

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 1, стр. 97-116
Новые палинологические данные из среднеэоценовых отложений опорного разреза Хеу, Кабардино-Балкария, Северный Кавказ
А. И. Яковлева 1, *, Д. Д. Вага 2, А. С. Андреева-Григорович 3, Э. П. Радионова 1
1 Геологический институт РАН
Москва, Россия
2 Department of Geology, University of Nairobi
Nairobi, Kenya
3 Институт геологических наук НАН
Киев, Украина
* E-mail: alina.iakovleva@gmail.com
Поступила в редакцию 02.05.2018
После доработки 22.05.2018
Принята к публикации 25.10.2018
Аннотация
Представлены результаты изучения палиноморф из средне-верхнеэоценового интервала опорного разреза Хеу. В исследованной части разреза выявлены интервалы диноцистовых зон Enneadocysta arcuata, Costacysta bucina, Enneadocysta pectiniformis, Rhombodinium draco, Reticulatosphaera actinocoronata, Talladinium? clathratum. На основе прямого сопоставления с данными по известковому наннопланктону установлены их стратиграфические границы, а также уточнены уровни появлений стратиграфически важных видов диноцист. Новые данные изучения микропланктона подтверждают раннелютетский возраст верхов черкесской свиты и керестинской свиты; кумская свита датируется второй половиной лютета–началом приабона, а низы белоглинской свиты – приабоном. Анализ соотношений водных и континентальных палиноморф в разрезе позволяет интерпретировать обстановки осадконакопления в течение лютета–приабона в исследуемой части Восточного Пери-Тетиса.
ВВЕДЕНИЕ
На протяжении 80 лет палеогеновые отложения Предкавказья привлекают исключительное внимание биостратиграфов и палеонтологов. Изучению центральной и восточной частей северного склона Кавказа посвящены многочисленные исследования, прежде всего, Н.С. Шатского, Н.Н. Субботиной, И.А. Коробкова, Е.К. Шуцкой, В.Г. Морозовой, Г.П. Леонова и В.П. Алимариной, В.А. Гроссгейма и многих других (Стратиграфия…, 1975). Северный Кавказ является одним из наиболее полно и детально изученных на территории бывшего СССР районов развития палеогеновых отложений. Разрезы Северного Кавказа известны своей стратиграфической полнотой и разнообразием фациальных типов отложений с многочисленными ископаемыми остатками.
В 1970-е годы и в начале 1980-х годов К.Е. Аристова, Л.А. Козяр, Л.А. Панова активно исследовали на Северном Кавказе пыльцу и споры наземных растений. Начало изучения органикостенного фитопланктона в регионе связано с именем К.Е. Аристовой (1973), а наибольший вклад в изучение палеогеновых диноцист здесь внесли А.С. Андреева-Григорович (1991) и Н.И. Запорожец (Ахметьев, Запорожец, 1996; Крашенинников и др., 1998; Запорожец, 2001). Так, А.С. Андреева-Григорович изучала наннопланктон и диноцисты в опорных разрезах по рекам Кубань и Белая, скв. Новопокровская-4 (Ставрополье) в интервале от границы мела/палеогена до границы олигоцена/миоцена. В свою очередь, Н.И. Запорожец исследовала диноцисты в более узком интервале (средний эоцен–олигоцен) в тех же опорных разрезах по рекам Кубань, Белая и Хеу (кумская свита).
Разрез по реке Хеу (рис. 1, 2), расположенный на окраине поселка Герпегеж в ~18 км юго-восточнее г. Нальчика в Кабардино-Балкарии, очень хорошо известен и неоднократно описывался в литературе (Шуцкая, 1956, 1970; Гроссгейм, 1960; Алимарина, 1963; Леонов, Алимарина, 1964). Он является наиболее полным из изученных на Северном Кавказе разрезов, имеет мощность более 300 м и охватывает стратиграфический интервал от верхов маастрихта до основания олигоцена. Первое зональное расчленение разреза Хеу по фораминиферам было проведено Н.Н. Субботиной еще в 1930-е годы, а затем В.А. Крашенинниковым (Крашенинников, Музылев, 1975). Особую роль в изучении разреза Хеу сыграл Н.Г. Музылев, который исследовал известковый наннопланктон из ~350 образцов и разработал первоначальное расчленение этого опорного разреза по наннопланктону (Крашенинников, Музылев, 1975; Музылев, 1980; Gavrilov et al., 2000). В последующие годы Е.А. Щербинина продолжила детальное изучение эоценового наннопланктона из разреза Хеу (Shcherbinina, 2000; Shcherbinina et al., in press), а для уровня перехода от палеоцена к эоцену ею получены данные высокой степени разрешаемости (Shcherbinina et al., 2016).
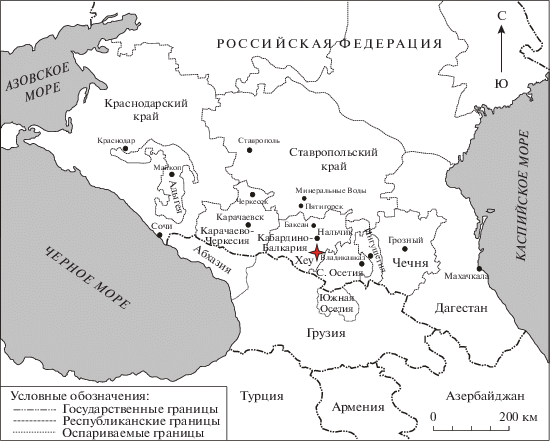
Рис. 2.
Разрез Хеу на палинспастической палеогеографической карте Западной Евразии в приабоне (Попов и др., 2009 с сокращениями). 1–3 – морские условия: 1 – склон и батиаль; 2 – глубоководный шельф; 3 – мелководный шельф; 4–6 – континентальные условия: 4 – возвышенности; 5 – низменности; 6 – пресноводные озера и лагуны; 7 – разломы; 8 – сбросы и флексуры, одновременные с седиментацией; 9 – синседиментационные надвиги.

Что касается диноцист, то до недавнего времени они изучались лишь из отдельных стратиграфических горизонтов разреза Хеу. Так, Н.И. Запорожец (2001) опубликованы данные по диноцистам из 10 образцов кумской свиты, которые исследовались ею в рамках комплексного анализа кумского горизонта на Северном Кавказе и в смежных областях (Ростовский выступ, Северные Ергени). Позднее диноцисты были изучены из палеоценовой и переходной от палеоцена к э-оцену частей разреза, но опубликованы лишь в кратком виде (Radionova et al., 2009). Наконец, недавно появились результаты комплексных микропалеонтологических и геохимических исследований (включая диноцисты) высокой степени разрешаемости из переходного интервала от палеоцена к эоцену (Shcherbinina et al., 2016). К настоящему времени проведено детальное комплексное микропалеонтологическое (диноцисты и наннопланктон) и геохимическое изучение нижнеэоценовых отложений (абазинская и черкесская свиты) в разрезе Хеу (Shcherbinina et al., in press). Целью же настоящей статьи явились детальный палиностратиграфический анализ средне-верхнеэоценовых отложений, сопоставление диноцистовых и наннопланктонных событий (датумов) в этом интервале разреза, восстановление обстановок осадконакопления и сравнение полученных данных с таковыми из одновозрастных отложений в соседних регионах.
МАТЕРИАЛ И МЕТОДЫ
В настоящей работе было изучено 47 образцов, отобранных из среднего эоцена и низов верхнего эоцена разреза Хеу в 2000 г. в рамках экспедиционных работ сотрудниками ГИН РАН Э.П. Радионовой, В.Н. Беньямовским и С.И. Ступиным.
Нижняя часть рассматриваемого интервала представлена верхами черкесской свиты мощностью более 12 м, сложенной зелеными и серо-зелеными с голубоватым оттенком мергелями. Выше по разрезу залегает керестинская свита (~13 м), представленная толщей зеленовато-серых мергелей, переходящих в чередующиеся светлые известняки и мергели. Керестинская свита перекрывается кумской свитой. Последняя в основании представлена единичными прослоями пеплов (бентониты); нижняя часть свиты сложена бурыми, темно-бурыми, кофейными мергелями (~10 м); выше следует задернованный участок мощностью 25–30 м; верхняя опробованная часть свиты представлена буровато-кофейным мергелем (1 м), сменяющимся переслаиванием бурых мергелей и беловато-серых известняков (13 м). Низы белоглинской свиты (>5 м) сложены голубовато-зеленовато-серыми щебенчатыми мергелями. Бóльшая часть свиты (общая мощность свиты ~100 м) в рамках настоящего исследования не опробовалась.
Палинологические исследования осуществлялись параллельно с изучением известкового наннопланктона, что позволило нам провести прямое сопоставление стратиграфического распределения видов диноцист и ключевых таксонов наннопланктона (рис. 3). Подробный анализ изучения наннопланктона и фораминифер из средне-вехнеэоценового интервала представлен в статье Waga et al. (in press).
Рис. 3.
Калибровка диноцистовых событий (датумов) и диноцистовых зон с датумами и зонами по наннопланктону в разрезе Хеу. НП-зоны – зоны по наннопланктону, Reticul. actinoc – Reticulatosphaera actinocoronata, Tallad.? clathrat – Talladinium? clathratum; полужирным шрифтом выделены наиболее важные виды.

Обработка палинологических образцов проведена в соответствии со стандартной методикой, принятой в Лаборатории палеофлористики ГИН РАН, и включает: (1) воздействие на образцы 10%-ной соляной кислотой (HCl) для удаления карбонатов; (2) воздействие горячим раствором пирофосфата натрия (Na4P2O7 · 10H2O) для дисперсии глинистого материала, а затем отмывку каждые 2 часа с целью удаления глинистых частиц; (3) центрифугирование образцов в тяжелой жидкости (K2CdI4) с плотностью 2.25 с целью отделения органической фракции от более тяжелых минеральных частиц; (4) воздействие 70%-ной плавиковой кислотой (HF) для растворения кремнистых компонентов; (5) воздействие 10%-ной соляной кислотой для удаления флюоросиликатных гелей; (6) отмывку образца в дистиллированной воде и заливку глицерином. Просеивание мацерата через сита не проводилось.
Таксономия диноцист соответствует индексу DINOFLAJ3 (Williams et al., 2017). Количественное палинологическое изучение материала проводилось в 2 этапа: (1) подсчет в каждом образце минимум 200–250 морских и наземных палиноморф (цисты динофлагеллат, акритархи, празинофиты, пыльца покрытосеменных, двухмешковая пыльца хвойных растений, споры наземных растений, внутренние камеры фораминифер); (2) дальнейший подсчет минимум 200 диноцист, после чего препарат просматривался до конца с целью обнаружения редко встречаемых таксонов. Палеоэкологическая интерпретация количественных флуктуаций в палинокомплексах основана в целом на работах (Brinkhuis, 1994; Powell et al., 1996; Crouch, Brinkhuis, 2005; Sluijs et al., 2005; Torricelli et al., 2006). Для интерпретации возможных палеообстановок цисты динофлагеллат были объединены в 18 групп морфологически близких таксонов: (1) Wetzelielloids; (2) Deflandrea; (3) Phthanoperidinium; (4) Lingulodinium-группа (Lingulodinium, Homotryblium, Heteraulacacysta, Polysphaeridium); (5) Areoligera-группа (Areoligera, Glaphyrocysta, Adnatosphaeridium, Membranophoridium, Emmetrocysta); (6) Cleistosphaeridium diversispinosum; (7) Enneadocysta/Areosphaeridium; (8) Thalassiphora; (9) Cordosphaeridium-группа (Cordosphaeridium, Fibrocysta, Muratodinium, Kenleyia); (10) Operculodinium; (11) Diphyes/Dapsilidinium; (12) Hystrichokolpoma; (13) Melitasphaeridium pseudorecurvatum; (14) Cribroperidinium; (15) Spiniferites-группа (Spiniferites, Spiniferella, Hystrichosphaeropsis, Hystrichostrogylon, Rottnestia); (16) Batiacasphaera; (17) Impagidinium; (18) Nematosphaeropsis labyrinthus; в 2 отдельные группы выделены оставшиеся перидиниоидные и гониаулакоидные диноцисты.
При описании палинологических результатов в изученных разрезах используется термин “диноцистовый интервал”, под которым понимается часть конкретного разреза с определенным комплексом диноцист (присутствие стратиграфических маркеров и количественные характеристики палинологических ассоциаций).
Коллекция образцов хранится в Лаборатории палеофлористики Геологического института РАН (Москва).
РЕЗУЛЬТАТЫ
Далеко не все образцы из изученного интервала разреза Хеу содержали количественно представительные палинологические комплексы, особенно в верхней части керестинской свиты. Все выявленные количественно представительные комплексы отличаются доминированием диноцист (от 60 до 100%); лишь на границе кумской и белоглинской свит наблюдается резкий и краткий скачок в содержании континентальных палиноморф, а количество диноцист сокращается до 40%. В отличие от нижнеэоценового интервала разреза Хеу (Shcherbinina et al., in press), в комплексах среднего эоцена в большей степени участвует двухмешковая пыльца хвойных (до 20–30%), а в кумской свите отмечаются частые находки внутренних камер фораминифер. Акритархи и празинофиты в верхах черкесской и низах керестинской свит составляют от 0 до 10% и почти отсутствуют в кумской свите.
На рис. 3–6 показаны стратиграфические результаты изучения диноцист и наннопланктона по разрезу, а также количественное распределение групп палиноморф и диноцистовых экогрупп. На фототабл. I–IV представлены наиболее характерные виды диноцист из изученных отложений.
Рис. 4.
Стратиграфическое распределение видов диноцист в разрезе Хеу. R. act. – Reticulatosphaera actinocoronata; Tall.? clath. – Talladinium? clathratum; Бел. – Белоглинская.
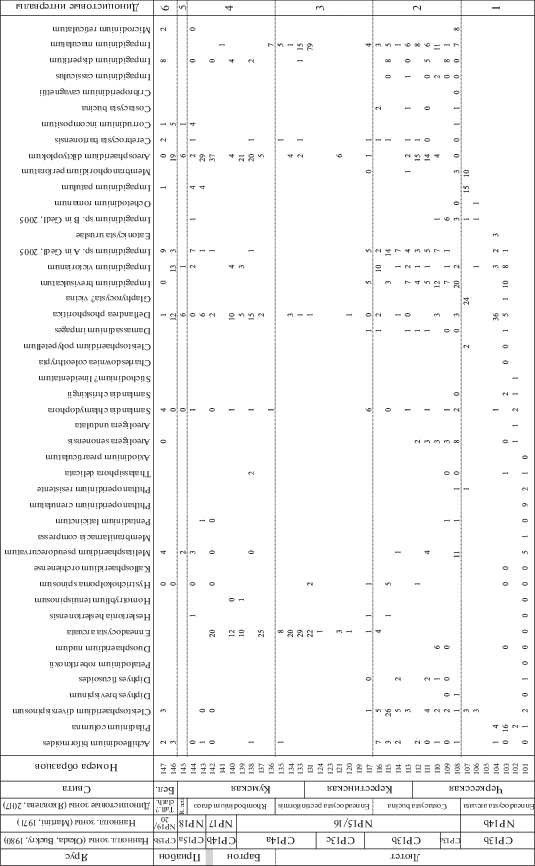
Рис. 4.
Окончание
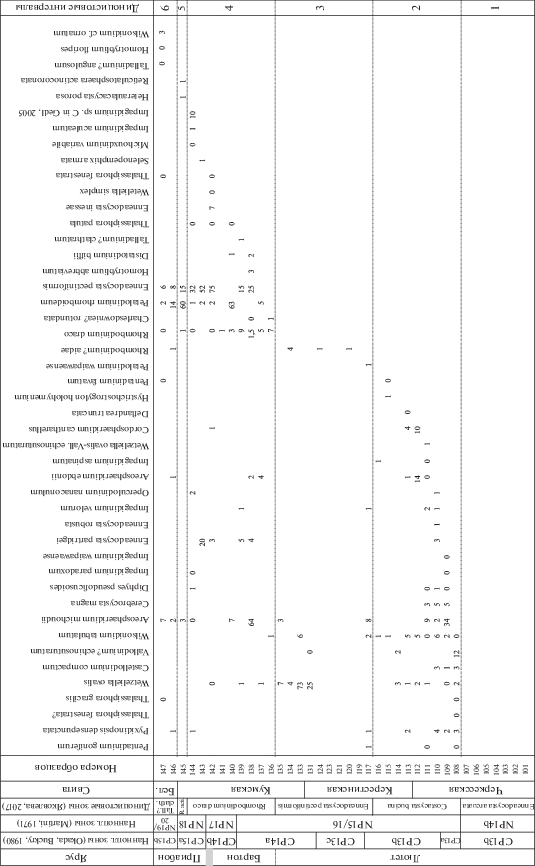
Рис. 5.
Количественное распределение основных групп палиноморф в разрезе Хеу. E. arc. – Enneadocysta arcuata; R. act. – Reticulatosphaera actinocoronata; палиноморфы неясн. сист. – палиноморфы неясной систематической принадлежности.
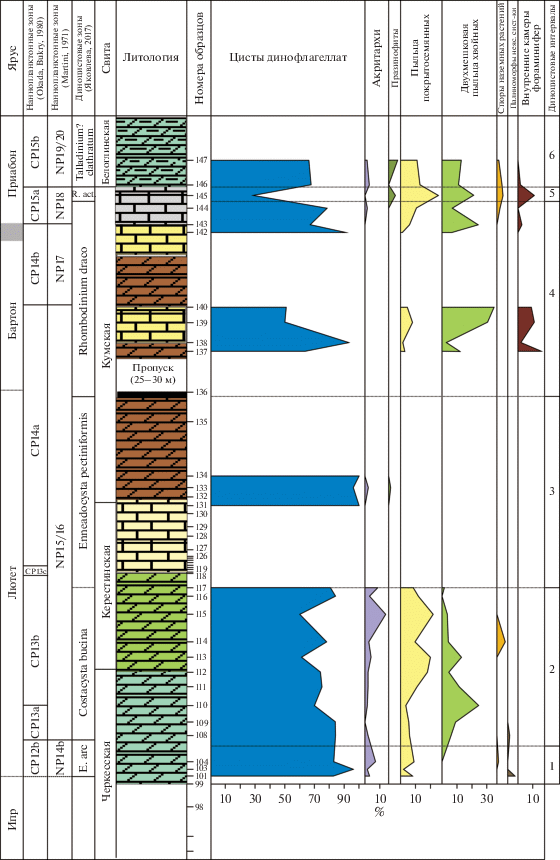
Рис. 6.
Количественное распределение экогрупп диноцист в разрезе Хеу. НП-зоны – наннопланктонные зоны, E. arc. – Enneadocysta arcuata; R. act. – Reticulatosphaera actinocoronata; Nematosph. labyrinthus – Nematosphaeropsis labyrinthus.
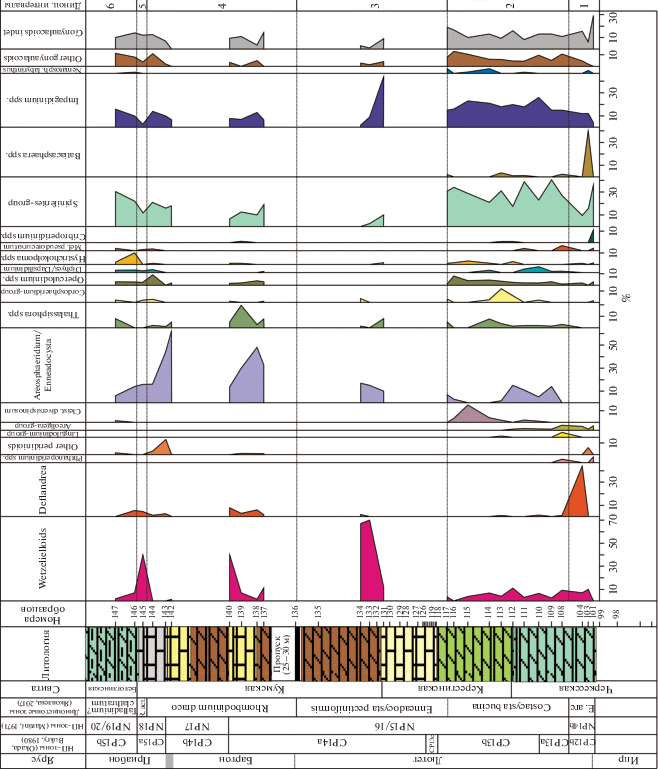
Таблица I. Диноцисты из эоценовых отложений разреза Хеу. 1 – Wilsonidium tabulatum, обр. 112; 2, 3 – Petalodinium rhomboideum, обр. 145; 4, 5 – Rhombodinium? aidae, обр. 146; 6, 10 – Wilsonidium tabulatum, обр. 116; 7–9 – Impagidinium dispertitum, обр. 142; 11, 12 – Corrudinium incompositum, обр. 144; 13, 14 – Areosphaeridium ebdonii, обр. 111; 15, 16 – Enneadocysta pectiniformis, обр. 144; 17, 18 – Areosphaeridium michoudii, обр. 109.
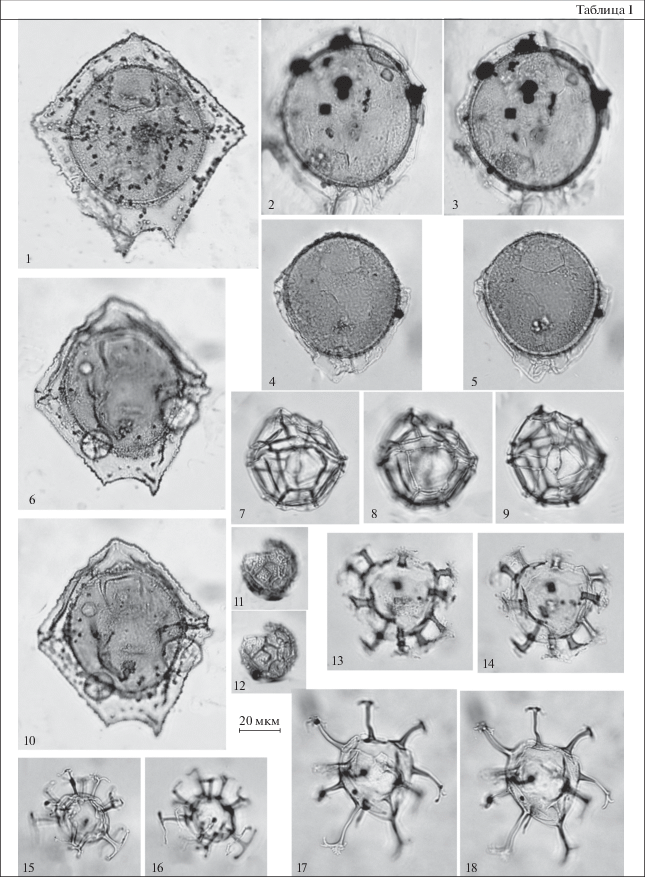
Таблица II. Диноцисты из эоценовых отложений разреза Хеу. 1, 2 – Wilsonidium tabulatum, обр. 111; 3 – Enneadocysta pectiniformis, обр. 142; 4, 5 – Vallodinium? echinosuturatum, обр. 111; 6–8 – Impagidinium maculatum, обр. 111; 9 – Impagidinium sp. A sensu Gedl, 2005, обр. 144; 10, 11 – Enneadocysta inessae, обр. 142; 12 – Cleistosphaeridium diversispinosum, обр. 108; 13 – Impagidinium dispertitum, обр. 111; 14, 17 – Impagidinium sp. B sensu Gedl, 2005, обр. 144; 15 – Impagidinium sp. C sensu Gedl, 2005, обр. 144; 16 – Corrudinium incompositum, обр. 144; 18, 19 – Impagidinium sp. A sensu Gedl, 2005, обр. 111; 20, 21 – Impagidinium brevisulcatum, обр. 108.
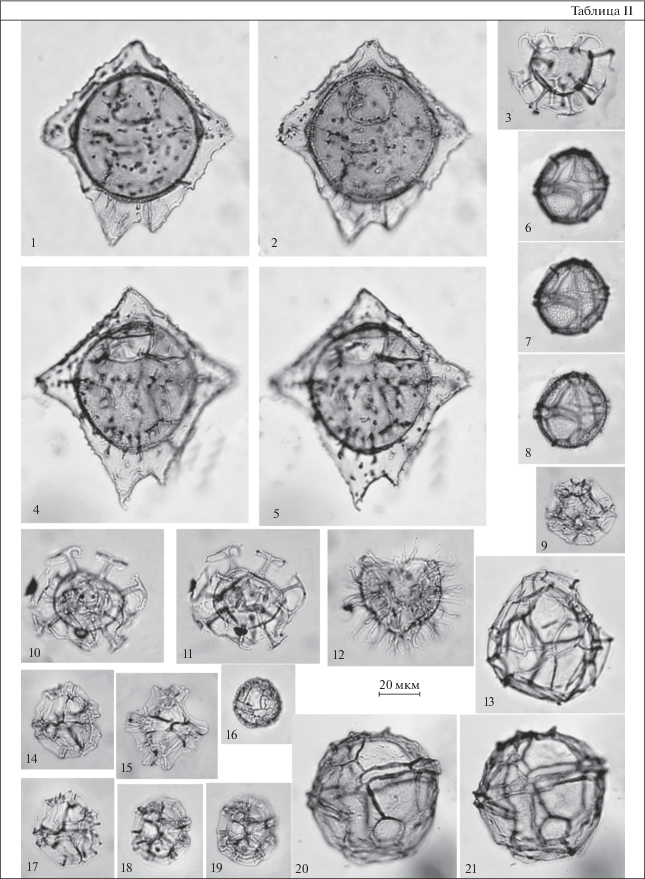
Таблица III. Диноцисты из эоценовых отложений разреза Хеу. 1–3, 6 – Impagidinium cassiculum, обр. 111; 4 – Wetzeliella ovalis-Vallodinium? echinosuturatum, обр. 111; 5 – Castellodinium compactum, обр. 108; 7 – Areosphaeridium diktyoplokum, обр. 147; 8 – Costacysta bucina, обр. 108; 9 – Rhombodinium draco, обр. 147; 10–12 – Impagidinium victorianum, обр. 144; 13, 14 – Microdinium ornatum, обр. 108.
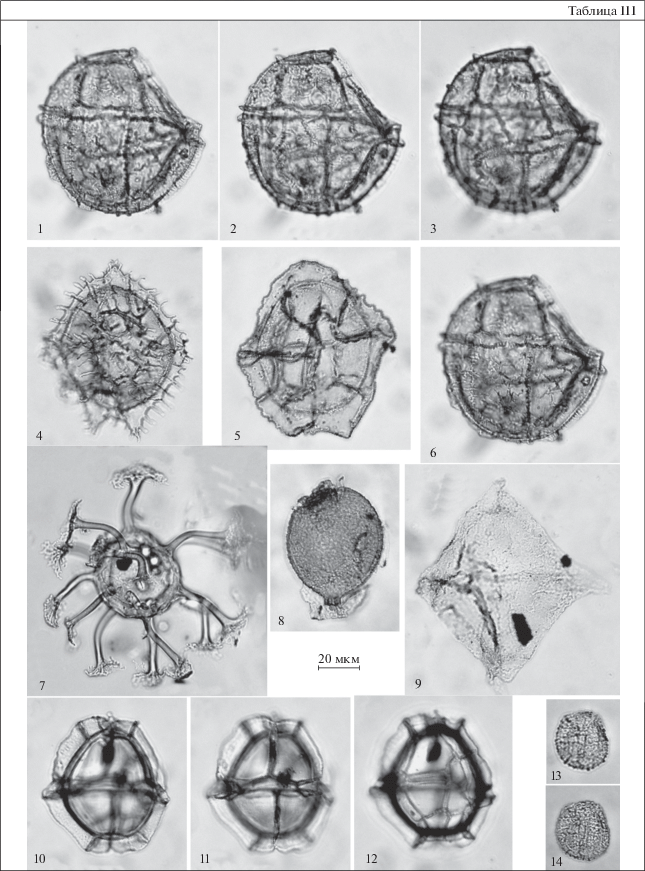
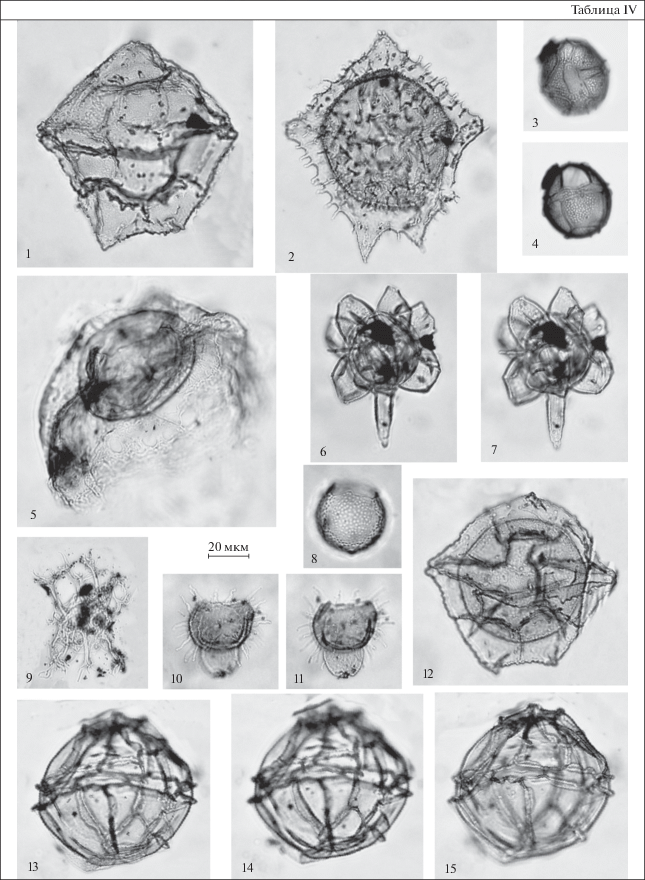
Таблица IV. Диноцисты из эоценовых отложений разреза Хеу. 1 – Castellodinium compactum, обр. 108; 2 – Wetzeliella ovalis, обр. 108; 3, 4 – Impagidinium maculatum, обр. 111; 5 – Thalassiphora fenestrata, обр. 108; 6, 7 – Hystrichokolpoma cinctum, обр. 147; 8 – Batiacasphaera compta, обр. 111; 9 – Distatodinium biffii, обр. 138; 10, 11 – Diphyes ficusoides, обр. 109; 12 – Castellodinium compactum, обр. 108; 13–15 – Impagidinium brevisulcatum, обр. 111.
Последовательное распределение стратиграфически важных видов диноцист, а также количественные флуктуации внутри палинологических ассоциаций позволяют выделить в средне-верхнеэоценовой части разреза Хеу шесть диноцистовых интервалов.
Первый диноцистовый интервал (верхняя часть черкесской свиты, обр. 101–107). Стратиграфически важный вид Enneadocysta arcuata обнаружен с основания рассматриваемой здесь части черкесской свиты (обр. 101), которая, согласно данным изучения наннопланктона, отнесена к интервалу подзон NP14b (лютетская часть) (Martini, 1971) и CP12b (Okada, Bukry, 1980). Следует отметить, что вид E. arcuata выявлен в разрезе Хеу чуть раньше первого появления (LO – lowermost occurrence) наннопланктона Nannotetrina quadrata (обр. 103), то есть однозначно ниже основания наннопланктонных зон NP15–16 и CP13a. Внутри этого интервала наблюдаются последовательные последние появления (HOs, highest occurrences) видов Petalodinium robertknoxii, Charlesdowniea coleothrypta, Piladinium columna и Eatonicysta ursulae. Указанный первый диноцистовый интервал среднеэоценовой части разреза Хеу (обр. 101–107) соответствует зоне Enneadocysta arcuata зональной шкалы Восточного Пери-Тетиса (Яковлева, 2017), чья нижняя граница устанавливается по LO E. arcuata. Возраст этой части черкесской свиты – начало лютета.
Палинологический комплекс в интервале обр. 101–107 характеризуется абсолютным доминированием диноцист при небольшом участии (5–10%) акритарх и пыльцы покрытосеменных. В диноцистовом комплексе можно отметить доминирование группы Spiniferites (>30%) в основании интервала, сменяющееся резким и кратковременным увеличением содержания Batiacasphaera (40%) и появлением Nematosphaeropsis, характерного для наиболее глубоководных обстановок обитания динофлагеллат; выше по разрезу наблюдается такое же резкое и кратковременное возрастание количества дефландреоидных (40%); Impagidinium составляет ~10% от комплекса. Исходя из указанных количественных характеристик палинокомплекса, мы предполагаем накопление нижнелютетской части черкесской свиты в открыто-морских условиях во внешней неритической зоне с последующим возможным переходом в более мелководные обстановки, где гетеротрофные диноцисты имели больший доступ к питательным веществам с суши. На прибрежно-мелководные условия также указывает уменьшение видового разнообразия наннопланктона и массовое развитие вида Zygrhablithus bijugatus.
Второй диноцистовый интервал (верхи черкесской–низы керестинской свит, обр. 108–116). Выявлен по первому появлению в основании интервала стратиграфически важных видов Costacysta bucina, Wetzeliella ovalis, Castellodinium compactum, Vallodinium? echinosuturatum. Указанные диноцистовые события совпадают в разрезе Хеу с первым появлением наннопланктона Nannotetrina fulgens. В основании интервала отмечено последнее появление вида Ochetodinium romanum, а выше по разрезу (обр. 110) фиксируется HO Duosphaeridium nudum, совпадающее с LO наннопланктона Chiasmolithus gigas (начало подзоны CP13b). Первые появления видов Enneadocysta partridgei и Enneadocysta robusta отмечаются в кровле черкесской свиты, чуть выше LO Chiasmolithus gigas; в обр. 112 впервые для данного разреза появляется вид Cordosphaeridium cantharellus (чуть ниже LO наннопланктона Discoaster tani nodifer). Рассматриваемый интервал разреза соответствует диноцистовой зоне Costacysta bucina зональной шкалы Восточного Пери-Тетиса, установленной в разрезе Актумсук в Узбекистане по первому появлению номинального вида, а также выявленной в скв. 230 в Днепровско-Донецкой впадине (Яковлева, 2017). Исходя из данных по известковому наннопланктону (зоны NP14b(верхи)–NP15/16 или CP12b(верхи)–CP13a), верхи черкесской свиты–низы керестинской свиты датируются частью лютета.
Палинологический комплекс характеризуется здесь общим доминированием диноцист (от 85 до 65%), однако внутри интервала наблюдаются определенные флуктуации в содержаниях различных групп палиноморф: пыльца покрытосеменных достигает 20%, а пыльца хвойных – 25%. В диноцистовом комплексе отмечается значительное содержание групп Spiniferites (20–40%) и Impagidinium (до 20%), исчезновение дефландреоидных и небольшое количество остальных перидиниоидных (ветзелиеллоидных и Phthanoperidinium). В верхней части интервала наблюдаются пик Cleistosphaeridium diversispinosum и совсем небольшой пик Nematosphaeropsis. Существенная роль Impagidinium, а также единичное присутствие Nematosphaeropsis свидетельствуют об открыто-морских обстановках осадконакопления, как минимум во внешней неритической зоне, что подтверждается и данными по наннопланктону: в ассоциациях преобладают роды открыто-океанической среды (Reticulofenestra, Cyclicargolithus, Chiasmolithus), реже представлены виды прибрежно-морских условий (Zygrhablithus bijugatus, Nannotetrina и Micrantholithus); здесь также отмечается рост видового разнообразия (до 37 видов).
Третий диноцистовый интервал (керестинская свита, обр. 117–135). Палинологические ассоциации внутри этого интервала разреза оказались крайне бедными, что не позволило нам диагностировать здесь все потенциально возможные диноцистовые события. В основании интервала отмечено появление вида Petalodinium waipawaense, совпадающее с HO наннопланктона Chiasmolithus gigas (основание подзоны CP13c), чуть выше по разрезу (обр. 120) выявлено первое появление диноцист Rhombodinium? aidae, совпадающее здесь с HO наннопланктона Nannotetrina quadrata (основание подзоны CP14a), чуть ниже LO наннопланктона Reticulofenestra umbilica. По присутствию видов Petalodinium waipawaense и Rhombodinium? aidae третий диноцистовый интервал в разрезе Хеу сопоставляется с диноцистовой зоной Enneadocysta pectiniformis зональной шкалы Восточного Пери-Тетиса (Яковлева, 2017). Возраст этой части керестинской свиты – поздний лютет.
Лишь средняя часть интервала охарактеризована количественно представительными палинологическими ассоциациями, почти полностью состоящими из диноцист. Образец 131 отличается доминированием (40%) Impagidinium; выше по разрезу преобладают ветзелиеллоидные (15–70%); представители группы Enneadocysta/Areosphaeridium составляют порядка 10–15%, при этом группа Spiniferites очень немногочисленна (от 10 до 0%). Скорее всего, в течение позднего лютета в рассматриваемой части морского бассейна произошли достаточно существенные изменения палеообстановок, выразившиеся в переходе от открыто-морских внешних неритических условий к более мелководным, возможно, с пониженной соленостью. Этим можно было бы объяснить исчезновение группы Impagidinium и доминирование ветзелиеллоидных (цист предположительно гетеротрофных видов) в обр. 134, в котором также наблюдается увеличение видового разнообразия наннопланктона и количества таких видов, как Cyclicargolithus floridanus и Ericsonia formosa, реже Thoracosphaera sp.
Четвертый диноцистовый интервал (кумская свита, обр. 136–144). Выявлен по появлению стратиграфически важного вида Rhombodinium draco в обр. 136 (зоны CP14a и NP15/16) и соответствует одноименной зоне шкалы Восточного Пери-Тетиса (Яковлева, 2017). Выше по разрезу последовательно появляются виды Petalodinium rhomboideum и Distatodinium biffii, при этом их появление отмечается ниже HOs наннопланктона Chiasmolithus solitus и Discoaster bifax. В верхней части кумской свиты (обр. 142), чуть ниже LO наннопланктона Chiasmolithus oamaruensis (основание зон CP15a и NP18), фиксируются первые появления диноцист Wetzeliella simplex, Enneadocysta inessae и Thalassiphora fenestrata. Первое появление вида Michouxdinium variabile (обр. 144) установлено здесь чуть выше LO наннопланктона Chiasmolithus oamaruensis (обр. 143). Исходя из данных изучения наннопланктона, часть кумской свиты, соответствующая четвертому диноцистовому комплексу, имеет бартонский и переходной бартон-приабонский возраст в понимании Ogg et al. (2016).
Палинологические ассоциации из количественно представительных образцов характеризуются существенным доминированием диноцист в обр. 137–138 (70–95%), тогда как в обр. 139–140 диноцисты не превышают 55%, но при этом значительно (до 35%) увеличивается количество двухмешковой пыльцы хвойных; в ассоциациях этого интервала постоянно отмечается присутствие (до 15%) внутренних камер фораминифер. Среди диноцист в обр. 137–138 преобладает группа Enneadocysta/Areosphaeridium (до 50%); Spiniferites, Impagidinium присутствуют постоянно (по 10–20%); в образце 140 резко увеличивается количество ветзелиеллоидных (до 35%). Скорее всего, эта часть кумской свиты могла формироваться в условиях прибрежной зоны, о чем свидетельствует существенная роль групп Enneadocysta/Areosphaeridium и ветзелиеллоидных, увеличение количества пыльцы хвойных деревьев, а также двукратное увеличение количества прибрежно-морского наннопланктона Zygrhablithus bijugatus.
Пятый диноцистовый интервал (кровля кумской свиты, обр. 145). Выявлен чуть ниже границы наннопланктонных зон NP18/NP19 или CP15ba/CP15b (LO Isthmolithus recurvus). Здесь отмечается первое появление диноцистового вида Reticulatosphaera actinocoronata, фиксирующего нижнюю границу одноименной зоны шкалы Восточного Пери-Тетиса (Яковлева, 2017).
Палинологический комплекс из обр. 145 отличается резким сокращением количества диноцист (35%) и существенным всплеском в содержании пыльцы покрытосеменных (25%) и хвойных (~20%), а также привносом празинофитов и спор наземных растений. Среди диноцист наблюдается резкий скачок в количестве ветзелиеллоидных (~40%), при этом группа Enneadocysta/Areosphaeridium составляет не более 15%, группа Spiniferites – не более 10%, Impagidinium и другие экогруппы – первые проценты. Исходя из этих характеристик палинологической ассоциации, можно предположить, что в конце кумского времени (начало приабона) продолжилось обмеление бассейна. В составе наннопланктонной ассоциации в этом интервале разреза доминируют роды Dictyococcites и Coccolithus, реже встречаются Discoaster и Sphenolithus, что, предположительно, может указывать на относительное снижение температуры поверхностных вод в конце кумского времени.
Шестой диноцистовый интервал (низы белоглинской свиты, обр. 146, 147) соответствует зоне Talladinium? angulosum Восточного Пери-Тетиса (Яковлева, 2017). Стратиграфически важный вид Talladinium? angulosum выявлен в обр. 147 выше LO наннопланктона Isthmolithus recurvus (основание зон NP19/20 и CP15b).
Низы белоглинской свиты характеризуются палинологическим комплексом, в котором вновь доминируют диноцисты (~80%), акритархи и празинофиты представлены первыми процентами, пыльца покрытосеменных и хвойных растений в целом составляет около 30%. В комплексе диноцист вновь увеличивается количество групп Spiniferites (30%) и Impagidinium (15%), почти отсутствуют перидиниоидные группы, что свидетельствует о начале накопления белоглинской свиты в период крупного трансгрессивного этапа. Подтверждением этому является и значительное возрастание видового разнообразия наннопланктона (61 видов) в обр. 146 с преобладанием среди них родов открыто-океанических условий (Dictyococcites, Discoaster, Coccolithus, Chiasmolithus и др.).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ И ЗАКЛЮЧЕНИЕ
Полученные в настоящей работе новые палинологические данные, сопоставленные с результатами изучения известкового наннопланктона, позволили выявить в разрезе интервалы шести диноцистовых зон шкалы Восточного Пери-Тетиса (Яковлева, 2017).
Так, в верхах черкесской свиты, соответствующей базальной части лютета (наннопланктонные подзоны NP14b, CP12a), установлена диноцистовая зона Enneadocysta arcuata, чья нижняя граница определяется первым появлением номинального вида. Первое появление Enneadocysta arcuata впервые использовалось в качестве важного события (датума) в Англии Дж. Итоном (Eaton, 1976), а затем стало определять основание зоны B-4 на юге Англии (Bujak et al., 1980). Согласно К. Кингу (King, 2016), LO Enneadocysta arcuata в Хэмпширском бассейне близко к границе наннопланктонных зон NP14/NP15. В Бельгии же первое появление этого вида отмечается внутри зоны NP14 (Heilmann-Clausen, Van Simaeys, 2005), что как раз соответствует данным, полученным в разрезе Хеу, и подтверждает возможную одновременность первого появления вида в самом начале лютета и в Пери-Тетисе, и в палеобассейне Северного моря.
Выше по разрезу, в кровле черкесской–низах керестинской свит раннелютетского возраста выявлен интервал диноцистовой зоны Costacysta bucina, установленной ранее в разрезе Актумсук в Узбекистане (Яковлева, 2017). Вид Costacysta bucina, описанный из отложений Дании (верхи Lillebælt Clay Formation и низы Søvind Marl Formation; Heilmann-Clausen, Van Simaeys, 2005), имеет лютетский интервал распространения и в бассейне Северного моря и на шельфе Фарерских островов (Waagstein, Heilmann-Clausen, 1995). Виды Wetzeliella ovalis, Castellodinium compactum, Vallodinium? echinosuturatum, появляющиеся внутри интервала зоны Costacysta bucina на Северном Кавказе и в Южном Устюрте, также отмечались ранее на юге бывшего СССР (Андреева-Григорович и др., 2011) и в разрезах Западной Европы (King, 2016) с раннего лютета (NP15), что свидетельствует о стратиграфическом потенциале использования LOs Wetzeliella ovalis, Castellodinium compactum и Vallodinium? echinosuturatum для датирования отложений и проведения межрегиональных корреляций. Следует отметить и появляющиеся чуть позднее, в интервале наннопланктонной подзоны CP13b, виды Enneadocysta partridgei и Enneadocysta robusta как на Северном Кавказе, так и в Южном Устюрте (Яковлева и др., 2019).
Исходя из данных изучения диноцист (по присутствию видов Costacysta bucina, Cordosphaeridium cantharellus, Wetzeliella ovalis), верхи черкесской–низы керестинской свит Северного Кавказа соответствуют нижней части бучакской свиты Днепровско-Донецкой впадины (Iakovleva, 2015), верхам тасаранской свиты в Северном Приаралье (Яковлева, 2017) а также части пачки А в разрезе Актумсук в Южном Устюрте (Яковлева и др., 2019).
Абсолютное доминирование диноцист над остальными группами палиноморф в комплексах раннелютетского возраста как в черкесской свите на Северном Кавказе, так и в нижней части бучакской свиты в Днепровско-Донецкой впадине и нижнелютетских отложениях Южного Устюрта позволяет говорить о крупном трансгрессивном этапе на территории Восточного Пери-Тетиса в начале лютета, который, вероятнее всего, последовал за регрессивным этапом на границе ипра/лютета, зафиксированном не только в Пери-Тетисе, но и в Западной Сибири (Яковлева, Александрова, 2014) и в палеобассейне Северного моря (King, 2006).
К сожалению, верхняя часть керестинской и нижняя часть кумской свит (диноцистовая зона Enneadocysta pectiniformis, наннопланктонные зоны NP15/16 и CP13c–CP14a) оказались неудачными для палинологического анализа: подавляющее большинство образцов либо содержали крайне малое количество палиноморф, либо вообще оказались стерильными. При этом отсутствие палиноморф в керестинской свите можно объяснить слишком большой карбонатностью отложений, что сказалось на сохранности фитопланктона, тогда как в кумской свите отмечается слишком большое количество аморфной органики. Тем не менее палинологический комплекс в интервале перехода от керестинской к кумской свите характеризуется абсолютным доминированием морского фитопланктона, однако в нем происходит четкая смена доминантов с гониаулакоидных (Impagidinium в кровле керестинской свиты) на перидиниоидные (wetzelielloids в основании кумской свиты), свидетельствующая о резкой смене обстановки осадконакопления.
Выше по разрезу в кумской свите установлена диноцистовая зона Rhombodinium draco. Следует отметить, что первое появление вида-индекса, фиксирующее нижнюю границу зоны, установлено здесь внутри интервала наннопланктонной зоны NP15/16 или подзоны CP14a, условно вблизи границы лютета/бартона. В Западной Европе LO Rhombodinium draco традиционно отмечают, начиная с зоны NP17 (Vandenberghe et al., 2012). Однако в Южной Англии первое появление этого вида происходит в зоне BAR-1 (Bujak et al., 1980) в слоях, отнесенных M.-P. Aubry (1983) к зоне NP16. Эту точку зрения подтверждают и новые данные А.И. Яковлевой, свидетельствующие о появлении Rhombodinium draco на юге Англии (Alum Bay) в конце интервала наннопланктонной зоны NP16 (Cotton et al., 2017). Что касается Пери-Тетиса, то LO Rhombodinium draco ранее уже было зафиксировано в лютетской части наннопланктонной зоны NP15/16 в отложениях бучакской свиты в Днепровско-Донецкой впадине (Iakovleva, 2015), а также в других разрезах Украины (Андреева-Григорович, неопубликованные данные).
Достаточно долго предполагалось, что стратиграфически важный вид Distatodinium biffii появляется в олигоцене (Van Simaeys et al., 2004). Однако по данным изучения опорных скважин Kysing-3 и Kysing-4 в Датском бассейне его первое появление было впоследствии зафиксировано стратиграфически гораздо ниже, в верхах Формации Søvind Marl под основанием формации Moesgaard Clay (приабон, уровень наннопланктонных зон NP19/20; Heilmann-Clausen, Van Simaeys, 2005), а позднее и в шубарсайской свите приабонского возраста в Прикаспийской впадине (при пропуске лютет-бартонской части керна) (Орешкина и др., 2015). В настоящее время наиболее раннее появление вида Distatodinium biffii отмечено внутри интервала зоны Rhombodinium draco в бартонских частях бучакской свиты в Днепровско-Донецкой скважине (Iakovleva, 2015) и кумской свиты в рассматриваемом здесь разрезе Хеу на Северном Кавказе.
Отдельного внимания заслуживает диноцистовая зона Reticulatosphaera actinocoronata, установленная в разрезе Хеу в кровле кумской свиты по первому появлению номинального вида. В Северо-Западной Европе, по данным из Датского бассейна, LO Reticulatosphaera actinocoronata фиксируется в настоящее время чуть выше основания наннопланктонной зоны NP19/20 (Heilmann-Clausen, Van Simaeys, 2005). В разрезе Хеу первое появление этого вида отмечается нами чуть ниже границы наннопланктонных зон NP18–NP19/20 или CP15a/CP15b, что совпадает и с данными из разреза Ланджар в Армении, где LO Reticulatosphaera actinocoronata выявлено немного выше LO Chiasmolithus oamaruensis (Щербинина и др., 2017).
Следует отметить, что в разрезе Хеу не выявлен интервал бартонской диноцистовой зоны Rhombodinium porosum, занимающей в шкале Восточного Пери-Тетиса (Яковлева, 2017) промежуточное положение между зонами Rhombodinium draco и Reticulatosphaera actinocoronata. Поскольку по данным изучения наннопланктона из отложений кумской свиты не зафиксировано существенного стратиграфического перерыва, вполне вероятно, что пропуск этой зоны связан с недостаточно детальным отбором образцов в этом интервале разреза Хеу.
Наконец, верхняя часть изученной нами эоценовой части разреза представлена низами белоглинской свиты, в которой установлена диноцистовая зона Talladinium? angulosum, соответствующая части наннопланктонных зон NP19/20 или CP15b. Белоглинская свита приабонского возраста коррелируется с одновозрастными обуховской свитой Украины (Андреева-Григорович и др., 2011; Iakovleva, 2015), чеганской свитой Приаралья и Устюрта (Запорожец, 1991; Яковлева, 1998), верхнетавдинской подсвитой Западной Сибири (Яковлева, Александрова, 2013) и отражает крупный трансгрессивный этап позднего эоцена.
Благодарности. Авторы благодарят Г.Н. Александрову (ГИН РАН) за помощь в химической обработке палинологических образцов, а также М.А. Ахметьева (ГИН РАН) и Н.К. Лебедеву (ИНГГ СО РАН) за конструктивные и полезные замечания.
Источники финансирования. Работа выполнена в рамках темы гос. заданий ГИН РАН №№ 0135-2016-0001, 0135-2018-0036, 0135-2019-0044, 0135-2019-0062.
Список литературы
Алимарина В.П. Некоторые особенности развития планктонных фораминифер в связи с зональным расчленением нижнего палеогена Северного Кавказа // Вопросы микропалеонтологии. 1963. Вып. 7. С. 158–197.
Андреева-Григорович А.С. Зональная стратиграфия палеогена юга СССР по фитопланктону (диноцисты и наннопланктон): Автореф. дисс. … д-ра геол.-мин. н-аук. Киев, 1991. 47 с.
Андреева-Григорович А.С., Запорожец Н.И., Шевченко Т.В., Александрова Г.Н., Васильева О.В., Яковлева А.И., Стотланд А.Б., Савицкая Н.А. Атлас диноцист палеогена Украины, России и сопредельных стран. Киев: Наукова Книга, 2011. 221 с.
Аристова К.Е. Пыльца и микропланктон из пограничных отложений эоцена и олигоцена южной части СССР // Палинология кайнофита. Москва: Наука, 1973. С. 117–121.
Ахметьев М.А., Запорожец Н.И. Смена диноцист в разрезах палеогена и нижнего миоцена Русской платформы, Крымско-Кавказской области и Туранской плиты как отражение экосистемных перестроек // Ископаемые микроорганизмы как основа стратиграфии, корреляции и палеобиогеографии фанерозоя. Ред. Кузнецова К.И., Музылев Н.Г. Вопросы микропалеонтологии. 1996. Вып. 31. С. 55–69.
Гроссгейм В.А. Палеоген Северо-Западного Кавказа // Тр. Краснодарского филиала Всесоюзного нефтегазового научно-исследовательского ин-та. 1960. Вып. 4. С. 3–190.
Запорожец Н.И. Палинокомплексы и фитопланктон верхнеэоценовых и олигоценовых отложений по опорной скважине № 3006 (Северо-Западное Приаралье) // Изв. АН КазССР. Сер. геол. 1991. № 1. С. 37–48.
Запорожец Н.И. Палинологические комплексы кумского горизонта среднего эоцена Кавказа и его возрастных аналогов в сопредельных регионах // Стратиграфия. Геол. корреляция. 2001. Т. 9. № 6. С. 83–103.
Крашенинников В.А., Музылёв Н.Г. Соотношение зональных шкал по планктонным фораминиферам и наннопланктону в разрезах палеогена Северного Кавказа // Вопросы микропалеонтологии. 1975. Вып. 18. С. 212–224.
Крашенинников В.А., Ахметьев М.А. (отв. редакторы). Геологические и биотические события позднего эоцена–раннего олигоцена на территории бывшего СССР. Часть II: Геологические и биотические события. М.: ГЕОС, 1998. 250 с. (Труды ГИН. Вып. 507).
Леонов Г.П., Алимарина В.П. Вопросы стратиграфии нижнепалеогеновых отложений Северо-Западного Кавказа. М.: Изд-во МГУ, 1964. 203 с.
Музылев Н.Г. Стратиграфия палеогена юга СССР по наннопланктону. М.: Наука, 1980. 83 с.
Орешкина Т.В., Яковлева А.И., Щербинина Е.А. Комплексный микропалеонтологический анализ эоценовых отложений восточного Прикаспия (скв. 57, Шубарсайская мульда, Казахстан) // Бюлл. МОИП. Отд. геол. 2015. Т. 90. Вып. 1. С. 42–80.
Попов С.В., Ахметьев М.А., Лопатин А.В., Бугрова Э.М., Сычевская Е.К., Щерба И.Г., Андреева-Григорович А.С., Запорожец Н.И., Николаева И.А., Копп М.Л. Палеогеография и биогеография бассейнов Паратетиса. Часть 1. Поздний эоцен–ранний миоцен. М.: Научный мир, 2009. 200 с.
Стратиграфия СССР. Палеогеновая система. Ред. Гроссгейм В.А., Коробков И.А. М.: Недра, 1975. 524 с.
Шуцкая Е.К. Стратиграфия нижних горизонтов палеогена Центрального Предкавказья по фораминиферам // Тр. Ин-та геол. наук АН СССР. Геол. сер. 1956. Вып. 164. № 71.
Шуцкая Е.К. Стратиграфия, фораминиферы и палеогеография нижнего палеогена Крыма, Предкавказья и западной части Средней Азии. М.: Недра, 1970. 255 с.
Щербинина Е.А., Яковлева А.И., Закревская Е.Ю. Зональная стратиграфия разреза Ланджар Южной Армении по наннопланктону, диноцистам и крупным фораминиферам // Стратиграфия. Геол. корреляция. 2017. Т. 25. № 5. С. 84–108.
Яковлева А.И. Эоценовые микрофоссилии из разреза Сары-Оба (северное Приаралье) // Бюлл. МОИП. Отд. геол. 1998. Т. 73. Вып. 3. С. 51–55.
Яковлева А.И. Детализация эоценовой диноцистовой шкалы для восточного Перитетиса // Бюлл. МОИП. Отд. геол. 2017. Т. 92. Вып. 2. С. 32–48.
Яковлева А.И., Александрова Г.Н. К вопросу об уточнении зонального деления по диноцистам палеоцен-эоценовых отложений Западной Сибири // Бюлл. МОИП. Отд. геол. 2013. Т. 88. Вып. 1. С. 59–82.
Яковлева А.И., Александрова Г.Н. Восстановление палеоэкологических обстановок люлинворского времени (эоцен) на юге Западно-Сибирского морского бассейна по палинологическим данным // Бюлл. МОИП. Отд. геол. 2014. Т. 89. Вып. 3. С. 33–50.
Яковлева А.И., Щербинина Е.А., Музылев Н.Г., Александрова Г.Н. Диноцисты среднего–верхнего эоцена разреза Актумсук, Устюрт, Узбекистан: биостратиграфия и палеообстановки // Стратиграфия. Геол. корреляция. 2019. Т. 27. № 6. С. 78–102.
Aubry M.-P. Biostratigraphie du Paléogène epicontinental de l’Europe du Nord-Ouest. Etude fondée sur les nannofossiles calcaires // Documents des Laboratoires de Géologie de Lyon 89. 1983. P. 1–317.
Brinkhuis H. Late Eocene to early Oligocene dinoflagellate cysts from the Priabonian type-area (Northeast Italy): biostratigraphy and paleoenvironmental interpretation // Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994. V. 107. P. 121–163.
Bujak J.P., Downie D., Eaton G.L., Williams G.L. Dinoflagellate cysts and acritarchs from the Eocene of southern England // Spec. Pap. Palaeontol. 1980. № 24. 100 p.
Cotton L.J., Riveiro Cuesta L., Franceschetti G., Iakovleva A., Hooker J., King Ch., Alegret L., Dinares-Turell J., Yager S., Fluegeman R., Monechi S. Reassessing the Bartonian unit stratotypes: an integrated approach // GSA 2017 Annual Meeting. Seattle, Washington, 2017. V. 49. № 6. https://doi.org/10.1130/abs/2017AM-305810
Crouch E.M., Brinkhuis H. Environmental change across the Paleocene–Eocene transition from eastern New Zealand: a marine palynological approach // Marine Micropaleontology. 2005. V. 56. P. 138–160.
Eaton G.L. Dinoflagellate cysts from the Bracklesham Beds (Eocene) of the Isle of Wight, southern England // Brit. Mus. (Nat. Hist.) Geol. Bull. 1976. V. 26. P. 227–332.
Gavrilov Yu.O., Shcherbinina E.A., Muzylöv N.G. A Paleogene sequence in central north Caucasus: a response to paleoenvironmental changes // GFF. 2000. V. 122. P. 51–53.
Heilmann-Clausen C., Van Simaeys S. Dinoflagellate cysts from the Middle Eocene to ?lowermost Oligocene succession in the Kysing Research borehole, central Danish Basin // Palynology. 2005. V. 29. P. 143–204.
Iakovleva A.I. Middle–late Eocene dinoflagellate cysts from NE Ukraine (Borehole No. 230, Dnepr-Donets Depression): stratigraphical and palaeoenvironmental approach // Acta Palaeobotanica. 2015. V. 55(1). P. 19–51.
King C. Paleogene and Neogene: uplift and a cooling climate // The Geology of England and Wales. Second edition. Geological Society, London, 2006. P. 395–427.
King C. A revised correlation of Tertiary rocks in the British Isles and adjacent areas of NW Europe // Geol. Soc. London. Spec. Rep. 2016. V. 27. 719 p.
Martini E. Standard Tertiary and Quaternary calcareous nannoplankton zonation // Proc. II Planktonic Conf., Roma, 1970. Rome: Edizioni Tecnoscienza, 1971. V. 2. P. 739–785.
Ogg J.G., Ogg F.M., Gradstein F.M. A concise Geologic Time Scale 2016. Elsevier, 2016. 229 p.
Okada H., Bukry D. Supplementary modification and introduction of code numbers of the low-latitude coccolith biostratigraphic zonation (Bukry 1973, 1975) // Marine M-icropaleontology. 1980. V. 5. P. 321–325.
Powell A.J., Brinkhuis H., Bujak J.P. Upper Paleocene–lower Eocene dinoflagellate cyst sequence biostratigraphy of southeast England // Correlation of the Early Paleogene in Northwest Europe. Geol. Soc. Spec. Publ. 1996. V. 101. P. 145–183.
Radionova E.P., Aleksandrova G.N., Gavtadze T.T., Stupin S.I., Khokhlova I.E. Analysis of Late Paleocene–Early Eocene Micropankton from the Kheu River Section, West Pre-Caucasus // Climatic and Biotic Events of the Paleogene (CBEP 2009). Int. Conf., Wellington, New Zealand, January 12–15, 2009. Extended Abstracts. Eds. Crouch E.M., Strong C.P., Hollis C.J. GNS Science Miscellaneous Series. 2009. V. 18. P. 111–116.
Shcherbinina E.A. Middle Eocene nannofossils and geological events of the northeastern Peri-Tethys // GFF. 2000. V. 122. P. 143–145.
Shcherbinina E., Gavrilov Yu., Iakovleva A., Pokrovsky B., Golovanova O., Aleksandrova G. Environmental dynamics during the Paleocene–Eocene thermal maximum (PETM) in the northeastern Peri-Tethys revealed by high-resolution micropalaeontological and geochemical studies of a Caucasian key section // Palaeogeogr. Palaeoclimat. Palaeoecol. 2016. V. 456. P. 60–81.
Shcherbinina E.A., Iakovleva A.I., Gavrilov Yu.O., Muzylöv N.G., Golovanova O.V. Lithological architecture and biostratigraphy of the lower Eocene sediments of the central northern Caucasus // Geologica Acta (in press).
Sluijs A., Pross J., Brinkhuis H. From greenhouse to icehouse: organic-walled dinoflagellate cysts as paleoenvironmental indicators in the Paleogene // Earth-Sci. Rev. 2005. V. 68. P. 281–315.
Torricelli S., Knezaurek G., Biffi U. Sequence biostratigraphy and paleoenvironmental reconstruction in the Early Eocene Figols Group of the Tremp–Graus Basin (south-central Pyrenees, Spain) // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006. V. 232(1). P. 1–35.
Van Simaeys S., De Man E., Vandenberghe N.L., Brinkhuis H., Suetrbaut E. Stratigraphic and palaeoenvironmental analysis of the Rupelian–Chattian transition in the type region: evidence from dinoflagellate cysts, foraminifera and calcareous nannofossils // Palaeogeogr. Palaeoclimat. Palaeoecol. 2004. V. 208. P. 31–58.
Vandenberghe N., Hilgen F.J., Speijer R.P. The Paleogene Period // The Geologic Time Scale 2012. Cambridge: Cambridge University Press, 2012. P. 855–921.
Waagstein R., Heilmann-Clausen C. Petrography and biostratigraphy of Palaeogene volcaniclastic sediments dredged from the Faeroes shelf // The Tectonics, Sedimentation and Palaeoceanography of the North Atlantic Region. Geol. Soc. Spec. Publ. 1995. № 90. P. 179–197.
Waga D.D., Andreeva-Grigorovich A.S., Mintuzova L.G., Mutterlose J., Suprun I.S., Bornemann A. Calcareous nannofossil and benthic/planktonic foraminifera biostratigraphy of Eocene sediments of the Kheu stratotype section of Northern Caucasus (Russia) // J. Nannoplankton Res. (in press).
Williams G.L., Fensome R.A., MacRae R.A. The Lentin and Williams index of fossil dinoflagellates 2017 edition // Am. Assoc. Stratigr. Palynol. Contrib. Ser. 2017. № 48.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция