

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 3, стр. 102-113
Особенности развития радиолярий в кампане и маастрихте и перспективы детальной стратиграфии
Л. Г. Брагина 1, *, Н. Ю. Брагин 1
1 Геологический институт РАН
Москва, Россия
* E-mail: l.g.bragina@mail.ru
Поступила в редакцию 23.10.2018
После доработки 12.02.2019
Принята к публикации 28.03.2019
Аннотация
Комплексы кампанских и маастрихтских радиолярий Тетической надобласти известны во многих регионах, но лишь для некоторых из них детально обоснован возраст и проведено полноценное палеонтологическое изучение. На основании данных по датированным и детально изученным кампан-маастрихтским радиоляриям Крыма, Кипра, Сербии и с привлечением материалов по Калифорнии, Мировому океану, Турции и другим районам проведен анализ развития группы. Показано, что в течение кампана и маастрихта происходило сравнительно медленное обновление сообществ, причем биоразнообразие на уровне родов постепенно сокращалось. История развития радиолярий в конце позднего мела разделена на этапы: позднесантонско-раннекампанский, раннекампанско-позднекампанский, позднекампанско-раннемаастрихтский и позднемаастрихтский.
ВВЕДЕНИЕ
В настоящее время для верхнего альба–сантона Тетических районов Евразии разработана детальная зональная шкала по радиоляриям (Брагина, 2016а). Верхней зоной этой шкалы является Afens perapediensis. Впервые изученные радиолярии и планктонные фораминиферы из пограничных отложений сантона и кампана Горного Крыма позволили расширить объем этой зоны до верхней части верхнего сантона–нижнего кампана (Брагина и др., 2016).
Между тем значительная часть кампанского яруса и весь маастрихт остаются за пределами новой зональной шкалы (рис. 1). Для расчленения здесь могут использоваться лишь разработанные несколько десятилетий назад схемы. Однако эти построения в ряде случаев имеют лишь региональное значение и не могут широко применяться (Pessagno, 1976) либо построены на недостаточном материале и являются неточными (Sanfilippo, Riedel, 1985). Детальный обзор ранее разработанных зональных схем верхнего мела по радиоляриям был проведен ранее Л.Г. Брагиной (2016а), показавшей невозможность практического применения этих схем для решения современных задач детальной стратиграфии и широкой межрегиональной корреляции верхнего мела по радиоляриям.
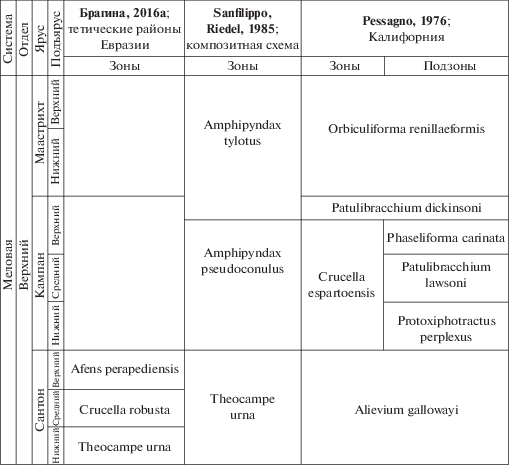
Поэтому важнейшей целью дальнейших исследований радиолярий мела является разработка зонального деления кампана и маастрихта на тех же принципах, которые использовались для альб-сантонской шкалы (Брагина, 2016а): таксономически полное изучение представительных комплексов радиолярий в разрезах разнофациальных толщ (от шельфовых до глубоководных) совместно с фоссилиями, обеспечивающими точную датировку и детальное расчленение разрезов. Наиболее перспективны здесь планктонные фораминиферы, которые изучаются давно, считаются одной из важнейших групп в детальной стратиграфии верхнего мела и широко распространены в разрезах, в том числе совместно с радиоляриями.
Основной сложностью в настоящее время является недостаточность материала. Авторы статьи пока не располагают фактическим материалом из разрезов кампана–маастрихта, в которых весь стратиграфический интервал был бы охарактеризован радиоляриями. Литературные данные также неполны. Поэтому на начальной стадии проведения исследования радиолярий требуется решить ряд задач, без которых дальнейший прогресс будет невозможен. К ним относятся следующие задачи:
1. Систематизация известных литературных, а также оригинальных данных по представительным комплексам радиолярий хорошей сохранности, встреченным совместно с планктонными фораминиферами или иными фоссилиями, позволяющими точно датировать обнаруженные радиоляриевые комплексы.
2. Анализ литературных и оригинальных данных, включающий общий обзор развития радиолярий в течение кампана–маастрихта, основные тенденции в морфофилогенезе важнейших таксонов и в изменениях общего биоразнообразия радиолярий.
3. Установление этапности развития радиолярий в кампане и маастрихте. Определение наиболее важных рубежей смены комплексов.
Решение этих задач позволит получить результаты, необходимые для разработки будущей схемы зонального деления кампана–маастрихта, и определить наиболее важные направления дальнейших работ, в частности выявление интервалов и уровней, где особенно важно получить дополнительный фактический материал. Данная статья посвящена обзору имеющихся данных по радиоляриям кампана и маастрихта и оценке перспектив построения новой детальной зональной шкалы для этого интервала.
ХАРАКТЕРНЫЕ КОМПЛЕКСЫ РАДИОЛЯРИЙ КАМПАНА И МААСТРИХТА
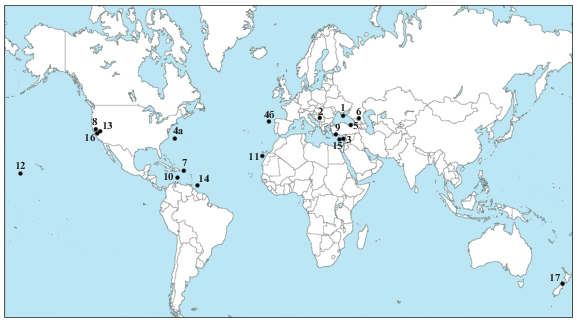
В настоящее время известно большое количество публикаций по радиоляриям кампана и маастрихта, однако данные, изложенные в них, неравноценны. Связано это с двумя факторами. Во-первых, многие комплексы обнаружены в толщах, не охарактеризованных другими фоссилиями, и потому их стратиграфическое положение не вполне ясно. Во-вторых, таксономический состав радиолярий в имеющихся публикациях часто ограничивается несколькими видами или десятком видов, а это означает, что комплекс беден и, следовательно, его ценность невелика. И наконец, в-третьих, иногда автор изображает или приводит в списке лишь некоторые виды из состава комплекса, и тогда эти данные трудно использовать, поскольку они неполные. Поэтому основное внимание в данном обзоре будет уделено детально изученным комплексам радиолярий, встреченным совместно с другими фоссилиями, особенно с ортостратиграфическими. Обзор будет ограничен тетическими и близкими к ним комплексами, наиболее важными для разработки зонального деления. Высокоширотные ассоциации представляют предмет отдельного анализа, выходящего за рамки данной работы. Обзор публикаций дополнен собственными материалами авторов, в результате чего охвачены многие разрезы и местонахождения из разных, нередко весьма удаленных друг от друга регионов (рис. 2).
