

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 4, стр. 96-110
Келловейские остракоды Центрального Дагестана: биостратиграфия, палеоэкология и хорологический анализ
Е. М. Тесакова 1, 2, *, Л. А. Глинских 3, 4
1 Московский государственный университет им. М.В. Ломоносова
Москва, Россия
2 Геологический институт РАН
Москва, Россия
3 Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
Новосибирск, Россия
4 Новосибирский государственный университет
Новосибирск, Россия
* E-mail: ostracon@rambler.ru
Поступила в редакцию 23.08.2019
После доработки 17.11.2019
Принята к публикации 20.11.2019
Аннотация
Анализ распределения келловейских остракод армхинской свиты по разрезу Цудахар (Центральный Дагестан) позволил установить последовательность из 6 комплексов, обусловленных палеоэкологией, и впервые провести биостратиграфическое расчленение келловея Дагестана по ракушковым ракам. В среднем келловее выделены слои с Pleurocythere khapissovi и установлена зона Neurocythere cruciata intermedia европейских остракодовых шкал, что облегчает межрегиональную корреляцию по остракодам. Палеоэкологический анализ показал формирование нижней части разреза (слои с P. khapissovi и зона N. cruciata intermedia) в прибрежной зоне верхней сублиторали с нормально-морской соленостью, хорошей аэрацией и низкими темпами осадконакопления. Верхняя часть разреза (палеоэкологические интервалы с Schuleridea translucida–Cytherella perennis и с C. perennis) накапливалась в нижней зоне верхней сублиторали с высокой скоростью седиментации, высокой (и постоянно растущей) эвтрофией и пониженными значениями pH. Анализ палеогеографического распространения изученных видов выявил связи Кавказского моря с Западной и Восточной Европой, а также со Средней Азией.
ВВЕДЕНИЕ
С точки зрения стратиграфии любая группа микрофауны имеет свои преимущества и ограничения, обусловленные биологией и характером скелета. Например, юрские морские остракоды не столь широко распространены в различных спектрах осадочных фаций, как фораминиферы, и далеко не так обильны и разнообразны. Это надолго задержало применение ракушковых раков в биостратиграфии (особенно в СССР), хотя их стратиграфический потенциал зачастую выше, чем у простейших (Тесакова, 2010). Так, немецкая юра обычно датируется именно по этой группе, поскольку фораминиферы не обеспечивают такую же детальность (например, Schudack, 1994; Schudack, Schudack, 2000; Franz et al., 2009). Остракоды также дали бóльшую детальность при расчленении нижнемелового разреза о. Колгуев (Баренцево море) (Куприянова, 2000) и верхнеюрского разреза Михаленино (Костромская обл.) (Tesakova et al., 2012). Поскольку они реагировали на палеособытия раньше, нижняя граница стратонов по ракушковым фиксировалась ниже границ фораминиферовых стратонов, и только в случае перерывов (явных или скрытых) границы комплексов обеих фаун совпадали.
За рубежом юрские остракоды используются в биостратиграфии с 1930-х годов, а в построении зональных шкал с 1960-х годов. Такие шкалы разработаны для морской юры Северной Франции (Bodergat, 1997), Шотландии и Англии (Bate, 2009; Wilkinson, Whatley, 2009), Северной Германии (Triebel, 1951; Wienholz, 1967; Brand, 1990; Schudack, 1994) и Польши (Bielecka et al., 1988a, 1988b). Остракодовые шкалы Сибири (Никитенко, 2009) и Русской плиты (Колпенская, 1999; Тесакова, 2014, 2015; Тесакова и др., 2017) созданы значительно позже. Стратиграфический и корреляционный потенциал таких шкал достаточно высок в каждой из перечисленных стран, но их прямое сопоставление крайне затруднено из-за различной изученности остракодовой фауны и разницы в принципах, положенных в основу выделения зон. В последнее время стала возможной частичная корреляция между упомянутыми шкалами Западной и Восточной Европы (в интервале средней юры) с помощью миграционных панъевропейских уровней, установленных по видам-космополитам, распространявшимся с трансгрессиями (Тесакова, 2014, 2015; Тесакова и др., 2017). С той же целью разрабатываются филогенетические последовательности по эволюции родов, широко расселившихся в Европе: Lophocythere Sylv.-Bradley, 1948, Camptocythere Triebel, 1950 и Palaeocytheridea Mandelstam, 1947 (Tesakova, 2013b, 2014; Тесакова, 2014; Shurupova, Tesakova, 2017a, 2017b, 2019).
Шкалы для Южной Германии и Швейцарии (Schudack, Schudack, 2000; Schudack et al., 2013; Franz et al., 2009; Beher et al., 2010; Dietze et al., 2017) отличаются от вышеперечисленных довольно сильно. Например, площадное распространение некоторых зональных таксонов ограничено только регионом Швабского Альба, а первое появление панъевропейских зональных индексов в разрезе южной части Западной Европы происходит раньше, чем в ее северной части. Поэтому корреляция по остракодам между северной и южной частями Западной Европы до сих пор не проводилась даже на отдельных уровнях.
Одним из обнажений, перспективным для подобного сопоставления, является опорный для келловея Центрального Дагестана разрез Цудахар (рис. 1) (Гаврилов и др., 2015), в котором, наряду с различными беспозвоночными, в том числе аммонитами, обнаружены ракушковые раки (рис. 2). Юрские остракоды этого региона до последнего времени практически не изучались. В предыдущей статье авторов (Глинских, Тесакова, 2020) выявлен их систематический состав (с опубликованием изображений) и дано полное описание нового вида. Настоящая работа посвящена биостратиграфии и палеоэкологии этого же разреза, выполненным по остракодам. Также в задачи настоящей статьи входит изучение корреляционного потенциала цудахарских остракод и их палеогеографический (хорологический) анализ.

Рис. 2.
Распределение остракод по разрезу Цудахар. Литологическая колонка и расчленение по аммонитам приведены по (Гаврилов и др., 2015, с изменениями). Условные обозначения: 1 – глина, 2 – глина алевритовая, 3 – алевролит, 4 – известняк, 5 – мергель. Цифры у залитых кружков означают число экземпляров вида на данном уровне разреза. а–з – фотоизображения остракод: а – Cytherella perennis Błaszyk, 1967 s.l., экз. № DZ-19/1, цр (целая раковина) самки слева, сл. 17, зона Coronatum; б – Praeschuleridea wartae Błaszyk, 1967 s.l., экз. № DZ-19/4, цр самца справа, сл. 7, зона Calloviense; в – Schuleridea translucida (Lyubimova, 1955) s.l., экз. № DZ-19/7, цр самца справа, сл. 7, зона Calloviense; г – Pleurocythere khapissovi Glinskikh et Tesakova, 2020, голотип № DZ-19/13, цр самки слева, сл. 2, зона Calloviense; д – Neurocythere rimosa (Dépêche, 1973), экз. № DZ-19/9, цр справа, сл. 2, зона Calloviense; е – Palaeocytheridea (P.) sp., экз. № DZ-19/12, цр слева, сл. 9, зона Jason; ж – Neurocythere cruciata intermedia (Lutze, 1960), экз. № DZ-19/11, цр самки слева, сл. 13, зона Coronatum; з – Bairdia pumicosa Sheppard in Franz et al., 2009, экз. № DZ-19/6, цр самки справа, сл. 9, зона Jason. Сокращения: В. цудах – Верхнецудахарская, Call. – Calloviense, Enod. – Enodatum, ?Med.–Jas. – ?Medea-Jason.

Разрез Цудахар представлен армхинской свитой (верхи нижнего–низы верхнего келловея, аммонитовые зоны Calloviense, Jason, Coronatum и Athleta), которая с несогласием подстилается цудахарской свитой (верхний байос–бат) и перекрывается иронской свитой (верхняя юра). Подробное описание литологии и комплексов ископаемых (аммониты, белемниты, двустворки), биостратиграфия, а также изображение характерных фораминифер и седиментологическая интерпретация приведены в работе (Гаврилов и др., 2015, с. 81–105).
Детальная характеристика материала, легшего в основу настоящей работы, опубликована в статье (Глинских, Тесакова, 2020).
Коллекция (№ DZ-19) хранится в лаборатории микропалеонтологии Института нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН.
БИОСТРАТИГРАФИЯ И КОРРЕЛЯЦИЯ
Крайне неравномерное распределение остракод по разрезу Цудахар позволило выделить шесть комплексов (рис. 2), которые характеризуют разные стратиграфические интервалы, установленные в разрезе по аммонитам М.А. Роговым и Д.Б. Гуляевым (Гаврилов и др., 2015). Описание комплексов производится снизу вверх.
Комплекс I выделяется в нижнем келловее (зона Calloviense, подзона Enodatum) и охватывает слои 1–7. Его систематический состав (Cytherella perennis Błaszyk, 1967 s.l., Praeschuleridea wartae Błaszyk, 1967 s.l., Schuleridea translucida (Lyubimova, 1955) s.l., Pleurocythere khapissovi Glinskikh et Tesakova, 2020 и Neurocythere rimosa (Dépêche, 1973)) почти полностью переходит в вышележащий комплекс, кроме последнего вида, единственный экземпляр которого встречен только в подошве комплекса I. Скорее всего, он переотложен из бата, на котором нижний келловей залегает с большим перерывом. В пользу аллохтонности N. rimosa свидетельствует приуроченность этого таксона именно к батскому интервалу (весь бат Южной Англии, нижний–средний бат Северо-Западной Франции и Южной Германии; табл. 1).
Таблица 1.
Палеогеографическое распространение цудахарских остракод (см. рис. 5)
| Вид | Распространение |
|---|---|
| B. pumicosa Sheppard in Franz et al., 2009 | Бат и келловей Ю Германии (Buck, 1954; Schudack, Schudack, 2000, p. 102, fig. 6 ; Franz et al., 2009, p. 132, pl. 1, fig. 8 , pl. 6, fig. 12 ; Beher et al., 2010, p. 50, text-fig. 3; Franz et al., 2014, fig. 15r ); бат Нормандии (Sheppard, 1981a, pl. 2, fig. 8 ; 1981b, p. 44, pl. 4, figs. 1–7 ; Dépêche, 1984, pl. 3, figs. 3, 4; 1985, pl. 29, fig. 1), нижний бат Дорсета (Sheppard, 1981a, pl. 2, fig. 8 ; 1981b, p. 44, pl. 4, figs. 1–7 ); нижний бат и нижний келловей Польши (Tesakova et al., 2008, pl. 1, fig. 3) |
| N. cruciata intermedia (Lutze, 1960) | Нижний келловей (начиная с зоны Koenigi)–нижний оксфорд Ю Германии (Buck, 1954; Huber et al., 1987, fig. 3; Franz et al., 2009, p. 147, pl. 5, fig. 8 ; Beher et al., 2010, p. 51, pl. 2, fig. 9 ; Franz et al., 2014, fig. 15p ; Dietze et al., 2017, pl. 12, fig. k); средний келловей восточной части Нидерландов и Днепровско-Донецкой впадины (Пяткова, Пермякова, 1978, с. 146, табл. 62 , фиг. 3–4; Herngreen et al., 1983, pl. 3, figs. 1–5); средний и верхний келловей СВ Германии (Lutze, 1960, p. 423, pl. 34, figs. 5–6 ); средний–верхний келловей и нижний оксфорд (зоны E. coronatum–C. cordatum) Шотландии и Англии (Whatley, 1970, p. 338, pl. 10, figs. 6, 7, 9–21 , pl. 11, figs. 1, 4; Kilenyi, 1978, pl. 11, figs. 9–12 ; Whatley et al., 2001, p. 153, pl. 5, figs. 4–9 ; Whatley, Ballent, 2004, p. 100; Wilkinson, Whatley, 2009, pl. 8, figs. 8–11 ); верхний келловей Парижского бассейна и Польши (Oertli, 1963, pl. 34, fig. 1f, pl. 35, figs. 1f, 2f; Bielecka et al., 1988a, p. 171, pl. 67, fig. 1; Olempska, Błaszyk, 2001, p. 568, figs. 9A–H ) |
| N. rimosa (Dépêche, 1973) | Нижний–средний бат Нормандии и весь бат Ю Англии (Dépêche, 1973, p. 218, pl. 1, figs. 9–14 ; 1984, p. 241, pl. 10, figs. 1–3; 1985, pl. 30, fig. 2; Sheppard, 1981b, p. 83, pl. 13, figs. 2–5; Bate, 2009, pl. 2, figs. 7–9 ); нижний–средний бат (зона Zigzag, подзона Macrescens–зона Morrisi) Ю Германии (Franz et al., 2014, fig. 13p ) |
| C. perennis Błaszyk, 1967 s.l. | Байос и бат Польши (Błaszyk, 1967, p. 16, pl. 2, figs. 1–11 , pl. 3, figs. 6, 7 ; Bielecka et al., 1988a, p. 169, pl. 65, fig. 3; Tesakova et al., 2008, pl. 1, fig. 1); верхний байос Днепровско-Донецкой впадины (Пяткова, Пермякова, 1978, с. 122, табл. 45 , фиг. 1); нижний–средний келловей Парижского бассейна (Лотарингия, Франция) (Dépêche, 1969, p. 264, pl. 3, figs. 7, 8 ); средний келловей–средний оксфорд Европейской России (Герасимов и др., 1996, табл. 3 , фиг. 2–3; Tesakova, 2003, p. 157, pl. 1, figs. 7–9 ; Tesakova, 2008, fig. 2 (2); Тесакова и др., 2017, табл. 2 , фиг. 2) |
| P. wartae Błaszyk, 1967 s.l. | Бат Польши (Błaszyk, 1967, p. 43, pl. 12, figs. 1–2; Bielecka et al., 1988a, p. 176, pl. 72, fig. 3); нижний келловей Белоруссии (Махнач, Тесакова, 2015, табл. 3 , фиг. 14–18 ); нижний–средний келловей (зоны Gowerianus–Jason) Европейской России (Tesakova, 2003, pl. 10, figs. 1–4; Тесакова и др., 2009, табл. 2 , фиг. 1–4; Tesakova, 2013a, pl. 6, fig. 17 ) |
| S. translucida (Lyubimova, 1955) s.l. | Нижний келловей Днепровско-Донецкой впадины (Пяткова, Пермякова, 1978, с. 138, табл. 53 , фиг. 10–11 ); средний–верхний келловей Европейской России (Любимова, 1955, с. 97, табл. 11 , фиг. 1; 1956, с. 564, табл. 3 , фиг. 7–8 ; Герасимов и др., 1996, табл. 3 , фиг. 5; Tesakova, 2003, p. 186, pl. 9, fig. 15 (non figs. 12–14, 16 ); Tesakova, 2008, fig. 3 (12); Тесакова и др., 2017, табл. 4 , фиг. 16–17 ) |
Стратиграфическое распространение других остракод (табл. 1) не противоречит раннекелловейскому возрасту комплекса I. Виды C. perennis, P. wartae и S. translucida являются фоновыми и самыми обильными в разрезе (рис. 3а, 3б) и в ассоциации с P. khapissovi облигатно входят в состав комплексов I, II и III.
Рис. 3.
Соотношение доминирующих видов (а) в экземплярах (общая численность = обилие) и (б) в процентах. Условные обозначения: 1 – видовое разнообразие, 2 – C. perennis, 3 – P. wartae, 4 – S. translucida, 5 – все остальные таксоны (B. pumicosa, N. cruciata intermedia, N. rimosa, P. khapissovi, Palaeocytheridea (P.) sp. и Cytheropteron sp.). Сокращения: Med.–Jas. – Medea-Jason, Phae. – Phaeinum, Pron. + Spin. – Proniae + Spinosum.
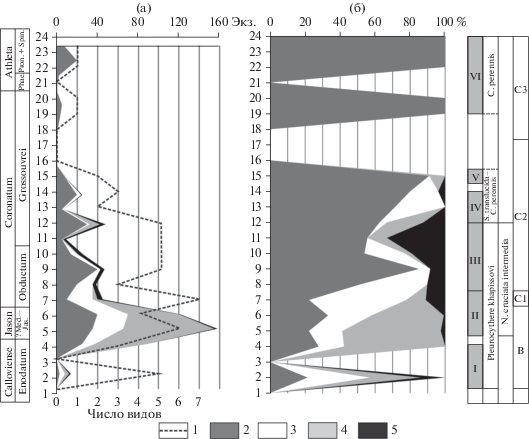
Комплекс II определен в среднем келловее (зоны Jason и Coronatum, с подзонами ?Medea–Jason и нижней частью подзоны Obductum) в слоях 9–12. Отличается от предыдущего комплекса тремя новыми таксонами: Bairdia pumicosa Sheppard in Franz et al., 2009, Neurocythere cruciata intermedia (Lutze, 1960) и Palaeocytheridea (P.) sp., по первому появлению которых проводится нижняя граница комплекса II. Появление в разрезе зонального вида N. cruciata intermedia (см. ниже) маркирует начало среднего келловея.
Комплекс III выделяется в слоях 13–26 среднего келловея (зона Coronatum, верхняя часть подзоны Obductum и нижняя часть подзоны Grossouvrei). Его состав почти идентичен составу предыдущего комплекса, за исключением того, что B. pumicosa исчезает из разреза вблизи нижней границы комплекса, и в комплексе встречен единственный экземпляр Cytheropteron sp., что важно с точки зрения палеоэкологии. Главным же отличием этого комплекса является другая палеоэкологическая особенность – смена доминантов с шулеридеид на цитереллиды – начало проявления Cytherella-сигнала (см. ниже).
Комплекс IV установлен в слоях 28–30 (зона Coronatum, средняя часть подзоны Grossouvrei). Он демонстрирует двукратное обеднение систематического состава, причем C. perennis и P. wartae относительно обильны, а P. (P.) sp. встречен в единственном экземпляре в верхней части комплекса.
Комплекс V выделяется в том же стратиграфическом интервале, в слое 36, и включает два вида – малочисленный C. perennis и единичные находки S. translucida.
Комплекс VI моновидовой, состоит из C. perennis, охватывает широкий интервал разреза – слои 48–58 – и датируется верхами зоны Coronatum (верхняя часть подзоны Grossouvrei) среднего келловея и зоной Athleta верхнего келловея (подзона Phaeinum и нижняя часть Proniae + Spinosum).
Распределение остракод по разрезу (рис. 2), с учетом вариаций их разнообразия, общей численности и смены доминантов (рис. 3а, 3б), четко разделяет разрез Цудахар на две части: нижнюю (комплексы I–III) и верхнюю (комплексы IV–VI). Нижняя часть полностью соответствует слоям с Pleurocythere khapissovi, которые определяются по распространению вида-индекса и выделяются в настоящей работе впервые. В будущем вид-индекс может приобрести зональный статус, поскольку является звеном неизвестной нам пока ветви филогенетического древа плевроцитер. Стратиграфический объем слоев – нижний–средний келловей (зоны Calloviense–Coronatum), а их географическое распространение пока ограничивается Центральным Дагестаном.
Кроме того, в нижней части разреза (в объеме комплексов II и III) установлена о-зона N. cruciata intermedia. Зональный или характерный для аналога этой зоны вид (рис. 4) почти всюду в Западной Европе появляется со среднего келловея и только в Южной Германии фиксируется раньше, с середины нижнего келловея (Franz et al., 2009). Одноименная о-зона в Южной Германии охватывает интервал аммонитовых зон Koenigi–Lamberti (Dietze et al., 2017).
Рис. 4.
Келловейские зональные шкалы по остракодам Западной и Восточной Европы. Интервал распространения N. cruciata intermedia выделен серым цветом.

В бореальной остракодовой стратиграфии N. cruciata intermedia используется как характерный сопутствующий вид английской зоны Lophocythere interrupta interrupta (рис. 4), отвечающей аммонитовым зонам Coronatum–Lamberti (Kilenyi, 1978; Wilkinson, Whatley, 2009). Та же зона L. interrupta interrupta, практически с тем же объемом (Jason–Lamberti), установлена в Парижском бассейне (Bodergat, 1997), но, описывая ее, Бодерга не упомянула о N. cruciata intermedia. Однако ранее этот вид отмечался в верхнем келловее Парижского бассейна как стратиграфически значимый (Dépêche, 1985, tabl. 6) и помещался в предполагаемую эволюционную филолинию N. cruciata cruciata (Triebel, 1951)–N. cruciata intermedia–N. cruciata oxfordiana (Lutze, 1960)–N. cruciata kimmeridgiensis (Guyader, 1967) (Dépêche, 1985, p. 121). Появление во Франции N. cruciata intermedia только с верхнего келловея, по-видимому, связано с недостаточной изученностью или с ухудшением условий существования остракод в среднем келловее, так как в этом интервале в Парижском бассейне отмечена всего пара стратиграфически значимых видов (Dépêche, 1985, tabl. 6). Анализ стратиграфического распространения остракод в польской юре (Bielecka et al., 1988a; Olempska, Błaszyk, 2001; Tesakova et al., 2008) позволил Тесаковой (2014, 2015) предложить зональное расчленение юрских отложений Польши по ракушковым. Начиная с зоны Jason появляются три характерных вида: N. cruciata intermedia, Lophocythere karpinskyi (Mand. in Lyub., 1955) и N. caesa (Triebel, 1951). Зональным, отвечающим интервалу Jason–нижняя половина Pollux [= Coronatum], был выбран второй вид, однако появление N. cruciata intermedia тоже может считаться индексом нижней границы среднего келловея в Польше. В зональных схемах Северной Германии (Triebel, 1951; Wienholz, 1967; Brand, 1990; Schudack, 1994) вид N. cruciata intermedia не фигурирует, но он впервые описан именно в Саксонии и появляется в немецком разрезе юры именно со среднего келловея, с зоны Castor et Pollux [верхняя часть Jason–нижняя часть Coronatum] (Lutze, 1960). В келловее Днепровско-Донецкой впадины этот вид также фиксируется со среднего подъяруса (Пяткова, Пермякова, 1978), но на территории Русской плиты встречен не был (Тесакова, 2014).
Таким образом, в южной части Западной Европы о-зона N. cruciata intermedia отвечает а-зонам Koenigi–Lamberti, а в северной части – интервалу зон Jason–Lamberti.
В разрезе Цудахар интервал зоны N. cruciata intermedia строго датируется по аммонитам как нижняя часть зоны Jason (индекс не встречен в зоне Calloviense)–средняя часть зоны Coronatum.
Ранее по остракодам подрода Palaeocytheridea (Palaeocytheridea) Mandelstam, 1947 были выделены четыре филозоны в средней юре Европы (Тесакова, 2014; Tesakova, 2014). Один из зональных видов – P. (P.) carinilia (Sylvester-Bradley, 1948) – известен из нижнего бата–нижнего келловея Северной Франции и Англии (Sylvester-Bradley, 1948; Dépêche, 1969, 1973, 1984; Sheppard, 1981b) и среднего бата Южной Германии (Malz, 1975; Liebau, 1987). Другой вид – P. (P.) pavlovi (Lyubimova, 1955) – встречается в нижнем келловее Поволжья (Любимова, 1955; Tesakova, 2013b, 2014; Тесакова, 2014). К сожалению, сохранность дагестанского материала не позволила определить до вида экземпляры P. (P.) sp. из комплексов II–IV и сопоставить этот интервал с западноевропейскими шкалами по палеоцитеридеям, в том числе с остракодовой шкалой Русской плиты.
Верхняя часть разреза Цудахар отличается бедностью систематического состава остракод. Комплексы IV и V объединяются в палеоэкологический интервал с S. translucida–C. perennis, который малоинформативен с точки зрения стратиграфии, так как виды-индексы имеют неоправданно широкий вертикальный диапазон (табл. 1), что, по-видимому, связано со сборными таксонами (Глинских, Тесакова, 2020). По аммонитам этот интервал сопоставляется с зоной Coronatum, подзоной Grossouvrei среднего келловея. Он является переходным к моновидовому комплексу VI, которому отвечает палеоэкологический интервал с Cytherella perennis. Последний также датируется по аммонитам верхами подзоны Grossouvrei зоны Coronatum среднего келловея–подзонами Phaeinum и Proniae+Spinosum зоны Athleta верхнего келловея.
Стратиграфическая несостоятельность интервалов с S. translucida–C. perennis и с C. perennis компенсируется их высоким палеоэкологическим значением (см. ниже).
ПАЛЕОЭКОЛОГИЯ
Анализ циклического строения келловейской толщи позволил Ю.О. Гаврилову (Гаврилов и др., 2015, с. 91–94) реконструировать в Цудахаре три крупных седиментационных цикла – один полный и два редуцированных (рис. 2). Редуцированные циклы B и D отвечают базальной (крайне мелководной) фазе трансгрессии при несохранившейся более высокой части, денудированной во время падения уровня моря перед последующими трансгрессиями. Оба цикла представлены известковистыми песчано-глинистыми алевролитами с несколькими пластами песчано-известковистых пород. Полный цикл C представлен всеми своими элементами. Начальной фазе цикла (C1) отвечает медленная трансгрессия с попеременным накоплением в мелководной обстановке глинистых и карбонатных пород. Чередование этих слоев (с преобладанием карбонатов) относят за счет частых флуктуаций уровня моря, связанных с прецессионными циклами Миланковича. Формирование карбонатных слоев (“мусорные известняки”) происходило в регрессивные эпизоды. Вторая фаза (C2), во время которой преимущественно накапливались глинистые породы, а число и мощность карбонатных прослоев сокращались, связывается с быстрой трансгрессией и редкими кратковременными эпизодами обмеления. Этот процесс сопровождался высокой скоростью осадконакопления, что препятствовало сильному углублению бассейна, и обильным поступлением органического вещества (ОВ), повышающего эвтрофию. Окисление ОВ приводило к снижению pH, а также могло спровоцировать углекислое заражение придонной воды (Гаврилов, 2005). Финальная стадия цикла (C3) ознаменовалась замедлением или остановкой трансгрессии, что привело к прогрессирующему обмелению бассейна и формированию карбонатной пачки.
Сравним палеоэкологические реконструкции, полученные методами литологического и остракодового анализа. Следует учитывать, что комплексы ракушковых были изучены только из алевролитовых прослоев, поскольку выделение их остатков из крепких известняков крайне затруднительно. Кроме того, заведомо мелководное происхождение “мусорных известняков” в обстановке высокой гидродинамики (Гаврилов и др., 2015, с. 89–91) не способствовало захоронению скелетов остракод (Захаров и др., 1988, с. 424; Tesakova, Shurupova, 2018, p. 1565).
Циклические флуктуации уровня моря закономерно отражаются в распределении бентоса на шельфе, в том числе остракод (Pokorný, 1971; Babinot, Lethiers, 1984; Андреев, 1988; Kolasa et al., 1998; Boomer, Eisenhauer, 2002; Tesakova, 2008, 2013a; Tesakova et al., 2016; Tesakova, Shurupova, 2018). Для прибрежно-мелководной обстановки (низкий уровень моря и близкий берег) характерно низкое таксономическое разнообразие, а общая численность, в зависимости от трофности, может быть скудная или высокая (последнее только за счет доминантов). Умеренно-глубоководная, удаленная от берега зона шельфа (высокий уровень моря и удаленный берег) обычно характеризуется высокими таксономическим разнообразием и общей численностью, но без выраженных доминантов (исключением может стать дизоксидная обстановка).
Соответственно, если в каком-то интервале разреза разнообразие и численность сильно варьируют, скорее всего, можно предположить обстановку верхней сублиторали и оценить изменение относительной глубины в ее пределах. Если же разнообразие и численность стабильны и высоки, можно предполагать обстановку нижней сублиторали.
Относительную глубину и ее вариации можно реконструировать методом размерных кластеров остракод. Верхняя сублитораль прекрасно устанавливается по высокому содержанию в комплексах крупноразмерных остракод (≈80–100%), к которым в юрское время относились формы с длиной раковины ≈0.48–1.2 мм. Мелкоразмерные остракоды (в юре ≈0.25–0.32 мм), напротив, существенно преобладали на нижней сублиторали. Мелкие вариации соотношения размерных кластеров позволяют судить об увеличении/уменьшении глубины в пределах каждой из этих биономических зон (Tesakova, Schurupova, 2018).
Другая методика разработана для оценки относительной глубины на нижней сублиторали (начиная с глубин 50–60 м). На шельфе с ростом глубины и уменьшением освещенности дна возрастают не только размер и выпуклость глазных бугорков, но и число таксонов с глазными бугорками и их общая численность (Карпук, 2016).
Можно также использовать и иные методы: анализ распространения по разрезу видов-космополитов, морфофункциональный анализ раковин остракод и анализ распространения по разрезу видов-индексов различных палеоэкологических обстановок и соотношения их доминирования (Шорников, Михайлова, 1990; Tesakova, Schurupova, 2018 и др.).
Следует иметь в виду, что описанные закономерности и их графическое отображение нарушаются при отклонении от нормальных условий обитания или захоронения остракод (высокая эвтрофия, сероводородное или углекислое заражение и др.).
В результате комплексного анализа, проведенного на основе некоторых из перечисленных методов, сделаны следующие реконструкции. Слои с P. khapissovi (рис. 2, 3) характеризуются невысоким таксономическим разнообразием морских остракод (3–7 видов) при периодическом возрастании общей численности за счет 2–3 доминантов. Обилие варьирует в широких пределах от 12 до 162 экз. на образец. Перечисленные характеристики присущи обстановкам верхней сублиторали, подверженной быстрым и частым изменениям факторов среды более, чем другие биономические зоны моря.
При этом в нижней части соответствующего графика (комплексы I и II) низкое разнообразие в сочетании с сильно варьирующей численностью выражено особенно ярко (рис. 3а), что говорит о периодическом обмелении верхней сублиторали, вплоть до прибрежья, и полностью согласуется с седиментологическими выводами об обстановках накопления циклов B и C1. Периодически существовавшая прибрежная обстановка в интервале комплексов I и II и циклов B и C1 подтверждается анализом таксономической структуры сообществ изученных остракод, а именно отчетливым доминированием двух таксонов-убиквистов – P. wartae и S. translucida (рис. 3б). Шулеридеиды, обитавшие только в условиях верхней сублиторали (Malz, 1966; Tesakova et al., 2012), давали вспышку численности в наиболее мелководных ее частях (Tesakova, 2013a, figs. 2, 5) за счет толерантности к солености (Wilkinson, 2008), поэтому доминирование этого семейства (“Schulerideidae-сигнал”) рассматривается в средней и поздней юре как маркер прибрежных обстановок. Об этом же говорит и полное отсутствие остракод мелкоразмерного кластера, в том числе таксонов, имеющих глазной бугорок (рис. 2).
Рис. 5.
Палеогеографическая схема Европы в среднем келловее (по Thierry, 2000) с распространением изученных остракод. 1 – размываемая суша, 2 – аккумулятивная суша, 3 – эпиконтинентальные моря, 4 – океанический бассейн, 5 – B. pumicosa, 6 – N. rimosa, 7 – N. cruciata intermedia, 8 – C. perennis s.l., 9 – P. wartae s.l., 10 – S. translucida s.l., 11 – P. khapissovi.

Находки разнообразных и порой обильных аммонитов, двустворок, брахиопод, гастропод, морских ежей, кораллов, мшанок и серпулид в нижнекелловейских слоях 1–7 (подзона Enodatum) и многочисленные аммониты из слоев 8–12 подтверждают наши выводы об очень мелководной, нормально-морской и хорошо аэрируемой обстановке (комплексы I и II).
Комплекс III (слои 13–26) мог формироваться в более глубоководных условиях, при удалении от берега, о чем свидетельствует некоторое выравнивание его таксономической структуры. Так, снизилась роль доминантов, представленных эврибионтами, не связанных с какими-либо определенными грунтами и обладающих гладкой, не скульптированной раковиной; процентное соотношение эврибионты/специалисты сместилось в сторону последних (рис. 3б). К тому же в этом интервале разреза появился один экземпляр мелкоразмерного таксона Cytheropteron sp. (рис. 2), тяготеющего к большим глубинам. Однако комплекс III по-прежнему находился в пределах верхней сублиторали, так как единичная находка Cytheropteron sp. не меняет картины абсолютного преобладания крупноразмерных остракод, а кривые разнообразия и численности имеют пики и демонстрируют низкие значения.
Снижение обилия остракод в фиксированной навеске породы (рис. 3а) может быть связано с разными причинами: с ухудшением условий обитания, растворением раковин при диагенезе, разубоживанием при высокой скорости осадконакопления (Гаврилов, 2005). Последнее представляется наиболее вероятным, так как комплекс III коррелирует с первой половиной фазы C2 – началом быстрой трансгрессии, при которой количество поступающего терригенного материала резко возросло.
Также весьма показательна смена доминантов в комплексах: если в комплексах I и II существенно преобладали шулеридеиды (маркеры близкого берега), то в комплексе III роль ведущего доминанта перешла к цитереллам. Ракушковые рода Cytherella Jones, 1849 освоили местообитания от лагун (Omatsola, 1971) до батиали (Тесакова, Савельева, 2005; Yasuhara et al., 2009 и др.) и выдерживали снижение солености до 16‰ (Wilkinson, 2008). Они также были способны прогонять воду сквозь раковину вдвое быстрее других остракод, поэтому оказались наиболее приспособленными к дизоксидным обстановкам (Whatley et al., 2003; Corbari et al., 2005). Их моновидовые ассоциации или существенное преобладание в комплексах называется “Cytherella-сигнал”, что указывает на эвтрофикацию (и, как возможное следствие, на снижение уровня кислорода) или на распреснение. В нашем случае Cytherella-сигнал может быть связан только с эвтрофией, которая возникла в фазу быстрой трансгрессии при поступлении большого количества ОВ с прибрежных маршей.
Во второй половине фазы C2 процесс сноса с берега ОВ и осадочного материала нарастал. И в дальнейшем в комплексах IV и V за счет первого фактора произошло исчезновение остракод-специалистов и усиление Cytherella-сигнала (рис. 3б), а за счет второго – снижение численности остракод в навесках породы (рис. 3а).
Выше в разрезе (слои 40–46) остракоды не найдены, и их предположительное исчезновение из палеобиотопов связывается с углекислым заражением придонных вод, которое вполне могло образоваться при окислении ОВ. А может быть, ракушковые исчезли из летописи из-за растворения их скелетов при диагенезе, благодаря высокому содержанию СО2 (Гаврилов, 2005).
Комплекс VI вновь демонстрирует “Cytherella-сигнал”, свидетельствующий об эвтрофной обстановке, но уже с нормальным уровнем СО2, что позволило остракодам сохраниться в разрезе. Следовательно, снизились поступление ОВ и скорость трансгрессии, что хорошо согласуется с реконструкцией седиментологической фазы C3.
Теперь оценим, насколько совпадают реконструкции по ракушковым и пелециподам. Биофациальный анализ, выполненный В.А. Захаровым по двустворчатым моллюскам, показал, что “бентосная биота (по крайней мере, в среднем келловее)… существовала в относительно мелководных условиях, преимущественно в пределах действия регулярных волн (выше уровня проникновения штормовых волн), в умеренно-теплой морской воде нормальной солености… Общая тенденция процесса седиментации свидетельствует о постепенном обмелении акватории в течение среднего келловея, приближении источника сноса терригенного материала и постепенной смене морских условий лагунно-морскими” (Гаврилов и др., 2015, с. 89–90).
Таким образом, результаты остракодового анализа в целом согласуются с выводами о мелководности, нормальной солености и хорошей аэрации палеобассейна Центрального Дагестана в среднем келловее, но позволяют детализировать эту реконструкцию. Во-первых, для раннего и начала среднего келловея (слои 2–12, комплексы I и II) доказывается обстановка верхней (прибрежной) части верхней сублиторали с перечисленными условиями среды. Во-вторых, выше по разрезу среднего келловея (слои 13–36, комплексы III, IV и V) показана обстановка нижней (условно глубокой) части верхней сублиторали с постепенно возрастающей эвтрофией, вплоть до растворения остракод при диагенезе (слои 38–46). В-третьих, верхи среднего келловея и нижняя часть верхнего келловея (слои 48–58, комплекс VI) характеризуются Cytherella-сигналом (высокой эвтрофией), но оценить по остракодам глубину бассейна, в котором формировался этот интервал разреза, и вероятность образования лагуны не представляется возможным.
АНАЛИЗ ПАЛЕОГЕОГРАФИЧЕСКОГО РАСПРОСТРАНЕНИЯ ИЗУЧЕННЫХ ВИДОВ
Общий список цудахарских остракод включает 9 видов, палеогеографическое распространение которых показано на рис. 5. Особый интерес представляют виды B. pumicosa, N. cruciata intermedia и N. rimosa, которые известны в Западной Европе, как в ее северной (Шотландия, Южная Англия, Нормандия, Парижский бассейн, Нидерланды, Северо-Восточная Германия), так и южной (Южная Германия, Швейцария) части, а также на западе Восточной Европы (Польша и Днепровско-Донецкая впадина) (табл. 1). При этом первое и последнее появление этих видов в летописи фиксируется именно в южной части ареала, а в более северные акватории Западной Европы они проникли позже и исчезли из них раньше. Широко расселившись в морях Западной Европы и на западе Восточной Европы, они не прошли на Русскую плиту в Среднерусское море. Следовательно, можно говорить об их происхождении, расселении и миграциях в пределах мелководных окраинных морей северной части океана Тетис и в эпиконтинентальных бассейнах Западной и частично Восточной Европы, где температурный режим был выше, чем в Среднерусском море.
Вид P. khapissovi пока следует считать эндемиком Центрального Дагестана, так как он, судя по его морфологии (Глинских, Тесакова, 2020), более всего тяготеет к плевроцитерам Узбекистана (Масумов, 1973). В меньшей степени наблюдается его сходство с плевроцитерами Западной и Восточной Европы (Triebel, 1951; Brand, Fahrion, 1962; Oertli, 1963; Błaszyk, 1967; Пяткова, Пермякова, 1978; Sheppard, 1981b; Dépeche, 1984; Liebau, 1987; Bielecka et al., 1988a; Brand, 1990). Еще дальше он отстоит от представителей рода из Южной Германии и Швейцарии (Ohmert, 2004; Franz et al., 2009, 2014, 2018; Beher et al., 2010; Dietze et al., 2017; Tesakova, 2017). И, как и в вышеописанных случаях, он не имеет ничего общего с филолиниями из Центральной России и Белоруссии (Тесакова и др., 2009; Tesakova, 2013a; Тесакова, 2014; Махнач, Тесакова, 2015). Этот вид, по-видимому, свидетельствует о связях со Средней Азией.
Особняком следует рассматривать таксоны C. perennis, P. wartae и S. translucida, которые населяли Польское и Среднерусское моря, более подверженные бореальному влиянию.
Для разреза Цудахар и Европы в целом весьма примечательно полное отсутствие общих видов с келловейскими остракодовыми фаунами южного побережья Тетис, которые проявляют высокий родовой эндемизм. Кстати, именно по этой причине в настоящей статье не рассматривались келловейские схемы по остракодам Саудовской Аравии (Dépéche et al., 1987), Израиля (Oertli, 1966), Иордании (Basha, 1980), Сирии (Кузнецова, Доброва, 1995, 1997; Доброва, 1997), Танзании (Bate, 1975, 1977), Конго (Grékoff, 1957) и Мадагаскара (Grékoff, 1963). Палеогеографическое разделение остракодовых фаун южного и северного побережья океана обусловлено биологическими особенностями группы. В онтогенезе остракод отсутствует планктонная личинка, поэтому они могут расселяться, передвигаясь только по дну и только в пределах характерного для каждого таксона батиметрического интервала. Сравнительно большие глубины, так же как и маленькие, являются естественным барьером для миграции шельфовых остракод (Tesakova, Shurupova, 2018).
ЗАКЛЮЧЕНИЕ
Анализ распространения остракод по разрезу позволил установить стратиграфическую последовательность из шести различных комплексов. Три первых объединяются распространением вида Pleurocythere khapissovi, по которому выделены одноименные слои в нижнем–среднем келловее (аммонитовые зоны Calloviense–Coronatum, подзоны Enodatum–нижняя часть Grossouvrei). Второй и третий комплексы относятся к среднекелловейской части (аммонитовые зоны Jason–нижняя часть Coronatum) зоны Neurocythere cruciata intermedia по присутствию зонального индекса остракодовых шкал Англии и Южной Германии. Четвертый и пятый комплексы отнесены к палеоэкологическому интервалу с S. translucida–C. perennis (зона Coronatum, средняя часть подзоны Grossouvrei); шестой комплекс – к интервалу с C. perennis (верхняя часть зоны Coronatum, верхи подзоны Grossouvrei–зона Athleta, подзоны Phaeinum и Proniae+Spinosum).
Условия формирования разреза Цудахар выявлены остракодовым кламп-анализом (комплексный палеоэкологический анализ с использованием разных остракодовых методик, примененных независимо). Нижний келловей и нижняя часть среднего келловея (нижняя половина слоев с P. khapissovi) накапливались в обстановке верхней (прибрежной) области верхней сублиторали с нормально-морской соленостью, хорошей аэрацией, высокой гидродинамикой и низкими темпами осадконакопления. Верхняя часть среднего келловея (верхняя половина слоев с P. khapissovi и палеоэкологический интервал с S. translucida–C. perennis) формировалась в нижней (условно глубокой) области верхней сублиторали, с высокой скоростью седиментации, высокой (и постоянно растущей) эвтрофией и пониженными значениями pH. Таксономическая бедность и малочисленность остракод этого интервала разреза объясняется разубоживанием комплексов в навесках породы за счет высоких скоростей осадконакопления. Отсутствие остракод в пробах может быть связано с низким pH и растворением при диагенезе. Верхи среднего и нижняя часть верхнего келловея (палеоэкологический интервал с C. perennis) характеризуются Cytherella-сигналом (снижение кислорода), но реконструкция глубины бассейна по остракодам в этом интервале затруднительна.
Хорологический анализ остракод показал, что все виды, кроме P. khapissovi, известны как из Западной, так и Восточной Европы. Но три из них (B. pumicosa, N. cruciata intermedia, N. rimosa), обитавшие исключительно в западноевропейских акваториях, могли попасть в Кавказское море лишь вдоль северной окраины океана Тетис или через южный пролив Польского моря, что доказывается отсутствием их находок на Русской плите. Три других вида (C. perennis, P. wartae и S. translucida) расселялись в Польском, Среднерусском и Кавказском морях через Прикаспийский бассейн. Таким образом, перечисленные остракоды имели два разных миграционных коридора, по которым происходил обмен биоты с Европой: западный – по южному краю Украинского щита в обход Русской плиты и северный – через Среднерусское море. А вид P. khapissovi, по-видимому, маркирует юго-восточное направление фаунистического обмена со Средней Азией.
Благодарности. Авторы глубоко признательны А.П. Ипполитову (ГИН РАН) за конструктивные замечания, существенно улучшившие нашу статью.
Источники финансирования. Работа выполнена в рамках тем госзадания №№ 0135-2019-0062 (ГИН РАН), АААА-А16-116033010096-8 (МГУ) и при частичной поддержке РФФИ (гранты №№ 18-05-00501, 19-05-00130) и проекта ФНИ № 0331-2019-0005.
Список литературы
Андреев Ю.Н. Меловые остракоды Средней Азии (состав, распространение, развитие, геологическое значение). Автореф. дисс. … д-ра геол.-мин. наук. М., 1988. 36 с.
Гаврилов Ю.О. Динамика формирования юрского терригенного комплекса Большого Кавказа: седиментология, геохимия, постседиментационные преобразования // Труды ГИН РАН. 2005. Вып. 555. 301 с.
Гаврилов Ю.О., Захаров В.А., Рогов М.А. и др. Цудахар // Юрские отложения центральной части Горного Дагестана. Путеводитель геологических экскурсий VI Всероссийского совещания “Юрская система России: проблемы стратиграфии и палеогеографии”, 15–20 сентября 2015 г. Ред. Черкашин В.И. Махачкала: АЛЕФ, 2015. С. 81–105.
Герасимов П.А., Митта В.В., Кочанова М.Д., Тесакова Е.М. Ископаемые келловейского яруса Центральной России. М.: Всеросс. научн.-исслед. геол. нефт. ин-т; Моск. городская станция юных натуралистов, 1996. 127 с.
Глинских Л.А., Тесакова Е.М. Первые данные об остракодах келловея Центрального Дагестана // Палеонтол. журн. 2020 (в печати).
Доброва М.Р. Стратиграфическое распространение остракод в морских отложениях Сирии и их использование для корреляции в пределах восточного Средиземноморья // Биостратиграфия и микроорганизмы фанерозоя Евразии. Труды XII Всероссийского микропалеонтологического совещания. Ред. Подобина В.М., Савина Н.И., Кузнецова К.И., Музылев Н.Г. М.: Геос, 1997. С. 113–118.
Захаров В.А., Мейен С.В., Очев В.Г., Янин Б.Т. Тафономические исследования // Современная палеонтология. Методы, направления, проблемы, практическое приложение. Справочное пособие. Ред. Меннер В.В., Макридин В.П. Т. 1. М.: Недра, 1988. С. 416–434.
Карпук М.С. Остракоды верхнего баррема–апта Горного Крыма: стратиграфическое значение и палеоэкология. Автореф. дисс. … канд. геол.-мин. наук. М.: ГИН РАН, 2016. 24 с.
Колпенская Н.Н. Юра // Практическое руководство по микрофауне. Том 7. Остракоды мезозоя. СПб.: Изд-во ВСЕГЕИ, 1999. С. 125–129.
Кузнецова К.И., Доброва М.Р. Соотношение зональных шкал юры восточного Средиземноморья по фораминиферам и остракодам // Докл. АН. 1995. Т. 340. № 2. С. 218–221.
Кузнецова К.И., Доброва М.Р. Среднеюрские бассейны восточного Средиземноморья и их микробиота (фораминиферы, остракоды) // Стратиграфия. Геол. корреляция. 1997. Т. 5. № 1. С. 32–41.
Куприянова Н.В. Нижнемеловые остракоды острова Колгуев // Стратиграфия и фауна палеозоя и мезозоя Арктики. Сб. научных трудов. СПб.: ВНИИОкеангеология, 2000. С. 92–98.
Любимова П.С. Остракоды мезозойских отложений Среднего Поволжья и Общего Сырта // Труды ВН-ИГРИ. Нов. сер. 1955. Вып. 84. С. 3–190.
Любимова П.С. Триасовые и юрские остракоды восточных районов Украины // Труды ВНИГРИ. Нов. сер. 1956. Вып. 98. С. 533–583.
Масумов А.С. Юрские остракоды Узбекистана. Ташкент: Фан, 1973. 197 с.
Махнач В.В., Тесакова Е.М. Палеогеографические реконструкции природной среды юго-восточной Белоруссии в батском–оксфордском веках // Вестник Московского университета. Сер. 4: Геология. 2015. № 2. С. 84–93.
Никитенко Б.Л. Стратиграфия, палеобиогеография и биофации юры Сибири по микрофауне (фораминиферы и остракоды). Новосибирск: Параллель, 2009. 680 с.
Пяткова Д.М., Пермякова М.Н. Фораминиферы и остракоды юры Украины. Киев: Науковая думка, 1978. 288 с.
Тесакова Е.М. Фораминиферы и остракоды: за кем будущее детальной стратиграфии? // Эволюция жизни на Земле. Материалы 4-го Международного симпозиума, 10–12 ноября 2010 г., Томск. Отв. ред. Подобина В.М. Томск: ТМЛ–Пресс, 2010. С. 411–414.
Тесакова Е.М. Юрские остракоды Русской плиты: стратиграфическое значение, палеоэкология и палеогеография. Дисс. … докт. геол.-мин. наук. М., 2014. 455 с. [неопубл.].
Тесакова Е.М. Корреляция средне-верхнеюрских остракодовых шкал Западной и Восточной Европы // Юрская система России: проблемы стратиграфии и палеогеографии. Шестое Всероссийское совещание, 15–20 сентября 2015 г., Махачкала. Научные материалы. Отв. ред. Захаров В.А. Махачкала: АЛЕФ, 2015. С. 268–272.
Тесакова Е.М., Савельева Ю.Н. Остракоды пограничных слоев юры и мела Восточного Крыма: стратиграфия и палеоэкология // Палеобиология и детальная стратиграфия фанерозоя. К 100-летию со дня рождения академика В.В. Меннера. Отв. ред. Алексеев А.С., Михайлова И.А. М.: Российская академия естественных наук, 2005. С. 135–155.
Тесакова Е.М., Cтреж А.С., Гуляев Д.Б. Новые остракоды из нижнего келловея Курской области // Палеонтол. журн. 2009. № 3. С. 25–36.
Тесакова Е.М., Шурупова Я.А., Устинова М.А. Стратиграфия келловея и нижнего оксфорда разреза Михайловцемент (Рязанская обл.) по микрофауне и наннопланктону // Труды ГИН РАН. 2017. Вып. 615. С. 264–300.
Шорников Е.И., Михайлова Е.Д. Остракоды Bythocytheridae раннего этапа развития. Сравнительная морфология, палеоэкология, пути эволюции. М.: Наука, 1990. 278 с.
Babinot J.-F., Lethiers F. Importance du taxon generique chez les Ostracodes fossiles // Bull. Soc. geol. Fr. 1984 (7). V. 26. № 4. P. 591–602.
Basha S.H. Ostracoda from the Jurassic system of Jordan (including a stratigraphical outline) // Rev. Esp. Micropaleontol. 1980. V. 12. № 2. P. 231–254.
Bate R.H. Ostracods from Callovian to Tithonian sediments of Tanzania, East Africa // Bull. Brit. Mus. Nat. Hist. Geol. 1975. V. 26. № 5. P. 161–223.
Bate R.H. Upper Jurassic Ostracoda from Tanzania, East Africa // VI Colloque African Micropaleontologie, Tunis. Ann. Min. Geol. Tunis. 1977. № 28. P. 163–183.
Bate R.H. Middle Jurassic (Aalenian–Bathonian) // Ostracods in British Stratigraphy. Eds. Whittaker J.E., Hart M.B. Micropaleontol. Soc. Spec. Publ. Geol. Soc. London, 2009. P. 199–223.
Beher E., Brand E., Franz M. Bathonian and Lower Callovian ostracods of Albstadt-Pfeffingen (Middle Jurassic, Baden-Württemberg, Germany) // Palaeodiversity. 2010. V. 3. P. 43–57.
Bielecka W., Styk O., Błaszyk J., Kopik J. Middle Jurassic. Ostracoda Latreille, 1806 // Geology of Poland. Atlas of guide and characteristic fossils. Ed. Malinowska L. Pt. 2b. Mesozoic Jurassic. Warsaw: Wydawnictwa Geologiczne, 1988a. P. 165–179.
Bielecka W., Styk O., Błaszyk J., Kopik J. Upper Jurassic. Ostracoda Latreille, 1806 // Geology of Poland. Atlas of guide and characteristic fossils. Ed. Malinowska L. Pt. 2b. Mesozoic Jurassic. Warsaw: Wydawnictwa Geologiczne, 1988b. P. 359–376.
Błaszyk J. Middle Jurassic ostracods of the Czestochowa region (Poland) // Acta Paleontol. Pol. 1967. V. 12. № 1. P. 1–75.
Bodergat A.-M. Les ostracodes marins du Jurassique Europeen. Utilisation stratigraphique // Bull. Centre Rech. Elf Explor. Prod. Mem. 1997. V. 17. P. 197–223.
Boomer I., Eisenhauer G. Ostracod faunas as palaeoenvironmental indicators in marginal marine environments // The Ostracoda: Applications in Quaternary Research. Eds. Chivas A., Holmes J. American Geophysical Monograph. 2002. № 131. P. 135–150.
Brand E. Biostratigraphische Untergliederung des Ober-Bathonium im Raum Hildesheim, Nordwestdeutschland mittels Ostracoden und Korrelation ihrer Vertikalreichweiten mit Ammoniten-Zonen // Geol. Jb. A 121. 1990. S. 119–273.
Brand E., Fahrion H. Dogger NW-Deutschlands // Arbeitskreis Deutscher Mikropalaontologen: Leitfossilien der Mikropalaontologie. Berlin: Borntraeger, 1962. P. 123–158.
Buck E. Stratigraphisch wichtige Ostrakoden im Lias u. Dogger SW-Deutschlands; Freiburg i. Br. 1954 [unpublished table].
Corbari L., Mesmer-Dudons N., Carbonel P., Massabuau J.-C. Cytherella as a tool to reconstruct deep-sea paleo-oxygen levels: the respiratory physiology of the platycopid ostracod Cytherella cf. abyssorum // Marine Biology. 2005. V. 147. P. 1377–1386.
Dépêche F. Description de quelques ostracodes nouveaux de bathonien Lorrain // Rev. Micropaleontol. 1969. № 2. P. 107–118.
Dépêche F. Etude des Ostracodes du Dogger du Bassin Parisien. I. Ostracodes du Bathonien inferieur et de la base du Bathonien moyen de Port-en-Bessin // Rev. Micropaleontol. 1973. № 4. P. 213–226.
Dépêche F. Les ostracodes d’une plate-forme continentale au Jurassique: recherches sur le bathonien du Bassin Parisien // Mém. Sci. Terre. Univ. Pierre-et-Marie-Curie, Paris. No 84 38. Paris, 1984. P. 1–419 [unpublished thesis].
Dépêche F. Lias supérieur, Dogger, Malm // Atlas des ostracodes de France. Bull. Centr. Rech. Explor.-Prod. Elf-Aquitaine, Mém. 1985. 9. P. 119–145.
Dépêche F., Le Nindre Y., Manivit J., Vaslet D. Les ostracodes du jurassique d’Arabie Saoudite centrale: systématique, répartition stratigraphique et paléogéographique // Geobios. Mém. spécial. 1987. № 9. P. 221–275.
Dietze V., Franz M., Kutz M., Waltschew A. Stratigraphy of the Middle Jurassic Sengenthal Formation of Polsingen-Ursheim (Nördlinger Ries, Bavaria, Southern Germany) // Palaeodiversity. 2017. V. 10. P. 49–95. https://doi.org/10.18476/pale.v10.a5
Franz M., Tesakova E., Beher E. Documentation and revision of the index ostracods from the Lower and Middle Jurassic in SW Germany according to BUCK (1954) // Palaeodiversity. 2009. V. 2. P. 119–167.
Franz M., Beher E., Dietl G. The Bathonian and Early Callovian Ostracoda of Baden-Wuerttemberg, southern Germany // N. Jb. Geol. Paläontol. Abh. 2014. V. 274. № 2/3. P. 149–185.
Franz M., Ebert M., Stulpinaite R. Aalenian–Lower Bajocian (Middle Jurassic) ostracods from the Geisingen clay pit (SW Germany) // Palaeodiversity. 2018. V. 11. P. 59–105.
Grékoff N. Ostracodes du Bassin du Congo. Crétacé inférieur du Nord du Bassin // Ann. Mus. Royal Congo Belge, 8, Sciences Geologiques. 1957. V. 19. № 1–10. P. 1–97.
Grékoff N. Contribution à l'étude des Ostracodes du Mésozoïque moyen (Bathonien – Valanginien) du Bassin de Majunga, Madagascar // Rev. I.F.P. Paris. 1963. P. 1709–1762.
Herngreen G.F.W., Lissenberg Th., de Boer K.F., Romein B.J., Wijker N.C. Middle callovian beds in the Achterhoek, Eastern Netherlands // Med. riks geol. dienst. 1983. V. 37. № 3. P. 1–29.
Huber B., Müller B., Luterbacher H. Mikropaläontologische Untersuchungen an der Callovien/Oxfordien-Grenze im Schweizer Jura und auf der Schwäbischen Alb (vorläufige Mitteilung) // Eclogae geol. Helv. 1987. V. 80. № 2. P. 449–459.
Kilenyi T. The Jurassic. Part III, Callovian–Portlandian // A stratigraphical index of British Ostracoda. Eds. Bate R., Robinson E. Geol. J. Spec. Iss. 1978. V. 8. P. 259–298.
Kolasa J., Hewitt C.L., Drake J.A. Rapoport’s rule: an explanation or a byproduct of the latitudinal gradient in species richness?? // Biodiversity and Conservation. 1998. V. 7. P. 1447–1455.
Liebau A. Skulptur-Beziehungen jurassischer Progonocytheridae (Ostracoden) // N. Jb. Geol. Paläontol. Abh. 1987. V. 176. № 1. S. 15–47.
Lutze G. F. Zur Stratigraphie und Palaontologie des Callovien und Oxfordien in Nordwest-Deutschland // Geol. Jahr. 1960. V. 77. P. 391–532.
Malz H. Zur Kenntnis einiger Ostracoden-Arte–n der Gattungen Kinkelinella und Praeschuleridea // Senckenbergiana lethaea. 1966. V. 47. № 4. P. 385–404.
Malz H. Ostracoden-Studien im Dogger, 8: Die Arten der Gattung Lophocythere, ihre stratigraphische und regionale Verbreitung // Senck. leth. 1975. V. 56. № 2–3. S. 123–145.
Oertli H. J. Faunes d’Ostracodes du Mesozoique de France. Leiden (Brill), 1963. 57 p.
Oertli H.J. On the Jurassic microfaunas of Israel; Ostracoda // Maync W. Microbiostratigraphy of the Jurassic of Israel. Geol. Surv. Israel. Bull. 1966. № 40. P. 10–22, 44–55.
Ohmert W. Ammoniten-Faunen im tiefen Unter-Bajocium des Reutlinger Gebiets (mittlere Schwäbische Alb) [mit einem Beitrag zur Ostracoden-Stratigraphie] // Jahreshefte des Landesamts für Geologie, Rohstoffe und Bergbau in Baden-Württemberg. 2004. V. 40. P. 9–141.
Olempska E., Błaszyk J. A Boreal Ostracod Assemblage from the Callovian of the Lukow Area, Poland // Acta Palaeontol. Pol. 2001. V. 46. № 4. P. 553–582.
Omatsola M.E. On the occurrence of Cytherellids (Ostracodes, Crustacea) in a brackish-water environment // Bull. Geol. Instit. Univ. Uppsala. New Ser. 1971. V. 2. № 10. P. 91–95.
Pokorný V. The diversity of fossil ostracode communities as an indicator of palaeogeographic conditions // Paléoécologie Ostracodes Pau 1970. Ed. Oertli H.J. Bull. Centre Rech. Pau-SNPA. 1971. V. 5. P. 45–61.
Schudack U. Revision, Dokumentation und Stratigraphie der Ostracoden des nordwestdeutschen Oberjura und Unter-Berriasium // Berliner geowiss. Abh. 1994. Ser. E. V. 11. 193 p.
Schudack U., Schudack M.E. Ostracods from the Upper Jurassic (Oxfordian–Tithonian) of southern Germany // J. Micropalaeontol. 2000. V. 19. P. 97–112.
Schudack U., Schudack M., Marty D., Comment G. Kimmeridgian (Late Jurassic) ostracods from Highway A16 (NW Switzerland): taxonomy, stratigraphy, ecology, and biogeography // Swiss J. Geosciences. 2013. V. 106. P. 371–395.
Sheppard L.M. Bathonian ostracod correlation north and south of the English Channel, with the description of two new Bathonian ostracods // Microfossils of Recent and Fossil Shelf Seas. Eds. Neale J.W., Brasier M.D. Chichester: Ellis Horwood, 1981a. P. 71–83.
Sheppard L.M. Middle Jurassic Ostracoda from Southern England and Northern France. Thesis Ph.D. University College London. 1981b. 214 p. [unpubl.]
Shurupova Ya.A., Tesakova E.M. Phylogeny of the subgenus Palaeocytheridea (Palaeocytheridea) (Progonocytheracea, Pleurocytheridae) from Middle Jurassic of Europe // 18th Int. Symp. on Ostracoda (ISO-18). University of California, Santa-Barbara, August 27–31, 2017a. Abstract book. P. 77–78.
Shurupova Y.A., Tesakova E.M. Detailed biostratigraphic scales as based on the palaeobiogenetical approach (an example of the Upper Bajocian–Lower Bathonian ostracod scale of the Russian Platform) // Volumina Jurassica. 2017b. V. 15. P. 1–17.
Shurupova Ya.A., Tesakova E.M. Species interrelatedness in the Genus Lophocythere Silvester-Bradley, 1948 (Ostracoda) in the Late Callovian of the Russian Plate // Paleontol. J. 2019. V. 53. № 9. P. 54–59. https://doi.org/10.1134/S0031030119090144
Sylvester-Bradley P.C. Bathonian ostracods from the Boueti Bed of Langton Herring, Dorset // Geol. Mag. 1948. V. 85. P. 185–204.
Tesakova E.M. Callovian and Oxfordian ostracods from the Central Region of the Russian Plate // Paleontol. J. 2003. V. 37. Suppl. 2. P. 107–227.
Tesakova E. Late Callovian and Early Oxfordian ostracods from the Dubki section (Saratov area, Russia): implications for stratigraphy, paleoecology, eustatic cycles and palaeobiogeography // Neues Jahrbuch für Geologie und Paläontologie. Abhandlungen. 2008. V. 249. № 1. P. 25–45.
Tesakova E.M. Ostracode-based reconstruction of paleodepths in the Early Callovian of the Kursk Region, Central Russia // Paleontol. J. 2013a. V. 47. № 10. P. 1–16.
Tesakova E.M. Ostracods of the genus Palaeocytheridea Mandelstam in the Middle and Upper Jurassic of Europe: 1. Development of ideas on the content of the genus and the results of its revision // Paleontol. J. 2013b. V. 47. № 3. P. 256–271.
Tesakova E.M. Ostracods of the genus Palaeocytheridea Mandelstam in the Middle and Upper Jurassic of Europe: 3. Stratigraphy and paleobiogeography // Paleontol. J. 2014. V. 48. № 1. P. 53–57.
Tesakova E.M. Biostratigraphie du Jurassique moyen des sondages Benken et Schlattingen-1 (Nord de la Suisse) ètablie sur la base de la répartition des ostracodes // NAGRA Arbeitsbericht NAB. 2017. V. 17–41. 79 p.
Tesakova E.M., Shurupova Ya.A. Ostracod analysis of Callovian and Lower Oxfordian deposits of the Mikhailovtsement Section (Ryazan Region): methods and results // Paleontol. J. 2018. V. 52. № 13. P. 1547–1568.
Tesakova E.M., Franz M., Baykina E., Beher E. A new view on Bathonian ostracods of Poland // Senckenbergiana lethaea. 2008. V. 88. № 1. P. 55–65.
Tesakova E.M., Demidov S.M., Guzhov A.V., Rogov M.A., Kiselev D.N. Middle Oxfordian–Lower Kimmeridgian ostracod zones from the Mikhalenino section (Kostroma region) and their comparison with synchronous strata of the Eastern and Western Europe // Neues Jahrb. Geol. Paläontol. Abh. 2012. V. 266. № 3. P. 239–249.
Tesakova E.M., Karpuk M.S., Brovina E.A. Palaeodepth reconstructions on inner and outer shelf using Mesozoic ostracodes // Second Meeting of Asian Ostracodologists (Second Asian Ostracodologists’ Meeting), Yunnan University, June 27–30, 2016. Abstract Book. Ed. Dayou Zhai. 2016. P. 69–71.
Thierry J. Middle Callovian // Atlas Peri-Tethys, Palaeogeographical maps. Explanatory notes. Coord. Crasquin S. Paris: CCGM/CGMW, 2000. P. 71–83.
Triebel E. Ostracoden aus dem hoheren Dogger Deutschlands // Abh. senck. naturf. Ges. 1951. V. 485. P. 87–101.
Whatley R.C. Scottish Callovian and Oxfordian Ostracoda // Bull. Brit. Mus. Nat. Hist. Geol. 1970. V. 19. № 6. P. 299–358.
Whatley R.C., Ballent S. A review of the Mesozoic ostracod genus Lophocythere and its close allies // Palaeontology. 2004. V. 47. № 1. P. 81–108.
Whatley R.C., Ballent S., Armitage J. Callovian Ostracoda from the Oxford Clay of southern England // Revista Española de Micropaleontologia. 2001. V. 33. № 2. P. 135–162.
Whatley R.C., Pyne R.S., Wilkinson I.P. Ostracoda and palaeo-oxygen levels, with particular reference to the Upper Cretaceous of East Anglia // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003. V. 194. P. 355–386.
Wienholz E. Neue Ostracoden aus dem norddeutschen Callovian // Freiberger Forschungshefte. Reihe C. 1967. № 213. P. 23–51.
Wilkinson I.P. The effect of environmental change on early Aptian ostracods faunas in the Wessex Basin, southern England // Rev. Micropaléontologie. 2008. V. 51. P. 259–272.
Wilkinson I.P., Whatley R.C. Upper Jurassic (Callovian–Portlandian) // Ostracods in British Stratigraphy. Eds. Whittaker J.E, Hart M.B. Micropaleontol. Soc. Spec. Publ. Geol. Soc. London, 2009. P. 241–287.
Yasuhara M., Okahashi H., Cronin T.M. Taxonomy of Quaternary deep-sea ostracods from the Western North Atlantic Ocean // Palaeontology. 2009. V. 52. Pt. 4. P. 879–931.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция