

Сенсорные системы, 2022, T. 36, № 2, стр. 109-123
Компонент N2pc в исследованиях зрительного внимания
А. С. Столетний 1, *, Д. С. Алексеева 1, В. В. Бабенко 1, П. В. Анохина 1, Д. В. Явна 1
1 Федеральное государственное автономное образовательное учреждение высшего образования “Южный федеральный университет”
344006 г. Ростов-на-Дону, ул. Большая Садовая, 105/42, Россия
* E-mail: asstoletniy@sfedu.ru
Поступила в редакцию 05.07.2021
После доработки 21.01.2022
Принята к публикации 01.02.2022
- EDN: CGQGIX
- DOI: 10.31857/S0235009222020068
Аннотация
В статье представлен обзор литературы, касающейся природы и свойств негативного отклонения связанных с событием потенциалов N2pc. Как правило, этот латерализованный компонент возникает спустя 180–300 мс после начала предъявления зрительного стимула и вычисляется как разница между мозговыми ответами, регистрируемыми в контралатеральных парието-окципитальных отведениях обоих полушарий. На сегодняшний день N2pc служит надежным индикатором вовлечения когнитивных ресурсов мозга в процесс идентификации местоположения цели. Изначально предполагалось, что N2pc отражает процесс подавления влияния нерелевантных стимулов в ходе восприятия, но впоследствии возникли и иные точки зрения на его функциональную роль, поскольку появились данные о его связи с процессами, обеспечивающими не только фильтрацию дистракторов, но и собственно обработку целевого стимула. Кроме того, особенности возникновения данного компонента позволяют исследовать механизмы развертывания внимания как у людей, так и у животных. N2pc нашел свое применение и в изучении внимания у людей с различными психическими и физическими расстройствами.
ВВЕДЕНИЕ
Метод связанных с событием потенциалов (ССП) традиционно используется в экспериментах, посвященных изучению когнитивных процессов и их электрографических коррелятов. Амплитуда и латентность данных волн или компонентов ССП выступают в качестве индикаторов когнитивных процессов, протекающих с момента подачи стимула до реакции на него, если таковая предусмотрена.
С точки зрения анализа ССП группа волн, отражающая ментальные операции, связанные с вниманием, характеризуется негативностью отклонения от изолинии и пиковой латентностью от 200 до 350 мс. Совокупность таких волн получила название компонент N2. Данную группу негативных отклонений потенциала в свою очередь можно разделить на подгруппы, имеющие различное функциональное значение – это субкомпоненты N2a, N2b, N2c и N2pc (Luck, Kappenman, 2012). Каждый из этих компонентов является инструментом исследования внимания (Luck 2012; Luck, Kappenman, 2012; Pazo-Alvarez et al., 2003; Пономарев 2016). В настоящей статье предметом анализа является такой феномен, как N2pc, представляющий собой часть задней группы компонентов N2c. Первоначально предполагалось, что этот компонент отражает процесс фильтрации, который запускается для подавления входных сигналов от иррелевантных объектов (дистракторов), окружающих потенциальную цель (Luck, Hillyard, 1994). Однако позднее было показано, что он может выступать индикатором не только процесса фильтрации дистракторов как такового, но целого комплекса операций, одна часть которых отвечает за подавление дистракторов, другая – за обработку целевого стимула (Hickey et al., 2009). В некотором роде это небольшое изменение в трактовке функциональной роли компонента N2pc позволило переосмыслить данные прошлых лет: N2pc стал считаться не только коррелятом восходящих информационных, но и нисходящих управляющих процессов, а также их взаимодействия. Кроме того, были выявлены и другие латерализованные компоненты ССП, такие как Pd – позитивность, вызванная дистрактором (Hickey et al., 2009; Sawaki, Luck, 2010), Ptc – поздний позитивный компонент (Hilimire et al., 2009), пост-N2pc компонент (PNP) (Papaioannou, Luck, 2020), SPCN – устойчивая задняя контралатеральная негативность (Robitaille, Jolicoeur, 2006).
ОБНАРУЖЕНИЕ КОМПОНЕНТА N2PC
Первой работой, в которой выявлен и проанализирован тогда еще не выделенный как отдельное явление компонент N2pc, была статья Лака и Хилларда (Luck, Hillyard, 1990), где авторы исследовали фокусировку внимания во время параллельного и последовательного зрительного поиска (visual search task) при помощи ССП. Когда участники эксперимента должны были обнаружить среди треугольников-дистракторов стрелку-цель, которая появлялась либо в правой, либо в левой части экрана, поиск был параллельным. В данном случае горизонтальная линия, образующая стержень стрелки, явно выделялась на экране, и испытуемые могли быстро и легко обнаружить цель. Когда цель была треугольником, а стрелки – дистракторами, поиск становился последовательным. В этом случае цель не содержала выразительной отличительной черты, значительно отличавшей ее от дистракторов, поэтому для нахождения цели испытуемым необходимо было выполнять медленный и тщательный поиск по массиву стимулов. В обеих задачах наблюдалось нарастание негативности потенциала в полушарии, контралатеральном относительно полуполя зрения, в котором находился целевой стимул. При параллельном поиске данная волна возникала в районе 200 мс и затухала уже к 300 мс с момента подачи целевого стимула, а при последовательном – появлялась примерно через 200 мс и имела длительность фактически до конца эпохи анализа (Luck, Kappenman, 2012).
В своей первой статье Лак и Хиллард дали название обнаруженному ими ранее компоненту – N2pc (Luck, Hillyard, 1994). Буква “N” обозначает отрицательную полярность, цифра “2” описывает его латентность в ряду компонентов ССП, а “рс” (сокращение от “posterior contralateral”) указывает на пространственную локализацию, так как негативность формируется в теменно-затылочных и окципитальных отведениях и более выражена в контралатеральном полушарии.
ХАРАКТЕРИСТИКА И ОСОБЕННОСТИ РЕГИСТРАЦИИ N2PC
Как уже было сказано выше, N2pc является одной из нескольких форм мозгового ответа на латерализованные зрительные целевые стимулы, т.е. находящиеся либо в левом, либо в правом полуполе зрения. Цель в свою очередь может задаваться цветом, ориентацией, размером, различной формой, буквой, словом, фигурами Канизса, выражением лица, направлением взора, движущимся объектом (Luck, 2012). Следует отметить, что целевые стимулы, расположенные в центре поля зрения, латерализованной активности в виде N2pc не вызывают (Hickey et al., 2009).
В типичной экспериментальной парадигме исследования внимания при помощи N2pc от испытуемых требуется искать на экране монитора целевой стимул среди отвлекающих, причем цель должна располагаться либо в левой, либо в правой части экрана (рис. 1), в то время как дистракторы могут находиться в том числе и на вертикальной средней линии дисплея (Hickey et al., 2009). Цель, как правило, должна существенно выделяться и быть легко заметной в массиве стимулов (“pop-out target”) (Luck, 2006). При обнаружении цели испытуемых обычно просят сигнализировать об этом нажатием на соответствующую клавишу; регламентация таких действий зависит от конкретной задачи, поставленной в эксперименте – например, можно связать обнаружение цели с простым нажатием на кнопку, или для целей с левой стороны определить левую кнопку мыши, а для целей справа – правую, и т.д. В инструкции испытуемых просят реагировать на стимулы как можно быстрее. Массивы стимулов предъявляются блоками, причем время экспозиции одного массива может колебаться от 200 мс до 1.5 с в зависимости от задачи (Brisson, Jolicœur, 2007; Girelli, Luck, 1997; Luck, Hillyard, 1994; Reutter et al., 2017).
Рис. 1.
Пример типичного экспериментального дисплея в N2pc-парадигме, содержащего массив стимулов с выделяющейся (pop-out) целью – белым прямоугольником, расположенным среди черных прямоугольников дистракторов.
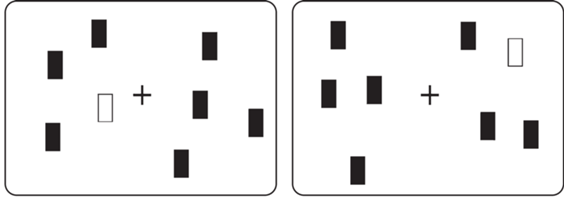
В центре экрана помещается точка фиксации, которая находится там постоянно в ходе предъявления стимулов, причем во многих экспериментах испытуемому давали указание фиксировать на ней свой взгляд как во время предъявления массива, так и при осуществлении зрительного поиска (Brisson et al., 2007). Как правило, в экспериментах с N2pc, помимо регистрации ЭЭГ, измеряется среднее время реакции участников на правильно распознанные целевые стимулы (Yuan et al., 2020).
Для обнаружения данного компонента сначала можно рассчитать контралатеральную волну (усреднение мозговых ответов на стимул из отведений правого полушария, когда он находится в левом полуполе зрения, и отведений левого полушария, когда стимул в правом полуполе) и ипсилатеральную волну (усреднение мозговых ответов на стимул из отведений правого полушария, когда стимул находится в правом полуполе зрения, и отведений левого полушария, когда стимул в левом полуполе). Получившееся в результате ССП анализируют двумя дополняющими друг друга путями – статистически и визуально сравнивают контралатеральный и ипсилатеральный ССП, и, вычитая второй из первого, получают разностный потенциал (Difference wave, Д-волна). На рис. 2 наглядно показаны два вида представления результатов для подобных расчетов: первый – когда на одном графике приводятся контра- и ипсилатеральные волны для отдельного экспериментального условия (go, nogo); второй – отображение разностных потенциалов, полученных с помощью описанного выше алгоритма для двух или более условий эксперимента (contra-ipsi).
Рис. 2.
Усредненные ССП из исследования (Barras, Kerzel, 2016), демонстрирующие вызванные ответы на контралатеральные (черная сплошная линия) и ипсилатеральные (прерывистая серая линия) стимулы в эксперименте с парадигмой go/nogo, в отведениях Po7 и Po8, и представление этих же результатов в виде разностных волн (зеленая go, красная – nogo).
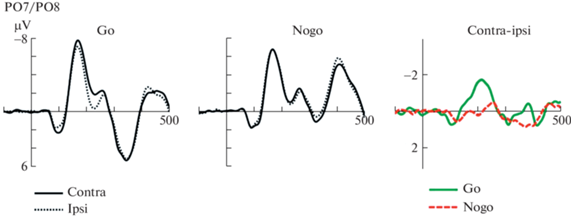
Первый тип представления позволяет оценить волновую форму и амплитуду компонентов ССП, в то время как второй хорошо подходит для анализа времени возникновения и завершения N2pc, а также его величины в случае, когда контра- и ипсилатеральные волны накладываются друг на друга. Построение разностных потенциалов также позволяет защитить N2pc от эффекта наложения других компонентов. При публикации результатов исследователи могут приводить ССП как из единичных симметричных отведений (рис. 2), так и усредняя волны по нескольким отведениям одного полушария (Brisson et al., 2007).
При записи ЭЭГ в экспериментах с латерализованной зрительной целью компонент N2pc может регистрироваться в отведениях задних отделов полушарий – P3, P4, Po3, Po4, P7, P8, Po7, Po8, среди которых наиболее информативными в плане амплитуды этого потенциала являются Po7 и Po8. Латентность возникновения негативной волны может колебаться от 180 до 300 мс, а продолжительность в некоторых случаях может превышать 200 мс (Luck, 2012). Амплитуда N2pc, как правило, не превышает 2 мкв.
Стоит отметить, что в работах некоторых авторов термин N2pc заменяется иным – Posterior Contralateral Negativity (PCN), очевидно, чтобы избежать ошибочных ассоциаций с N2 (Töllner et al., 2011).
ФУНКЦИОНАЛЬНАЯ РОЛЬ КОМПОНЕНТА N2PC
Чаще всего парадигма N2pc используется для исследования фокусировки внимания в условиях зрительного поиска заданной цели в массиве из множества изображений объектов. При этом считается, что момент возникновения потенциала соответствует времени, когда мозг начинает вычислять местоположение цели (Luck, 2012). Например, если N2pc начинается на латентности 200 мс, можно сделать вывод, что внимание было перемещено на цель на двухсотой миллисекунде после ее появления на экране. Однако отсутствие искомого компонента рассматривается как свидетельство того, что сдвига внимания не произошло (Ansorge et al., 2011; Burra et al., 2016; Burra, Kerzel, 2013; Eimer, Kiss 2008; Hilimire, Corballis 2014; Ikeda et al., 2013; Kiss et al., 2008; Lien et al., 2011; X. Liu et al., 2017; Laura Lorenzo-López et al., 2008; Woodman, Luck, 1999). Таким образом, можно сказать, что N2pc является своеобразным индикатором динамики фокусировки внимания (Kiss et al., 2009; Laura Lorenzo-López et al., 2008). Безусловно, раннее смещение внимания к цели может быть модулировано работой не только вентрального зрительного пути, на поздних этапах которого, по всей видимости, и формируется N2pc, но и иных мозговых систем. Так, в работе (Cohen et al., 2009) на обезьянах было показано, что еще до возникновения N2pc некая селективная обработка производится в передних глазодвигательных областях (восьмое поле по Бродману). Таким образом, возможно, что компонент N2pc формируется не на самом раннем этапе фокусировки внимания на потенциальной цели в принципе, но отражает самый ранний момент, когда обработка местоположения цели начинается в вентральном зрительном пути.
На то, какие процессы лежат в основе феномена N2pc, в литературе существует несколько точек зрения. Наиболее общепринятой считается та, которая гласит, что N2pc вероятнее всего отражает сосредоточение внимания на потенциальной цели и фильтрацию окружающих элементов – дистракторов (Luck, Hillyard, 1994; Luck, 2012). Также существует гипотеза о приоритете процессов выбора и обработки цели над фильтрацией (Eimer 1996; Eimer, Kiss, 2008; Mazza et al., 2009a), и гипотеза, объединяющая в себе обе вышеозначенные точки зрения (Hickey et al., 2009). Рассмотрим каждую из них подробнее.
В 1994 г. Лак и Хиллард предположили, что волна N2pc отражает процесс фильтрации, подавляющей обработку окружающих цель дистракторов с целью уменьшения их влияния на идентификацию целевого объекта, сформулировав таким образом гипотезу подавления дистракторов (distractor-suppression hypothesis) (Luck, Hillyard, 1994b). В качестве доказательства своей точки зрения авторы провели серию экспериментов, в которых манипулировали характеристиками стимульного материала, что в свою очередь должно было влиять на упомянутый выше процесс. Полученные результаты позволили им сформулировать четыре основных аргумента в пользу того, что компонент N2pc непосредственно отражает процесс зрительной пространственной фильтрации. Во-первых, появление волны N2pc зависит от того, распознается ли объект как потенциальная цель, а не от того, является ли он целью на самом деле – например, в упомянутых выше экспериментах N2pc наблюдался как для целей (target), так и для нецелевых стимулов, очень похожих на цель (difficult non-target). Во-вторых, волна N2pc сама по себе не отражает процесс идентификации как таковой – компонент проявляется только тогда, когда цель представлена на дисплее вместе с дистракторами. В-третьих, N2pc возникает, когда необходимо распознать цель среди таких дистракторов, которые конкурируют за внимание испытуемого – если дистракторы в массиве стимулов идентичны заданной цели, N2pc не регистрируется, т.е. его роль шире, чем просто углубленная обработка признаков объектов. В-четвертых, N2pc генерируется, когда нерелевантная информация от окружающих (отвлекающих объектов) может помешать идентификации цели. Также стоит добавить, что при росте количества дистракторов происходит увеличение амплитуды N2pc (Luck et al., 1997; Mazza et al., 2009b). На основе приведенных выше аргументов Стивен Лак предположил, что избирательность внимания работает в ключе уменьшения или устранения влияния неподходящих по инструкции стимулов, оставляя подходящие доступными для обработки. Таким образом, амплитуда N2pc отражает величину потребности в таком уменьшении (подавлении), что происходит, например, при увеличении количества дистракторов или их сходства с целью. Следовательно, чем больше вышеозначенная потребность, тем выше амплитуда N2pc. Очевидная связь между селективностью и компонентом N2pc привела к созданию теории, в которой подавление дистракторов играет ключевую роль в избирательности внимания – “теория разрешения неопределенности” (“ambiguity resolution theory”) (Luck et al., 1997). Согласно этой теории, внимание подавляет неопределенность в привязке признаков к тому или иному стимулу путем подавления информации от стимулов, на которых не было сфокусировано внимание, а N2pc отражает работу данного подавляющего процесса.
Вторая гипотеза предполагает, что N2pc отражает процессы выбора цели (target-selection hypothesis), когда зрительное внимание усиливает обработку целевых или подобных целям стимулов в поле зрения, т.е. в процесс селективного внимания вовлечены и другие механизмы помимо подавления дистракторов. В своей статье 1996 г. Эймер представил данные о том, что компонент N2pc проявляется не только в типичных задачах зрительного поиска, но также в ситуациях, когда цель представлена вместе с одним дистрактором, и они расположены в противоположных полуполях зрения. Таким образом, N2pc как коррелят работы механизма избирательности внимания генерируется не только в случае, когда необходимо обнаружить и идентифицировать цель среди множества смежных дистракторов, но и тогда, когда необходимость отфильтровывать нерелевантную или противоречивую информацию сводится к минимуму. Эймер пришел к выводу, что N2pc в первую очередь отражает операцию выбора цели, которая не зависит от количества и расположения дистракторов, а не процесс подавления нерелевантной информации (Eimer, 1996). Подобный вывод можно сделать и из работы (Shedden, Nordgaard, 2001), в которой N2pc наблюдался в отведениях контралатерального полушария для одиночного латерализованного стереоскопического стимула при отсутствии каких-либо дистракторов. В этом случае амплитуда N2pc будет отражать селективность внимания относительно релевантных задаче стимулов.
В своей работе Хики и соавт. приводят результаты экспериментов с ССП, целью которых было выяснить, отражает ли N2pc один или несколько механизмов внимания, и тем самым попытаться объединить представления об N2pc, имевшиеся на тот момент (Hickey et al., 2009). В ходе исследования зрительного поиска при помощи разных конфигураций стимульного материла авторам удалось выделить два независимых компонента ССП, которые они назвали Nt и Pd. Помимо того, что эти компоненты появлялись при предъявлении массивов с разным расположением стимулов, они отличались своей полярностью и корковой локализацией. Несмотря на то что оба компонента регистрируются в задних отделах коры, Pd возникает в более медиальных и дорсальных областях, в то время как Nt – в более латеральных и вентральных. Хики и соавт. связали этот факт с гипотезой о том, что Pd и Nt являются маркерами процессов обработки информации в дорсальном и вентральном путях зрительной обработки (Milner, Goodale, 2006). Pd, по-видимому, привязан к позиции дистрактора, а Nt, напротив, связан с местоположением цели. Вместе оба компонента отражают два различных процесса, участвующих в зрительном поиске: один, пространственный по своей природе, ответственен за обработку дистракторов, другой же связан с характеристиками объектов и принимает участие в обработке целевых стимулов. На основе этих фактов Хики и соавт. сделали вывод, что N2pc представляет собой не единую волну, а суперпозицию двух независимых компонентов ССП, указанных выше. Тем самым авторы показали, что селективность внимания работает не как отдельный процесс подавления дистракторов, а как совокупность по крайней мере двух процессов – обработки цели и подавления дистракторов, и N2pc является отражением такой совокупной работы.
Как видно из представленной выше информации, природа компонента N2pc в некоторой степени остается предметом дискуссии среди исследователей, хотя большинство авторов признают, что N2pc является надежным нейронным маркером выбора цели. В дополнение к этому стоит упомянуть некоторые работы, которые не вполне согласуются с изложенными выше теоретическими представлениями.
Значительный интерес представляет статья Теувеса, посвященная восходящим и нисходящим механизмам управления вниманием, в которой он подчеркивает, что выбор того или иного визуального объекта определяется исключительно восходящими (bottom-up) механизмами, которые не зависят от намерений наблюдателя. Этот автор предполагает, что выбор целей внимания определяется картами салиентности (salience maps), которые вычисляются на этапе параллельной зрительной обработки, и что данный процесс не может быть прерван или изменен какими-либо нисходящими (top-down) воздействиями. На основе указанных аргументов и ряда экспериментальных данных, автор оспаривает представление о N2pc как о маркере процессов отбора информации и отождествляет его с аттентивной обработкой зрительных признаков объекта, которая имеет место уже после отбора стимула (Theeuwes, 2010).
В исследовании (Zivony et al., 2018) ставится под сомнение предположение, что отсутствие N2pc является свидетельством отсутствия сдвига внимания. В сложном эксперименте использовался эффект мерцания внимания при демонстрации стимулов с временными интервалами в 300 и 800 мс. Было показано, что мерцание внимания не повлияло на сам факт сдвига внимания, но подавило связанный с ним N2pc. На этом основании Зивоны и соавт. предположили, что N2pc отражает не смещение внимания как таковое, а временное улучшение обработки (transient enhancement of processing), которое происходит в пространственном фокусе внимания и способствует дальнейшей, уже высокоуровневой обработке. Исследователи посчитали, что такой вывод требует переосмысления результатов, приведенных в литературе по захвату внимания, где N2pc использовался как индикатор селективности внимания.
Безусловно, говоря о компоненте N2pc, следует упомянуть и о теориях захвата внимания, в исследованиях которого широко применяется этот компонент. Частично они уже упоминались выше, когда речь шла о процессах, лежащих в основе N2pc. Традиционно выделяют две группы таких теорий (Gaspelin, Luck, 2019). Первая – это теории восходящего управления вниманием (bottom-up), которые предполагают, что салиентные стимулы привлекают внимание автоматически и независимо от наших знаний и задач. Вторая – это теории нисходящего управления вниманием (top-down). Такие теории предполагают, что салиентные элементы не оказывают особого влияния на распределение внимания, если только они не соответствуют ожидаемым признакам цели поиска или предыдущему опыту. Спор между этими двумя теоретическими позициями продолжается уже несколько десятилетий, причем обе теории подтверждаются многочисленными исследованиями, что и привело в итоге к теоретическому тупику (Gaspelin, Luck, 2019). В недавних работах были представлены экспериментальные доказательства того, что восходящий захват внимания, вызванный салиентным стимулом, может подавляться при помощи нисходящих ингибирующих механизмов. Важно подчеркнуть, что такое подавление происходит до первоначального переключения внимания и является активным процессом с заранее заданными характеристиками признаков, которые следует игнорировать при выборе цели среди других объектов. Такие данные позволили в некоторый степени объединить описанные выше теории в новую – “гипотезу подавления” (signal suppression hypothesis), наладив эмпирическую связь между двумя описанными выше теоретическими подходами. В работах (Gaspelin, Luck, 2019; Luck et al., 2021) приведена подробная аргументация состоятельности “гипотезы подавления” на основе данных исследований из области психофизики, окулографии и электрофизиологии, в том числе N2pc и Pd.
С точки зрения функциональной роли N2pc нельзя не упомянуть ряд исследований, посвященных взаимодействию этого компонента и зрительной рабочей памяти. Очевидно, что память играет важную роль в распределении внимания, предоставляя зрительной системе ограниченный по времени доступ для сохранения недавно обнаруженной информации. При этом объекты конкурируют в пределах зрительной сцены, чтобы в дальнейшем “получить доступ” к более высокому уровню обработки. В ходе этой конкуренции рабочая память может инициировать сдвиг внимания в пользу тех элементов, что ранее были сохранены в памяти (Hollingworth, Luck, 2009).
Так, в работе (Kumar et al., 2009) было выдвинуто предположение, что содержание рабочей памяти может определять лидера в борьбе за захват внимания, и результат этой конкуренции отражается в параметрах N2pc. По результатам своей работы Эстл и соавт. пришли к выводу, что информация, хранящаяся в памяти, оставляет след, который содержит первоначальное пространственное положение элементов в массиве стимулов, и что этот след сдвигает внимание автоматически всякий раз, когда находится соответствие между элементом в сохраненном массиве стимулов и объектом, который предъявляется испытуемому в данный момент (Astle et al., 2009). Также N2pc наблюдался в похожих экспериментах, где цель определялась различием между представленным массивом стимулов и массивом, ранее сохраненным в рабочей памяти (Eimer, Mazza, 2005; Hyun et al., 2009; Schankin, Wascher, 2008). В статье Крзан было показано, что активное поддержание образа объекта в памяти вызывает автоматическое смещение внимания к целевому стимулу, который совпадает с хранящимся образом, что в свою очередь вызывает N2pc (Krzan, 2015). Индивидуальные отличия в зрительной и пространственной рабочей памяти исследовались в статье Куперуса и соавт. Было показано, что больший объем пространственной рабочей памяти напрямую связан с большей точностью выполнения зрительного поиска и выражается в большей амплитуде N2pc (Couperus et al., 2021). Похожие данные приводятся и в работе (Heuer, Schubö, 2016).
Приведенные выше результаты исследований свидетельствуют о возможности применения компонента N2pc в изучении как зрительного пространственного внимания, так и зрительной рабочей памяти. Необходимо тщательно продумывать дизайн эксперимента в данной парадигме, учитывая, что в N2pc может отражаться работа нескольких механизмов обработки информации.
ВЛИЯНИЕ РАЗЛИЧНЫХ ФАКТОРОВ НА АМПЛИТУДНЫЕ И ВРЕМЕННЫЕ ПАРАМЕТРЫ N2PC
Как уже указывалось выше, компонент N2pc может иметь как относительно короткую, в 100–300 мс, так и существенно большую продолжительность, вплоть до конца эпохи анализа ССП. По всей видимости, продолжительный N2pc является следствием применения последовательного зрительного поиска в ходе эксперимента, в котором цель не имела выраженного отличия от дистракторов, и испытуемым приходилось долго и тщательно вести поиск по всему массиву стимулов, чтобы найти цель. Однако в задачах, где цель заметно отличалась от нецелей, участники эксперимента быстро и без затруднений отыскивали цель. Длительность N2pc является индикатором того, какой тип зрительного поиска используется для выполнения задачи – параллельный (короткий N2pc) или последовательный (длинный N2pc) (Luck, Hillyard, 1990; Wolber, Wascher, 2003).
Стоит упомянуть влияние свойств самих стимулов на N2pc. Бриссон и соавт. провели серию экспериментов, в которых исследовали влияние длительности (Brisson, Jolicœur, 2007) и яркости (Brisson et al., 2007) стимулов на характеристики компонента N2pc. В первом случае испытуемым предлагалось отслеживать целевые стимулы заданного цвета (например, целью могли выступать красные цифры), которые в случайном порядке появлялись вместе с похожими на них дистракторами (зелеными цифрами, красными буквами) на правой и левой половинах экрана монитора. Длительность предъявления стимулов составляла 50, 200 и 350 мс для разных блоков стимулов. Анализ результатов опыта показал, что большая длительность стимула (350 мс) отразилась в снижении амплитуды N2pc по сравнению с более короткими предъявлениями. Значения амплитуды для стимулов длительностью 50 и 200 мс оставались одинаковыми. Кроме того, были представлены методические рекомендации для длительности стимулов в подобных опытах (Brisson, Jolicœur, 2007).
В другом исследовании (Brisson et al., 2007) авторы манипулировали яркостью зрительных стимулов. Целевые стимулы представляли собой появляющиеся в левой и правой части экрана монитора квадраты зеленого и красного цветов, построенные из линий малой, средней или большой ширины, при этом у каждого квадрата имелся разрыв на одной из сторон. Задачей испытуемых было определить, с какой из сторон целевого (красного или зеленого, в зависимости от заданной инструкции) стимула был разрыв, нажав соответствующую клавишу на клавиатуре. Анализ результатов показал, что несмотря на рост амплитуды P1 в ответ на утолщение линий представленных стимулов, значимого изменения амплитуды N2pc не наблюдалось. Однако компонент N2pc в ответ на стимулы с большей толщиной линий появляется значительно раньше, чем на стимулы с малой и средней. Также они отметили, что небольшое изменение амплитуды компонента P1 серьезно повлияло на латентность N2pc, демонстрируя низкую эффективность стимулов с тонкими линиями в задачах, связанных с избирательностью внимания. По всей видимости, именно интенсивность стимула в значительной мере определяет тот момент, когда внимание направляется к целевой зоне (Brisson et al., 2007).
Разумеется, на проявление N2pc в ходе зрительного поиска влияет салиентность цели. Так, например, в статье, где исследовалось зрительное внимание при шизофрении, было показано, что и у экспериментальной, и у контрольной групп N2pc формировался примерно на 50 мс ранее на красный целевой стимул, чем на зеленый (Luck et al., 2006). В работе Толлнера и соавт. изучалось влияние салиентности на скорость преаттентивной обработки при восприятии стимулов низкой, средней и высокой салиентности. В опытах использовались две характеристики стимульных объектов, по которым варьировалась салиентность – цвет и ориентация в пространстве. Результаты экспериментов показали, чем выше салиентность (как по цвету, так и по ориентации), тем раньше возникает компонент и тем больше амплитуда самого пика (Töllner et al., 2011).
Есть сведения и о влиянии на N2pc цветового несоответствия стимулов (Zhao et al., 2011). В ходе опыта исследователи управляли цветовой интенсивностью цели, которая задавалась разными оттенками синего цвета, а также степенью сложности различения – низкой и высокой, которая задавалась разной геометрической формой цели. Дистракторы же имели белый цвет и постоянную форму. Результаты показали, что повышение амплитуды N2pc имело место при увеличении цветовой разницы между целью и дистракторами. При этом не было выявлено никаких достоверных отличий в характеристиках компонента N2pc, которые были бы связаны со степенью сложности различения, основанной на изменении формы цели. Таким образом, амплитуда N2pc может модулироваться физическим несоответствием, в данном случае цветовым, между целью и дистракторами непосредственно в ходе зрительного поиска (Zhao et al., 2011).
Имеются данные о влиянии эксцентриситета стимулов на компонент N2pc. Шаффер и соавт. в своем исследовании манипулировали эксцентриситетом отдельного участка изображения, текстура которого отличалась от фона, в пределах от 2 до 7°, и сообщили, что амплитуда N2pc немного уменьшалась с увеличением эксцентриситета. Однако целевой участок физически значительно отличался от фона, поэтому трудно определить, что отражает наблюдаемый эффект – связанные с вниманием изменения в N2pc или же латерализованную сенсорную активность низкого уровня в том же временном окне (Schaffer et al., 2011).
Папойоанну и Лак в своей статье представили результаты исследования влияния эксцентриситета целевых стимулов на амплитуду компонента N2pc (Papaioannou, Luck, 2020). Ими был проведен эксперимент, в котором участники решали задачу различения стимулов, предъявляемых с одним из пяти значений эксцентриситета (0, 0.05, 1, 2, 4 и 8°). Было обнаружено, что амплитуда N2pc оставалась относительно постоянной при всех значениях, кроме 8°. При максимальном эксцентриситете в восемь угловых градусов амплитуда N2pc уменьшалась более чем на 50% по сравнению с другими значениями эксцентриситета. Папойоанну и Лак сделали методический вывод в пользу того, что в экспериментах с применением N2pc можно располагать стимулы близко к средней линии экрана без риска существенно повлиять на амплитуду компонента, и одновременно с этим следует избегать большой удаленности стимулов от средней оси (Papaioannou, Luck, 2020).
Из результатов приведенных выше исследований следует, что при подготовке стимульного материала, как собственно цели, так и дистракторов, следует учитывать нюансы, которые могут повлиять на амплитуду, латентность, длительность и сам момент формирования N2pc.
НЕЙРОНАЛЬНЫЕ ГЕНЕРАТОРЫ N2PC
Волна N2pc считается компонентом ССП, отражающим процессы аттентивной обработки, поэтому изучение ее генераторов может рассматриваться как важный шаг в исследованиях нейрофизиологических механизмов внимания.
Как уже указывалось выше, сдвигу внимания при зрительном поиске обычно соответствует инициация нейрональной активности в префронтальной и задней теменной коре, после чего она концентрируется в вентральном зрительном пути, на промежуточных и поздних участках которого и генерируется N2pc (Buschman, Miller, 2007). В работе Хопфа и соавт. регистрировались ССП и МЭГ в парадигме, почти полностью повторяющей эксперимент (Luck, Hillyard, 1994a). Применение МЭГ позволило разделить N2pc (или mN2pc – магнитоэнцефалографический эквивалент N2pc) на две части: ранний компонент отражает активность нейронов теменной доли, в то время как поздний – работу латеральной части задних отделов, что скорее всего соотносится с верхней затылочной и задней нижневисочной зрительными областями. Хопф и соавт. предположили, что теменной источник активности ответственен за инициацию пространственного сдвига внимания к местоположению цели, в то время как затылочно-височный генератор отражает механизм отбора релевантной информации уже после сдвига внимания. Данная работа была первой, в которой четко показано, что N2pc является отражением контура активности теменных и затылочно-височных отделов (Hopf et al., 2000). В своей следующей работе с использованием несколько иной методики, которая предполагала предсказуемое расположение цели, Хопф и соавт. получили схожие результаты – в заданиях с одним и двумя дистракторами нейрональные генераторы N2pc располагались в вентральных затылочно-височных отделах обоих полушарий (Hopf et al., 2002).
В статье (Hopf, 2006) была дана более точная оценка местоположения нейрональных генераторов N2pc. В проведенном опыте испытуемые занимались обнаружением больших и малых целевых стимулов на изображениях разного масштаба. Такая парадигма позволила выявить быстрые изменения в фокусе зрительного внимания, которые соответствуют пространственному масштабу предъявляемых изображений. Оказалось, что генератор компонента меняется в соответствии с масштабом изображения. Когда изображение имело большой масштаб и включало в себя множество объектов, N2pc генерировался в латеральном затылочном комплексе, в то время как при работе с изображением малого масштаба, тоже включающим в себя некоторое множество объектов, генератор перемещался в корковую зону V4.
Исследование (Lorenzo-López et al., 2011) было посвящено возрастным изменениям в зрительно-пространственном внимании и электрофизиологическим коррелятам этого процесса. Активность мозга в их работе исследовалась при помощи МЭГ, где было обнаружено, что источник нейронной активности, лежащей в основе mN2pc, как у молодых, так и у пожилых испытуемых, располагается в затылочных областях – 17, 18, 19 полях по Бродману, – и нижних височных отделах – в 20, 21, 36 и 37 полях. Приведенные выше результаты воспроизводились и в работе (Boehler et al., 2011).
Значительный интерес в контексте обсуждаемой проблематики представляют результаты ряда исследований нейрофизиологических механизмов зрительного поиска у животных, полученные как при помощи регистрации потенциалов с поверхности головы, так и с применением техники микроэлектродной регистрации активности нервных клеток. В работе (Young et al., 2011) была проверена гипотеза о схожести нейронных сетей обезьяны, отвечающих за N2pc, с таковыми у человека. Вычислив при помощи томографии низкого разрешения (LORETA) плотность распределения токов в коре в ходе выполнения эксперимента, Янг и соавт. определили, что источник активности N2pc лежит в экстрастриарной зрительной коре, что в целом соотносится с данными, полученными на людях.
В работе (Westerberg et al., 2021) изучался корковый нейронный ансамбль, по всей видимости, отвечающий за генерирование электрической активности, связанной с вниманием и регистрируемой в задних отделах мозга у человека и обезьян. Исследование проводилось на макаках, обученных выполнять зрительный поиск в экспериментальных условиях. Вначале при выполнении задания записывалась ЭЭГ, на втором этапе животные выполняли зрительный поиск уже с имплантированными им в зону V4 микроэлектродами. Исходя из полученных результатов и данных о функциональной роли электрической активности (рис. 3), отождествляемой с вниманием, Вестерберг и соавт. предположили, что N2pc возникает в ходе процесса взаимодействия нескольких областей коры. Учитывая повсеместное распространение колончатой архитектуры в сенсорной коре и специфику репрезентации зрительных признаков в различных областях, электрические диполи, формирующиеся в этих областях, могут осуществлять различный вклад в итоговую активность в зависимости от того, какой признак в данный момент воспринимается. Более того, вклад других областей, помимо V4, весьма вероятен, поскольку предыдущие нейрофизиологические исследования на макаках показали участие в зрительном поиске височных (Sato, 1988), теменных (Bisley, Mirpour, 2019) и лобных отделов (Thompson et al., 2005; Zhou, Desimone, 2011). Наряду с этим следует отметить, что исследования у людей указывают на вклад в генерацию N2pc задней теменной коры (Hopf et al., 2000) и передних глазодвигательных областей, которые обнаруживают цель среди дистракторов даже раньше возникновения самого N2pc (Purcell et al., 2013). Учитывая такую взаимосвязь передних областей и V4 (Gregoriou et al., 2012; Ninomiya et al., 2012), Вестерберг и соавт. предположили, что лобная доля может быть функциональным источником активности, являющейся нейрональным коррелятом селективности внимания, которая затем передается в V4 и другие задние области (Gregoriou et al., 2012; Popov et al., 2017), и которые в свою очередь генерируют N2pc (Westerberg, Schall, 2021).
Рис. 3.
Локализация N2pc на примере иллюстрации из работы (Westerberg et al., 2021). Трехмерная локализация распределения плотности токов на поверхности правого полушария мозга макаки, согласующаяся по времени с N2pc в момент, когда цель поиска находилась в левом полуполе зрения. Максимальная плотность тока сосредоточена под электродом P6 каудальнее лунной борозды и в области V4 на угловой извилине.
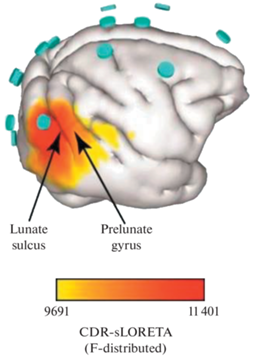
Таким образом, можно сделать общий вывод о том, что с точки зрения нейрофизиологии компонент N2pc – сложное явление, которое представляет собой результат взаимодействия нескольких отдельных нейрофизиологических процессов и работы множества областей коры.
ПРАКТИКА ПРИМЕНЕНИЯ ПАРАДИГМЫ N2PC В ИССЛЕДОВАТЕЛЬСКОЙ РАБОТЕ
Регистрация N2pc широко используется в исследованиях прикладного характера, в которых данный компонент выступает маркером организации процессов, связанных с избирательным вниманием. Такие работы охватывают разные возрастные группы, а также людей, страдающих различными заболеваниями и психическими отклонениями. Ярким примером может служить статья Лоренцо-Лопес и соавт., в которой показано, что амплитуда N2pc у пожилых людей снижена, а время возникновения и пик волны компонента имеют несколько большую латентность, чем у более молодых. Тем самым авторы продемонстрировали влияние старения на механизмы распределения внимания, которые отражает N2pc (Lorenzo-López et al., 2008b). Время возникновения волны N2pc и латентность пика также увеличены у пациентов с печеночной энцефалопатией – нейродегенеративным заболеванием, являющимся следствием печеночной недостаточности. Лоренцо-Лопес и соавт. объясняют это нарушением функционального взаимодействия нисходящих и восходящих процессов обработки поступающей информации, ссылаясь на исследование (Schiff et al., 2006). Однако никаких изменений в амплитуде или латентном периоде N2pc не было выявлено в экспериментах с больными шизофренией (Luck et al., 2006), болезнью Паркинсона (Praamstra, Plat, 2001), у спортсменов, многократно получавших сотрясение мозга (Beaumont et al., 2007).
Применяется N2pc и в исследованиях, связанных с психологическими особенностями людей. Так, Крусэрмак и соавт. представили данные о том, что у испытуемых с высокой выраженностью психопатии (измеренной при помощи метода PCL-R) конгруэнтная фокусу внимания информация вызывает достоверно больший N2pc, чем у испытуемых с низкой и средней психопатизацией. Авторы связали этот эффект с усилением у таких людей нисходящих влияний на ранних стадиях зрительного внимания (Krusemark et al., 2016). В исследовании Ли и соавт. было показано, что у испытуемых с низкой самооценкой N2pc на стимулы-подсказки, нерелевантные по отношению к инструкции, имел большую амплитуду, чем у испытуемых с высокой самооценкой (Li et al., 2012). У людей с выраженной социальной тревожностью выше амплитуда и короче латентность N2pc при восприятии лиц с угрожающим или гневным выражением лица. Стоит отметить, что чем тяжелее такое тревожное состояние, тем больше растет амплитуда и уменьшается латентность компонента, таким образом делая из N2pc электрофизиологический маркер описанного состояния (Reutter et al., 2017b). В экспериментах со зрительным поиском целей, вызывающих страх (изображения пауков), у людей с арахнофобией наблюдался достоверно более выраженный N2pc по сравнению с контрольной группой, что интерпретировалось Веймаром и соавт. как нейрональный маркер усиления избирательного внимания к стимулам, связанным со страхом (Weymar et al., 2013). У людей с фобией вида крови N2pc возникал в ответ на демонстрацию изображений травм, причем он был больше, чем в ответ на изображения бескровных драк. Исследователи пришли к выводу, что такое смещение внимания связано с ранним зрительно-пространственным отбором релевантных фобии стимулов и отчетливо отражается в изменении амплитуды N2pc (Buodo et al., 2010).
В последние годы появилось множество работ с регистрацией компонента N2pc, целью которых является исследование механизмов, ответственных за распознание эмоционального выражения лица. Например, в статье (Yuan et al., 2020) статистически закономерные отличия были обнаружены в амплитуде N2pc при демонстрации статичного лица с выражением отвращения: у участников с повышенной социальной тревожностью амплитуда N2pc была выше по сравнению с контрольной группой. При осмотре анимированных лиц с тем же выражением отличий обнаружено не было. Юань и соавт. сделали вывод, что у испытуемых экспериментальной группы экзогенное внимание с большей вероятностью будет захвачено лицом с ярко выраженной, “пиковой” эмоцией из-за их преаттентивной настройки на постоянный поиск внешних стимулов-угроз (Yuan et al., 2020). В работе (Liu et al., 2021), посвященной гипотезе “превосходства гнева” (“anger superiority”) – предположению о том, что выражение гнева на лице лучше захватывает внимание на ранней стадии зрительного поиска, чем счастливое или нейтральное лицо, – был проведен метаанализ исследований, в которых применялась N2pc-парадигма. По итогам проведенной работы Лю и соавт. констатировали, что амплитуда N2pc достоверно больше на лица с выражением счастья или гнева по сравнению с нейтральными. При этом разница эффектов для счастливых и гневных лиц была статистически незначимой, в то время как для самого N2pc она модулировалась возрастом испытуемых, количеством предъявлений, реальным или схематичным изображением лиц. Такой результат привел к выводу о том, что результаты исследований с N2pc на здоровых людях не подтверждают описанную выше гипотезу о “превосходстве гнева”. Необходимо упомянуть также использование парадигмы N2pc при исследовании аспектов захвата внимания угрожающими и счастливыми выражениями лица (Burra et al., 2016) и эмоционально окрашенными сценами (Kim et al., 2017).
Кроме перечисленного выше, в последние годы компонент N2pc использовался для изучения следующих аспектов внимания: контроля пространственного внимания (Lien et al., 2011); развертывания зрительного внимания в процессе временной интеграции информации (Akyürek, Meijerink, 2012); захвата внимания в ходе зрительного поиска заданной и случайной целей (Burra, Kerzel, 2013); управления вниманием на основе категорий (Nako et al., 2014); скорости последовательного смещения внимания при зрительном поиске (Grubert, Eimer, 2016); скрытого внимания (Xu et al., 2016); нейрональных механизмов селективности внимания (Bachman et al., 2020). Следует отметить использование N2pc при исследовании закономерностей группирования признаков в объекты и объектов в категории (Wu et al., 2016); обнаружения зрительных синглтонов (Tay et al., 2019); обнаружения лиц (Bola et al., 2021).
ЗАКЛЮЧЕНИЕ
В заключение представляется целесообразным остановиться на некоторых возможных областях применения N2pc. Перспективным видится использование этого компонента для изучения приоритета различных стимулов для зрительного внимания. Поскольку у компонента N2pc, помимо амплитуды, может модулироваться латентность пика и, следовательно, момента возникновения самой волны, такое изменение можно трактовать как нейрофизиологический маркер приоритета стимула – чем он важнее для системы, тем раньше мозг начнет вычислять его местоположение. Такой подход будет весьма актуален, например, при исследовании внимания к признакам (feature attention). Также стоит отметить возможность применения N2pc в исследованиях процессов категоризации: существующие работы показали принципиальную возможность того, что N2pc может отражать использование мозгом категориальной информации наряду с визуальными различиями во время зрительного поиска (Moon et al., 2022).
Актуальными представляются также дальнейшие исследования нейрофизиологических основ N2pc как важной части мозговых систем внимания. Современные нейрофизиологические исследования, выполненные на мозге приматов с использованием техники микроэлектродной регистрации клеточной активности (Bichot et al., 2015, 2019; Westerberg et al., 2021), выявили ряд важных аспектов взаимодействия некоторых фронтальных структур (VPA, FEF) со зрительными областями, расположенными дорзальнее (V4 и другие). Сопоставление нейрофизиологических результатов с данными ЭЭГ-исследований человека может явиться ключом к пониманию механизмов генерации N2pc и процессов, которые он отражает.
Подводя итог, следует еще раз подчеркнуть, что компонент N2pc является надежным индикатором, а возможно, и ключевым электрофизиологическим маркером процесса выбора цели в зрительном поиске. Особенности его возникновения позволяют исследовать как отдельно взятые операции селекции, так и свойства нисходящих и восходящих процессов управления вниманием, некоторые аспекты зрительной рабочей памяти, а также целый спектр задач, связанных с эмоциональным распознаванием и реагированием на эмоционально-валентные стимулы, как было указано выше.
Таким образом, описанный в статье компонент ССП является эффективным инструментом исследования механизмов зрительного внимания.
Список литературы отсутствует.
Дополнительные материалы отсутствуют.
Инструменты
Сенсорные системы