.И., Белокопытов А.В., Грачева М.А. Загадки слепой зоны и кольца повышенной плотности колбочек на крайней периферии сетчатки. Сенсорные системы. 2016. Т. 30. № 4. С. 263–281.
Рожкова Г.И., Белокопытов А.В., Иомдина Е.Н. Современные представления о специфике периферического зрения человека. Сенсорные системы. 2019. Т. 33. № 4. С. 306–332. https://doi.org/10.1134/S0235009219040073
Рожкова Г.И., Ярбус А.Л. Особенности восприятия человеком объектов в условиях работы ограниченных центральных участков сетчатки. Физиология человека. 1977. Т. 3. № 6. С. 1119–1127.
Самарин А.И., Подладчикова Л.Н., Петрушан М.В., Шапошников Д.Г., Гаврилей Ю.К. Алгоритмы активного пространственно-неоднородного зрения. Ростов-на-Дону: Изд-во ЮФУ, 2020. 104 с.
Хьюбел Д. Глаз, мозг, зрение. М.: Мир, 1990. 240 с.
Ярбус А.Л. Роль движений глаз в процессе зрения. М.: Наука, 1965. 166 с.
Ярбус А.Л., Рожкова Г.И. Особенности восприятия объектов на периферии поля зрения. Сенсорные системы. СПб., 1977. С. 64–73.
Anstis S.M. A chart demonstrating variations in acuity with retinal position. Vision Research. 1974. V. 14. № 7. P. 589–592. https://doi.org/10.1016/0042-6989(74)90049-2
Baddeley R.J., Tatler B.W. High frequency edges (but not contrast) predict where we fixate: A Bayesian system identification analysis. Vision research. 2006. V. 46. № 18. P. 2824–2833. https://doi.org/10.1016/j.visres.2006.02.024
Baveye Y., Dellandrea E., Chamaret C., & Chen L. LIRIS-ACCEDE: A Video Database for Affective Content Analysis. IEEE Transactions on Affective Computing. 2015. V. 6. № 1. P. 43–55. https://doi.org/10.1109/TAFFC.2015.2396531
Burnat K. Are visual peripheries forever young? Neural Plasticity. 2015. V. 15. Art. 307929. https://doi.org/10.1155/2015/307929
Cajar A. Eye-movement control during scene viewing: the roles of central and peripheral vision. PhD dissertation. Universität Potsdam, Humanwissenschaftliche Fakultät, Potsdam, 2016. 145 p.
Cajar A., Engbert R., Laubrock J. Spatial frequency processing in the central and peripheral visual field during scene viewing. Vision Research. 2016. V. 127. P. 186–197. https://doi.org/10.1016/j.visres.2016.05.008
Cajar A., Schneeweiß P., Engbert R., Laubrock J. Coupling of atten-tion and saccades when viewing scenes with central and peripheral degradation. Journal of Vision. 2016. V. 16. № 2. P. 8–12. https://doi.org/10.1167/16.2.8
Calvo M.G., Fernandez-Martin A., Nummenmaa L. Facial expression recognition in peripheral versus central vision: role of the eyes and the mouth. Psychological research. 2014. V. 78. № 2. P. 180–195. https://doi.org/10.1007/s00426-013-0492-x
Carrasco M. Visual attention: the past 25 years. Vision Research. 2011. V. 51. P. 1484–1525. https://doi.org/10.1016/j.visres.2011.04.012
Chambers C.D., Allen C.P.G., Maizey L., & Williams M.-A. (2013). Is delayed foveal feedback critical for extra-foveal perception? Cortex. 2013. V. 49. № 1. P. 327–335. https://doi.org/10.1016/j.cortex.2012.03.007
Cornelissen F.W., Bruin K.J., Kooijman C. Eye movements during search with artificial scotomas. Optometry and Vision Science. 2005. V. 82 №1. P. 27–35.
De Weerd P. Perceptual filling-in: more than the eye can see. Progress in Brain Research. 2006. V. 154. P. 227–245. https://doi.org/10.1016/s0079-6123(06)54012-9
Foulsham T., Kingstone A. Modelling the influence of central and peripheral information on saccade biases in gaze-contingent scene viewing. Visual Cognition. 2012. V. 20. № 4–5. P. 546–579. https://doi.org/10.1080/13506285.2012.680934
Germeys F., Graef P., Eccelpoel C., & Verfaillie K. The visual analog: Evidence for a preattentive representation across saccades. Journal of Vision. 2010. V. 10. № 10: 9. P. 1–28. https://doi.org/10.1167/10.10.9
Gloriani A.H., & Schütz A.C. Humans trust central vision more than peripheral vision even in the dark. Current Biology. 2019. V. 29. № 7. P. 1206–1210.e4. https://doi.org/10.1016/j.cub.2019.02.023
Hennig M., Wörgötter F. Eye micro-movements improve stimulus detection beyond the Nyquist limit in the peripheral retina. Advances in neural information processing systems. 2003. V. 16. P. 1475–1482.
Higgins E., & Rayner, K. Transsaccadic processing: stability, integration, and the potential role of remapping. Attention, Perception, & Psychophysics. 2014. V. 77. № 1. P. 3–27. https://doi.org/10.3758/s13414-014-0751-y
Hughes A.E., Southwell R.V., Gilchrist I.D., Tolhurst D.J. Quantifying peripheral and foveal perceived differences in natural image patches to predict visual search performance. Journal of vision. 2016. V. 16. № 10. P. 18–19. https://doi.org/10.1167/16.10.18
Irwin D., Zacks J., & Brown J. Visual memory and the perception of a stable visual environment. Perception & Psychophysics. 1990. V. 47. № 1. P. 35–46. https://doi.org/10.3758/BF03208162
Kragic D., Bjorkman M. Strategies for object manipulation using foveal and peripheral vision. Fourth IEEE Intern. Conf. Comp. Vision Systems. 2006. P. 50–52.
Kuraguchi K., Ashida H. Beauty and cuteness in peripheral vision. Frontiers in psychology. 2015. V. 6. P. 566–569. https://doi.org/10.3389/fpsyg.2015.00566
Larson A.M., Loschky L.C. The contributions of central versus peripheral vision to scene gist recognition. Journal of Vision. 2009. V. 9. № 10. P. 1–16. https://doi.org/10.1167/9.10.6
Laubrock J., Cajar A., Engbert R. Control of fixation duration during scene viewing by interaction of foveal and peripheral processing. Journal of Vision. 2013. V. 13. № 12. P. 11–15. https://doi.org/10.1167/13.12.11
Lomakina O., Podladchikova L., Shaposhnikov D., Koltunova T. Spatial and temporal parameters of eye movements during viewing of affective images. Advances in Intelligent Systems and Computing. 2016. V. 449. P. 127–133. https://doi.org/10.1007/978-3-319-32554-5_17
Levi D.M. Crowding–An essential bottleneck for object recognition: A mini-review. Vision research. 2008. V. 48. № 5. P. 635–654. https://doi.org/10.1016/j.visres.2007.12.009
Loschky L.C., Nuthmann A., Fortenbaugh F.C., Levi D.M. Scene perception from central to peripheral vision. Journal of vision. 2017. V. 17. № 1. P. 6–8. https://doi.org/10.1167/17.1.6
Loschky L.C., Szaffarczyk S., Beugnet C., Young M.E., Boucart M. The contributions of central and peripheral vision to scene-gist recognition with a 180° visual field. Journal of Vision. 2019. V. 15. № 12. P. 1–21. https://doi.org/10.1167/19.5.15
Ludwig C.J.H., Davies J.R., Eckstein M.P. Foveal analysis and peripheral selection during active visual sampling. Proceedings of the National Academy of Sciences. 2014. V. 111. № 2. P. E291–E299. https://doi.org/10.1073/pnas.1313553111
McCamy M.B., Otero-Millan J., Macknik S.L., Yang Y., Troncoso X.G., Baer S.M., Crook S.M., Martinez-Conde S. Microsaccadic efficacy and contribution to foveal and peripheral vision. J. Neurosci. 2012. V. 32. № 27. P. 9194–9204. https://doi.org/10.1523/JNEUROSCI.0515-12.2012
Parkhurst D.J., Niebur E. Variable-resolution displays: A theoretical, practical, and behavioral evaluation. Human Factors. 2002. V. 44. № 4. P. 611–615.
Podladchikova L., Shaposhnikov D., Koltunova T., Lazurenko D., Kiroy V. Spatial and temporal properties of gaze return fixations while viewing affective images. Advances in Applied Physiology. 2020. V. 5. № 2. P. 42–47. https://doi.org/10.11648/j.aap.20200502.16
Podladchikova L.N., Shaposhnikov D.G., Kozubenko E.A. Towards neuroinformatic approach for second-person neuroscience. Stud.Computat. Intelligence. 2021a. V. 925. P. 143–148. https://doi.org/10.1007/978-3-030-60577-3_16
Shaposhnikov D., Podladchikova L., Lazurenko D., Kiroy V., Search for guantitative prameters of scan path of image viewing by biologically motivated model. Advances in Applied Physiology. 2021b. V. 6. № 1. P. 9–13. https://doi.org/10.11648/j.aap.20210601.12
Podladchikova L.N., Shaposhnikov D.G., Koltunova T.I. Spatial and temporal properties of gaze return fixations while viewing affective images. Russian Journal of Physiology. 2018. V. 104. № 2. P. 245–254 (in Russian). https://rusjphysiol.org/index.php/rusjphysiol/article/view/242/44
Podladchikova L.N., Shaposhnikov D.G., Tikidgji-Hamburyan A.V., Koltunova T.I., Tikidgji-Hamburyan R.A., Gusakova V.I., Golovan A.V. Model-based approach to sudy of mechanisms of complex image viewing. Optical Memory and Neural Networks. 2009. V. 18. № 2. P. 114–121. https://doi.org/10.3103/S1060992X09020088
Pointer J., Hess R. The contrast sensitivity gradient across the human visual field: with emphasis on the low spatial frequency range. Vision Research. 1989. V. 29. № 9. P. 1133–1151. https://doi.org/10.1016/0042-6989(89)90061-8
Privitera C.M., Stark L.W. Scanpath theory, attention, and image processing algorithms for predicting human eye fixations. Neurobiol. Attention. 2005. P. 296–299.
Reingold E.M., Reichle E.D., Glaholt M.G., Sheridan H. Direct lexical control of eye movements in reading: Evidence from a survival analysis of fixation durations. Cognitive psychology. 2012. V. 65. № 2. P. 177–206. https://doi.org/10.1016/j.cogpsych.2012.03.001
Ringer R.V., Throneburg Z., Johnson A.P., Kramer A.F., Loschky L.C. Impairing the useful field of view in natural scenes: Tunnel vision versus general interference. Journal of Vision. 2016. V. 16. № 2. P. 7–9. https://doi.org/10.1167/16.2.7
Rosenholtz R. Capabilities and limitations of peripheral vision. Annual Review of Vision Science. 2016. V. 2. P. 437–457. https://doi.org/10.1146/annurev-vision-082114-035733
Ryu D., Abernethy B., Mann D.L., Poolton J.M., Gorman A.D. The role of central and peripheral vision in expert decision making. Perception. 2013. V. 42. P. 591–607. https://doi.org/10.1068/p7487
Samarin A., Koltunova T., Osinov V., Shaposhnikov D., Podladchikova L. Scanpaths of complex image viewing: insights from experimental and modeling studies. Perception. 2015. V. 44. № 8–9. P. 1064–1076. https://doi.org/10.1177/0301006615596872
Samarin A.I., Podladchikova L.N., Petrushan M.V., Shaposhnikov D.G. Active vision: from theory to application. Optical Memory and Neural Networks. 2019. V. 28. № 3. P. 185–191. https://doi.org/10.3103/S1060992X19030068
Samarin A., Koltunova T., Osinov V., Shaposhnikov D., Podladchikova L. Scanpaths of complex image viewing: insights from experimental and modeling studies. Perception. 2015. V. 44. № 8–9. P. 1064–1076. https://doi.org/10.1177/0301006615596872
Shaposhnikov D.G., Gizatdinova Yu.F., Podladchikova L.N. The peculiarities of the visual perception in the peripheral vision field. Pattern Recognition and Image Analysis. 2001. V. 11. № 2. P. 376–378.
Stephenson C.M., Knapp A.J., Braddick O.J. Discrimination of spatial phase shows a qualitative difference between foveal and peripheral processing. Vision Research. 1991. V. 31. № 7–8. P. 1315–1326. https://doi.org/10.1016/0042-6989(91)90053-8
Stewart E.E.M., Valsecchi M., Schütz A.C. A review of interactions between peripheral and foveal vision. Journal of Vision. 2020. V. 20. № 12: 2. P. 1–35. https://doi.org/10.1167/jov.20.12.2
Strasburger H., Rentschler I., Jüttner M. Peripheral vision and pattern recognition: A review. Journal of Vision. 2011. V. 11. № 5. P. 13. https://doi.org/10.1167/11.5.13
Tanrıkulu O.D., Chetverikov A., Kristjansson A. Encoding perceptual ensembles during visual search in peripheral vision. J. Vision. 2020. V. 20. № 8. P. 20–22. https://doi.org/10.1167/jov.20.8.20
To M.P.S., Gilchrist I.D., Troscianko T., Tolhurst D.J. Discrimination of natural scenes in central and peripheral vision. Vision Research. 2011. V. 51. № 14. P. 1686–1698. https://doi.org/10.1016/j.visres.2011.05.010
Torralba A., Oliva A., Castelhano M.S., Henderson J.M. Contextual guidance of eye movements and attention in real-world scenes: the role of global features in object search. Psychological review. 2006. V. 113. № 4. P. 766–769. https://doi.org/10.1037/0033-295X.113.4.766
Trukenbrod H.A., & Engbert R. ICAT: A computational model for the adaptive control of fixation durations. Psychonomic bulletin & review. 2014. V. 21. № 4. P. 907–934. https://doi.org/10.3758/s13423-013-0575-0
Van Diepen P., d’Ydewalle G. Early peripheral and foveal processing in fixations during scene perception. Visual Cognition. 2003. V. 10. № 1. P. 79–100. https://doi.org/10.1080/713756668
Wiecek E.W., Pasquale L.R., Fiser J., Dakin S., Bex P.J. Effects of peripheral visual field loss on eye movements during visual search. Frontiers in Psychology. 2012. V. 3. P. 472–475. https://doi.org/10.3389/fpsyg.2012.00472
Yu Q., Shim W.M. Modulating foveal representation can influence visual discrimination in the periphery. J.Vision. 2016. V. 16. № 3. P. 15–19. https://doi.org/10.1167/16.3.15
Дополнительные материалы отсутствуют.
Инструменты
Сенсорные системы