Рис. 2.
Важнейшие местонахождения радиолярий кампана и маастрихта в Тетической палеобиогеографической надобласти. 1 – Горный Крым, верхний сантон–нижний кампан (зона Afens perapediensis) (Корчагин и др., 2012; Брагина, 2016а; Брагина и др., 2016); 2 – Сербия, верхний сантон–нижний кампан (зона Afens perapediensis) (Брагина и др., 2014, 2018; Брагина, 2016а; Брагина и др., в печати); 3 – Кипр, верхний сантон–нижний кампан (зона Afens perapediensis) (Bragina, 2012; Брагина, 2016а); 4 – Северная Атлантика (4а – континентальный склон восточнее мыса Гаттерас, Северная Каролина; 4б – Галисийское плато), верхний сантон–нижний кампан (зона Afens perapediensis) (Thurow, 1988; Брагина, 2016а); 5 – Северо-Восточная Турция, нижний кампан (Soycan, Hakyemez, 2018); 6 – Северный Кавказ, р. Урух, предположительно нижний кампан (Вишневская, 2001); 7 – Пуэрто-Рико, нижний кампан (Pessagno, 1963); 8 – Калифорния, кампан и маастрихт (Pessagno, 1976); 9 – Южная Турция, кампан (Moix et al., 2009); 10 – Карибское море, скв. 146, кампан (Riedel, Sanfilippo, 1974); 11 – Атлантический океан, скв. 369, кампан и маастрихт (Foreman, 1978); 12 – Тихий океан, скв. 313, верхний кампан (Empson-Morin, 1981); 13 – Калифорния, верхний кампан?–нижний маастрихт (Campbell, Clark, 1944); 14 – Атлантический океан, скв. 144, маастрихт (Petrushevskaya, Kozlova, 1972); 15 – Кипр, верхний кампан–нижний маастрихт (Urquhart, Banner, 1994; Брагина, Брагин, 1995); 16 – Калифорния, верхний маастрихт (Foreman, 1968); 17 – Новая Зеландия, маастрихт (Hollis, 1997).
Нижний рубеж кампана по радиоляриям в настоящее время проследить сложно. В недавно разработанной зональной схеме (Брагина и др., 2018) граница сантона и кампана проходит внутри зоны Afens perapediensis. Соответственно, обзор следует начать с известных комплексов этой зоны, имеющих позднесантонский–раннекампанский возраст.
Комплексы позднесантонского–раннекампанского возраста (зона Afens perapediensis) известны на территории Горного Крыма (Корчагин и др., 2012; Брагина, 2016а; Брагина и др., 2016) (рис. 2). В нижней части зоны (верхний сантон) комплекс содержит более 60 видов. Среди них присутствуют таксоны, существование которых, как ранее считалось, начинается с кампана: Conocaryomma dauerhafta Empson-Morin, Spongosaturninus parvulus var. lateralispinosus Campbell et Clark, Clathropyrgus titthium Riedel et Sanfilippo, Dictyomitra koslovae Foreman, Eucyrtidium (Eucyrtis) carnegiense Campbell et Clark, Theosyringium jugosum Campbell et Clark, Pseudoaulophacus pargueraensis Pessagno.
Раннекампанский комплекс, за исключением Alievium superbum (Squinabol) и Crucella messinae Pessagno, охарактеризован теми же видами, что и позднесантонский, а также вновь появившимися: Prunobrachium sp. ex gr. P. crassum (Lipman), P. sp. ex gr. P. spongiosum (Lipman), Spongosaturninus parvulus Campbell et Clark, Diacanthocapsa acanthica Dumitrica, Dictyocephalus (Dictyocryphalus) (?) legumen Campbell et Clark, ?Sciadiocapsa causia Foreman, Stichomitra cechena Foreman и Stichopilium (Stichopilidium) teslaense Campbell et Clark, при отсутствии вида Amphipyndax pseudoconulus Pessagno.
Разрезы Горного Крыма особенно ценны тем, что в них имеются многочисленные уровни, охарактеризованные и радиоляриями, и фораминиферами (планктонными и бентосными) хорошей сохранности, причем все эти группы микрофоссилий, как правило, встречаются совместно, в одних и тех же образцах. К сожалению, более молодые кампанские и маастрихтские отложения Горного Крыма не содержат радиолярий.
Кроме того, комплексы зоны Afens perapediensis известны на территории Сербии (Брагина и др., 2014, 2018; Брагина, 2016а; Брагина и др., в печати) (рис. 2). Следует особо отметить, что эти комплексы почти полностью представлены видами, распространенными и в разрезах Горного Крыма. Планктонные фораминиферы также подтверждают верхнесантонско-нижнекампанский стратиграфический интервал. Более молодые комплексы радиолярий в Сербии пока не известны.
Заслуживает внимания то, что и в Сербии в низах кампана отсутствует вид Amphipyndax pseudoconulus (Pessagno). Некоторые авторы считают, что первое появление данного вида связано с основанием кампана (Hollis, Kimura, 2001; Moix et al., 2009), но это не подтверждается результатами, полученными при совместном изучении радиолярий и планктонных фораминифер Горного Крыма и Сербии. Вероятно, вид Amphipyndax pseudoconulus появляется позднее, в течение кампана.
Особняком стоит чрезвычайно богатый комплекс зоны Afens perapediensis на Кипре (Bragina, 2012; Брагина, 2016а) (рис. 2), насчитывающий около 100 видов, причем здесь встречаются практически все виды, известные в Крыму. Однако на Кипре данная зона охарактеризована только радиоляриями, и поэтому не вполне ясно, включает ли она нижний кампан.
Зона A. perapediensis прослеживается и в Северной Атлантике, в районе Галисийского плато и на континентальном склоне восточнее мыса Гаттерас, Северная Каролина, где представлена типичнейшими для этой зоны видами (Thurow, 1988) (рис. 2): Alievium gallowayi (White), Crucella (?) sp. A = Cr. robusta Bragina, Patellula verteroensis (Pessagno), Pseudoaulophacus floresensis Pessagno, Pseudoaulophacus lenticulatus (White), Afens liriodes Riedel et Sanfilippo = Afens perapediensis Bragina, Clathropyrgus titthium Riedel et Sanfilippo, Dictyomitra koslovae Foreman, Eucyrtidium (?) sp. C. = Pseudoeucyrtis belogorskensis Bragina, Heliocryptocapsa sp. A, Obesacapsula (?) sp. A. (= Stichomitra sp. A; Корчагин и др., 2012). Данный комплекс отнесен Ю. Туровым (Thurow, 1988) к зоне кампана Amphipyndax pseudoconulus (Sanfilippo, Riedel, 1985). Однако следует заметить, что экземпляр, определенный как Amphipyndax pseudoconulus (Pessagno) (Thurow, 1988, pl. 1, fig. 8 ), не имеет характерных для этого вида отчетливых узелков на межкамерных пережимах и весьма сходен с морфотипами, известными в среднем–верхнем сантоне и нижнем кампане Горного Крыма, Сербии и Кипра, например с Amphipyndax sp. A (Корчагин и др., 2012, табл. IX , фиг. 6 ; Брагина и др., 2014, табл. I , фиг. 11, 12 ; Брагина и др., в печати, табл. I , фиг. 6–8 ). Из-за этого интерпретация возраста комплекса, изученного Ю. Туровым в Северной Атлантике, спорна.
Большую ценность представляют работы, в которых интерпретации возраста по радиоляриям подтверждены находками ортостратиграфических фоссилий. В недавней публикации по Северо-Восточной Турции (Soycan, Hakyemez, 2018) (рис. 2) представлены раннекампанские радиолярии и планктонные фораминиферы, найденные совместно на нескольких уровнях разреза Койонджулар. К сожалению, судя по приведенным данным, комплекс радиолярий не отличается значительным таксономическим разнообразием и содержит не более двух десятков видов, среди которых отсутствуют виды-индексы зональной шкалы (Sanfilippo, Riedel, 1985), в частности Amphipyndax pseudoconulus (Pessagno).
К нижнему кампану, вероятно, принадлежат комплексы, описанные В.С. Вишневской в разных районах Большого Кавказа: на р. Урух (рис. 2), р. Туапсе и р. Агура (Вишневская, 2001). Они отнесены к радиоляриевой зоне Dictyomitra torquata, которая характеризуется видами Dictyomitra torquata Foreman, D. koslovae Foreman, Stichomitra livermorensis (Campbell et Clark). В этом небогатом комплексе отсутствуют характерные виды зоны Afens perapediensis, за исключением Dictyomitra koslovae и Eucyrtis sp. = Pseudoeucyrtis belogorskensis Bragina, и поэтому точно сопоставить его с другими раннекампанскими комплексами пока сложно.
Раннекампанский возраст имеет комплекс радиолярий из Пуэрто-Рико (Pessagno, 1963) (рис. 2), характеризующийся присутствием Amphipyndax pseudoconulus совместно с Pseudoaulophacus floresensis Pessagno, P. pargueraensis Pessagno, P. lenticulatus (White), P. gallowayi (White), Holocryptocapsa (?) sphaerica (White) = Cryptamphorella sphaerica (White). Судя по присутствию A. pseudoconulus, данный комплекс может быть моложе комплексов верхов зоны Afens perapediensis и, вероятно, встречается в верхней части нижнего кампана–среднем кампане.
В Калифорнии выделяются последовательные комплексы радиолярий кампана (Pessagno, 1976) (рис. 2). Нижнекампанский комплекс описан из нижней части зоны Crucella espartoensis и характеризуется появлением, наряду с видом-индексом, таких таксонов, как Protoxiphotractus perplexus Pessagno, P. kirbyi Pessagno, Parvicuspis colusaensis Pessagno, Sciadiocapsa (?) rumseyensis Pessagno. Здесь необходимо отметить, что Э. Пессаньо (Pessagno, 1971) указывал на присутствие самых ранних представителей Crucella espartoensis в среднем сантоне, другие же виды слабо известны за пределами Северной Америки. В более молодых комплексах кампана в Калифорнии представлены Pseudoaulophacus delvallensis Pessango, Spongosaturninus ellipticus Campbell et Clark, S. hueyi Pessagno, Peritiviator labyrinthi Pessagno, Neosciadiocapsa diabloensis Pessagno, Bisphaerocеphalina (?) amazon Foreman (Pessagno, 1976).
Недавно был обнаружен хорошо сохранившийся и таксономически разнообразный кампанский комплекс в Южной Турции, на побережье Анталии (Moix et al., 2009) (рис. 2). Это типичный тропический комплекс, включающий вид-индекс и сопутствующие виды зоны Amphipyndax pseudoconulus (Sanfilippo, Riedel, 1985), в том числе Pseudoaulophacus floresensis Pessagno, P. pargueraensis Pessagno, Clathropyrgus titthium Riedel et Sanfilippo, Dictyomitra koslovae Foreman, а также характерные для кампана Botryometra heros (Campbell et Clark), Dictyoprora salillum (Foreman) и Foremanina schona Empson-Morin. Однако в этом местонахождении неизвестны другие фоссилии, которые бы подтверждали возрастную интерпретацию комплекса. Можно лишь предполагать, что этот комплекс относится к более высокой части кампана, и об этом говорит присутствие типичных Amphipyndax pseudoconulus.
Кампанские комплексы с Amphipyndax pseudoconulus известны из многих местонахождений донных осадков Мирового океана (Riedel, Sanfilippo, 1974; Foreman, 1978; Sanfilippo, Riedel, 1985). В качестве примера можно привести скв. 146 в Карибском море (Riedel, Sanfilippo, 1974) (рис. 2). Здесь, помимо вида-индекса, отмечаются Pseudoaulophacus floresensis Pessagno, P. pargueraensis Pessagno, Theocapsomma comys Foreman, Theocampe apicata Foreman и другие виды. В этом интервале наблюдается также последнее появление Clathropyrgus titthium Riedel et Sanfilippo. Необходимо отметить недостаточную изученность как самих комплексов (обычно изображенных лишь в проходящем свете, что не всегда обеспечивает уверенное определение видов), так и стратиграфического распространения этих видов, что уже отмечалось (Брагина, 2016а).
Несомненно позднекампанским является комплекс из скв. 313 Центральной Пацифики (Empson-Morin, 1981). Его возраст подтвержден совместным нахождением с фораминиферами, отнесенными к подзоне Globotruncana elevatа и зонуле Globotruncana calcarata (Pessagno, 1967), а также с позднекампанским наннопланктоном. В составе этого комплекса радиолярий встречаются: Pseudoaulophacus vielseitigus Empson-Morin, Theocampe altamontensis (Campbell et Clark), T. apicata Empson-Morin, T. salilllum Foreman = Dictyoprora salillum (Foreman), T. ixys Riedel et Sanfilippo, Krempelinella kronenartiga Empson-Morin, Rhopalosyringium kleinum Empson-Morin, R. magnificum Campbell et Clark, Foremanina shona Empson-Morin и другие. Как можно видеть, в позднем кампане состав комплексов радиолярий значительно обновляется.
Комплексы радиолярий верхов кампана и маастрихта из донных осадков океанов известны во многих регионах, но пока недостаточно изучены. В верхах кампана–маастрихте в тропических областях Мирового океана выделена зона Amphipyndax tylotus (Sanfilippo, Riedel, 1985). В составе комплекса этой зоны представлены, помимо вида-индекса, Archaeodictyomitra lamellicostata (Foreman), Siphocampe bassilis Foreman, Amphipyndax pseudoconulus (Pessagno), Theocampe salilum Foreman = Dictyoprora salillum (Foreman), T. apicata Foreman, Pseudoaulophacus floresensis Pessagno, Clathropyrgus titthium Riedel et Sanfilippo, Afens lirioides Riedel et Sanfilippo и др. (Foreman, 1968, 1978; Riedel, Sanfilippo, 1974; Sanfilippo, Riedel, 1985).
Достаточно полное представление о раннемаастрихтских (возможно, позднекампанских) комплексах можно получить из ранней работы по верхнемеловым радиоляриям Калифорнии (Campbell, Clark, 1944). Следует заметить, что специалисты, занимающиеся кампаном и маастрихтом, постоянно цитируют эту работу, так как в ней приводится описание видов, которые широко распространены за пределами Калифорнии, где они были впервые описаны, например: Lithomelissa (Micromelissa) heros Campbell et Clark = Botryometra heros (Campbell et Clark), Rhopalosyringium magnificum Campbell et Clark, Tricolocampe (Tricolocamptra) altamontensis Campbell et Clark = Siphocampe altamontensis (Campbell et Clark), Theocampe vanderhoofi Campbell et Clark, T. latipunctata Campbell et Clark, Lithomitra (Lithometrissa) regina Campbell et Clark = Archaeodictyomitra regina (Campbell et Clark), Lithocampe (Lithocampanula) andersoni Campbell et Clark = Dictyomitra andersoni (Campbell et Clark), Artocapsa livermorensis Campbell et Clark = Stichomitra livermorensis (Campbell et Clark). Таким образом, этот комплекс можно сопоставить по видовому составу с комплексом зоны Orbiculiforma renillaeformis (маастрихт; Pessagno, 1976).
Маастрихтский комплекс радиолярий известен в экваториальной части западной и восточной Атлантики (Petrushevskaya, Kozlova, 1972) (рис. 2). Здесь сосуществуют Amphipyndax enessefi Foreman = Amphipyndax pseudoconulus (Pessagno), Lithomelissa hoplites Foreman, Rhopalosyringium colpodes Foreman, Theocampe bassilis Foreman, Theocapsomma comys Foreman, Theocampe ? daseia Foreman. К сожалению, в этой работе радиолярии сфотографированы в проходящем свете и нуждаются в переизучении.
Комплексы позднего кампана–раннего маастрихта описаны на Кипре (Urquhart, Banner, 1994; Брагина, Брагин, 1995, 2006) (рис. 2). Здесь совместно с планктонными фораминиферами из верхов верхнего кампана и нижнего маастрихта (Корчагин, 2011) встречаются Alievium gallowayi (White), A. praegallowayi Pessagno, A. vielseitigus Empson-Morin, Archaeospongoprunum nishiyamae Nakaseko et Nishimura, A. salumi Pessagno, Hexapyramis sp. cf. H. perforatum Bragina, Patellula euessceei Empson-Morin, P. verteroensis (Pessagno), Praeconocaryomma universa Pessagno, Pseudoaulophacus floresensis Pessagno, P. lenticulatus (White), P. pargueraensis Pessagno, P. praefloresensis Pessagno, P. riedeli Pessagno, Afens liriodes Riedel et Sanfilippo, Amphipyndax sp. ex gr. A. pseudoconulus (Pessagno), A. tylotus Foreman, Artostrobium urna Foreman = Theocampe urna (Foreman), Botryometra heros (Campbell et Clark), Clathropyrgus titthium Riedel et Sanfilippo, Cryptamphorella macropora Dumitrica, Lithocampe eureia Foreman, Podocapsa (?) topferia Empson-Morin, Siphocampe daseia (Foreman) и другие виды. Кроме того, кипрские комплексы имеют большое сходство с известными низкоширотными ассоциациями донных осадков океанов и Калифорнии. Интересно отметить, что в значительно более северных, чем Кипр, районах (юг России, Ростовская область) в пределах верхнего кампана–нижнего маастрихта прослеживаются некоторые виды, характерные для низких широт и включающие вид-индекс зоны Amphipyndax tylotus (Беньямовский и др., 2012).
Достаточно полное представление о самых молодых мезозойских комплексах Калифорнии дает публикация (Foreman, 1968) (рис. 2), посвященная позднемаастрихтским радиоляриям. Эта работа до сих пор остается наиболее значимой (описано 8 родов и 77 видов, из них 56 – новые) и постоянно цитируемой. Для упомянутых комплексов характерно присутствие таких видов, как Acidnomelos apteron Foreman, Clathrocyclas diceros Foreman, C. hyronia Foreman, Dictyomitra lamellicostata Campbell et Clark = Archaeodictyomitra lamellicostata (Campbell et Clark), Myllocercion acineton Foreman, Stichomitra alamedaensis (Campbell et Clark), S. livermorensis (Campbell et Clark), Theocampe altamontensis (Campbell et Clark) = Siphocampe altamontensis (Campbell et Clark), Theocapsomma amphora Campbell et Clark.
Необходимо отметить многочисленные публикации по радиоляриям кампана и маастрихта Японии (Okamura, 1980; Nakaseko, Nishimura, 1981; Iwata, Tajika, 1986; Yamasaki, 1987; Takahashi, Ishii, 1995; Hashimoto, Ishida, 1997 и многие другие). Среди них наибольший интерес вызывают работы по южной части Японии, где выявлены таксоны, характерные для Тропической надобласти, а потому прослеживаются зоны Amphipyndax pseudoconulus и Amhipyndax tylotus (Sanfilippo, Riedel, 1985). К сожалению, сложное геологическое строение Японии и, как правило, отсутствие других групп микрофоссилий не позволяют уточнить стратиграфическое распространение видов и стратиграфическое положение комплексов.
Более северные районы Японии характеризуются комплексами радиолярий, близкими к калифорнийским. Например, на севере Хоккайдо кампанские комплексы содержат Archaeodictyomitra regina (Campbell et Clark), Cornutella californica Campbell et Clark, Dictyomitra cf. andersoni Foreman, Spongosaturnalis spinifer Campbell et Clark, Stichomitra livermorensis (Campbell et Clark) (Iwata, Tajika, 1986). Специфичность комплексов заключается в отсутствии видов, позволяющих напрямую соотнести их с одновозрастными комплексами низких широт.
Особый интерес вызывает комплекс радиолярий верхов кампана и маастрихта Новой Зеландии (Hollis, 1997) (рис. 2). Хотя он найден за пределами Тетической надобласти, его следует рассмотреть, поскольку это на сегодняшний день единственная находка маастрихтского комплекса, сменяемого нижнепалеоценовым в непрерывном разрезе пограничных слоев мела и палеогена. Новозеландский комплекс характеризуется присутствием ряда видов, широко распространенных в верхнем кампане–маастрихте Калифорнии и тропической части Пацифики, в том числе Orbiculiforma renillaeformis (Campbell et Clark), Dictyomitra andersoni (Campbell et Clark), Lithomelissa? hoplites Foreman, Siphocampe altamontensis (Campbell et Clark), Theocapsomma amphora (Campbell et Clark), T. erdnussa (Empson-Morin) и Myllocercion acineton Foreman (Campbell, Clark, 1944; Foreman, 1968; Pessagno, 1976; Empson-Morin, 1981). В разрезах Новой Зеландии в интервале распространения данного комплекса выделяется интервал-зона по радиоляриям Lithomelissa? hoplites, которая коррелируется с зоной Abathomphalus mayoroensis (планктонные фораминиферы). Необходимо подчеркнуть, что в пределах зоны Lithomelissa? hoplites продолжают существование многие виды, известные с конца кампана, некоторые из них даже продолжают встречаться в низах палеоцена.
РАЗВИТИЕ РАДИОЛЯРИЙ В КАМПАНЕ И МААСТРИХТЕ
При рассмотрении динамики развития групп органического мира и их биоразнообразия часто применяются количественные показатели. Этот метод используется и в данной статье.
Темпы изменений таксономического состава комплексов и их количественные показатели в кампане и маастрихте уже подсчитаны для высоких широт, где ассоциации радиолярий не столь таксономически разнообразны, как в низких широтах: с начала кампана к границе с маастрихтом резко возрастает количество видов, с начала маастрихта к концу маастрихта видовое разнообразие постоянно снижается и становится меньше, чем в начале кампана (Вишневская, 2009).
Для низких широт, а также для областей, испытывающих их непосредственное влияние, в последние десятилетия продолжается постоянное и значительное пополнение данных по видовому разнообразию ассоциаций позднего сантона–кампана и маастрихта (Брагина, Вишневская, 2007; Брагина, 2010, 2014; Bragina, Bragin, 2018; Soycan, Hakyemez, 2018 и другие). Время существования новых видов требует уточнения. Недостаточная изученность видового состава большинства известных кампан-маастрихтских комплексов, а также неясность стратиграфического распространения многих видов делает пока невозможным анализ их развития на видовом уровне. Поэтому приходится ограничиваться самыми общими данными на родовом уровне, привлекая виды лишь в тех случаях, когда речь идет о стратиграфически значимых из них (например, виды рода Amphipyndax).
В последнее десятилетие было проанализировано родовое разнообразие кампанских и маастрихтских радиолярий (O’Dogherty et al., 2009; Брагина, 2016б). Показатели изменений таксономического состава комплексов в кампане (число появляющихся, число вымирающих таксонов и их соотношения) невысокие. В течение кампана и маастрихта не появляются какие-либо новые таксоны радиолярий высокого ранга (семейство и выше), за исключением Miropylidae; более того, даже количество впервые появившихся новых родов сравнительно невелико, особенно в маастрихте. В целом развитие радиолярий в кампане и маастрихте наследует темпы, наблюдавшиеся в сантоне и более ранних веках (Брагина, 2016б), при этом происходит постепенное сокращение разнообразия на уровне родов.
Кампан характеризуется последним присутствием типично мезозойских представителей отряда Entactinaria: семейства Centrocubidae, Polyentactiniidae и Quinquecapsulariidae вымирают на рубеже кампана и маастрихта. В течение кампана появляются роды отряда Nassellaria: Bothryometra, Clathrocyclas, Homeoarchicorys, Immersothorax, Lithomelissa, Krempelinella, Myllocercion, Schaumellus, Foremanina и роды отряда Spumellaria: Amphibrachium, Amphymenium, Bipylomella, Cenellipsis, Miropyle, Pseudobrachium, Rhombastrum и Spongurus (O’Dogherty et al., 2009; Вишневская, 2011; Брагина, 2016б).
В это же время вымирают 10 родов из отряда Nassellaria: Annikaella, Cyprodictyomitra, Dictyodedalus, Dorypyle, Heliocryptocapsa, Homeoarchicorys, Krempelinella, Pseudoeucyrtis, Sandovalella, Saturniforma, Schaumellus, Spongocapsula и Thanarla и 7 родов отряда Spumellaria: Acastea, Acaeniotyle, Falsocromyodrimus, Paronaella, Parvicuspis, Quadrigastrum и Triactoma (O’Dogherty et al., 2009; Брагина, 2016б).
В кампане род Siphocampe, к которому в настоящее время относят многие виды, ранее считавшиеся принадлежащими к роду Theocampe, испытывает всплеск видообразования (Campbell, Clark, 1944; Empson-Morin, 1981). Увеличивается разнообразие рода Amphipyndax, остаются разнообразными представители рода Dictyomitra. В то же время наблюдается существенное изменение видового состава представителей семейства Pseudoaulophacidae (Alievium и Pseudoaulophacus) (Bragina, Bragin, 2018).
В маастрихте, по данным Л. О’Догерти с коллегами (O’Dogherty et al., 2009), в отряде Nassellaria появляется 6 родов (Acidnomelos, Botryocylinder, Calyptocoryphe, Clathrocyclas, Cornutovum, Eribotrys), а в отряде Spumellaria – 5 новых родов (Catenopyle, Heliosestarium, Pseudosaturnalis, Tholodiscus). Большая часть этих родов (Acidnomelos, Botryocylinder, Calyptocoryphe, Cornutovum, Eribotrys, Catenopyle, Pseudosaturnalis) вымирает к концу маастрихта. Кроме того, в маастрихте вымерли еще 19 родов (O’Dogherty et al., 2009) из отрядов Nassellaria (Botryometra, Cryptamphorella, Diacanthocapsa, Distylocapsa, Ectonocorys, Eostichomitra, Gongylothorax, Pseudotheocampe, Kuppelella, Microsciadiocapsa, Rhopalosyringium) и Spumellaria (Acanthocircus, Alievium, Miropyle, Neoparonaella, Patulibracchium, Prunobrachium, Orbiculiforma и Vitorfus). Количество вымирающих родов по сравнению с кампаном становится более значительным. В то же время трудно говорить о каком-либо массовом вымирании радиолярий на рубеже мела и палеогена, тем более что результаты детальных исследований радиолярий пограничных слоев маастрихта и палеоцена Новой Зеландии (Hollis, 1997) убедительно свидетельствуют о том, что многие меловые роды и виды радиолярий продолжали существовать в начале палеоцена. Следовательно, смена мезозойских комплексов радиолярий кайнозойскими являлась постепенной.
В развитии большинства родов радиолярий кампанского–маастрихтского времени наряду со снижением таксономического разнообразия наблюдается и морфологическое упрощение. Так, среди циртоидных насселлярий все реже встречаются морфотипы со сложной скульптурой внешней поверхности раковины (как у родов Pseudodictyomitra, Novixitus, Cyprodictyomitra). Упрощение внешнего строения до округлой формы в течение маастрихта наблюдается в филогенетической линии Pseudoaulophаcus stellatus–P. circularis (Bragina, Bragin, 2018). Аналогичная тенденция прослежена и для рода Alievium (Bragina, Bragin, 2018).
Итак, можно сказать, что в течение кампана и маастрихта развитие радиолярий сохранило черты, присущие ему с начала позднего мела (Брагина, 2016б). В целом кампан и маастрихт являются временем завершения позднемеловой стадии развития радиолярий, характеризующейся сравнительно медленными темпами эволюции и постепенным снижением биоразнообразия на уровне родов. Наблюдается высокая степень преемственности кампанских–маастрихтских комплексов радиолярий на уровне родов, в то время как на видовом уровне состав этих комплексов значительно обновляется. Все это сопровождается процессами морфологического упрощения в развитии ряда таксонов.
ОСНОВНЫЕ РУБЕЖИ И ЭТАПЫ РАЗВИТИЯ РАДИОЛЯРИЙ В КАМПАНЕ И МААСТРИХТЕ
Наблюдаемые тенденции сокращения таксономического и морфологического разнообразия группы, установленные, несмотря на пока еще недостаточную изученность видового состава комплексов радиолярий кампана и маастрихта, позволяют подойти к предварительному суммированию данных по этапности развития радиолярий в кампане и маастрихте. Такой анализ имеет большое значение для будущих зональных построений. Однако в настоящее время он может быть лишь весьма осторожным и не претендующим на завершенность. Ряд рубежей, которые могут представлять интерес при разработке биостратиграфии этого интервала по радиоляриям, являются сами по себе еще недостаточно изученными; положение многих из них пока не может быть установлено с полной достоверностью. Связано это, прежде всего, как с недостаточным числом уверенно датированных комплексов радиолярий, так и с отсутствием детально изученных разрезов, в которых была бы установлена последовательность радиоляриевых комплексов.
Первым этапом следует считать интервал нижнего кампана, входящий в состав радиоляриевой зоны Afens perapediensis (рис. 3). Он может приблизительно соответствовать нижней части зоны по планктонным фораминиферам Globotruncanita elevata (нижний кампан–низы верхнего кампана) (Robaszynski, Caron, 1995), однако верхняя граница зоны Afens perapediensis пока не установлена. В составе типичного тетического комплекса этого времени наблюдаются разнообразные представители родов Pseudoaulophacus и Alievium (Bragina, Bragin, 2018), постоянно встречаются Crucella espartoensis Pessagno, Afens perapediensis Bragina, Pseudoeucyrtis belogorskensis Bragina, а также Amphipyndax sp. А (Корчагин и др., 2012; Брагина и др., в печати). Последний таксон очень важен и нуждается в дополнительном изучении. Биостратиграфически он предшествует виду Amphipyndax pseudoconulus, но морфологически более схож с переходными формами от A. pseudoconulus к A. tylotus.
Рис. 3.
Важнейшие рубежи в развитии радиолярий в кампане и маастрихте. 1 – появление таксона; 2 – исчезновение таксона; 3 – этапы. Рубежи этапов показаны точечными линиями.
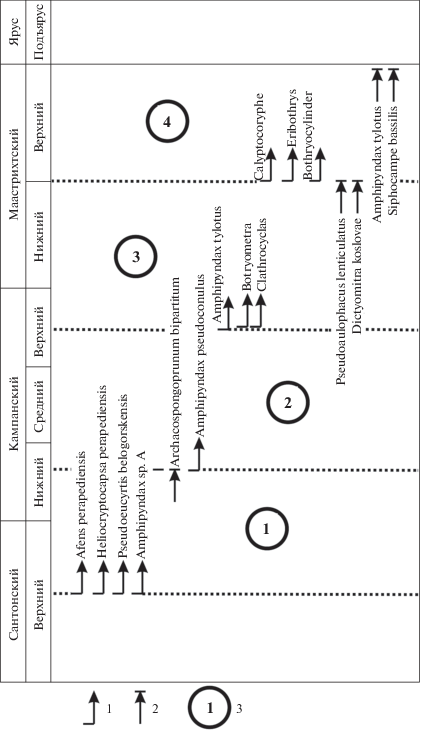
Появление Amphipyndax pseudoconulus можно считать важным рубежом первой половины кампана (рис. 3). Ранее считалось, что данный вид появляется на рубеже сантона и кампана (Sanfilippo, Riedel, 1985), однако убедительного подтверждения этого нет. Более того, в составе датированных комплексов радиолярий из низов нижнего кампана Amphipyndax pseudoconulus отсутствует (Брагина и др., 2016; Брагина и др., в печати). С появления A. pseudoconulus, приуроченного к нижнему кампану и, возможно, совпадающего или близкого по времени к исчезновению Archaeospongoprunum bipartitum Pessagno, начинается следующий этап в развитии тетических радиоляриевых комплексов кампана–маастрихта. Этот этап, вероятно, охватывает интервал от второй половины раннего кампана (или даже начала среднего кампана) до начала позднего кампана (рис. 3).
О составе комплексов этого интервала можно судить по материалам из Южной Турции (Moix et al., 2009). Здесь встречаются Amphipyndax pseudoconulus, Pseudoaulophacus floresensis, P. pargueraensis Pessagno, Clathropyrgus titthium, Dictyomitra koslovae, Botryometra heros, Dictyoprora salillum, Theocapsomma comys, Theocampe apicata и Foremanina schona. Нельзя исключить и присутствия Afens perapediensis. Внутри данного интервала могут быть рубежи появления и исчезновения некоторых таксонов, таких как Krempelinella, Myllocercion, Schaumellus, однако их точное положение пока неизвестно.
Следующим значительным рубежом в развитии радиолярий можно предположить появление Amphipyndax tylotus, приуроченное к позднему кампану (рис. 3). В это же время (не обязательно одновременно, но в пределах позднего кампана) появляются представители рода Clathrocyclas, широко распространенные в маастрихте Японии (Takahashi, Ishii, 1995; Hollis, Kimura, 2001) и Корякско-Камчатского региона (Вишневская, 2001). Рубеж кампана и мастрихта, возможно, не отмечается какими-либо значимыми событиями в развитии радиолярий. Косвенно на это указывает высокая степень сходства комплексов конца позднего кампана и начала раннего маастрихта, наблюдаемая на Кипре (Urquhart, Banner, 1994; Брагина, Брагин, 1995).
Сравнительно слабая изученность радиолярий маастрихта пока не позволяет с достоверностью наметить этапы развития радиолярий в это время. Можно лишь с осторожностью говорить о вероятных изменениях комплексов в средней части маастрихта, когда предположительно появляются первые Calyptocoryphe, несколько видов рода Eribothrys, а также представители Botryocylinder (Foreman, 1968; O’Dogherty et al., 2009). Вероятно, в это же время исчезают Pseudoaulophacus floresensis и Dictyomitra koslovae (Sanfilippo, Riedel, 1985). Таким образом, намечаются два этапа: конец позднего кампана–ранний маастрихт и поздний маастрихт (рис. 3).
Некоторые виды, широко распространенные в маастрихте, преодолевают рубеж маастрихта и дания, например Amphipyndax stocki, Myllocercion acineton, Theocampe vanderhoofi, Theocapsomma amphora, T. erdnussa и другие (Hollis, 1997). Степень изменений комплексов на этой границе не свидетельствует о массовом вымирании радиолярий. Тем не менее можно с уверенностью отметить позднемаастрихтский этап как завершающий в развитии мезозойской фауны радиолярий.
ЗАКЛЮЧЕНИЕ
1. Комплексы радиолярий кампана и маастрихта в низких палеоширотах характеризуются богатым и разнообразным составом. Однако степень их изученности в настоящее время недостаточна. Требуются значительные работы по таксономии с целью получения адекватного представления об их родовом и видовом составе, а также работы по биостратиграфии с целью уточнения распространения таксонов радиолярий путем сопоставления их с данными по другим группам фоссилий.
2. Развитие радиолярий в кампане и маастрихте наследует черты, присущие ему в докампанское время, и характеризуется относительно медленными темпами смены сообществ при постепенном снижении биоразнообразия на уровне родов. На рубеже маастрихта и дания происходит постепенная смена мезозойских комплексов кайнозойскими, часть мезозойских видов продолжает существовать и в палеоцене.
3. На основе данных по развитию радиолярий кампан-маастрихтский интервал может быть подразделен на несколько этапов: 1) позднесантонско-раннекампанский (во временнóм интервале зоны Afens perapediensis); 2) раннекампанско-позднекампанский; 3) позднекампанско-раннемаастрихтский; 4) позднемаастрихтский. На этой основе, при условии продолжения детальных исследований развития радиолярий, возможно построение новой зональной схемы.
4. Наиболее важными задачами будущих исследований могут быть: (1) уточнение первого появления видов Amphipyndax pseudoconulus и A. tylotus; (2) детальный анализ морфофилогенеза рода Amphipyndax; (3) уточнение рубежей появления маастрихтских родов Calyptocoryphe, Eribothrys, а также других представителей позднемеловых радиолярий; (4) установление таксономического состава датированных маастрихтских комплексов радиолярий и их палеонтологическое описание.
Благодарности. Автор выражает признательность М.С. Афанасьевой, В.С. Вишневской и В.А. Захарову за ценные советы и замечания.
Источники финансирования. Работа выполнена в соответствии с планами научно-исследовательской работы ГИН РАН (Брагина Л.Г. – тема № 0135-2018-0036) и при частичной финансовой поддержке гранта РФФИ (проект № 16-05-00363).
Список литературы
Беньямовский В.Н., Алексеев А.С., Овечкина М.Н., Вишневская В.С., Подгаецкий А.В., Пронин В.Г. Верхний кампан–нижний маастрихт северо-запада Ростовской области. Статья 1. Характеристика разрезов и палеонтологических комплексов, лито-биостратиграфия // Стратиграфия. Геол. корреляция. 2012. Т. 20. № 4. С. 33–67.
Брагина Л.Г. Новые виды меловых радиолярий рода Afens Riedel et Sanfilippo // Палеонтол. журн. 2010. № 1. С. 13–16.
Брагина Л.Г. Новые виды радиолярий отряда Nassellaria из коньяка–сантона формации Перапеди (Южный Кипр) // Палеонтол. журн. 2014. № 2. С. 3–10.
Брагина Л.Г. Зональная схема меловых (альб-сантонских) отложений тетических районов Евразии по радиоляриям // Стратиграфия. Геол. корреляция. 2016а. Т. 24. № 2. С. 41–66.
Брагина Л.Г. Развитие радиолярий в позднем альбе–кампане // Стратиграфия. Геол. корреляция. 2016б. Т. 24. № 5. С. 91–112.
Брагина Л.Г., Брагин Н.Ю. Радиолярии и стратиграфия кампан-маастрихтских отложений Юго-Западного Кипра // Стратиграфия. Геол. корреляция. 1995. Т. 3. № 2. С. 53–61.
Брагина Л.Г., Вишневская В.С. Новые виды меловых радиолярий рода Multastrum Vishnevskaya и их палеобиогеографическое распространение // Палеонтол. журн. 2007. Т. 41. № 6. С. 1–6.
Брагина Л.Г., Брагин Н.Ю., Джерич Н., Гаич В. Позднемеловые радиолярии и уточнение возраста субфлишевых отложений разреза Струганик, Западная Сербия // Стратиграфия. Геол. корреляция. 2014. Т. 22. № 2. С. 90–107.
Брагина Л.Г., Бенямовский В.Н., Копаевич Л.Ф. Радиолярии, фораминиферы и биостратиграфическое расчленение коньяка–кампана разреза Алан-Кыр, Горный Крым // Стратиграфия. Геол. корреляция. 2016. Т. 24. № 1. С. 44–63.
Брагина Л.Г., Брагин Н.Ю., Копаевич Л.Ф., Беньямовский В.Н. Сантонские радиолярии и фораминиферы в разрезе Брэждэ, Себрия // Вестник МГУ. Сер. 4. Геология. 2018. № 3. С. 12–24.
Брагина Л.Г., Брагин Н.Ю., Копаевич Л.Ф., Джерич Н., Герзина Н. Стратиграфия и микрофауна (радиолярии и фораминиферы) верхнемеловых (верхний сантон–нижний кампан) карбонатных отложений района села Струганик, Западная Сербия // Стратиграфия. Геол. корреляция (в печати).
Вишневская В.С. Радиоляриевая биостратиграфия юры и мела России. М.: ГЕОС, 2001. 376 с.
Вишневская В.С. Эволюция видового разнообразия меловых радиолярий высокоширотных палеобиохорий // Стратиграфия. Геол. корреляция. 2009. Т. 17. № 1. С. 110–121.
Вишневская В.С. Новые радиолярии семейства Prunobrachidae из верхов верхнего мела восточного склона Полярного Урала // Палеонтол. журн. 2011. № 4. С. 19–25.
Корчагин О.А. Планктонные фораминиферы и биостратиграфия верхнего кампана–нижнего маастрихта формации Мони, Южный Кипр // Стратиграфия. Геол. корреляция. 2011. Т. 19. № 5. С. 57–77.
Корчагин О.А., Брагина Л.Г., Брагин Н.Ю. Планктонные фораминиферы и радиолярии сантонских отложений горы Ак-Кая, Горный Крым, Украина // Стратиграфия. Геол. корреляция. 2012. Т. 20. № 1. С. 1–25.
Bragina L.G. Radiolarian biostratigraphy of the Perapedhi Formation (Cyprus): implications for the geological evolution of the Troodos Ophiolite // Bull. Soc. Geol. France. 2012. V. 183. № 4. P. 347–353.
Bragina L.G., Bragin N.Yu. Family Pseudoaulophacidae (Radiolaria) from the Upper Cretaceous (Coniacian–Maastrichtian) of Cyprus // Revue de micropaléontologie. 2018. V. 61. P. 55–79.
Campbell A.S., Clark B.L. Radiolaria from the Upper Cretaceous of Middle California // Spec. Pap. Geol. Soc. Am. 1944. № 57. P. 1–61.
Empson-Morin K.M. Campanian Radiolaria from DSDP Site 313, mid-Pacific Mountains // Micropaleontology. 1981. V. 27. № 3. P. 249–292.
Foreman H. Upper Maastrichtian Radiolaria of California // Spec. Pap. Paleontol. 1968. № 3. P. 1–82.
Foreman H. Cretaceous Radiolaria in the eastern South Atlantic, Deep Sea Drilling Project, Leg 40 // Initial Reports of the Deep Sea Drilling Project. Eds. Bolli H.M., Ryan W.B.F. Washington, DC: U.S. Government Printing Office, 1978. V. 40. P. 839–843.
Hashimoto H., Ishida K. Correlation of selected radiolarian assemblages of the Upper Cretaceous Izumi and Sotoizumi Groups and Shimanto Supergroup, in Shikoku // News of Osaka Micropaleontologists. 1997. Spec. V. 10. P. 245–257.
Hollis C.J. Cretaceous–Paleocene Radiolaria from Eastern Marlborough, New Zealand // Institute of Geological & Nuclear Sciences. Monograph. 1997. № 17. P. 1–152.
Hollis C.J., Kimura K. A unified radiolarian zonation for the Late Cretaceous and Paleocene of Japan // Micropaleontology. 2001. V. 47. № 3. P. 235–255.
Iwata K., Tajika J. Late Cretaceous radiolarians of the Yubetsu Group, Tokoro Belt, northeast Hokkaido // J. Fac. Sci. Hokkaido Univ. 1986. Ser. IV. V. 21. № 4. P. 619–644.
Moix P., Goričan Š., Marcoux J. First evidence of Campanian radiolarians in Turkey and implications for the tectonic setting of the Upper Antalya Nappes // Cretaceous Res. 2009. V. 30. P. 952–960.
Nakaseko K., Nishimura A. Upper Jurassic and Cretaceous Radiolaria from the Shimanto Group in Southwest Japan // Sci. Rept. College Gen. Educ. Osaka Univ. 1981. V. 30. № 2. P. 133–203.
O’Dogherty L., Carter E.S., Dumitrica P. et al. Catalogue of Mesozoic radiolarian genera. Pt. 2: Jurassic–Cretaceous // Geodiversitas. 2009. V. 31. № 2. P. 271–356.
Okamura M. Radiolarian fossils from the Northern Shimanto Belt (Cretaceous) in Kochi Prefecture, Shikoku // Geology and paleontology of the Shimanto Belt. Eds. Taira A., Tashiro M. Rinyakyosaikai, Kochi, 1980. P. 153–178.
Pessagno Jr., E.A. Upper Cretaceous Radiolaria from Puerto Rico // Micropaleontology. 1963. V. 9. № 2. P. 197–214.
Pessagno Jr., E.A. Jurassic and Cretaceous Hagiastridae from the Blake-Bahama Basin (Site 5A, JOIDES Leg I) and the Great Valley sequence, California Coast Ranges // Bull. Am. Paleontol. 1971. V. 60. № 264. P. 5–83.
Pessagno Jr., E.A. Radiolarian zonation and stratigraphy of Upper Cretaceous portion of Great Valley Sequence // Micropaleontology. 1976. Spec. Publ. № 2. P. 1–96.
Petrushevskaya M.G., Kozlova G.E. Radiolaria: Leg 14 // Initial Reports of the Deep Sea Drilling Project. Eds. Bolli H.M., Ryan W.B.F. Washington, DC: U.S. Government Printing Office, 1972. V. 14. P. 495–532.
Riedel W.R., Sanfilippo A. Radiolaria from the Southern Indian Ocean, DSDP Leg 26 // Initial Reports of the Deep Sea Drilling Project. Washington: U.S. Government Printing Office, 1974. V. 26. P. 771–813.
Robaszynski F., Caron M. Foraminifѐres planctoniques du Crétacé: commentaire de la zonation Europe-Mediterranée // Bull. Soc. géol. France. 1995. V. 166. № 6. P. 681–692.
Sanfilippo A., Riedel W.R. Cretaceous Radiolaria // Plankton stratigraphy. Eds. Bolli H.M., Saunders J.B., Perch-Nielsen K. Cambridge: Cambridge University Press, 1985. P. 573–630.
Soycan H., Hakyemez A. The first calibration of radiolarian biochronology with Late Cretaceous (latest Coniacian–Santonian to early Campanian) planktonic foraminifera in the volcano-sedimentary sequences of the Eastern Pontides, NE Turkey // Cretaceous Res. 2018. V. 85. P. 319–348.
Takahashi O., Ishii A. Radiolarian assemblage-zones in the Jurassic and Cretaceous sequence in the Kanto Mountains, central Japan // Memoirs of the Faculty of Science, Kyushu University. Series D. Earth and Planetary Sciences. 1995. V. 24. P. 49–85.
Thurow J. Cretaceous radiolarians of the North Atlantic Ocean; ODP Leg 103 (sites 638, 640, and 641) and DSDP legs 93 (Site 603) and 47B (Site 398) // Eds. Boilot G., Winterer E.L. Proc. Ocean Drilling Program, Scientific Results. 1988. V. 103. P. 379–418.
Urquhart E., Banner F.T. Biostratigraphy of the supraophiolite sediments of the Troodos Massif, Cyprus: the Cretaceous Perapedhi, Kannaviou, Moni and Kathikas formations // Geol. Mag. 1994. V. 131. № 4. P. 499–518.
Yamasaki T. Radiolarian assemblages of the Izumi Group in Shikoku and western Awaji Island, southwest Japan // J. Geol. Soc. Japan. 1987. V. 93. P. 403–417.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция