

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 6, стр. 125-170
Новые седиментологические, магнитостратиграфические и биостратиграфические данные по разрезу кампана–маастрихта горы Бешкош, Юго-Западный Крым
Е. Ю. Барабошкин 1, *, А. Ю. Гужиков 2, Г. Н. Александрова 3, В. А. Фомин 2, Б. Г. Покровский 3, В. А. Грищенко 2, А. Г. Маникин 2, Е. В. Наумов 2
1 Московский государственный университет им. М.В. Ломоносова
Москва, Россия
2 Саратовский государственный университет им. Н.Г. Чернышевского
Саратов, Россия
3 Геологический институт РАН
Москва, Россия
* E-mail: barabosh@geol.msu.ru
Поступила в редакцию 18.10.2019
После доработки 15.11.2019
Принята к публикации 20.11.2019
Аннотация
Проведено комплексное исследование кампан-маастрихтских отложений горы Бешкош (Юго-Западный Крым). Изучены находки головоногих моллюсков, диноцисты, впервые получены изотопные, палео- и петромагнитные данные; проведен ихнофациальный и седиментологический анализ. Макро- и микропалеонтологические данные позволили уточнить стратиграфическое расчленение разреза, подтвердив позднекампанский–маастрихтский возраст пород. Палеомагнитные исследования не выявили аналогов магнитного хрона C31r. На фоне доминирующей нормальной полярности установлена единственная магнитозона обратного знака, предположительно идентифицированная как аналог хрона C30r. Изучение диноцист выявило резкую смену их комплексов в нижней части разреза, выше которой встречен аммонит Pachydiscus (P.) neubergicus neubergicus (Hauer) – вид-маркер границы кампанского и маастрихтского ярусов. Совместный анализ био-, магнитостратиграфических и седиментологических данных позволил установить уровень кампан-маастрихтской границы в разрезе и предположить присутствие неизвестного ранее перерыва в осадконакоплении в пограничном интервале. Данные по стабильным изотопам согласуются с предполагаемым уровнем перерыва.
ВВЕДЕНИЕ
Разрезы кампана–маастрихта в междуречье Чурюк-Су–Бодрак хорошо известны в литературе. К ним принадлежит разрез юго-западного склона горы Бешкош, краткое описание которого приведено в работах (Маслакова, Липник, 1971; Заклинская, Найдин, 1985; Alekseev, Kopaevich, 1997). Это один из наиболее полных и мощных разрезов терминального кампана и маастрихта Юго-Западного Крыма. В работах предшественников (Алексеев, 1989; Alekseev, Kopaevich, 1997) в нем выделяется пять литологических пачек, прослеживаемых и в других разрезах данного региона. Эти пачки были отнесены к верхнему кампану (пачка XIX), нижнему маастрихту (пачки XX–XXI) и верхнему маастрихту (XXII–XXIII) по комплексам макрофаунистических остатков (аммониты, белемниты, двустворки: Маслакова, Липник, 1971; Заклинская, Найдин, 1985; Алексеев, 1989; Alekseev, Kopaevich, 1997), а также фораминифер (Маслакова, Липник, 1971; Alekseev, Kopaevich, 1997). Спорово-пыльцевые комплексы из верхнемаастрихтских отложений впервые были описаны Е.Д. Заклинской и Д.П. Найдиным (1985).
Целью настоящего исследования являлось комплексное изучение разреза для обоснования положения ярусных и подъярусных границ кампана и маастрихта, что обусловлено необходимостью разработки современной стратиграфической схемы.
Е.Ю. Барабошкиным, А.Ю. Гужиковым, А.Г. Маникиным, В.А. Грищенко и Е.В. Наумовым проведено полевое изучение и опробование разреза. Ориентированные штуфы для палеомагнитного и петромагнитного анализа взяты со 134 стратиграфических уровней, при общей мощности разреза 163 м. Палинологические пробы взяты с 60 уровней, а шлифы изготовлены с 25 уровней. Изотопные определения получены по 14 образцам из пограничного интервала кампана–маастрихта. Разные виды анализа выполнены по системе “образец в образец”. При магнитостратиграфическом анализе учтены палеомагнитные и петромагнитные определения по 42 образцам, отобранным В.А. Фоминым и М.В. Пименовым (Саратовский государственный университет – СГУ) в 2008 г.
Описание разреза, седиментологические, петрографические, ихнологические исследования, а также изучение остатков головоногих моллюсков и остальной макрофауны проведены Е.Ю. Барабошкиным (Московский государственный университет им. Ломоносова – МГУ), магнитостратиграфические данные получены А.Ю. Гужиковым, В.А. Грищенко, Е.В. Наумовым и В.А. Фоминым (СГУ), палинологический анализ выполнен Г.Н. Александровой (Геологический институт РАН – ГИН РАН), изотопные определения – Б.Г. Покровским (ГИН РАН).
СТРОЕНИЕ РАЗРЕЗА
Разрез расположен на юго-западе Крымского полустрова, к западу от города Бахчисарай (рис. 1а, 1б). Слои залегают согласно и наклонены к СЗ. Согласно данным разных групп исследователей (включая наши измерения), азимуты падения варьируют от 280° до 315°, а углы падения от 6°30′′ до 15°. Разрез изучался вдоль большой промоины на юго-западном склоне горы, его начало имеет координаты 44°44′04.3′′ с.ш., 43°55′22.1′′ в.д., высота над уровнем моря 327 м.
Рис. 1.
(а, б) Местонахождение разреза и (в) литологическая характеристика разреза, распределение важнейших ихнофоссилий и находок головоногих моллюсков.

Разрез сложен мергелями, переходящими вверх в алевритистые мергели, а затем в песчаники. Отложения полностью биотурбированы, биотурбационный индекс (Bi) равен 6. Породы формируют характерный рельеф горы, состоящий из пологих и крутых склонов, в разной степени задернованных, и субвертикальных обрывов. Типичной чертой рельефа является его ступенчатый характер, связанный с горизонтами цементации, включая уровни твердого дна, к которым приурочены многочисленные норы Thalassinoides, в разной степени окремнелые. Сочетание состава пород и рельефа позволило А.С. Алексееву и Л.Ф. Копаевич (Alekseev, Kopaevich, 1997) наметить расчленение разреза, которого мы старались в основном придерживаться. Мощности пачек измерялись в поле и сравнивались с данными фототеодолитных измерений, предоставленных М.Ю. Никитиным (МГУ). В разрезе снизу вверх обнажается следующая последовательность пачек (рис. 1в, 2).
Пачка ХIХ. Пачка разделяется нами на две подпачки. Нижняя подпачка XIXa (видимая мощность 15 м) отличается от верхней подпачки XIXb (мощность 18.5 м) тем, что в ее разрезе чередуются прослои более глинистых мергелей светло-серого цвета, мощностью по 1–2 м и менее глинистых почти белых мергелей примерно такой же мощности. Верхняя подпачка (XIXb) менее глинистая, белого цвета. В шлифах породы обеих подпачек – это фораминиферовые вакстоуны с примесью мелких фрагментов иглокожих, единичных сферических радиолярий и неопределимых биокластов (табл. I , фиг. А). Присутствуют единичные мелкоалевритовые зерна кварца. Планктонные глобигериноподобные фораминиферы преимущественно мелкие, доминируют над бентосными секреционными (отношение планктон/бентос (П/Б) в 5 полях зрения при объективе ×5 составляет 90–95%), многие из них окремнены или пиритизированы. Встречаются остатки губок, призматический слой иноцерамов. Из биотурбаций определены Zoophycos и Planolites. Из данной пачки указываются находки Belemnitella mucronata senior Now., Pachydiscus koeneni Gross., Baculites аnсерs Lam. (Маслакова, Липник, 1971, с. 97), которые датированы верхним кампаном (Заклинская, Найдин, 1985, с. 80). Породы слагают сильно задернованный и террасированный склон в основании горы, местами обнажаясь в промоинах. Суммарная видимая мощность пачки около 33.5 м.
Таблица I . Петрографическая характеристика разреза. А – обр. 3110-10, фораминиферовый вакстоун, пачка XIXa; Б – обр. 3110-30, фораминиферовый вакстоун, пачка XXb; В – обр. 3110-60, фораминиферовый вакстоун, пачка XXb; Г – обр. 3136-14, алевритистый фораминиферовый вакстоун, пачка XXс; Д – обр. 3136-30, окремнелый алевритистый фораминиферовый вакстоун, нижняя часть пачки XXII; Е – обр. 3136-45, окремнелый песчаный фораминиферовый вакстоун, верхняя часть пачки XXII; Ж – обр. 3171-15, окремнелый песчаный фораминиферовый вакстоун, пачка XXIII; З – обр. 3171-20, глауконит-кварцевый песчаник, пачка XXV. Сокращения: биокласты: а – водоросли; b – двустворки; br – мшанки; e – иглокожие, фораминиферы: bf – бентосные, pf – планктонные; p – пелоиды; другие зерна: q – кварц; gl – глауконит. Все фото сделаны в неполяризованном свете на микроскопе Zeiss Axio Lab A1pol камерой Axiocam ICc5.

Таблица II. Ихнологическая характеристика разреза. А – Zoophycos isp. (zo) и Chondrites isp. (ch), пачка XXI; Б – Pilichnus isp. (p), пачка XXb; В – Lepidenteron lewesiensis (Mant.), подчеркнутый чешуей и костями рыб (пунктир), пачка XXIII; Г – Arenicolites isp. (a), пачка XXb; Д – окремненные норы Thalassinoides suevicus (Reith) (t) в горизонте твердого дна, пачка XXI; Е – Schaubcylindrichnus isp. (s), пачка XXIII; Ж – нора Thalassinoides isp. (пунктир), заполненная раковинами Mimachlamys sp., пачка XXIII; З – темпеститы, состоящие из одиночных створок Mimachlamys sp., содержащие устрицу Pycnodonte mirabile (Rous.) (pm), пачка XXIII. Фото Е.Ю. Барабошкина, 2017–2019 гг.

Таблица III. Находки головоногих моллюсков из нижнего маастрихта разреза Бешкош. 1 – Pachydiscus (P.) neubergicus neubergicus (Hauer, 1858), обр. МЗ МГУ 137/1: а – поперечное сечение, б – сбоку, в – вентральная сторона; осыпь в нижней части подпачки ХХа; сборы Н.Г. Зверькова (МГУ), 2016 г.; 2 – Pachydiscus (P.) armenicus Atabekian et Akopian, 1969, обр. МЗ МГУ 137/2, сбоку; слой 3136/14, 2 м ниже кровли подпачки ХХc; сборы Е.Ю. Барабошкина, 08.08.2017 г.; 3–4 – Belemnella gracilis (Arkhangelsky, 1912): 3 – обр. МЗ МГУ 137/3: а – вентральная сторона, б – сбоку, в – брюшная сторона, г – строение брюшной щели; 4 – молодой экземпляр, обр. МЗ МГУ 137/4: а – вентральная сторона, б – сбоку, в – брюшная сторона; слой 3136/7-8, 7 м выше подошвы подпачки ХХc; сборы Е.Ю. Барабошкина, 09.08.2019 г. Образцы покрыты хлористым аммонием. Фото Е.Ю. Барабошкина. Все образцы хранятся в Музее землеведения МГУ, коллекция 137.

Таблица IV. Палиноморфы разреза Бешкош. Все фигуры в одном увеличении. 1 – Eatonicysta hapala Schiøler et Wilson, 1993, т.н. 3110, проба 26; 2 – Hystrichosphaeropsis sp., т.н. 3110, проба 26; 3 – Conneximura fimbriata (Morgenroth, 1968) May, 1980, т.н. 3110, проба 26; 4 – Rottnestia wetzelii (Deflandre, 1937) Slimani, 1994, т.н. 3110, проба 26; 5 – Isabelidinium sp., т.н. 3110, проба 26; 6 – Exochosphaeridium cf. muelleri Yun Hyesu, 1981, т.н. 3110, проба 31; 7 – Hystrichosphaeridium tubiferum (Ehrenberg, 1837) Deflandre, 1937, т.н. 3110, проба 31; 8 – Cribroperidinium sp., т.н. 3110, проба 27; 9 – Cribroperidinium cf. cooksoniae Norvick, 1976, т.н. 3110, проба 31; 10 – Exochosphaeridium sp., т.н. 3110, проба 19; 11 – Circulodinium distinctum (Deflandre et Cookson, 1955) Jansonius, 1986, т.н. 3110, проба 28; 12 – cf. Cassidium fragile (Harris, 1965) Drugg, 1967, т.н. 3110, проба 35; 13 – Leberidocysta chlamydata (Cookson et Eisenack, 1962) Stover et Evitt, 1978, т.н. 3110, проба 27; 14 – Pervosphaeridium cf. tubuloaculeatum Slimani, 1994, т.н. 3110, проба 33; 15, 20 – Palaeostomocystis foveolata Slimani, 1994, т.н. 3110, проба 27; 16 – Cladopyxidium velatum Marheinecke, 1992, т.н. 3110, проба 42; 17 – Areoligera senonensis Lejeune-Carpentier, 1938, т.н. 3110, проба 40; 18, 19 – Triblastula wilsonii Slimani, 1994, т.н. 3110, проба 40; 21 – Cladopyxidium paucireticulatum Slimani, 1994, т.н. 3110, проба 45; 22 – Cladopyxidium saeptum (Morgenroth, 1968) Stover et Evitt, 1978, т.н. 3110, проба 40; 23 – Alisogymnium euclaense (Cookson et Eisenack, 1970) Lentin et Vozzhennikova, 1990, т.н. 3136, пр-оба 25; 24, 31 – Batiacasphaera sp.: 24 – т.н. 3110, проба 42, 31 – т.н. 3136, проба 25; 25 – Impagidinium sp., т.н. 3110, проба 45; 26, 27 – Membranigonyaulax wilsonii Slimani, 1994: 26 – т.н. 3110, проба 42, 27 – т.н. 3110, проба 45; 28 – Palaeocystodinium bulliforme Ioannides, 1986, т.н. 3110, проба 42; 29 – Fromea sp. 1, т.н. 3110, проба 33; 30 – Fromea cf. laevigata (Drugg, 1967) Stover et Evitt, 1978, т.н. 3110, проба 45; 32 – Cymatiosphaera sp., т.н. 3110, проба 45; 33 – Neosphaerodictyon filosum Slimani, 2003, т.н. 3110, проба 45; 34 – Trigonopyxidia ginella (Cookson et Eisenack, 1960) Downie et Sarjeant, 1965, т.н. 3136, проба 25.

Таблица V. Палиноморфы разреза Бешкош. Все фигуры в одном увеличении. 1, 2 – Cerodinium leptodermum (Vozzhennikova, 1963) Lentin et Williams, 1987, т.н. 3110, проба 45; 3 – Cerodinium albertii (Corradini, 1973) Lentin et Williams, 1987, т.н. 3110, проба 45; 4, 10 – Isabelidinium cooksoniae (Alberti, 1959) Lentin et Williams, 1977: 4 – т.н. 3110, проба 45, 10 – т.н. 3110, проба 47; 5 – Chatangiella sp., т.н. 3110, проба 49; 6, 7 – Cerodinium diebelii (Alberti, 1959) Lentin et Williams, 1987: 6 – т.н. 3110, проба 45, 7 – т.н. 3110, проба 26; 8 – Florentinia ferox (Deflandre, 1937) Duxbury, 1980, т.н. 3110, проба 49; 9 – Microdinium carinatum (Below, 1987) Lentin et Williams, 1989, т.н. 3110, проба 49; 11 – Dinogymnium acuminatum Evitt et al., 1967, т.н. 3110, проба 45; 12 – Trithyrodinium sp., т.н. 3110, проба 63; 13 – Alterbidinium recticorne Vozzhennikova, 1967, т.н. 3110, проба 49; 14 – Alterbidinium acutulum (Wilson, 1967) Lentin et Williams, 1985, т.н. 3136, проба 23; 15 – Alterbidinium minus (Alberti, 1959) Lentin et Williams, 1985, т.н. 3110, проба 49; 16 – Microdinium carpentierae Slimani, 1994, т.н. 3110, проба 45; 17 – Gillinia hymenophora Cookson et Eisenack, 1960, т.н. 3110, проба 45; 18, 19 – Alisocysta circumtabulata (Drugg, 1967) Stover et Evitt, 1978: 18 – т.н. 3110, проба 45, 19 – т.н. 3110, проба 47; 20 – Phanerodinium sp., т.н. 3110, проба 49; 21 – Alterbidinium varium Kirsch, 1991, т.н. 3110, проба 45; 22 – Pterodinium cingulatum (Wetzel, 1933) Below, 1981, т.н. 3110, проба 45; 23 – Lanternosphaeridium lanosum Morgenroth, 1966, т.н. 3110, проба 45; 24 – Turnhosphaera hypoflata (Yun Hyesu, 1981) Slimani, 1994, т.н. 3110, проба 45; 25 – Leberidocysta? microverrucosa Slimani, 1994 – Leberidocysta? flagellichnia Schiøler, 1993, т.н. 3110, проба 45; 26 – Fromea laevigata (Drugg, 1967) Stover et Evitt, 1978, т.н. 3110, проба 49; 27, 28 – Palaeostomocystis reticulata Deflandre, 1937, т.н. 3110, проба 45; 29 – Fromea fragilis (Cookson et Eisenack, 1962) Stover et Evitt, 1978, т.н. 3110, проба 49; 30 – Spiniferites ramosus (Ehrenberg, 1837) Mantell, 1854, т.н. 3110, проба 45; 31 – Pervosphaeridium monasteriense Yun Hyesu, 1981, т.н. 3110, проба 45; 32 – Palaeocystodinium sp., т.н. 3110, проба 45; 33 – Operculodinium sp., т.н. 3110, проба 35.

Таблица VI. Палиноморфы разреза Бешкош. Все фигуры в одном увеличении. 1 – Trithyrodinium suspectum (Manum et Cookson, 1964) Davey, 1969, т.н. 3110, проба 49; 2 – Palaeoperidinium pyrophorum (Ehrenberg, 1837) Sarjeant, 1967, т.н. 3110, проба 63; 3 – Spongodinium delitiense (Ehrenberg, 1837) Deflandre, 1936, т.н. 3110, проба 51; 4 – Membranilarnacia liradiscoides (Wetzel, 1933) Downie et Sarjeant, 1965, т.н. 3136, проба 9; 5 – Glaphyrocysta ordinata (Williams et Downie, 1966) Stover et Evitt, 1978, т.н. 3110, проба 51; 6 – Cannosphaeropsis utinensis Wetzel, 1933, т.н. 3136, проба 23; 7 – Pierceites pentagonus (May, 1980) Habib et Drugg, 1987, т.н. 3136, проба 11; 8 – Senegalinium obscurum (Drugg, 1967) Stover et Evitt, 1978, т.н. 3110, проба 51; 9 – Phanerodinium? sonciniae Marheinecke, 1992, т.н. 3110, проба 49; 10 – Palaeocystodinium australinum (Cookson, 1965) Lentin et Williams, 1976, т.н. 3110, проба 63; 11 – Fromea chytra (Drugg, 1967) Stover et Evitt, 1978, т.н. 3110, проба 51; 12 – Microdinium sp., т.н. 3136, проба 11; 13, 14, 15, 16 – Riculacysta? pala Kirsch, 1991, 13, 14 – т.н. 3110, проба 31, 15, 16 – т.н. 3110, проба 28; 17 – Exochosphaeridium phragmites Davey et al., 1966, т.н. 3110, проба 45; 18 – Operculodinium? severinii (Cookson et Cranwell, 1967) Islam, 1983, т.н. 3110, проба 49.

Пачка ХХ. В работе (Alekseev, Kopaevich, 1997) пачка XX разделена на три подпачки. Нами они также выделяются, однако изучение пород в шлифах показало существенные отличия подпачки XXa от остальных подпачек.
Подпачка ХХа. Белые биотурбированные мергели, аналогичные подпачке XIXb, чередующиеся с мергелями, содержащими алевритовую примесь зерен кварца и единичные зерна глауконита алевритовой размерности. Мощности прослоев 1–2 м. Алевритовая примесь появляется на уровне обр. 3010/45, а начиная с обр. 3010/54 она присутствует регулярно, и поэтому граница между подпачками XXa и XXb проводится на уровне обр. 3010/54. В шлифах мергели представляют собой фораминиферовые вакстоуны с фрагментами иглокожих, единичных спикул и радиолярий, неопределимых биокластов (табл. I , фиг. Б). Алевритистые мергели аналогичны подпачке XXb (см. ниже). Мелкие планктонные фораминиферы продолжают доминировать над бентосными. Часто первые окремнены и составляют иногда значительную часть “алевритовой примеси”, выделенной при растворении образцов (Alekseev, Kopaevich, 1997). На уровне нижней части пачки в осыпи был встречен Pachydiscus (P.) neubergicus neubergicus (Hauer, 1858) (табл. III, фиг. 1). Среди биотурбаций в разрезе хорошо различимы только Zoophycos, мелкие Palaeophycus и Chondrites. Из данной пачки указываются находки Belemnitella mucronata minor Jel., Pachydiscus koeneni Gross. (Маслакова, Липник, 1971, с. 97), а также Baculites anceps Lam. и Hauericeras sulcatum (Kner) (Заклинская, Найдин, 1985, с. 81). Породы слагают сильно задернованный склон, верхняя часть подпачки задернована полностью. Мощность 9 м.
Подпачка ХХb. Светло-серые алевритистые биотурбированные мергели с единичными пятнами окремнения. В шлифах это фораминиферовые вакстоуны с фрагментами иглокожих, спикулами губок и неопределимыми биокластами (табл. I , фиг. В). Планктонные фораминиферы доминируют над бентосными (П/Б = 95–100%); часть из них окремнена. Количество алевритовой примеси кварца (~1–5%), присутствующей в шлифах, возрастает вверх по разрезу. Встречаются немногочисленные зерна глауконита алевритовой размерности и мелкий фитодетрит. Из биотурбаций в разрезе выделяются Zoophycos, Planolites, Chondrites, мелкие Palaeophycus, редкие пиритизированные Pilichnus (табл. II, фиг. Б) и единичные U-образные норы Arenicolites isp. (табл. II, фиг. Б). Примерно в 8 м выше подошвы присутствует линзовидное скопление многочисленных створок иноцерамов, пиритизированных губок, единичных мелких двустворок Nucula, Modiolus, Limatula, гастропод и неопределимых до вида Baculites. Многочисленные фрагменты губок продолжают встречаться и выше. По данным (Alekseev, Kopaevich, 1997), из основания этой пачки происходит Diplomoceras cylindraceum (Defr.), указанный в работах предшественников (Маслакова, Липник, 1971, с. 98) вместе с другими аммонитами из более широкого интервала. Пачка образует крутой задернованный склон с небольшими обрывами, экспонированными вдоль промоины. Мощность 17.5 м.
Подпачка ХХс. Серые алевритистые биотурбированные мергели с пятнами окремнения. Подпачка резко отличается от предыдущей появлением цементированных горизонтов твердого дна, к которым приурочены многочисленные норы Thalassinoides, частично окремнелые. Эти горизонты расположены нерегулярно, с разными промежутками, создавая ступенчатый рельеф на крутом склоне. В шлифах породы представлены алевритовыми биокластовыми пак-вакстоунами с биокластами планктонных и бентосных (секреционных и агглютинированных) фораминифер, иглокожих, спикул губок (табл. I , фиг. Г). Размер кварцевых (10–20%) и глауконитовых (1–3%) зерен увеличивается вверх по разрезу, достигая крупноалевритового. Помимо Thalassinoides среди биотурбаций определены Zoophycos и Planolites. Во всей подпачке содержится большое количество фрагментов губок, панцирей Echinocrys, мелких неопределимых Baculites. В 7 м выше подошвы встречен маркирующий прослой с достаточно многочисленными Belemnella gracilis (Arkhangelsky) (табл. III, фиг. 3–4), прослеживающийся и в других разрезах, а в 2 м ниже кровли встречены остатки Pachydiscus (P.) cf. armenicus Atabekian et Akopian, 1969 (табл. III, фиг. 2). Остатки крупной жилой камеры неопределимого до вида Pachydiscus отмечены нами и в 4 м выше подошвы. Из пачки указываются редкие находки Belemnella lanceolata (Schloth.), которые вверх по разрезу сменяются Belemnella sumensis sumensis Jeletzky и B. sumensis praearkhangelskii Naidin (Заклинская, Найдин, 1985, с. 81). Мощность 21 м.

Рис. 3.
Биотические события по диноцистам и результаты биостратиграфического расчленения разреза Бешкош. Условные обозначения см. рис. 1.
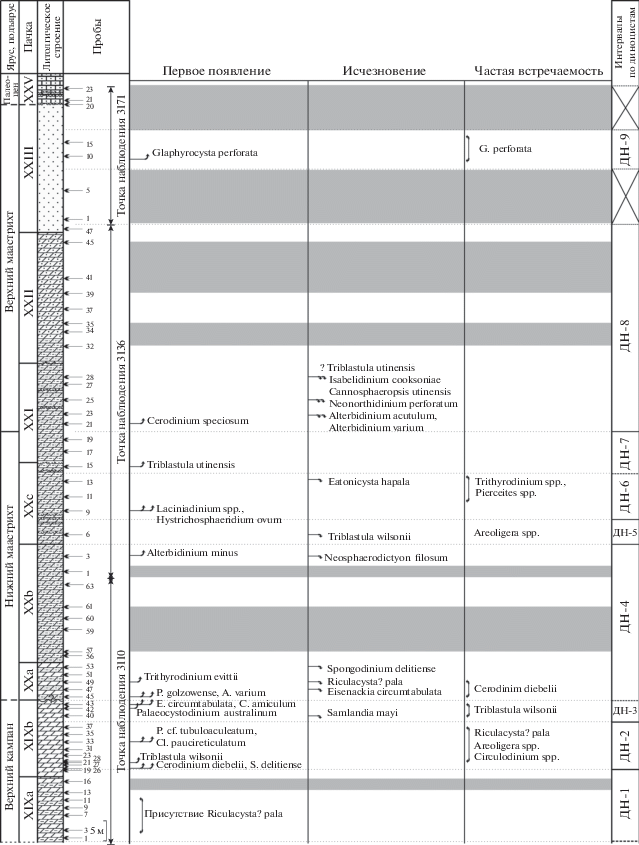
Рис. 4.
Количественное распределение в разрезе Бешкош экогрупп диноцист, основных групп палиноморф и реконструируемые палеообстановки формирования пород. Условные обозначения см. рис. 1.

Пачка XXI. Породы близки к подпачке XXc (табл. I , фиг. Д), но слагают хорошо выраженный вертикальный обрыв и крутую часть экспонированного склона над ним. Как и остальные породы, они полностью биотурбированы, но в ряде случаев отчетливо заметны ходы Zoophycos и Chondrites (табл. II, фиг. А). В толще нерегулярно прослеживаются горизонты выветрелых пиритовых конкреций и хардграунды, вблизи кровли образующие выступающие карнизы, пронизанные окремнелыми норами Thalassinoides (табл. II, фиг. Д). Из пачки указываются Belemnella sumensis, переходные формы между В. sumensis и Neobelemnella kazimiroviensis (Skolozd.), ядра Phyllopachyceras surya (Forb.), Hoploscaphites cf. constrictus (J. Sow.), Hauericeras sulcatum (Kner) и Acanthoscaphites ех gr. tridens (Kner) (Заклинская, Найдин, 1985, с. 81). Мощность 24.5–25 м.
Пачка XXII. Мергели желтовато-серые, биотурбированные, алевритовые, до тонкопесчаных вверху (табл. I , фиг. Е), с окремнением, приуроченным к норам Thalassinoides, развитым вдоль горизонтов твердого дна. Пачка очень плохо обнажена и образует крутой заросший склон. Мощность 30–31.5 м.
Пачка ХХIII. Глауконит-кварцевые биотурбированные известняки и известковые песчаники с многочисленными линзовидными и протяженными горизонтами цементации, в том числе твердого дна. Очень редко присутствуют участки разреза (мощностью в первые сантиметры) с реликтами бугорчатой и градационной слоистости. Породы содержат скопления раковин устриц и пектенид, количество которых заметно увеличивается вверх по разрезу. Обычно створки в них разноориентированные, но вблизи кровли пачки Mimachlamys и мелкие устрицы образуют сплошные протяженные прослои с раковинами, ориентированными выпуклостью вверх. Достаточно часто с ними ассоциируют крупные двустворчатые Pycnodonte (табл. II, фиг. З). В шлифах (табл. I , фиг. Ж) видно, что размер зерен кварца (30–40%) и глауконита (5–10%) увеличивается вверх до мелкопесчаного. Присутствуют биокласты планктонных и бентосных фораминифер (последние преобладают, П/Б = 30% и менее), иглокожих, двустворок, единичные спикулы. Среди биотурбаций доминируют Thalassinoides и Schaubcylindrichnus (табл. II, фиг. Е), реже присутствуют Skolithos, в основании встречаются единичные Zoophycos. Из пачки указываются Pachydiscus (P.) neubergicus (Hauer), Р. (P.) colligatus (Вinkhоrst), Hoploscaphites constrictus (Sow.) (Маслакова, Липник, 1971, с. 98), Neobelemnella kazimiroviensis (Skolozd.), Hoploscaphites cf. constrictus (Заклинская, Найдин, 1985, с. 83). Породы образуют почти вертикальный уступ с резко выраженной нишей в самой кровле пачки. Кровля представляет собой горизонт “зрелого” твердого дна, пронизанного норами Thalassinoides, местами фосфатизированными и окремненными, уходящими на глубину до 50–60 см, иногда заполненными битой ракушей (табл. II, фиг. Ж). Большинство нор выполнено зеленоватыми песчаниками вышележащего слоя, в них встречаются зерна и мелкая галька фосфоритов. Мощность 22 м.
Пачка XXV. Согласно (Алексеев, 1989), пачка XXIV в данном разрезе отсутствует, но, возможно, часть ее сохранилась от размыва (Яковишина и др., 2008). Светло-серые известняки, в основании (1.5 м) с прослоями зеленоватого мелко-среднезернистого глауконит-кварцевого и кварцево-глауконитового песчаника, содержащего несколько уровней твердого дна с норами Thalassinoides. В шлифах (табл. I , фиг. З), помимо зерен кварца и глауконита мелко- и среднепесчаной размерности, отмечаются биокласты бентосных фораминифер, зеленых водорослей, крупные фрагменты иглокожих, двустворок, мшанок и серпулид, фосфатные и карбонатные пелоиды (?пеллеты). Песчаная примесь быстро убывает вверх по разрезу. В нижних 0.5 м разреза указываются переотложенные ростры Neobelemnella kazimiroviensis (Skolozdr.), раковины Entolium sp., Pycnodonte mirabile (Rouss.) и другие двустворки (Заклинская, Найдин, 1985, с. 83).
УСЛОВИЯ ОСАДКОНАКОПЛЕНИЯ
Петрографическая характеристика
Микроскопическое строение пород еще раз подтверждает отчетливый тренд обмеления в течение кампана–маастрихта, выявленный ранее (Маслакова, Липник, 1971; Заклинская, Найдин, 1985; Алексеев, 1989; Alekseev, Kopaevich, 1997 и др.).
Состав мергелей пачки XIX свидетельствует об их формировании в условиях пелагического бассейна. Это полностью биотурбированные фораминиферовые вакстоуны с доминированием мелких глобигериноподобных форм. Согласно модели Э. Флюгеля (Flügel, 2010), породы отвечают стандартной микрофации 3: “пелагический мад-, вак-, пакстоун с планктонными микрофоссилиями”. Подобные отложения характеризуют бассейновые фации или глубокий шельф. Несмотря на постоянное присутствие остатков губок в разрезе, в шлифах спикулиты не встречены, хотя отдельные спикулы отмечаются. Единичные зерна угловатого кварца имеют мелкоалевритовый размер (0.04–0.06 мм) и, очевидно, являются следствием эолового разноса.
В основании подпачки XXа (обр. 3110/45) появляется, а с подошвы подпачки XXb (обр. 3110/54) постоянно присутствует алевритовая примесь кварца, размер которой увеличивается вверх по разрезу от 0.06–0.08 до 0.08–0.1 мм, а количество – с 1–5% в подошве пачки XX до 10–15% в пачке XXI. Горизонты твердого дна за счет роста карбонатного цемента содержат меньше алеврита. Одновременно появляется глауконит, размер зерен которого сопоставим с размером зерен кварца, а количество примерно в 3–4 раза меньше. Сами карбонаты при этом почти не меняются – несколько увеличивается количество биокластов бентосных организмов, появляются агглютинирующие фораминиферы.
В пачке XXII появляется кварцевая примесь мелкопесчаной размерности, а примерно с середины пачки ее количество составляет около 5% и появляются единичные зерна плагиоклазов и метаморфических пород. Размер и количество обломочных зерен и глауконита увеличиваются до 50% в подошве пачки XXIII и продолжают расти вверх по разрезу. Одновременно до первых процентов сокращается количество планктонных фораминифер и растет доля биокластов бентосных организмов.
В подошве палеоцена (пачка XXV) размер зерен кварца и глауконита увеличивается до среднепесчаного. При этом кварц остается плохо- и среднеокатанным, что свидетельствует о сравнительно недалеких источниках сноса.
Таким образом, тренд обмеления подтверждается изменением количества и размера обломочной составляющей. Об этом же говорит и характер изменения соотношения П/Б для фораминифер: от 100–95% в пачке XIX до 30% в пачке XXIII и до 0 на границе с пачкой XXV.
Стоит отметить, что оценки количества обломочной составляющей и отношение планктон/бентос для фораминифер в шлифах в настоящей работе отличаются от данных (Alekseev, Kopaevich, 1997, рис. 4). Для обломочной примеси это объясняется присутствием многочисленных раковин фораминифер и других биокластов, замещенных халцедоном и входящих в нерастворимый остаток, а также самим методом измерения. Различие отношений П/Б также можно объяснить методикой измерений, в которой подсчитывалось количество фораминифер всех размеров в нескольких полях зрения, а не число выделенных фораминифер определенной фракции.
Ихнологическая характеристика
Как уже упоминалось, все породы разреза полностью биотурбированы, реликты первичных текстур отмечены лишь в верхней части пачки XXIII. На фоне биотурбированных пород в нижней части разреза (пачка XIX–подпачки XXa, XXb) наиболее часто наблюдаются поперечные сечения Zoophycos isp. (табл. II, фиг. А). Ихнокомплекс дополняется Planolites isp., Palaeophycus isp., Chondrites isp. Редко присутствуют Pilichnus isp. Ихнокомплекс в целом обычен для ихнофации Zoophycos пелагических бассейнов (в том числе внешнего шельфа) (Knaust, Bromley, 2012) и характеризуется обилием червей и червеподобных организмов-грунтоедов. Светлая окраска пород, отсутствие значительной биотурбации Chondrites и Pilichnus свидетельствуют о нормальной аэрации осадка.
Вблизи подошвы подпачки XXc к этому ихнокомплексу добавляется еще один важный и многочисленный элемент – Thalassinoides, норы ракообразных, с которыми ассоциируется формирование хардграундов (табл. II, фиг. Д). Очень редко отмечаются вертикальные трубки Arenicolites (табл. II, фиг. Г), характерные в основном для мелководья. Ассоциация Zoophycos и Thalassinoides, с учетом привноса обломочной составляющей, отвечает ихнофации Cruziana открыто-морских мелководных бассейнов или внутреннего шельфа (Knaust, Bromley, 2012).
Начиная с подошвы пачки XXIII, а возможно, и с верхней части пачки XXII, в разрезе появляются норы Schaubcylindrichnus (табл. II, фиг. Г), также рассматриваемые обычно как норы полихет или кишечножаберных (Knaust, 2017). Их широкое развитие типично для проксимальной части ихнофации Cruziana или для ихнофации Skolithos и характеризует мелководные условия, чаще всего нижнюю предфронтальную зону (Knaust, 2017). Редкость нор Skolithos в этой части разреза предполагает принадлежность данного интервала к проксимальной части ихнофации Cruziana. Lepidenteron lewesiensis (табл. II, фиг. В), представляющие собой скопление остатков рыб и обычно интерпретируемые как копролиты, в настоящее время рассматриваются как выполнение нор хищных червей (Jurkowska, Uchman, 2013).
В кровле пачки XXIII, как уже отмечалось, сохранились реликты горизонтальной и бугорчатой слоистости, типичные для штормовых отложений – темпеститов. Здесь доминируют Thalassinoides, встречаются последние Schaubcylindrichnus, но сохранность отдельных нор весьма плохая на фоне широкого развития ракушняковых прослоев, количество и протяженность которых увеличиваются к кровле (табл. II, фиг. З). Все это вместе указывает на формирование пачек XXIII и XXV в условиях интенсивного волнового воздействия нижней предфронтальной зоны.
Суммируя сказанное, изученный разрез можно интерпретировать следующим образом. Седиментация происходила в условиях постепенного обмеления бассейна. На начальном этапе, отвечающем пачке XIX–подпачкам XXa, XXb, здесь существовали условия, сопоставимые с внешним шельфом. В интервале, отвечающем подпачке XXc–пачке XXII, происходило медленное осадконакопление в условиях, сопоставимых с внутренним шельфом и переходной зоной пляжа (глубины порядка 50 м). Оно характеризовалось постепенным увеличением привноса обломочного материала и сопровождалось перерывами, во время которых происходило формирование хардграундов. В интервале, соответствующем пачке XXIII, продолжалось обмеление бассейна до глубин нижней предфронтальной зоны (15–20 м), подверженной периодическому штормовому воздействию. Это, видимо, явилось причиной сокращения и исчезновения диноцистовых комплексов (см. ниже). За этим последовал продолжительный перерыв, во время которого формировались глаукониты и фосфориты. Судя по находкам костей сухопутных динозавров из основания датских известняков (Рябинин, 1946) в разрезе Бешкош и в соседних разрезах (Лопатин, Аверьянов, 2019), можно допустить осушение бассейна в конце маастрихтского века. Водную массу морского бассейна можно охарактеризовать как умеренно теплую, типичную для бассейнов “Европейской палеогеографической области”. Результаты изотопных анализов (см. ниже) позволяют оценить температуру воды ~18–19°С. Полученные данные частично подтверждают выводы, сделанные ранее (Яковишина и др., 2008; Габдуллин и др., 2015).
БИОСТРАТИГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА
Головоногие моллюски
Несмотря на то, что из разреза Бешкош в литературе указывается достаточно большое количество находок головоногих моллюсков, судить о достоверности определений не представляется возможным, поскольку их фотографии отсутствуют. Нам удалось найти только изображения ростров Neobelemnella kazimiroviensis (Skolozdrowna, 1932), предположительно, из пачки XXIII в работе (Найдин, 1975, табл. XV , фиг. 4–5), определение которых не вызывает больших возражений (Keutgen et al., 2017, p. 21). Это типично верхнемаастрихтские формы.
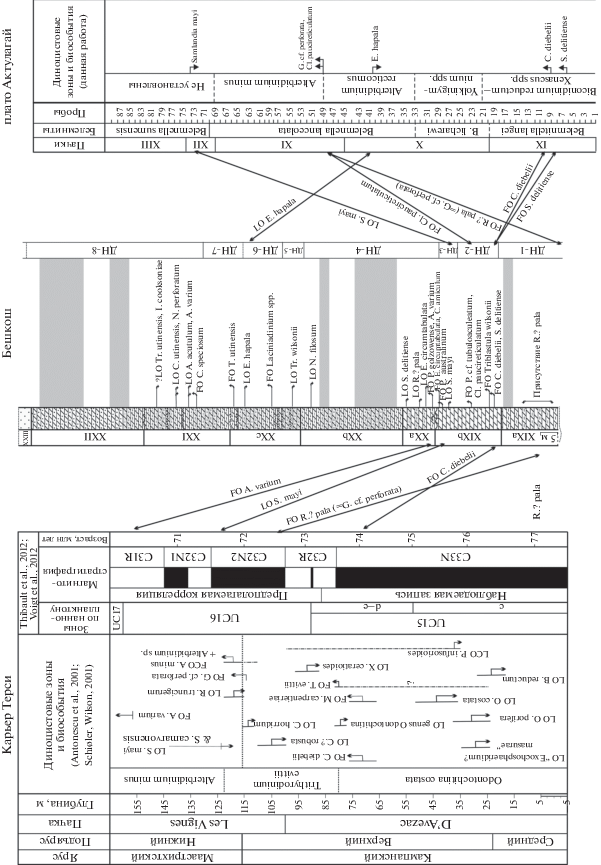
Рис. 5.
Корреляция биособытий по диноцистам разрезов карьера Терси, Бешкош и плато Актолагай. Условные обозначения см. рис. 1. Сокращения: LO – последнее присутствие, FO – первое присутствие, FCO – первая частая встречаемость.
Находка Phyllopachyceras surya (Forbes), изображенная в (Найдин, 1959, с. 176, табл. 15 , фиг. 3), упоминающаяся в (Заклинская, Найдин, 1985, с. 81) и хранящаяся в Музее землеведения МГУ (обр. № 1/17), происходит из Белогорского района, с. Курское. Находки других аммонитов и белемнитов из окрестностей г. Бахчисарай, изображенные в (Найдин, 1959; Йолкичев, Найдин, 1999), не имеют, к сожалению, привязки к конкретным разрезам и слоям. Поэтому мы не беремся их интерпретировать применительно к рассматриваемому разрезу. В то же время известно, что большинство палеонтологов подтверждают определения аммонитов, опубликованных в (Найдин, 1959). Поэтому можно допустить высокую степень достоверности цитируемых находок, перечисленных ниже.
Pachydiscus (P.) koeneni De Grossouvre в настоящее время рассматривается либо как самостоятельный вид (Wiedmann, Schmidt, 1993), либо как младший синоним Pachydiscus (P.) haldemsis (Schlueter) (Kennedy, Summesberger, 1984). В любом случае он указывает на кампанский возраст пачки XIX и подпачки XXa.
Pachydiscus (P.) neubergicus (Hauer) указан из пачки XXIII (Маслакова, Липник, 1971). Данный вид является индексом подошвы маастрихта и распространен до нижней части верхнего маастрихта (Kennedy, Summesberger, 1986; Niebuhr, 2003; Jagt, Felder, 2003).
В пачке XXIII встречается и Pachydiscus (P.) colligatus (Вinkhоrst). Это, пожалуй, единственный аммонит из разреза Бешкош, с определением которого не согласен У. Кеннеди (Kennedy, 1987). Данный аммонит был переопределен как раннемаастрихтский Pachydiscus (P.) epiplectus (Redtenbacher) (Kennedy, Summesberger, 1986).
Hauericeras sulcatum (Kner, 1848) является раннемаастрихтским видом (Kennedy, Summesberger, 1987), поэтому его присутствие в подпачке XXa и пачке XXI указывает на принадлежность к маастрихту.
Baculites anceps Lamarck обычен для верхнего маастрихта, хотя появляется в нижнем маастрихте (Klinger, Kennedy, 2001), что говорит о возможности проведения границы кампанского и маастрихтского ярусов в интервале пачки XIX–подпачки XXa.
Diplomoceras cylindraceum (Defrance) распространен в верхах кампана–маастрихте (Klinger, Kennedy, 2003), в том числе и в лимитотипе (Odin, Lamaurelle, 2001). Сообщения о раннекампанских находках D. cylindraceum нуждаются в подтверждении. Согласно (Alekseev, Kopaevich, 1997), в данном разрезе этот вид происходит из подпачки XXb.
Hoploscaphites constrictus (J. Sowerby) и H. cf. constrictus указаны из пачек XXI и XXIII. Вид характеризует маастрихтский ярус (Kennedy, Summesberger, 1987; Niebuhr, 2003), появляясь, возможно, в самой кровле кампана (Odin, Lamaurelle, 2001).
Наконец, вид Acanthoscaphites (A.) gr. tridens (Kner) также указан из пачки XXI. Он характерен для нижнего маастрихта (Kennedy, Summesberger, 1987; Niebuhr, 2003).
Таким образом, на основе цитированных находок аммонитов пачки XXI–XXIII несомненно имеют маастрихтский возраст, а граница кампанского и маастрихтского ярусов предположительно может располагаться в подпачке XXa.
С белемнитами дело обстоит сложнее, поскольку, судя по спискам определений, они несколько раз пересматривались Д.П. Найдиным. Поэтому мы остановимся только на наиболее поздних определениях, приведенных в (Заклинская, Найдин, 1985) и воспроизведенных в (Alekseev, Kopaevich, 1997).
Belemnitella mucronata senior Now., отмеченная в пачке XIX (Маслакова, Липник, 1971), является верхнекампанским видом.
Из подпачки XXc указываются Belemnella lanceolata (Schlotheim), Belemnella sumensis sumensis Jeletzky и B. sumensis praearkhangelskii Naidin (Заклинская, Найдин, 1985). Появление Belemnella lanceolata считается дополнительным маркером границы между кампаном и маастрихтом в “безаммонитовых” разрезах Западной Европы (Odin, Lamaurelle, 2001). Данная концепция сохраняется и по сей день (Remin, 2018). В то же время для востока Русской плиты и Прикаспия нами было показано, что эта граница проходит не в подошве, а внутри зоны Belemnella lanceolata (Беньямовский и др., 2016; Baraboshkin et al., 2017).
Оба подвида Belemnella sumensis характеризуют нижний маастрихт Восточной Европы (Alekseev et al., 1999) и восточной части Западной Европы, где в последнее время они рассматриваются в качестве самостоятельных видов (Remin, 2018).
Из пачки XXI указываются Belemnella sumensis, переходные формы между В. sumensis и Neobelemnella kazimiroviensis (Skolozd.). Последний вид присутствует в пачке XXIII, переотложен в подошве пачки XXV и, как отмечено выше, характеризует верхний маастрихт (Найдин, 1975; Alekseev et al., 1999; Keutgen et al., 2017).
Таким образом, на основе цитированных находок белемнитов можно считать, что граница кампанского и маастрихтского ярусов должна проводиться ниже подпачки XXc, а граница нижнего и верхнего маастрихта – в интервале пачек XXI–XXIII.
Немногочисленные остатки головоногих моллюсков, имеющиеся в нашем распоряжении, позволяют сделать следующие выводы.
Ядро фрагмокона Pachydiscus (P.) neubergicus neubergicus (Hauer) встречено в осыпи в нижней части подпачки XXa (табл. III, фиг. 1). Он может характеризовать только подпачку XXa, о чем свидетельствует выполняющая его порода, не содержащая алевритовой примеси. Хотя аммонит слабо деформирован (сплюснут с боков), его диагностические признаки близки к лектотипу (см. Kennedy, Summesberger, 1986). Раковина полуэволютная; диаметр ядра 110 мм; на половине оборота присутствует 8 слабоизогнутых главных ребер, утолщенных на умбиликальном перегибе; им соответствует 26 вторичных ребер, сглаживающихся в месте ветвления, чуть выше середины боков. Вторичные ребра почти радиальны, немного наклонены вперед.
Данный вид является биостратиграфическим маркером подошвы маастрихта (Odin, Lamaurelle, 2001), хотя в различных разрезах появляется на разных уровнях и имеет значительную диахронность (Jagt, Felder, 2003).
Таким образом, подошва маастрихта должна находиться ниже кровли подпачки XXa.
Pachydiscus cf. (P.) armenicus Atabekian et Akopian, 1969 из подпачки XXс, несмотря на фрагментарную сохранность, определен достаточно уверенно. Это сильно деформированный (сплющенный) обломок полуэволютного фрагмокона высотой 30 мм. Главные ребра начинаются на умбиликальном перегибе, где они утолщены. Чуть ниже середины боков они делятся на 3 ветви, слегка понижаясь в месте ветвления. Cлабо изгибаясь вперед, они без понижения пересекают вентральную сторону. Данный вид известен из маастрихта и, вероятно, верхней части кампана, наиболее часто встречаясь в верхнем маастрихте (Machalski, 2012). Поэтому данная находка не помогает уточнить положения ярусных и подъярусных границ.
В подпачке XXс нами встречен уровень с белемнитами Belemnella gracilis (Arkh.) (табл. III, фиг. 3–4), который, вслед за (Schulz, 1979; Christensen, 1997; Remin, 2018), мы рассматриваем как самостоятельный вид, а не подвид. Это крупные (до 131 мм), почти гладкие субцилиндрические ростры с большим удлинением (10) и нулевым индексом Шатского, что отличает их от близких Belemnella praearkhangelskii Naidin. Вероятно, подобные ростры и определялись Д.П. Найдиным как Belemnella sumensis praearkhangelskii Naidin (Заклинская, Найдин, 1985). По существующим данным эта форма характеризует нижний маастрихт, а согласно (Christensen, 1997), она распространена вместе с B. praearkhangelskii Naidin в средней–верхней частях нижнего маастрихта.
Суммируя изложенный материал по находкам головоногих моллюсков, можно заключить, что граница кампанского и маастрихтского ярусов должна находиться ниже кровли подпачки XXa; граница подъярусов маастрихта – в интервале пачек XXI–XXIII. Граница мела и палеогена в разрезе хорошо известна и проводится по горизонту твердого дна в подошве пачки XXV (Маслакова, Липник, 1971; Заклинская, Найдин, 1985; Алексеев, 1989; Alekseev, Kopaevich, 1997).
ПАЛИНОЛОГИЧЕСКИЙ АНАЛИЗ
Методы и материал исследований
Мацерация проб проводилась по методике, принятой в лаборатории палеофлористики Геологического института РАН и являющейся модификацией сепарационного метода (Гричук, 1940). Последовательность химической обработки следующая: 1) обработка 10%-ным раствором HCl для удаления карбонатов; 2) отмывка проб дистиллированной водой до нейтральной реакции (каждые 2 ч); 3) обработка проб горячим 10%-ным раствором Na2HPO4; 4) отмучивание пробы от глинистых частиц дистиллированной водой (каждые 2 ч); 5) центрифугирование полученного осадка в растворе тяжелой жидкости [K2(CdI4)] с удельным весом 2.25 для извлечения ископаемых палиноморф; 6) отмывка органического мацерата от тяжелой жидкости дистиллированной водой и его сбор в пробирки; 7) обработка полученного мацерата концентрированной HF; 8) отмывка мацерата в центрифуге от плавиковой кислоты дистиллированной водой до нейтральной PH. Полученный мацерат заливается глицерином и хранится в пробирках.
Изучение препаратов производилось в оптическом микроскопе Axiostar plus (Carl Zeiss), при рабочем увеличении ×400 и ×600. Определялись все имеющиеся в пробе виды палиноморф (диноцисты, акритархи, празинофиты, споры и пыльца растений). Минимально подсчитывалось 200 экземпляров палиноморф на одну пробу, после чего препарат просматривался до конца с целью обнаружения редко встречаемых таксонов. В случае малого количества палиноморф подсчитывались все встреченные экземпляры. Фотографии выполнены на микроскопе Axiostar plus (Carl Zeiss) с фотонасадкой на базе фотоаппарата Canon PC1200 и программы визуализации AxioVision Rel. 4.7. (Carl Zeiss).
При подсчете в палинологических спектрах за 100% принималась сумма всех палиноморф. Выделение палинологических ассоциаций было основано (1) на выявлении последовательности стратиграфически важных уровней первого появления, исчезновения, частой встречаемости видов диноцист; (2) на количественных соотношениях различных групп палиноморф (диноцист, акритарх, празинофитов) и уровней их изменений; (3) на изменениях таксономического состава и количественных соотношений различных экологических групп диноцист.
Материалом для палинологического исследования послужили 60 проб из разреза Бешкош, взятых с соответствующих уровней палеомагнитных проб в точках 3110, 3136 и 3171 (рис. 3).
Установлено, что палиноморфы распределены по разрезу неравномерно. Они отсутствуют или встречены в незначительном количестве (менее 15 экз.) в точке наблюдения (т.н.) 3110 – пробы 3, 13–16, 56–61, т.н. 3136 – пробы 1, 32–35, 39–45 и т.н. 3171 – пробы 1, 5, 20–21, 23. Остальные пробы содержат многочисленные диноцисты, акритархи, празинофиты, при незначительном участии спор и пыльцы растений, что указывает на формирование отложений в открыто-морских, удаленных от берега палеообстановках.
Стратиграфическое распределение палиноморф приведено в Приложении 1 ; количественное распределение различных групп палиноморф и экогрупп диноцист – на рис. 3, 4. Изображения характерных таксонов помещены на фототаблицах IV–VI.
Расчленение по палинологическим данным
Приведенные Е.Д. Заклинской и Д.П. Найдиным (1985) палинологические данные по разрезу Бешкош получены из одной пробы, взятой в основании зоны Neobelemnella kazimiroviensis. Этими авторами было показано преобладание в спектре пыльцы покрытосеменных стемм Normapolles Pflug и Postnormapolles Pflug, с участием ключевых таксонов – Ulmodeipites spp., Myricites krempii Pflug, Nudopollis terminalis (Pflug et Thomson) Pflug и других. Данный спектр был соотнесен с верхней частью фазы “с” второго этапа кайнофита (верхний маастрихт) Европейско-Туранской области палеофлористического царства “Norma” (Zaklinskaya, 1981). Также были приведены сведения о наличии в пробе диноцист (как планктонных водорослей) Hafnisphaera fluens Hansen., Palynodinium grallator Gocht, находки которых дали основание отнести изученный образец к верхнемаастрихтской зоне Palynodinium grallator схемы (Hansen, 1977). Однако приведенные в работе (Заклинская, Найдин, 1985) фотографии P. grallator не подтверждают сделанный данными авторами вывод, поскольку на них изображены другие таксоны.
Выявленные диноцистовые события позволили выделить 9 диноцистовых интервалов, установленных по присутствию или появлению руководящих таксонов и/или по количественным характеристикам ассоциаций палиноморф.
Диноцистовый интервал 1 (ДН-1) (подпачка XIXа, т.н. 3110, пробы 1–19). Комплекс диноцист чрезвычайно бедный в количественном и видовом отношении, таксоны встречаются в виде редких (чаще единичных) экземпляров плохой сохранности (Приложение 1 ). В этом интервале обнаружены Circulodinium distinctum (Deflandre et Cookson) Jansonius, Riculacysta? pala Kirsch, cf. Cassidium fragile (Harris) Drugg, Exochosphaeridium sp., Ex. bifidum (Clarke et Verdier) Clarke et al., Exochosphaeridium cf. mullieri Yun Hyesu, Cladopyxidium saeptum (Morgenroth) Stover et Evitt, Palaeostomocystis reticulata Deflandre, Hystrichosphaeridium sp., Xenikoon australis Cookson et Eisenack, Membranigonyaulax wilsonii Slimani, Areoligera sp., Lanternosphaeridium lanosum Morgenroth, Leberidocysta chlamydata (Cookson et Eisenack) Stover et Evitt, Leberidocysta? microverrucosa Slimani – Leb.? flagellichnia Schiøler.
Диноцистовый интервал 2 (ДН-2) (нижняя–средняя части подпачки XIXb, т.н. 3110, пробы 21–37). Комплекс диноцист сохраняет в своем составе ранее встречавшиеся таксоны и обогащается за счет появления новых таксонов.
В этом интервале отмечено появление C. diebelii (Alb.) Lent. et Will., Spongodinium delitiense (Ehrenberg) Deflandre, Triblastula wilsonii Slimani, Caligodinium amiculum Drugg, Conneximura fimbriata (Morgenroth) May, Cladopyxidium paucireticulatum Slimani, Cribroperidinium ventriosum (Wetzel) Lentin et Williams, Eatonicysta hapala Schiøler et Wilson, Hystrichosphaeridium proprium Marheinecke, Neosphaerodictyon filosum Slimani, Pervosphaeridium cf. tubuloaculeatum Slimani, Isabelidinium sp., Areoligera spp., Phanerodinium sp., P. sonciniae Marheinecke и др. (Приложение 1 , рис. 3, 4). Только на этом уровне присутствуют Areoligera tenuicapillata (Wetzel, ex Deflandre) Lejeune-Carpentier, Glaphyrocysta expansa (Corradini) Roncaglia et Corradini. В кровле интервала отмечено последнее присутствие Circulodinium distinctum.
В ассоциации палиноморф преобладают представители диноцист семейства Areoligeraceae (Riculacysta? pala, Areoligera spp., Circulodinium distinctum) и таксоны неясного систематического положения, предположительно акритархи Palaeostomocystis reticulata, Fromea chytra, Fromea cf. laevigata. Относительно часты диноцисты Hystrichosphaeridium spp., Cribroperidinium, Exochosphaeridium spp., Coronifera sp., Xenikoon australis; остальные таксоны немногочисленны (рис. 4).
Диноцистовый интервал 3 (ДН-3) (терминальная часть подпачки XIXb, т.н. 3110, пробы 40–43). На этом стратиграфическом уровне отмечено появление диноцист Palaeocystodinium australinum (Cookson) Lent. et Will., Eisenackia circumtabulata Drugg, акритарх Paralecaniella indentata (Deflandre et Cookson) Cookson et Eisenack и последняя постоянная встречаемость диноцист Riculacysta? pala.
Ассоциация микрофитопланктона характеризуется преобладанием диноцист Triblastula wilsonii и Hystrichosphaeridium spp., сокращением количества Riculacysta? pala, Areoligera spp., Cladopyxidium saeptum, акритарх Fromea spp. и увеличением – Spiniferites spp., Membranigonyaulax wilsonii; постоянным присутствием Spongodinium delitiense (Приложение 1 , рис. 3, 4).
Диноцистовый интервал 4 (ДН-4) (подпачки ХХa–XXb, т.н. 3110, пробы 45–63, т.н. 3136, проба 3). Наиболее полная характеристика получена для подпачки ХХа, выше палиноморфы встречаются дискретно (Приложение 1 , рис. 3). В основании интервала ДН-4 комплекс диноцист значительно обогащается (Приложение 1 ), что предполагает наличие перерыва в разрезе. В интервале впервые встречаются: Cerodinium sp., C. albertii (Corradini) Lent. et Will., C. leptodermum (Vozzhennikova) Lent. et Will., Alterbidinium sp., A. acutulum (Wilson) Lent. et Will., A. varium Kirsch, Isabelidinium cooksoniae (Alb.) Lent. et Will., Neonorthidium perforatum Marheinecke, Palaeocystodinium golzowense Alberti, Turnhosphaera hypoflata (Yun Hyesu) Slimani, Fibradinium annetorpense Morgenroth, Membranosphaera maastrichtica Samoilovitch, Microdinium bensonii Slimani, M. carinatum (Below) Lent. et Will., Pterodinium cingulatum (Wetzel) Below, Cannosphaeropsis utinensis Wetzel, Gillinia hymenophora Cookson et Eisenack, Kallosphaeridium? cf. granulatum (Norvick) Stover et Evitt, Rhiptocorys veligera (Deflandre) Lejeune-Carpentier et Sarjeant, Florentinia ferox (Deflandre) Duxbury, Senegalinium obscurum (Drugg) Stover et Evitt, Spinidinium cf. eggerii Kirsch, Trigonopyxidia ginella (Cookson et Eisenack) Downie et Sarjeant, а также акритархи Fromea fragilis (Cookson et Eisenack) Stover et Evitt и др.
В интервале ДН-4 отмечается последнее присутствие диноцист Spongodinium delitiense, Riculacysta? pala, Neosphaerodictyon filosum, Eisenackia circumtabulata, Caligodinium amiculum, Leberidocysta? microverrucosa – Leb.? flagellichnia, Lanternosphaeridium lanosum.
В интервале ДН-4 выявлено существенное количество (не менее 15%) перидиниоидных таксонов (Cerodinium spp., Alterbidinium spp., Isabelidinium spp., Senegalinium obscurum). Также обильны Spiniferites, Pterodinium cingulatum, Microdinium spp., Exochosphaeridium cf. mullieri, Membranosphaera maastrichtica и акритархи Palaeostomocystis reticulata. В комплексе палиноморф относительно часты (в среднем 10%) споры и пыльца растений, постоянно встречаются колониальные зеленые водоросли Palambages morulosa, в нижней части интервала – органические оболочки фораминифер (рис. 4).
Диноцистовый интервал 5 (ДН-5) (основание подпачки ХХс, т.н. 3136, проба 6). В ассоциации диноцист значительно сокращается таксономическое разнообразие, доминируют хоратные таксоны Areoligera spp., Exochosphaeridium spp., часты Triblastula wilsonii, Hystrichosphaeridium spp., остальные таксоны малочисленны (Приложение 1 ). На этом уровне установлено последнее присутствие Triblastula wilsonii, последняя частая встречаемость Exochosphaeridium cf. mullieri.
Диноцистовый интервал 6 (ДН-6) (средняя часть подпачки ХХс, т.н. 3136, пробы 9–13). Комплекс диноцист значительно обогащается, что предполагает наличие перерыва в разрезе (Приложение 1 ). В основании этого интервала (проба 9) зафиксировано присутствие различных видов Laciniadinium, первое появление Tanyosphaeridium variecalamum Davey et Williams. В кровле интервала исчезает Eatonicysta hapala.
Интервал характеризуется постепенным увеличением количества диноцист (снизу вверх по разрезу) Spiniferites spp., Membranigonyaulax wilsonii, Phanerodinium spp., Microdinium spp., Membranosphaera maastrichtica. В значительном количестве встречаются Xenikoon australis и акритархи Fromea spp., Paralecaniella indentata, cf. Prolixosphaeridium? nanus (Wetzel ex Lentin et Williams) Sarjeant. В основании интервала встречены частые акантоморфные акритархи (Micrhystridium sp.), выше – обилие перидиниоидных таксонов диноцист (Trithyrodinium spp., Pierceites spp.) (рис. 4).
Диноцистовый интервал 7 (ДН-7) (терминальная часть подпачки ХХс–нижняя часть пачки XXI, т.н. 3136, пробы 15–19). Диноцисты неразнообразны и в большей части интервала малочисленны (Приложение 1 ).
В основании интервала фиксируется первое появление Triblastula utinensis Wetzel, Rigaudella apenninica (Corradini) Below, а в целом доминирование Spiniferites spp. и водорослей Palambages morulosa.
Диноцистовый интервал 8 (ДН-8) (средняя часть пачки XXI–пачка XXII, т.н. 3136, пробы 21–47). Наиболее полная характеристика получена для средней части пачки XXI (Приложение 1 , рис. 3).
Интервал характеризуется первым появлением Cerodinium speciosum (Alberti) Lentin et Williams, последним присутствием Alterbidinium acutulum, Isabelidinium cooksonae, Cannosphaeropsis utinensis, Triblastula utinensis. Преобладают Spiniferites spp., Cerodinium spp., Phanerodinium sp., Microdinium spp., Membranosphaera maastrichtica, Xenikoon australis, cf. Prolixosphaeridium? nanus, акритархи Paralecaniella indentata, Fromea spp.
Диноцистовый интервал 9 (ДН-9) (средняя часть пачки XXIII, т.н. 3171, пробы 10, 15). Большая часть пачки XXIII и изученная часть пачки XXV не содержат палиноморф (Приложение 1 , рис. 3).
Интервал характеризуется первым появлением и массовой встречаемостью диноцист Glaphyrocysta perforata, при единичной встречаемости Areoligera senonensis, cf. Cassidium fragile.
Возраст по палинологическим данным
В результате анализа палинологических данных в разрезе Бешкош выявлены уровни появления, исчезновения, массовой встречаемости стратиграфически значимых таксонов диноцист (рис. 3). Основываясь на присутствии в разрезе ряда зональных и руководящих видов, представляется возможным сопоставить полученные палинологические данные с имеющимися зональными схемами по диноцистам кампана–маастрихта Западной Европы (Hulthberg, Malmgren, 1987; Kirsch, 1991; Marheinecke, 1992; Schiøler, Wilson, 1993; Roncaglia, Corradini, 1997a; Slimani, 2000, 2001), Израиля (Hoek et al., 1996), Западной Сибири (Lebedeva, 2006; Лебедева и др., 2013; Лебедева, Кузьмина, 2018), Тургайского прогиба (Васильева, Левина, 2007), Поволжья (Александрова и др., 2012).
В видимом основании разреза Бешкош (в подпачке XIXа), охарактеризованном ДН-1, установлено низкое таксономическое разнообразие диноцист, представленных преимущественно таксонами широкого стратиграфического распространения, за исключением Riculacysta? pala. В стратотипе нижней границы маастрихта появление Riculacysta? pala (=Glaphyrocysta cf. perforatа) выявлено в терминальном кампане, на уровне 114.8 м, что близко к границе кампана и маастрихта, установленной на уровне 115.2 м (Antonescu et al., 2001; Schiøler, Wilson, 2001) (рис. 5). Первое появление Riculacysta? pala на юге Германии (Гельветские и Ультрагельветские покровы Альп, Верхняя Бавария) (Kirsch, 1991) зафиксировано в основании зоны Cerodinium diebelii раннего маастрихта (рис. 6). В разрезах Бельгии (Slimani, 2001) это событие указывается на более высоком стратиграфическом уровне – в подзоне Cladopyxidium paucireticulatum средней части раннего маастрихта (рис. 7). В разрезе плато Актолагай (Казахстан) это событие установлено на уровне средней части белемнитовой зоны Belemnella lanceolata в слоях с Alterbidinium minus по диноцистам, отвечающих переходному этапу от кампана к маастрихту (неопубликованные данные Г.Н. Александровой) (рис. 5).

Рис. 7.
Корреляция биособытий по диноцистам в Израиле, разрезе Бешкош и Бельгии. Условные обозначения см. рис. 1. Сокращения: CO – частая встречаемость, FO Av – FO A. varium; FO Clp – FO C. paucireticulatum; FO Cp – FO C. paucireticulatum; FO Te – FO T. evittii; FO Cd – FO C. diebelii; FO Cs – FO C. speciosum; FO Pa – FO P. australinum; FO Pg – FO P. golzowense; LO Aa – LO A. acutulum; LO Ic – LO I. cooksaniae; Форам. – фораминиферы; Нанн. – наннопланктон; ниж. – нижний.

В нижней и средней частях подпачки XIXb, соответствующих интервалу ДН-2, выявлены уровни появления зональных и характерных видов диноцист – Cerodinium diebelii, Spongodinium delitiense, Cladopyxidium paucireticulatum, Pervosphaeridium cf. tubuloaculeatum, Glaphyrocysta expansa, Triblastula wilsonii, а также массовая встречаемость представителей семейства Areoligeraceae (Areoligera spp., R.? pala, Circulodinium spp.).
Наиболее ранние находки Spongodinium delitiense указываются в раннем кампане Норвежского моря (Radmacher et al., 2015), Бельгии (Slimani, 2000, 2001) (рис. 7), в позднем кампане Атлантического побережья США (Aurisano, 1989). На плато Актолагай (Казахстан) это событие установлено в кампане на уровне зоны Belemnitella langei (неопубликованные данные Г.Н. Александровой) (рис. 5).
В разрезах Гельветских и Ультрагельветских покровов Альп (Kirsch, 1991) появление S. delitiense, C. diebelii, присутствие R.? pala, наблюдаемые в интервале ДН-2, установлены в основании зоны Cerodinium diebelii (рис. 6). Нижняя граница зоны C. diebelii отвечает основанию нижнего маастрихта (Kirsch, 1991) и приурочена к подошве зоны Globotruncana falsostuarti по планктонным фораминиферам (по шкале Robaszynski et al., 1984).
В разрезе плато Актолагай (Казахстан) появление Cladopyxidium paucireticulatum установлено в слоях с Alterbidinium minus, отвечающих переходному этапу от кампана к маастрихту (Baraboshkin et al., 2017) (рис. 5).
В разрезах Италии Glaphyrocysta expansa имеет узкий стратиграфический диапазон – пограничные отложения нижнего/верхнего маастрихта, и ее первое появление фиксирует нижнюю границу одноименной подзоны (Roncaglia, Corradini, 1997a).
В разрезах Бельгии в течение всего раннего маастрихта отмечается последовательное появление видов Cladopyxidium paucireticulatum, Pervosphaeridium tubuloaculeatum, Triblastula wilsonii (Slimani, 2000, 2001) (рис. 7). В разрезе Бешкош не установлены различные уровни появлений C. paucireticulatum, P. tubuloaculeatum, а выявлена их совместная встречаемость в основании интервала ДН-2, отмечаемая в разрезах Бельгии с основания зоны Pervosphaeridium tubuloaculeatum второй половины раннего маастрихта. Стратиграфический диапазон распространения Triblastula wilsonii в Бельгии охватывает конец раннего–начало позднего маастрихта (Slimani, 2001).
Первое появление Cerodinium diebelii установлено в верхнем кампане в стратотипе нижней границы маастрихта (Antonescu et al., 2001; Schiøler, Wilson, 2001), в разрезах Бельгии (Slimani, 2000, 2001), Марокко (Slimani et al., 2016), а также плато Актолагай (Казахстан) (неопубликованные данные Г.Н. Александровой) (рис. 5, 7), а его постоянное присутствие характерно для нижнего маастрихта. В разрезах юга Германии (Kirsch, 1991), Северного моря (Schiøler, Wilson, 1993), Италии (Roncaglia, Corradini, 1997a), Израиля (Hoek et al., 1996) (рис. 6, 7), Западной Сибири (Lebedeva, 2006; Лебедева и др., 2013) (рис. 8), Испании (Radmacher et al., 2014) данный таксон впервые появляется в нижнем маастрихте, в Тургайском прогибе – в верхнем маастрихте (Васильева, Левина, 2007). Следует отметить, что C. diebelii в разрезе Бешкош имеет разорванный диапазон распространения, его единичная встречаемость отмечена в основании интервала ДН-2 (проба 26, т.н. 3110), а постоянная частая встречаемость наблюдается выше – с уровня ДН-4.

Ассоциация диноцист с массовой встречаемостью Areoligeraceae ранее была выявлена в пограничном кампан-маастрихтском интервале в Бельгии и Нидерландах (Robaszynski et al., 1985), Израиле (Hoek et al., 1996), Нижнем Поволжье (Александрова и др., 2012). В Израиле это биособытие установлено в начале раннего маастрихта на уровне верхней части наннопланктонной зоны Q. trifidum, в низах зоны Globotruncana falsostuarti по фораминиферам (рис. 7). В Поволжье массовое развитие представителей семейства Areoligeraceae в пограничных кампан-маастрихтских отложениях зафиксировано на уровне слоев с Senoniasphaera microreticulata, отвечающих зонам Neoflabellina praereticulata/Neoflabellina reticulata (LC19)–Falsoplanulina complanata (LC20) по бентосным фораминиферам, верхней части слоев с Prunobrachium articulatum и слоям c Orbiculiforma renillaeformis по радиоляриям (Александрова и др., 2012).
В терминальной части подпачки XXb, соответствующей интервалу ДН-3, выявлены уровни появления/исчезновения диноцист Samlandia mayi, Alterbidinium varium, Palaeocystodinium australinum, Eisenackia circumtabulata, Caligodinium amiculum, используемых для зонального расчленения верхнемеловых пород различных регионов.
В стратотипе нижней границы маастрихта в разрезе Терси исчезновение Samlandia mayi установлено в базальной части нижнего маастрихта (Antonescu et al., 2001; Schiøler, Wilson, 2001) (рис. 5), а ее стратиграфическое распространение в разрезах Бельгии ограничено верхним кампаном–нижней частью нижнего маастрихта (Slimani, 2000, 2001) (рис. 7). В нижнем маастрихте последнее присутствие S. mayi ранее было установлено в Поволжье на уровне нижней части слоев с Senoniasphaera microreticulata (Александрова и др., 2012), в разрезе плато Актолагай в нижней части зоны Belemnella sumensis, а также на юге Западной Сибири стратиграфически ниже зоны CC24/UC18 по наннопланктону – в нижней части слоев с Cerodinium diebelii (Лебедева и др., 2013) (рис. 5, 8).
Следует отметить, что в ДН-3 и в нижней части ДН-4 наблюдаются частые находки S. delitiense, что в разрезах Бельгии характеризует зону Membranilarnacia liradiscoides нижнего маастрихта (рис. 7).
Первое появление видов рода Palaeocystodinium приурочено к верхнему кампану в Израиле (Hoek et al., 1996), Тургайском прогибе (Васильева, Левина, 2007), Датском бассейне (Surlyk et al., 2013), к основанию маастрихта на юге Германии (Kirsch, 1991), в Испании (Radmacher et al., 2014), к нижнему маастрихту в Италии (Roncaglia, Corradini, 1997a), Бельгии (Slimani, 2000), Западной Сибири (Lebedeva, 2006) (рис. 6, 7, 8).
Наиболее раннее появление Eisenackia circumtabulata, выявленное в разрезе Бешкош в ДН-3, в Европейской палеогеографической области зафиксировано в нижней части нижнего маастрихта на юге Германии (Kirsch, 1991) и на севере Италии (Roncaglia, 2002).
Данные по диноцистам для северо-запада Германии (разрез Hemmoor, Нижняя Саксония) имеются для верхов нижнего маастрихта–нижней части верхнего маастрихта (Marheinecke, 1992). Сопоставление показывает, что в разрезе Бешкош выявляется только один корреляционный уровень – нижняя граница подзоны 2 зоны А, которая проводится по появлению видов Eisenackia circumtabulata и Caligodinium amiculum (рис. 6). Первая совместная встречаемость этих видов в разрезе Бешкош установлена в терминальной части пачки ХIХb (проба 43, т.н. 3110, кровля ДН-3). В разрезе Hemmoor основание подзоны 2 зоны А приурочено к верхней части зоны B. sumensis по белемнитам (по шкале Schulz, Schmid, 1983), которая датируется второй половиной раннего маастрихта. Обращает на себя внимание узкий диапазон распространения Eisenackia circumtabulata в разрезе Бешкош, в то время как в разрезе Hemmoor она присутствует вплоть до нижней части верхнего маастрихта.
В ДН-4 выявлено присутствие Alterbidinium minus, Alterbidinium varium, совместная встречаемость которых в стратотипе нижней границы маастрихта в разрезе Терси установлена в нижнем маастрихте (Antonescu et al., 2001; Schiøler, Wilson, 2001) (рис. 5). Появление Trithyrodinium evittii, выявленное в нижней части ДН-4, в разрезе Терси (Antonescu et al., 2001; Schiøler, Wilson, 2001) приурочено к терминальной части кампана. В Бельгии это событие распознано в основании зоны Membranilarnacia liradiscoides нижнего маастрихта (Slimani, 2000, 2001) (рис. 7).
Сравнение ассоциаций диноцист разреза Бешкош с комплексами диноцист, выявленными на юге Западной Сибири (Лебедева и др., 2013), показывает, что на основании присутствия Cerodinium diebelii, Palaeocystodinium golzowense, Cladopyxidium spp., Alterbidinium varium, Alterbidinium acutulum диноцистовые интервалы ДН-4–ДН-7 разреза Бешкош могут быть сопоставлены со слоями с Cerodinium diebelii, а интервал ДН-8, исходя из присутствия Cerodinium speciosum, отсутствия Alterbidinium acutulum, – со слоями с Cerodinium speciosum из ганькинской свиты, охарактеризованными наннопланктоном зон СС24–CC25a (Perch-Nielsen, 1985)/UC18–UC19 (Burnett, 1998) нижнего маастрихта (рис. 8).
Сопоставление полученных данных по диноцистам из разреза Бешкош с зональной схемой по диноцистам для юга Германии (Kirsch, 1991) показывает, что интервалы ДН-4–ДН-7 (пачка XX–нижняя часть пачки XXI) разреза Бешкош уверенно коррелируются с подзонами Spongodinium delitiense и Alterbidinium varium зоны Cerodinium diebelii нижнего маастрихта–нижней части верхнего маастрихта. Их сближает совместная встречаемость зональных и руководящих видов – Cerodinium diebelii, Spongodinium delitiense, Alterbidinium varium и Riculacysta? pala. Граница между подзонами Spongodinium delitiense и Alterbidinium varium, фиксируемая по исчезновению S. delitiense (Kirsch, 1991), в разрезе Бешкош установлена в нижней части ДН-4 в терминальной части подпачки ХХа (т.н. 3110, проба 53) (рис. 3, 6). Более высокая часть разреза Бешкош (верхняя часть пачки XXI–пачка XXII) на основании присутствия в ДН-8 Cerodinium speciosum может быть сопоставлена с подзоной Deflandrea galeata зоны Cerodinium diebelii, объем которой охватывает верхнюю часть зоны G. gansseri по планктонным фораминиферам и датируется средней частью маастрихта (Kirsch, 1991) (рис. 6).
Наблюдаемые в интервале ДН-4 совместная встречаемость видов Cladopyxidium paucireticulatum, Spongodinium delitiense, Cerodinium alberti, Palaeocystodinium golzowense, Riculacysta? pala, Alterbidinium varium, исчезновение Samlandia mayi установлены в разрезах Бельгии в подзоне “а” зоны Pervosphaeridium tubuloaculeatum (Slimani, 2000, 2001), объем которой охватывает зоны Belemnitella sumensis–Belemnitella fastigata верхней части нижнего маастрихта (рис. 7).
Верхняя граница подзоны “а” зоны Pervosphaeridium tubuloaculeatum установлена по исчезновению Alterbidinium acutulum, Isabelidinium bujakii (= I. cooksoniae), Neosphaerodictyon filosum (Slimani, 2000, 2001). Первые два биособытия в разрезе Бешкош установлены в нижней части ДН-8 (середина пачки XXI), а исчезновение Neosphaerodictyon filosum – на более низком стратиграфическом уровне, в кровле ДН-4 (граница подпачек XXb и ХХс). На основании появления в ДН-8 Cerodinium speciosum (верхняя часть пачки XXI–пачка XXII) эта часть разреза Бешкош может быть сопоставлена с зоной Deflandrea galeata, объем которой в Бельгии охватывает верхнюю часть белемнитовой зоны Belemnitella junior и датируется первой половиной позднего маастрихта.
В бассейне Северного моря, на юге Скандинавии, зональное расчленение верхнего мела ограничено маастрихтом (Hansen, 1977, 1979; Hulthberg, Malmgren, 1987; Schiøler, Wilson, 1993) (рис. 8). Вид-индекс зоны Triblastula utinensis (Schiøler, Wilson, 1993), охватывающей в Северном море зоны по фораминиферам Neoflabellina reticulata (верхняя часть)–Gavelinella danica (нижняя часть) (по шкале Koch, 1977) и зоны Belemnetella lanceolata (верхняя часть)–Belemnitella junior (середина) по белемнитам нижнего маастрихта – нижней части верхнего маастрихта, появляется в разрезе Бешкош в интервале ДН-7 (терминальная часть подпачки ХХс).
Зона Triblastula utinensis в бассейне Северного моря подразделена на три подзоны, верхние границы которых фиксируются по последнему присутствию Eatonicysta hapala (маркирует кровлю зоны по белемнитам Belemnella lanceolata), Alterbidinium acutulum (несколько ниже кровли зоны Belemnella occidentalis), Triblastula utinensis (в средней части Belemnitella junior) соответственно (Schiøler, Wilson, 1993). При всей кажущейся сходимости последовательности этих биособытий в разрезе Бешкош и бассейне Северного моря, выявляются нюансы. Так, Eatonicysta hapala в разрезе Бешкош встречается спорадически в виде единичных экземпляров, и ее исчезновение зафиксировано в кровле ДН-6, т.е. раньше уровня первого появления Triblastula utinensis. Исчезновение Alterbidinium acutulum установлено в нижней части ДН-8 (средняя часть пачки XXI) (рис. 8).
Уровень исчезновения Triblastula utinensis, являющийся маркером верхней границы одноименной зоны (Schiøler, Wilson, 1993), предположительно приходится на нижнюю часть пачки XXII. Вышележащая зона Isabelidinium cooksoniae, объем которой определяется как интервал от последнего присутствия Triblastula utinensis до последнего присутствия Isabelidinium cooksoniae (Schiøler, Wilson, 1993), в разрезе Бешкош уверенно не распознается, что, скорее всего, обусловлено отсутствием диноцист в большей части пачки XXII. Последнее присутствие данных таксонов выявлено в нижней части ДН-8, а выше по разрезу Isabelidinium cooksoniae не обнаружен.
Следует обратить внимание на распространение видов рода Triblastula и в других регионах. Так, появление Triblastula wilsonii, T. utinensis в разрезах Бельгии установлено практически на одном стратиграфическом уровне и приурочено к зоне Pervosphaeridium tubuloaculeatum, а исчезновение этих видов фиксируется в зоне Deflandrea galeata нижней части верхнего маастрихта (Slimani, 2000, 2001) (рис. 7). В Тургайском прогибе данные виды рода Triblastula также встречены совместно в верхней части журавлевской свиты на уровне находок белемнитов Neobelemnella kazimiroviensis Skołozdrowna и моллюсков Oxytoma danica (J. Ravn) нижнего маастрихта (Васильева, Левина, 2007). На юге Западной Сибири (Лебедева и др., 2013) Triblastula wilsonii, T. utinensis появляются в нижнем маастрихте на разных стратиграфических уровнях. Сначала на уровне наннопланктонной зоны СС24 (Perch-Nielsen, 1985)/UC18 (Burnett, 1998) и слоев с Cerodinium diebelii появляется T. utinensis, а позже, в подзоне СС25а (Perch-Nielsen, 1985)/зоне UC19 (Burnett,1998) и слоях с Cerodinium speciosum, появляется Triblastula wilsonii (рис. 8). В разрезе Бешкош данные виды имеют различные диапазоны распространения, но не встречены совместно и характеризуются обратной динамикой появления: Triblastula wilsonii присутствует в нижней части разреза (интервал ДН-2–ДН-5), Triblastula utinensis – в верхней (интервал ДН-7–ДН-8 (часть)) (рис. 3, 4).
На основе приведенных данных можно предположить одновременное появление в раннем маастрихте Triblastula utinensis в Бореальной области и Triblastula wilsonii в Тетической области. Как следствие, распространение данных таксонов в различных регионах было обусловлено палеогеографическим положением. С проникновением теплых тетических водных масс в Бореальную область происходила миграция Triblastula wilsonii, что можно наблюдать в разрезах Бельгии, Тургайского прогиба, Западной Сибири. Соответственно, с внедрением бореальных вод в Тетическую облать (что, как правило, вело к понижению температуры бассейна) происходила миграция на юг Triblastula utinensis. Учитывая, что появление Triblastula utinensis в разрезе Бешкош происходит до исчезновения Alterbidinium acutulum, установленного синхронно в различных регионах Евразии (Kirsch, 1991; Schiøler, Wilson, 1993; Slimani, 2000, 2001; Лебедева и др., 2013), можно предположить, что во второй половине раннего маастрихта на данную территорию произошло проникновение бореальных вод и, как следствие, началось похолодание.
Зональных таксонов, характеризующих в различных регионах Евразии верхнюю часть верхнего маастрихта, – Palynodinium grallator, Thalassiphora pelagica, Hystrichostrogylon borissii и др. (Hansen, 1977, 1979; Hultberg, Malmgren, 1987; Kirsch, 1991; Schiøler, Wilson, 1993; Schiøler et al., 1997; Brinkhuis, Schiøler, 1996; Nøhr-Hansen, 1996, 2012; Williams et al., 2004; Slimani et al., 2011, 2016 и др.) – в разрезе Бешкош не установлено. В средней части пачки XXIII в интервале ДН-9 присутствует Glaphyrocysta perforata, чье появление в Датском бассейне зафиксировано на уровне подзоны Cannosphaeropsis utinensis зоны T. utinensis (Schiøler, Wilson, 1993; Surlyk et al., 2013). Однако, учитывая выявленную в ДН-9 массовую встречаемость Glaphyrocysta perforata, которая в Датском бассейне зафиксирована на уровне нижней части наннопланктонной зоны Nephrolithus frequens и нижней части подзоны Rugoglobigerina rugosa–Rugoglobigerina rugosa зоны Heterohelix striata–Globigerinelloides multispina по планктонным фораминиферам (Hultberg, Malmgren, 1987), эта часть разреза Бешкош может быть датирована средней частью позднего маастрихта.
Таким образом, сравнение биособытий по диноцистам, установленных в ДН-1–ДН-3, с аналогичными биособытиями, выявленными в разрезах Западной Европы, показывает, что первые приурочены к позднему кампану–началу раннего маастрихта. Это позволяет отнести пачку XIX и нижнюю часть пачки XX разреза Бешкош к пограничному интервалу кампана и маастрихта. Если опираться на критерии распознавания нижней границы маастрихта по диноцистам, выявленные в разрезе Терси (Odin, Lamaurelle, 2001), то этот уровень в разрезе Бешкош приурочен к границе ДН-2 и ДН-3. Исходя из приведенных данных, нижнюю часть разреза, включающую интервалы ДН-1 и ДН-2, можно датировать терминальным кампаном, интервал ДН-3 – ?началом раннего маастрихта, ДН-4–ДН-7 – концом раннего маастрихта, ДН-8 – началом позднего маастрихта, ДН-9 – средней частью позднего маастрихта (рис. 9).

Палеообстановки
Ранее для различных регионов было установлено, что повышение уровня моря в неритической зоне, связанное с трансгрессивными трактами (в терминах сиквенс-стратиграфии), вызывает массовое развитие динофлагеллат семейства Areoligeraceae в обстановках внутреннего шельфа (например, Brinkhuis, 1994; Schiøler et al., 1997; Iakovleva et al., 2001; Pross, Brinkhuis, 2005; Crouch, Brinkhuis, 2005; Sluijs et al., 2008). Обилие в ДН-2 таксонов этого семейства, а также постоянная встречаемость Cribroperidinium spp. свидетельствуют о формировании отложений, отвечающих нижней–средней частям подпачки XIXb, в пределах внешнего шельфа в обстановках с высокой гидродинамикой во время трансгрессивной фазы развития бассейна (рис. 4). Здесь же фиксируется значительное участие акритарх Fromea, Paleostomocystis reticulata, что может свидетельствовать о стратификации водного столба (Sluijs et al., 2005; Prauss, 2007). Крайне малое количество и спорадическая встречаемость представителей перидиниоидных диноцист указывают на низкую продуктивность.
Количественные изменения различных групп диноцист в ДН-3 (терминальная часть подпачки XIXb) – постепенное уменьшение количества Areoligeraceae на фоне возрастания Spiniferites spp., Hystrichosphaeridium spp. – свидетельствуют о сохранении обстановок внешней зоны шельфа. Заметное присутствие в ДН-3 представителей открыто-морских и океанических экогрупп диноцист (Spiniferites spp., Pterodinium cingulatum, Impagidinium sp.) позволяет реконструировать уровень высокого стояния моря на этом этапе развития бассейна и формирование подпачки XIXb в условиях внешнего шельфа (Brinkhuis, 1994; Brinkhuis, Schiøler, 1996; Schiøler et al., 1997; Pross, Brinkhuis, 2005).
Для времени осадконакопления подпачки ХХa (интервал ДН-4) фиксируется разнообразие и высокое содержание перидиниоидных таксонов (Cerodinium spp., Alterbidinium spp., Isabelidinium spp., Senegalinium obscurum); это свидетельствует о палеообстановках внутреннего шельфа, росте эвтрофикации, вероятно за счет апвеллинга, что косвенно подтверждается появлением зерен глауконита.
Выше, в основании подпачки ХХс (интервал ДН-5) вновь установлено значительное участие видов Areoligera, что указывает на новый трансгрессивный импульс во время формирования пород.
В целом таксономически схожие с ДН-3–ДН-5 комплексы диноцист известны из нижнего маастрихта Италии (Corradini, 1973), юга Германии – Гельветские и Ультрагельветские покровы Альп (Kirsch, 1991), отчасти Бельгии (Slimani, 2000, 2001), что позволяет предположить их существование в единой палеобиогеографической области. Это подтверждается и находками макрофауны.
Весьма интересным является интервал ДН-6, соответствующий средней части пачки ХХ. В основании интервала выявлено присутствие различных видов Laciniadinium, частая встречаемость которых характерна для верхнемеловых пород Западной Сибири и Полярного Предуралья (Лебедева, 2005; Lebedeva, 2006; Лебедева, Кузьмина, 2018). Этот факт свидетельствует об изменении циркуляции и, вероятно, о понижении температуры палеобассейна в связи с внедрением бореальных водных масс, что подтверждается находками белемнитов. По мере развития трансгрессии, отражающейся в прогрессирующем увеличении содержания рода Spiniferites spp., обитавшего в открыто-морских условиях, в верхней части ДН-6 отмечена частая встречаемость перидиниоидных таксонов (Trithyrodinium spp., Pierceites spp.), что указывает на увеличение поступления питательных веществ в море (эвтрофикацию) и обмеление. В то же время с этого уровня и выше, включая пачки XXI–XXII, в разрезе отмечается постоянная относительно частая встречаемость акритарх Fromea spp., Paralecaniella indentata, празинофитов, что косвенно свидетельствует о стратификации водного столба (Prauss, 2007; Sluijs et al., 2005).
В нижней части пачки XXI (интервал ДН-7) установлено снижение разнообразия диноцист, появление высокоширотного (бореального) вида Triblastula utinensis, что позволяет предполагать прогрессирующее понижение температуры воды в морском бассейне в конце раннего маастрихта.
Верхняя часть пачки XXI–нижняя часть пачки XXII (нижняя часть интервала ДН-8) характеризуются возрастанием разнообразия диноцист и количества перидиниоидных таксонов, снижением количества Spiniferites spp., частой встречаемостью акритарх Paralecaniella indentata, Fromea spp., празинофитов. Такой состав ассоциации свидетельствует о формировании пород в мелководном морском бассейне (Brinkhuis, Schiøler, 1996; Schiøler et al., 1997). Эти изменения фиксируют понижение уровня моря.
Выше, в средней части пачки ХХIII (интервал ДН-9) установлено обилие Glaphyrocysta perforata, что указывает на высокую гидродинамику и трансгрессивный импульс во время формирования отложений.
МАГНИТОСТРАТИГРАФИЯ
Петромагнитные и магнито- минералогические исследования
Петромагнитные данные необходимы для обоснования вида и генезиса минералов-носителей естественной остаточной намагниченности и выяснения особенностей их распределения по разрезу. Эта информация используется для оценки пригодности образцов к палеомагнитным исследованиям и доказательства возраста намагниченности. Петромагнитные данные имеют самостоятельную ценность, как инструмент для дополнительного расчленения и корреляции разрезов. В Стратиграфическом кодексе России (2019) петромагнитные подразделения (магнитозоны, выделенные по численным магнитным характеристикам, не связанным с древним геомагнитным полем), наряду с магнитополярными (палеомагнитными), фигурируют как магнитостратиграфические подразделения. По сути, они являются разновидностью литостратиграфических подразделений, поскольку так же, как и последние, выделяются по особенностям вещественного состава (а именно, по характеру ферромагнитной фракции) пород. Вариации петромагнитных свойств отражают многие особенности формирования отложений, основные принципы их геологической интерпретации разработаны ранее и многократно апробированы на объектах различного возраста и генезиса (Молостовский, Храмов, 1997; Evans, Heller, 2003; Гужиков, 2013 и многие другие).
В разрезе Бешкош у образцов со 176 стратиграфических уровней измерены следующие параметры (рис. 10): K – магнитная восприимчивость и ее анизотропия (АМВ); Kt (термокаппа) – магнитная восприимчивость после прогрева пород при температуре 500°C в течение часа (прирост dK = = Kt – K при этом отражает содержание тонкодисперсного пирита в образце, благодаря фазовому переходу немагнитного FeS2 в сильномагнитный Fe3O4 при температуре свыше 400°C); Jn – естественная остаточная намагниченность; Jrs – остаточная намагниченность насыщения (максимально возможная остаточная намагниченность в образце, создаваемая искусственным магнитным полем); Bcr – поле, соответствующее остаточной коэрцитивной силе, которую нужно приложить, чтобы полностью разрушить Jrs образца. Помимо характеристик, установленных опытным путем, рассчитывались параметры: фактор Q (параметр Кенигсбергера), равный отношению Jn к индуктивной намагниченности; параметр магнитной жесткости S = |Jr(–300)|/Jrs (где Jr(–300) – остаточная намагниченность после воздействия полем 300 мТл, обратном направлению поля насыщения) и отношение K/Jrs. Повышенные значения фактора Q указывают на более высокую степень упорядоченности магнитных моментов частиц. Величины параметра S, близкие к нулю, свидетельствуют о доминировании в образце магнитожесткой фазы, а близкие к единице – о преобладании магнитомягких минералов. Отношение K/Jrs пропорционально среднему размеру ферромагнитных зерен.
Рис. 10.
Магнитостратиграфический разрез Бешкош. Показаны палеомагнитная колонка, графики палеомагнитного склонения (D), наклонения (I) и петромагнитных параметров. Условные обозначения: 1, 2 – геомагнитная полярность: 1 – нормальная, 2 – обратная; 3 – отсутствие данных о полярности; 4 – данные по образцам из точек наблюдения (т.н.) 3110, 3136, 3171; 5 – данные по образцам из точки наблюдения 2561 (отбор В.А. Фомина); 6 – уровни, на которых смещение Jn в ходе магнитных чисток происходит по дугам больших кругов. Остальные условные обозначения см. на рис. 1.

Выборочные образцы, характеризующие различные литологические и петромагнитные разности, подвергались термомагнитному анализу (ТМА), заключавшемуся в исследованиях зависимостей намагниченности (J) или К от температуры (T) и исследованиям на коэрцитивном спектрометре J_meter (в Казанском федеральном университете – КФУ, Казань). В ТМА диагностическими признаками являются температура Кюри (выше которой исчезают ферромагнитные свойства вещества) или температуры фазовых переходов, уникальные для каждого минерала.
Измерения K велись на каппабридже MFK1-FB, измерения Jn – на спин-магнитометре JR-6 и криогенном магнитометре 2G-Enterprices (в Институте физики Земли – ИФЗ РАН, Москва). Гистерезисные характеристики (Jrs, Bcr) получены с помощью регулируемого электромагнита с максимальной интенсивностью поля 700 мТл (поэтому в данной работе за Jrs условно принята остаточная намагниченность после воздействия полем интенсивностью 700 мТл, заведомо достаточным для насыщения магнитомягких образцов). dK измерялось после нагрева образцов в печи СНОЛ-6/11-В. Исследования J(T) проводились на термоанализаторе фракций ТАФ-2 (“магнитные весы”), K(T) – на каппабридже MFK1-FA с приставкой CS3 (в ИФЗ РАН, Москва). Анализ зависимостей K(T) и данных АМВ проводился с помощью программ Cureval 8.0.2 и Anisoft 4.2 соответственно.
Изученные отложения, несмотря на малые значения К и Jn (K = 0.3–5.8 × 10–5 ед. СИ и Jn = = 0.006–0.52 × 10–3 А/м), хорошо дифференцированы по магнитным свойствам, вариации которых обнаруживают очевидные закономерности в распределении по разрезу (рис. 10).
Прежде всего, следует отметить закономерные тренды к увеличению вверх по разрезу величин параметров, обусловленных концентрацией ферромагнитных минералов в образце: K, Jn, Jrs (рис. 10). Эти тренды сопровождаются возрастанием количества терригенных зерен и глауконита, заметным визуально и подтвержденным данными количественного анализа – увеличением доли нерастворимого остатка (Alekseev, Kopaevich, 1997). Однако связь между возрастаниями значений K, Jn, Jrs и увеличением концентраций крупнообломочного материала неочевидна, потому что наблюдаемый спад отношения K/Jrs вверх по разрезу (за исключением пограничного маастрихтского–датского интервала), напротив, указывает на уменьшение среднего размера ферромагнитных зерен. О снижении размерности ферромагнитной фракции свидетельствует и возрастание Q вверх по разрезу, вследствие лучшего упорядочивания магнитных моментов у тонкодисперсных частиц, чем у крупных (рис. 10).
Ранее идентичный петромагнитный облик маастрихтских отложений был зафиксирован в разрезах Чахмахлы и Такма, расположенных в долине р. Бодрак, к ССЗ от г. Бешкош, примерно в 5 и 8 км соответственно (Гужикова, 2018). Исследования под микрозондом образцов из разреза Чахмахлы показали, что ферромагнитная фракция представлена в них субмикронными железистыми чешуйками (химический состав – Fe, Ni, Cr) и магнетитовыми микросферулами, характерными для метеорной пыли (Гужикова, 2018). Поэтому изменения магнитных свойств, свидетельствующие о возрастании тонкодисперсной ферромагнитной фракции вверх по разрезу, были связаны с повышенным поступлением метеоритного вещества на Землю в конце мелового периода (Гужикова, 2018).
Результаты магнито-минералогических анализов выборочных образцов по разрезу Бешкош согласуются с предположением о космогенном веществе в виде магнетитовых микросферул, как наиболее вероятном носителе намагниченности. В изученных образцах, за исключением образцов с двух уровней (3110-41 и 3110-44), фиксируется магнитомягкая фаза: Bcr = 15–44 мТл, S ~ 1, Jrs приобретается в полях до 200 мТл (рис. 11а). Подобная картина характерна для тонкодисперсного магнетита. Данные коэрцитометрии указывают на псевдооднодоменное и многодоменное состояние ферромагнитных частиц (Dunlop, 2002), характерное для магнетитовых сферул космического происхождения (рис. 11б). Интерпретация кривых ТМА, к сожалению, невозможна ввиду малой намагниченности образцов, с величиной которой сопоставим уровень приборных помех.
Рис. 11.
Результаты магнито-минералогических анализов. (а) – кривые магнитного насыщения и разрушения; (б) – диаграмма Дея: SD, PSD и MD – области однодоменных, песводооднодоменных и многодоменных частиц соответственно; (в) – данные по анизотропии магнитной восприимчивости: стереограммы проекций длинных (K1), средних (K2) и коротких (K3) осей АМВ в палеогеографической системе координат и диаграммы P–T (области положительных и отрицательных значений T соответствуют уплощенным и удлиненным формам ферромагнитных частиц соответственно). Условные обозначения: 1, 2, 3 – средние направления с овалами доверия K1, K2 и K3 соответственно.

Нельзя исключить, что обогащение тонкодисперсными ферромагнетиками связано с изменением состава и количества терригенной фракции и глауконита. Например, имеются сведения об образовании биогенного магнетита за счет железа, содержащегося в глауконите (Chaudhuri et al., 2001). Единичные зерна глауконита, действительно, появляются в XX пачке, а их количество возрастает к верхам разреза, что совпадает с трендом к увеличению тонкодисперсных магнитомягких частиц. Характерный размер частиц биогенного магнетита составляет 0.01 мкм, а максимальный – не превышает 0.1 мкм, но для них характерно однодоменное состояние, что не согласуется в полной мере с коэрцитометрическими данными (рис. 11б).
Наиболее выразительный рубеж, подразделяющий разрез на два петромагнитных комплекса – нижний (ПК-1) и верхний (ПК-2), расположен между обр. 3110-44 и 3110-45 (рис. 10). Он соответствует кровле пачки XIX и приурочен к границе диноцистовых интервалов ДН-3 и ДН-4. Этот уровень маркируется аномальными величинами Bcr = 300 мТл и S = 0.004 (обр. 3110-44), свидетельствующими, в совокупности с характером магнитного насыщения (рис. 11а), о доминировании магнитожесткой фазы – гидроокислов железа и/или гематита. На кривых K, Jn, Q и Jrs этот рубеж заметен по окончанию постепенного возрастания вверх по разрезу значений петромагнитных параметров: показатели K (на уровне обр. 3110-44), Jn и Q (на уровне обр. 3110-45) падают примерно в 2 раза, а Jrs (на уровне обр. 3110-45) – почти в 4 раза. Значимые изменения на этом рубеже происходят и в характере распределений величин K/Jrs по разрезу: ниже уровня обр. 3110-45 значения параметра сильно варьируют, а выше плавно убывают вверх по разрезу.
Разница между нижним и верхним петромагнитными комплексами наиболее очевидна на графике дисперсий K/Jrs, рассчитанном методом скользящего окна размером в три образца, перемещаемого по разрезу с шагом в один образец (рис. 10). Из отличительных особенностей ПК-1 следует отметить также ничтожно малые концентрации ферромагнетиков, судя по минимальным для всего разреза значениям K, Jn, Jrs (рис. 10).
Вышележащие отложения уникальны тем, что в них на фоне спада или практического отсутствия прироста магнитной восприимчивости (dK варьирует от (–0.8) до 0.4 × 10–5 ед. СИ) рельефно выделяются два интервала (между обр. 3110-48 и 3110-54 и обр. 3136-9 и 3136-20), внутри которых встречаются значения dK от 1 до 7 × 10–5 ед. СИ, указывающие на присутствие тонкодисперсного пирита (рис. 10). Кровля разреза (обр. с 3171-18 по 3171-22) выделяется максимальными для всего разреза значениями K > 3 × 10–5 ед. СИ (и, вследствие этого, повышенными значениями K/Jrs), связанными, очевидно, с высокими концентрациями парамагнитного глауконита в пограничном интервале маастрихта–дания. ПК-2 очень однороден по Bcr и S, но характерные изменения в трендах кривых Jrs, K/Jrs (на уровнях между обр. 3136-2 и 3136-3 и между обр. 3136-30 и 3136-31), термокаппаметрические вариации в его пределах и граница между ПК-1 и ПК-2 (рис. 10) могут стать ценными корреляционными реперами, при условии их латеральной устойчивости.
Значительные различия в петромагнитном облике ПК-1 и ПК-2 указывают на то, что между обр. 3110-44 и 3110-45 располагается уровень, разделяющий комплексы пород, формировавшихся в существенно разных условиях. Подобные уровни, как правило, связаны с поверхностями перерывов в осадконакоплении, для которых также характерно обогащение гидроокислами железа и/или гематитом.
Распределение осей эллипсоидов магнитной восприимчивости в изученных отложениях, в первом приближении, соответствует первичной осадочной текстуре (при которой проекции коротких осей группируются в центре полярной стереопроекции, а проекции длинных и средних осей равномерно распределены по экватору), но проекция среднего направления K3 смещена к северу от центра приблизительно на 20° (рис. 11в). Магнитная текстура отложений практически не изменяется после прогрева пород до 500°C и одинакова в разных пачках (за исключением верхов пачки XXIII, обогащенных глауконитом). Отклонение проекции среднего направления K3 может быть обусловлено наклоном дна палеобассейна к югу и вязкопластичными смещениями осадка в этом направлении.
Палеомагнитные исследования
Из каждого ориентированного штуфа выпиливалось по 3–4 образца кубической формы, размером 2 × 2 × 2 см, предназначенных для лабораторных измерений. Палеомагнитные исследования проводились по стандартной методике (Молостовский, Храмов, 1997), включавшей в себя измерения Jn образцов на спин-магнитометре JR-6 (в СГУ) и криогенном магнитометре (SQUID) 2G-Enterprices (в ИФЗ РАН) после серии последовательных магнитных чисток переменным полем (до 50–100 мТл с шагом 3–5 мТл). Размагничиванию переменным полем подвергались 1–2 образца с каждого уровня. Для компонентного анализа использовалась программа Remasoft 3.0.
В образцах со 131 уровня (что составляет ~75% от общего объема палеомагнитной коллекции) выделены характеристические компоненты Jn (ChRM), максимальный угол отклонения (мера точности, с которой прямая линия аппроксимирует набор точек на диаграмме Зийдервельда) которых не превышает 15° (рис. 12).
Рис. 12.
Результаты компонентного анализа. Слева направо представлены: стереографические изображения изменения векторов Jn в процессе магнитных чисток, диаграммы Зийдервельда (в стратиграфической системе координат), графики размагничивания образцов. Условные обозначения: 1, 2 – проекции Jn на нижнюю (1) и верхнюю (2) полусферы; 3, 4 – проекции Jn на горизонтальную (3) и вертикальную (4) плоскости.

Проекции палеомагнитных векторов, группирующиеся в северных румбах нижней полусферы (рис. 13а), интерпретированы нами как отражение нормальной полярности (N) геомагнитного поля. Совокупность этих направлений характеризуется относительно большим разбросом, одной из причин которого, вероятно, является невозможность точного определения элементов залегания слоев на каждом уровне. Среднее направление совокупности статистически совпадает с направлением современного поля и значимо отличается от направления маастрихтского поля, пересчитанного для района Бешкоша из стандартных полюсов с возрастом 70 млн лет для стабильной Европы (Besse, Courtillot, 2002) (рис. 13б). Эти обстоятельства являются аргументами в пользу вторичной природы Jn, но не доказательствами полного перемагничивания пород. ChRM может представлять собой сумму первичной и вторичной компонент, разделить которые невозможно ввиду близости направлений позднемелового и современного поля. Результаты сравнения палеомагнитных данных по разрезу Бешкош со стандартными полюсами не исключают первичности намагниченности, потому что геодинамическая позиция Горного Крыма изменялась в кайнозое относительно внеальпийской Европы. К сожалению, надежные палеомагнитные данные по маастрихту Юго-Западного Крыма (как и по всему верхнему мелу Крыма), которые можно было бы использовать для корректного сопоставления с целью проверки первичности намагниченности, в настоящее время отсутствуют.
Рис. 13.
Стереопроекции в стратиграфической системе координат. (а, б) – ChRM, соответствующие нормальной полярности (а) и предположительно обратной полярности (б); (в) – направления перемагничивания современным полем (I), среднее направление ChRM, соответствующее нормальной полярности (II), и направления, пересчитанного для района разреза Бешкош из стандартного полюса (70 млн лет) для стабильной Европы (III); (г) – таблица с угловыми расстояниями, образуемыми векторами I, II и III. Ошибки (±), определяемые статистикой этих векторов, приведены согласно (Debiche, Watson, 1995). Если угол больше ошибки, то вектора различаются значимо, если наоборот, то вектора статистически совпадают. Для современного поля за α95 принята максимальная амплитуда вековой вариации (Бахмутов, 2006). Палеомагнитная статистика: N – число образцов в выборке, D, I – среднее палеомагнитное склонение и наклонение соответственно, k – палеомагнитная кучность, α95 – радиус круга доверия вектора. Условные обозначения: 1 – средние палеомагнитные направления, 2 – круг доверия (α95). Остальные условные обозначения см. на рис. 12.

На некоторых уровнях обнаруживаются направления, характерные для обратной полярности (R), или аномальные направления (имеющие, например, пологие наклонения в южных румбах), или смещения проекций Jn в процессе магнитных чисток по дугам больших кругов. Последние два случая характерны для частично перемагниченных образцов, в которых ChRM представляет собой векторную сумму вторичной и не полностью уничтоженной первичной компонент. В разрезе зафиксировано 17 подобных уровней, 6 из которых спорадически рассеяны по разрезу, а 11 группируются в интервале на границе пачек XXI и XXII (между обр. 3136-24 и 3136-35) (рис. 10), что вряд ли можно считать случайностью. Более вероятным выглядит предположение о частичной сохранности в этом интервале древней компоненты Jn, соответствующей древнему полю. Данный участок выделен нами как магнитозона предположительно обратной полярности – R(?)-зона, мощностью ~20 м (рис. 10).
Многочисленные магнитостратиграфические данные, полученные по маастрихтским отложениям разных континентов и осадочного слоя океанской коры (Гужиков и др., 2007; Guzhikov et al., 2020; Thibault et al., 2012a, 2012b; Thibault, 2016; Гнибиденко и др., 2014 и др.), принципиально не отличаются друг от друга: повсеместно низам маастрихта соответствует магнитозона обратной полярности (аналог хрона C31r), верхам яруса – магнитозона нормальной полярности (аналог хрона C30n), граница маастрихта–дания отмечена магнитозоной обратной полярности (аналог хрона C29r). Эти представления обобщены в магнитохронологической шкале (Ogg et al., 2016), и при сопоставлении с ней палеомагнитной колонки Бешкоша очевидно, что R(?)-зона может быть только аналогом кратковременного (0.173 млн лет) хрона С30r (рис. 14). Иные варианты сопоставления не представляются возможными. Основание хрона C31r расположено вблизи подошвы маастрихтского яруса, поэтому отождествление R-зоны с C31r означало бы обоснование границы кампана–маастрихта вблизи подошвы пачки XXI, что противоречит всем существующим макро- и микрофаунистическим данным. Корреляция R(?)-зоны с хроном C29r исключена, потому что последний соответствует пограничному интервалу мела и палеогена.
Рис. 14.
Палеомагнитное и хемостратиграфическое сопоставление разреза Бешкош с GSSP маастрихта (разрез Терси, Франция) и шкалой геомагнитной полярности (GPTS). Условные обзначения см. на рис. 1 и 10.
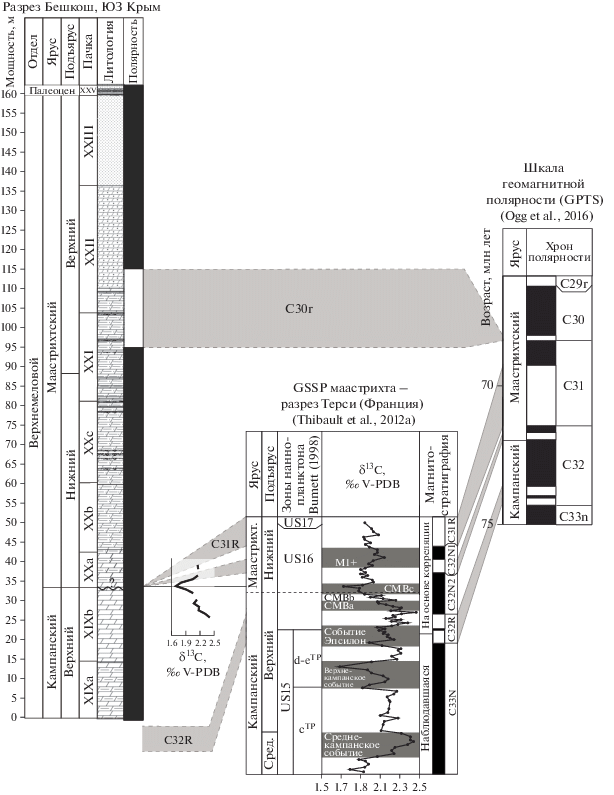
Отношение мощности магнитозоны обратного знака к длительности хрона C30r дает оценку средней скорости осадконакопления – 11.6 см/тыс. лет. На первый взгляд, это высокая скорость, но она совпадает со средней скоростью осадконакопления (11.7 см/тыс. лет), полученной при циклостратиграфическом анализе петромагнитных данных маастрихтских отложений в соседних разрезах Чахмахлы и Такма (расположенных к ЮВ от разреза Бешкош, в ~5 и ~8 км соответственно, см. рис. 1б) (Суринский, Гужиков, 2019). Интервал разреза, соответствующий R(?)-зоне, не уникален по литологическим, петромагнитным или иным признакам (рис. 10), поэтому нет оснований предполагать, что процессы перемагничивания затронули его в меньшей степени. Если же интенсивность перемагничивания всех пород была примерно одинаковой, то, кроме R(?)-зоны, других интервалов, формировавшихся в эпоху обратной полярности, скорее всего, в разрезе не было. Однако тезис об одинаковой степени перемагничивания всех изученных отложений не бесспорен, и, таким образом, следует признать, что имеющиеся в распоряжении материалы не позволяют однозначно обосновать первичный или вторичный генезис намагниченности, направление которой соответствует нормальной полярности поля. Поэтому нельзя постулировать изначальное отсутствие палеомагнитной записи хрона C31r в разрезе. Возможно, соответствующий ему интервал оказался впоследствии полностью перемагничен.
ИЗОТОПНО-ГЕОХИМИЧЕСКИЙ АНАЛИЗ
Для определения изотопного состава углерода и кислорода в 14 образцах карбонатных пород, отобранных из пограничного интервала кампана–маастрихта, был использован комплекс аппаратуры корпорации Thermoelectron, включающий масс-спектрометр Delta V Advаntage и установку Gas-Bench-II. Разложение карбонатов проводилось в 100%-ной ортофосфорной кислоте при 50°С. Значения δ13С и δ18О даны в промилле относительно стандарта V-PDB. Для привязки результатов анализов к V-PDB использовались сверочные пробы IAEA С-О-1 и NBS-19. Точность определения δ18О и δ13C находится в пределах ±0.2‰ и ±0.1‰ соответственно.
Значения δ13С варьируют от 1.8 до 2.4‰ (табл. 1, рис. 14). В средней части опробованного интервала отчетливо выделяется отрицательный экскурс δ13С с амплитудой ~0.4‰. Этот изотопный минимум зафиксирован в разрезе Бешкош в пограничном интервале кампана–маастрихта, что обосновано биостратиграфическими данными, и может быть сопоставлен с аналогичными отрицательными пиками δ13С, наблюдаемыми несколько выше подошвы маастрихта в разрезе GSSP яруса в Юго-Восточной Франции (Thibault et al., 2012a) (рис. 14) и в других опорных разрезах верхнего кампана–маастрихта, например Губбио в Италии (Coccioni, Premoli-Silva, 2015), Кронсмоор в Северной Германии (Niebuhr et al., 2011), скважинах глубоководного бурения в Атлантическом (Thibault, Gardin, 2007) и Индийском океанах (Thibault et al., 2012b). Исчерпывающие сводки и сопоставления изотопно-геохимических данных по этим и многим другим разрезам верхнего кампана–маастрихта приведены в работах Н. Тиболта и его соавторов (Thibault et al., 2012a, 2012b; Thibault, 2016).
Таблица 1.
Результаты определений изотопного состава углерода и кислорода в разрезе Бешкош
| Номер образца | δ13C, ‰ V-PDB | δ18О, ‰ V-PDB | Т, °С |
|---|---|---|---|
| 3110-35 | 2.4 | –1.5 | 23.1 |
| 3110-36 | 2.3 | –1.4 | 22.8 |
| 3110-37 | 2.2 | –1.3 | 22.4 |
| 3110-39 | 2.1 | –1.3 | 22.2 |
| 3110-40 | 2.1 | –1.4 | 22.7 |
| 3110-42 | 2.0 | –1.2 | 21.7 |
| 3110-43 | 1.9 | –1.4 | 22.8 |
| 3110-45 | 1.8 | –1.5 | 23.4 |
| 3110-46 | 2.0 | –1.3 | 22.5 |
| 3110-47 | 2.2 | –1.3 | 22.3 |
| 3110-49 | 2.2 | –1.3 | 22.4 |
| 3110-50 | 2.2 | –1.8 | 24.7 |
Вариации δ18О в 11 из 12 проанализированных образцов (–1.2…–1.5‰) не выходят за пределы аналитической ошибки. Немного ниже эта величина только в обр. 3110-50 (–1.8‰). Корреляция δ13С и δ18О отсутствует, что говорит о весьма слабых постседиментационных изменениях и синседиментационной природе отрицательного С-изотопного экскурса. Изотопно-кислородные температуры формирования карбонатов, рассчитанные при условии равновесия с современной средней океанической водой (δ18О = 0‰), соответствуют 22–23°С (Epstein et al., 1953). Если предположить, что вода была на 1‰ обеднена тяжелым изотопом кислорода (Ice-Free Earth), рассчитанные температуры можно оценить величинами ~18–19°С, которые представляются вполне реалистичными.
ЗАКЛЮЧЕНИЕ
Существующие представления о стратиграфии разреза горы Бешкош, согласно которым пачка XIX относилась к верхнему кампану, пачки XX–XXI – к нижнему маастрихту, а пачки XXII–XXIII – к верхнему маастрихту, базировались на результатах анализа находок белемнитов, аммонитов, достоверность которых трудно проверить из-за отсутствия изображений, и фораминифер (Заклинская, Найдин, 1985; Алексеев, 1989; Alekseev, Kopaevich, 1997).
Находка аммонита Pachydiscus (P.) neubergicus neubergicus (Hauer) позволила уточнить возможное положение кампан-маастрихтской границы. Новые находки других головоногих моллюсков важны для корреляции, но детализировать положение ярусных или подъярусных границ по ним нельзя. Анализ существующих данных по распространению остатков головоногих моллюсков в разрезе позволяет заключить, что граница кампанского и маастрихтского ярусов должна находиться не выше подпачки XXa, а граница подъярусов маастрихта – в интервале пачек XXI–XXIII.
Полученные данные по стратиграфическому распространению диноцист позволяют предполагать наличие перерывов между диноцистовыми интервалами ДН-3 и ДН-4, ДН5 и ДН-6, объем которых на данной стадии изученности диноцист региона точно оценить невозможно. Исходя из приведенных корреляций по диноцистам с разрезами Западной Европы, можно предположить, что перерыв между ДН-3 и ДН-4 охватывает интервал значительной части нижнего маастрихта, хотя не исключено, что столь резкие изменения таксономического состава диноцист обусловлены палеоэкологическими факторами. Наиболее вероятным представляется, что перерыв между интервалами ДН-5 и ДН-6 обусловлен сменой циркуляции водных масс, что привело к размыву слабо консолидированных осадков.
На основании палинологических данных возраст подпачки XIXa и большей части подпачки XIXb принимается как вторая половина позднего кампана, терминальная часть подпачки XIXb с долей условности датируется ранним маастрихтом, пачка ХХ и нижняя часть пачки XXI – ранним маастрихтом, верхняя часть пачки XXI–пачка XXIII – поздним маастрихтом (без терминальной части, которая размыта).
Таким образом, подтверждаются данные по макрофауне о положении границы кампана и маастрихта ниже кровли пачки ХХ. Однако требуется дополнительное изучение ортостратиграфических групп – планктонных фораминифер и наннопланктона, которое позволит более точно установить положение ярусной границы и ценить объем выявленного перерыва.
Данные по стабильным изотопам хорошо согласуются с заключением о положении ярусной границы вблизи подошвы пачки XX. На кривой δ13C этому уровню соответствует отчетливый отрицательный пик, аналогичный изотопным маркерам, фиксируемым выше подошвы маастрихта в GSSP яруса (Thibault et al., 2012 a) (рис. 14) и в других разрезах (Thibault et al., 2012a, 2012b; Thibault, 2016).
Магнитные свойства пород позволили получить нетривиальную геологическую информацию, отражающую особенности их вещественного состава, обусловленные седиментацией. Выделенные петромагнитные подразделения являются, по сути, петромагнитными фациями. Выявление уровня, обогащенного гидроокислами железа и/или гематитом, к которому приурочено резкое изменение большинства петромагнитных кривых, является косвенным подтверждением стратиграфического перерыва в подошве пачки XX. Петромагнитные подразделения, установленные в разрезе Бешкош, латерально устойчивы в пределах Второй гряды Крымских гор (Гужиков и др., 2018) и могут быть использованы для детального расчленения и корреляции разрезов.
В верхней части разреза (верхи пачки XXI и низы пачки XXII) зафиксирован интервал мощностью ~20 м, где, вероятно, частично сохранилась древняя компонента, соответствующая обратной полярности поля. Это единственная магнитозона обратного знака, выделенная нами в разрезе и предположительно идентифицированная как аналог хрона C30r. В остальной части разреза, характеризующейся нормальной полярностью, на основании имеющихся материалов невозможно доказать ни первичность намагниченности, ни, наоборот, частичное или полное перемагничивание отложений более поздним полем. Решение этой проблемы возможно на основе комплексного био- и магнитостратиграфического изучения одновозрастных отложений соседних разрезов.
Для того чтобы согласовать все имеющиеся данные по разрезу Бешкош, необходимо признать наличие перерыва в подошве пачки XX, в пользу которого свидетельствуют и петромагнитные, и палинологические, и литологические (внезапное появление обломочной составляющей) данные. Не прибегая к гипотезе перемагничивания, отсутствие аналогов хрона C31r можно объяснить крупным гиатусом в разрезе, допустив, что кровля C31r древнее подъярусной границы маастрихта, а принятые в GTS (Gradstein et al., 2012) представления о палеомагнитной характеристике подъярусов нуждаются в корректировке. Такой вариант не является чисто умозрительным. Как отмечалось ранее (Гужиков и др., 2017), положение границы нижнего и верхнего маастрихта в GTS нельзя считать однозначно определенным, а вопрос о выборе критерия для прослеживания подошвы верхнего маастрихта, по аналогии с потенциальным стратотипом подъярусной границы в Испании (Odin et al., 1996), остается открытым. Новые магнитостратиграфические данные по маастрихтским отложениям Поволжья свидетельствуют о том, что длительность хрона С31 в магнитохронологической шкале, возможно, преувеличена (Guzhikov et al., 2020).
Таким образом, новые данные подтверждают проведение границы кампанского и маастрихтского ярусов в разрезе Бешкош в подошве пачки XX (Alekseev, Kopaevich, 1997), но предполагают наличие в пограничном интервале значительного перерыва. Граница между нижним и верхним маастрихтом принимается нами в средней части пачки XXI – ниже, чем проводилась ранее (Алексеев, 1989; Яковишина и др., 2008).
По седиментологическим, ихнологическим и палинологическим данным установлено, что происходила закономерная смена обстановок от открыто-морских (внешний шельф) в начале маастрихта до прибрежных (предфронтальная зона пляжа) в конце маастрихта, завершившаяся, вероятно, кратковременным осушением бассейна на рубеже мела и палеогена. Палинологические данные, кроме того, позволили выявить ряд трансгрессивных импульсов и проникновение бореальных водных масс.
Благодарности. Мы признательны А.С. Алексееву и Л.Ф. Копаевич (МГУ), Елене Ягт-Языковой (University of Opole) и Джону Ягту (Natuurhistorisch Museum, Maastricht) за обсуждение полученных результатов и Н.Г. Зверькову (МГУ) за переданную нам находку стратиграфически важного аммонита Pachydiscus (P.) neubergicus neubergicus. Авторы благодарят М.Ю. Никитина (МГУ) за предоставленные фотоматериалы и данные фотограмметрии; В.Э. Павлова, Р.В. Веселовского и других сотрудников лаборатории Главного геомагнитного поля и петромагнетизма ИФЗ РАН за предоставление возможности измерений на криогенном магнитометре; Д.К. Нургалиева, Д.М. Кузину (КФУ) и А.М. Суринского (СГУ) за содействие в работе на коэрцитометре; П.А. Прошину (ГИН РАН, МГУ) за пробоподготовку образцов для изотопного анализа. Мы весьма признательны М.А. Рогову (ГИН РАН), благодаря которому рукопись была существенно улучшена.
Источники финансирования. Работа выполнена при финансовой поддержке РФФИ (проект №18-05-00784-а).
Список литературы
Александрова Г.Н., Беньямовский В.Н., Вишневская В.С., Застрожнов А.С. Новые данные по биостратиграфии верхнего мела Нижнего Поволжья // Стратиграфия. Геол. корреляция. 2012. Т. 20. № 5. С. 25–64.
Алексеев А.С. Верхний мел // Геологическое строение Качинского поднятия Горного Крыма. Стратиграфия мезозоя. Ред. Мазарович О.А., Милеев В.С. М.: Изд-во Моск. ун-та, 1989. С. 123–157.
Бахмутов В.Г. Палеовековые геомагнитные вариации. Киев: Наукова думка, 2006. 295 с.
Беньямовский В.Н., Гужиков А.Ю., Барабошкин Е.Ю., Александрова Г.Н., Первушов Е.М., Сельцер В.Б., Овечкина М.Н., Калякин Е.А., Копаевич Л.Ф., Вишневская В.С., Покровский Б.Г., Яковишина Е.В. Значение результатов комплексного изучения опорных разрезов Вольских карьеров Саратовского Поволжья и на плато Актолагай (Западный Казахстан) для установления нижней границы маастрихта на Восточно-Европейской платформе // Меловая система России и ближнего зарубежья: проблемы стратиграфии и палеогеографии. Ред. Барабошкин Е.Ю. Материалы Восьмого Всероссийского совещания, 26 сентября–3 октября 2016 г., Республика Крым. Симферополь: Издательский Дом Черноморпресс, 2016. С. 59–60.
Васильева О.Н., Левина А.П. Органикостенный микрофитопланктон в верхнемеловых и палеогеновых отложениях разреза Кушмурун Тургайского прогиба (Казахстан) // Бюлл. МОИП. Отд. геол. 2007. № 2. С. 40–56.
Габдуллин Р.Р., Самарин Е.Н., Иванов А.В., Бадулина Н.В., Афонин М.А., Игтисамов Д.В., Фомин Е.Ю., Юрченко А.Ю. Литолого-геохимическая и палеоэкологическая характеристика условий осадконакопления в Горном Крыму в маастрихтском веке // Вестник Московского университета. Сер. 4: Геология. 2015. № 2. С. 39–56.
Гнибиденко З.Н., Лебедева Н.К., Шурыгин Б.Н. Региональный магнитостратиграфический разрез верхнемеловых отложений юга Западной Сибири (Омская впадина) // Докл. АН. 2014. Т. 458. № 1. С. 83–87.
Гричук В.П. Палинологический анализ отложений // Проблемы физической географии. М.–Л.: Изд-во АН СССР, 1940. Т. VIII. С. 53–57.
Гужиков А.Ю. Геологическая информативность магнетизма керна и шлама осадочных пород, полученных при бурении разведочных скважин // Приборы и системы разведочной геофизики. 2013. № 4(46). С. 51–61.
Гужиков А.Ю., Барабошкин Е.Ю., Фомин В.А. Магнитостратиграфическая шкала меловой системы: современное состояние, проблемы построения и перспективы развития // Меловая система России и ближнего зарубежья: проблемы стратиграфии и палеогеографии. Сб. науч. трудов. Ред. Первушов Е.М. Саратов: Изд-во Саратовского университета, 2007. С. 69–86.
Гужиков А.Ю., Барабошкин Е.Ю., Беньямовский В.Н., Вишневская В.С., Копаевич Л.Ф., Первушов Е.М., Гужикова А.А. Новые био- и магнитостратиграфические данные по кампанским–маастрихтским отложениям классического разреза Нижняя Банновка (юг Саратовского Правобережья) // Стратиграфия. Геол. корреляция. 2017. Т. 25. № 1. С. 24–61.
Гужиков А.Ю., Гужикова А.А., Александрова Г.Н., Грищенко В.А., Маникин А.Г., Фомин В.А. Предварительные данные по магнитостратиграфии кампана–маастрихта Юго-Западного Крыма // Меловая система России и ближнего зарубежья: проблемы стратиграфии и палеогеографии. Материалы IX Всероссийского совещания, 17–21 сентября 2018 г., Белгород. Ред. Барабошкин Е.Ю., Липницкая Т.А., Гужиков А.Ю. Белгород: ПОЛИТЕРРА, 2018. С. 113–116.
Гужикова А.А. Первые магнитостратиграфические данные по маастрихту Горного Крыма (Бахчисарайский район) // Изв. Сарат. ун-та. Нов. сер. Сер. Науки о Земле. 2018. Т. 18. Вып. 1. С. 41–49.
Заклинская Е.Д., Найдин Д.П. К палинологической характеристике верхнемаастрихтских отложений Крыма // Бюлл. МОИП. Отд. геол. 1985. Т. 60. Вып. 1. С. 80–87.
Йолкичев Н.А., Найдин Д.П. Верхний мел Северной Болгарии, Крыма и Мангышлака. Статья 2. Стратиграфия верхнего мела юго-западной части Горного Крыма // Бюлл. МОИП. Отд. геол. 1999. Т. 74. Вып. 5. С. 48–59.
Лебедева Н.К. Биостратиграфия верхнемеловых отложений в бассейне р. Уса (Полярное Предуралье) по диноцистам // Стратиграфия. Геол. корреляция. 2005. Т. 13. № 3. С. 114–131.
Лебедева Н.К., Кузьмина О.Б. Палиностратиграфия верхнемеловых и палеогеновых отложений юга Западной Сибири на примере скважин Русско-Полянского района, Омский прогиб // Стратиграфия. Геол. корреляция. 2018. Т. 26. № 1. С. 85–114.
Лебедева Н.К., Александрова Г.Н., Шурыгин Б.Н., Овечкина М.Н., Гнибиденко З.Н. Палеонтологическая и магнитостратиграфическая характеристика верхнемеловых отложений, вскрытых скважиной 8 Русско-Полянского района (юг Западной Сибири) // Стратиграфия. Геол. корреляция. 2013. Т. 21. № 1. С. 43–73.
Лопатин А.В., Аверьянов А.О. О находках динозавров в Крыму // Изв. ВУЗов. Геол. и разведка. 2019. № 1. С. 67–71.
Маслакова Н.И., Липник E.C. Экскурсия “D” // Путеводитель экскурсий. XII Европейский микропалеонтологический коллоквиум. Часть 1. Крым. М.: Ротапринт Гос. библ. им. В.И. Ленина, 1971. С. 95–113.
Молостовский Э.А., Храмов А.Н. Магнитостратиграфия и ее значение в геологии. Саратов: Изд-во Саратовского университета, 1997. 180 с.
Найдин Д.П. Подкласс Endocochlia. Внутреннераковинные // Атлас верхнемеловой фауны Северного Кавказа и Крыма. Ред. Москвин М.М. М.: Гостоптехиздат, 1959. С. 198–209
Найдин Д.П. Позднемаастрихтские белемнителлиды Евразии // Развитие и смена органического мира на рубеже мезозоя и кайнозоя. Новые данные о развитии фауны. Ред. Шиманский В.Н., Соловьев А.Н. М.: Наука, 1975. С. 91–108.
Рябинин А.Н. Новые находки ископаемых рептилий в Крыму // Природа. 1946. № 11. С. 65–66.
Стратиграфический кодекс России. Издание третье, исправленное и дополненное. СПб.: Изд-во ВСЕГЕИ, 2019. 96 с.
Суринский А.М., Гужиков А.Ю. Циклостратиграфический анализ петромагнитных данных по маастрихту долины р. Бодрак (Юго-Западный Крым) // Изв. Саратовского ун-та. Нов. сер. Сер. Науки о Земле. 2019. Т. 19. Вып. 3. С. 206–211.
Яковишина Е.В., Копаевич Л.Ф., Беньямовский В.Н., Соколова Е.А. Генетические типы верхнемаастрихтских отложений Горного Крыма // Вестник Московского ун-та. Сер. 4. Геология. 2008. № 3. С. 11–23.
Alekseev A.S., Kopaevich L.F. Foraminiferal biostratigraphy of the uppermost Campanian-Maastrichtian in SW Crimea (Bakhchisaray and Chakhmakhly sections) // Bulletin de l’Institut Royal des Sciences Naturelles de Belgique Sciences de la Terre. 1997. V. 67. P. 103–118.
Alekseev A.S., Kopaevich L.F., Ovechkina M.N., Olferiev A.G. Maastrichtian and Lower Palaeocene of Northern Saratov Region (Russian Platform, Volga River): Foraminifera and calcareous nannoplankton // Bulletin de l’Institut Royal des Sciences Naturelles de Belqique, Sciences de la Terre. 1999. V. 69. Supplement-A. P. 15–45.
Antonescu E., Foucher J.-C., Odin G.S., Schiøler P., Siegl-Farkas A., Wilson G.J. Dinoflagellate cysts in the Campanian-Maastrichtian succession of Tercis-les Bains (Landes, France), a synthesis // The Campanian-Maastrichtian Stage Boundary: characterisation at Tercis les Bains (France): correlation with Europe and other continents. Ed. Odin G.S. IUGS Spec. Publ. (monograph) Ser. 36; Developments in Palaeontology and Stratigraphy Ser. 19. Amsterdam: Elsevier Science Publ., 2001. P. 253–264.
Aurisano R.W. Upper Cretaceous dinoflagellate biostratigraphy of the subsurface Atlantic coastal plain of New Jersey and Delaware, U.S.A. // Palynology. 1989. V. 13. № 1. P. 143–179. https://doi.org/10.1080/01916122.1989.9989359
Baraboshkin E.Yu., Benyamovski V.N., Guzhikov A., Aleksandrova G., Pervushov E., Selzer V.B., Ovechkina M.N., Kaljakin E.A., Kopaevich L., Vishnevskaya V.S., Guzhikova A., Pokrovsky B., Baraboshkin E.E., Iakovishina E.V. Integrated study of Campanian/Maastrichtian boundary interval at Volga region (Russia) and Aktolagay Plateau (West Kazakhstan) of the Russian Platform // Berichte der Geologischen Bundesanstalt. 2017. V. 120. P. 22.
Besse J., Courtillot V. Apparent and true polar wander and the geometry of the geomagnetic field over the last 200 Myr // J. Geophys. Res. 2002. V. 107. № 11. P. 1–31.
Birkelund T. Upper Cretaceous belemnites of Denmark // Biologiske Skrifter, Det Kongelige Danske Yidenskabernes Selskab. 1957. V. 9. P. 1–69.
Brinkhuis H. Late Eocene to Early Oligocene dinoflagellate cysts from the Priabonian type-area (Northeast Italy): biostratigraphy and palaeoenvironmental interpretation // Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994. V. 107. P. 121–163.
Brinkhuis H., Schiøler P. Palynology of the Geulhemmerberg Cretaceous-Tertiary boundary section (Limburg, SE Netherlands) // Geol. Mijnbouw. 1996. V. 75. P. 193–213.
Burnett J. Upper Cretaceous // Calcareous Nannofossil Biostratigraphy. Ed. Bown P.R. London: Chapman and Hall/Kluwer Academic Publ., 1998. P. 132–199.
Chaudhuri S.K., Lack J.G., Coates J.D. Biogenic Magnetite Formation through 11 Anaerobic Biooxidation of Fe(II) // Applied and Environmental Microbiology. 2001. V. 67. № 6. P. 2844–2848.
Christensen W.K. The Late Cretaceous belemnite family Belemnitellidae. Taxonomy and evolutionary history // Bull. Geol. Soc. Denmark. 1997. № 44. P. 59–88.
Christensen W.K. Upper Campanian and Lower Maastrichtian belemnites from the Mons Bassin, Belgium // Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la terre. 1999. V. 69. P. 97–131.
Coccioni R., Premoli-Silva I. Revised Upper Albian–Maastrichtian planktonic foraminiferal biostratigraphy and magneto-stratigraphy of the classical Tethyan Gubbio section (Italy) // Newsl. Stratigr. 2015. V. 48. № 1. P. 47–90.
Corradini D. Non-calcareous microplankton from the Upper Cretaceous of the northern Apennines // Bollettino della Società paleontologica italiana. 1973. V. 11. № 2. P. 119–197.
Crouch E.M., Brinkhuis H. Environmental change across the Paleocene-Eocene transition from eastern New Zealand: a marine palynological approach // Marine Micropaleontol. 2005. V. 56. P. 138–160.
Debiche M.G., Watson G.S. Confidence limits and bias correction for estimating angles between directions with applications to paleomagnetism // J. Geophys. Res. 1995. V. 100. № B12. P. 24405–24430.
Dunlop D. Theory and application of the Day plot (Mrs/Ms versus Hcr/Hc) 1. Theoretical curves and tests using titanomagnetite data // J. Geophys. Res. 2002. V. 107. № B3. https://doi.org/10.1029/2001JB000487
Epstein S., Buchsbaum R., Lowenstam H. A., Urey H. C. Revised carbonate-water isotopic temperature scale // Geol. Soc. Am. Bull. 1953. V. 64. № 11. P. 1315–1326.
Evans M.E., Heller F. Environmental magnetism. Principles and applications of enviromagnetics. San Diego: Academic Press, 2003. 311 p.
Flügel E. Microfacies Analysis of Limestones. Analysis, Interpretation, and Application. Berlin: Springer, 2010. 894 p.
Gradstein F., Ogg J. G., Schmitz M. D., Ogg G. M. The Geologic Time Scale. Amsterdam: Elsevier, 2012. 1144 p.
Guzhikov A.Y., Guzhikova A.A., Manikin A.G., Grishchenko V.A. Magnetostratigraphy of the Maastrichtian from Volga Right Bank Area Near Saratov (Lower Volga Region) // Problems of Geocosmos–2018. Eds. Yanovskaya T., Kosterov A., Bobrov N., Divin A., Saraev A., Zolotova N. Springer Proceedings in Earth and Environmental Sciences. Springer, Cham., 2020. P. 83–105.
Hansen J.M. Dinoflagellate stratigraphy and echinoid distribution in Upper Maastrichtian and Danian deposits from Denmark // Bull. Geol. Soc. Den. 1977. V. 26. P. 1–26.
Hansen J.M. A new dinoflagellate zone at the Maastrichtian/Danian Boundary in Denmark // Danmarks Geologiske Undersøgelse Arbog. 1979. P. 131–140.
Hoek R.P., Eshet Y., Almogi-Labin A. Dinoflagellate Cyst Zonation of Campanian-Maastrichtian Sequences in Israel // Micropaleontology. 1996. V. 42. № 2. P. 125–150.
Hultberg S.U., Malmgren B.A. Quantitative Biostratigraphy Based on Late Maastrichtian Dinoflagellates and Planktonic Foraminifera from Southern Scandinavia // Cretaceous Res. 1987. V. 8. P. 211–228.
Iakovleva A.I., Brinkhuis H., Cavagnetto C. Late Palaeocene–Early Eocene dinoflagellate cysts from the Turgay Strait, Kazakhstan: correlations across ancient seaways // Palaeogeogr. Palaeoclimat. Palaeoecol. 2001. V. 172. P. 43–268.
Jagt J.W.M., Felder W.M. The stratigraphic range of the index ammonite Pachydiscus neubergicus (von Hauer, 1858) in the type area of the Maastrichtian Stage // Netherlands J. Geosciences/Geologie en Mijnbouw. 2003. V. 82. № 3. P. 261–268.
Jeletzky J.A. Die Stratigraphie und Belemnitenfauna des Obercampan und Maastricht Westfalens, Nordwestdeutschlands und Danemarks, sowie einige allgemeine Gliederungsprobleme der jüngeren borealen Oberkreide Eurasiens // Beihefte Geologisches Jahrbuch. 1951. V. 1. P. 1–142.
Jurkowska A., Uchman A. The trace fossil Lepidenteron lewesiensis (Mantell, 1822) from the Upper Cretaceous of southern Poland // Acta Geol. Polon. 2013. V. 63. № 4. P. 611–623.
Kennedy W.J. The ammonite fauna of the type Maastrichtian with a revision of Ammonites colligatus Binkhorst, 1861 // Bulletin de I’lnsritut Royal des Sciences Naturelles de Belgique 56. Sciences de la Terre 56. 1987. P. 151–267.
Kennedy W.J., Summesberger H. Upper Campanian ammonites from the Gschliefgraben (Ultrahelvetic, Upper Austria) // Beitraege zur Paläontologie von Öesrerreich. 1984. V. 11. P. 149–206.
Kennedy W.J., Summesberger H. Lower Maastrichtian ammonites from Neuberg, Steiermark, Austria // Beitraege zur Paläontologie von Öesrerreich. 1986. V. 12. P. 181–242.
Kennedy W.J., Summesberger H. Lower Maastrichtian ammonites from Nagoryany, Ukrainian SSR // Beitraege zur Paläontologie von Oesrerreich. 1987. V. 13. P. 25–78.
Keutgen N., Remin Z., Jagt J.W.M. The late Maastrichtian Belemnella kazimiroviensis group (Cephalopoda, Coleoidea) in the Middle Vistula valley (Poland) and the Maastricht area (the Netherlands, Belgium) – taxonomy and palaeobiological implications // Palaeontologia Electronica. 2017. V. 20. № 2.38A. P. 1–29.
Kirsch K.-H. Dinoflagellatenzysten aus der Oberkreide des Helvetikums und Nordultrahelvetikums von Oberbayern // Münchner Geowissenschaftliche Abhandlungen, A. Geologie und Paläontologie. 1991. V. 22. P. 1–306.
Klinger H.C., Kennedy W.J. Stratigraphic and geographic distribution, phylogenetic trends and general comments on the ammonite family Baculitidae Gill, 1871 (with an annotated list of species referred to the family) // Annals of the South African Museum. 2001. V. 107. Pt. 1. 290 p.
Klinger H.C., Kennedy W.J. Observations on the systematics, geographic and stratigraphic distribution, and origin of Diplomoceras cylindraceum (Defrance, 1816) (Cephalopoda: Ammonoidea) // Annals of the South African Museum. 2003. V. 110. Pt. 4. P. 171–198.
Knaust D. Atlas of Trace Fossils in Well Core: Appearance, Taxonomy and Interpretation. Springer Int. Publ., 2017. 209 p.
Knaust D., Bromley R.G. (Eds.). Trace fossils as indicators of sedimentary environments // Developments in Sedimentology. Amsterdam: Elsevier, 2012. V. 64. 960 p.
Koch W. Biostratigraphie in der Oberkreide und Taxonomie von Foraminiferen // Geol. Jahrb. A. 1977. V. 38. P. 11–123.
Lebedeva N.K. Dinocyst Biostratigraphy of the Upper Cretaceous of Northern Siberia // Paleontol. J. 2006. V. 40. № 5. P. S604–S621.
Machalski M. Stratigraphically important ammonites from the Campanian–Maastrichtian boundary interval of the Middle Vistula River section, central Poland // Acta Geol. Polon. 2012. V. 62. № 1. P. 91–116.
Marheinecke U. Monographie der Dinozysten, Acritarcha und Chlorophyta des Maastrichtium von Hemmoor (Niedersachsen) // Palaeontographica. 1992. B. 227. P. 1–173.
Niebuhr B. Late Campanian and Early Maastrichtian ammonites from the white chalk of Kronsmoor (northern Germany) – taxonomy and stratigraphy // Acta Geol. Polon. 2003. V. 53. № 4. P. 257–281.
Niebuhr B., Hampton M.J., Gallagher L.T., Remin Z. Integrated stratigraphy of the Kronsmoor section (northern Germany), a reference point for the base of the Maastrichtian in the Boreal Realm // Acta Geol. Polon. 2011. V. 61. № 2. P. 193–214.
Nøhr-Hansen H. Upper Cretaceous dinoflagellate cyst stratigraphy, onshore West Greenland // Bull. Grønl. Geol. Unders. 1996. V. 170. P. 1–104.
Nøhr-Hansen H. Palynostratigraphy of the Cretaceous–lower Palaeogene sedimentary succession in the Kangerlussuaq Basin, southern East Greenland // Rev. Palaeobot. Palynol. 2012. V. 178. P. 59–90.
Odin G.S., Lamaurelle M.A. The global Campanian–Maastrichtian Stage boundary // Episodes. 2001. V. 24. № 4. P. 229–238.
Odin G.S., Hancock J.M., Antonescu E., Bonnemaison M., Caron M., Cobban W.A., Dhondt A.V., Gaspard D., Ion J., Jagt J.W.M., Kennedy W.J., Melinte M., Neraudeau D., von Salis K.A., Ward P.D. Definition of a Global Boundary Stratotype Section and Point for the Campanian/Maastrichtian boundary // Bull. de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre. 1996. V. 66 (Suppl.). P. 111–117.
Ogg J.G., Ogg G.M., Gradstein F.M. A Concise Geologic Time Scale. Elsevier, 2016. 242 p.
Perch-Nielsen K. Mesozoic calcareous nannofossils // Plankton Stratigraphy. Eds. Bolli H.S. Cambridge University Press, 1985. P. 329–426.
Prauss M. Availability of reduced nitrogen chemospecies in photic-zone waters as the ultimate cause of fossil prasinophyte prosperity // Palaios. 2007. V. 22. P. 489–499.
Pross J., Brinkhuis H. Organic-walled dinoflagellate cysts as paleoenvironmental indicators in the Paleogene; a synopsis of concepts // Palaontologische Zeitschrift. 2005. V. 79. P. 53–59.
Radmacher W., Perez-Rodríguez I., Arz J.A., Pearce M.A. Dinoflagellate biostratigraphy at the Campanian–Maastrichtian boundary in Zumaia, northern Spain // Cretaceous Res. 2014. V. 51. P. 309–320.
Radmacher W., Mangerud G., Tyszka J. Dinoflagellate cyst biostratigraphy of Upper Cretaceous strata from two wells in the Norwegian Sea // Rev. Palaeobot. Palynol. 2015. V. 216. P. 18–32.
Remin Z. Understanding coleoid migration patterns between eastern and western Europe – belemnite faunas from the upper lower Maastrichtian of Hrebenne, southeast Poland // Cretaceous Res. 2018. V. 87. P. 368–384.
Robaszynski F., Caron M., Gonzales J.M., Wonders A. Atlas of Late Cretaceous planktonic foraminifera // Rev. micropalèontol. 1984. V. 26. № 3–4. P. 145–305.
Robaszynski F., Bless M.J.M., Felder P.J.S., Foucher J.-C., Legoux O., Maniytt H., Meessen J.P.M.Th., Van der Tuuk L.A. The Campanian–Maastrichtian Boundary in the chalky facies close to the type-Maastrichtian area // Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine. 1985. V. 9 (1). P. 1–113.
Roncaglia L. Lower Maastrichtian dinoflagellates from the Viano Clay Formation at Viano, northern Apennines, Italy // Cretaceous Res. 2002. V. 23. P. 65–76.
Roncaglia L., Corradini D. Upper Campanian to Maastrichtian dinoflagellate zonation in the northern Apennines, Italy // Newsletters on Stratigraphy. 1997a. V. 35. P. 29–57.
Roncaglia L., Corradini D. Correlation of key dinoflagellate events with calcareous nannoplankton and planktonic foraminiferal zones in the Solignano Formation (Maastrichtian, Late Cretaceous) northern Apennines, Italy // Rev. Palaeobot. Palynol. 1997b. V. 97. P. 177–196.
Schiøler P., Wilson G.J. Maastrichtian dinoflagellate zonation in the Dan Field, Danish North Sea // Rev. Palaeobot. Palynol. 1993. V. 78. P. 321–351.
Schiøler P., Wilson G.J. Dinoflagellate biostratigraphy around the Campanian–Maastrichtian boundary at its type section (Tercis Quarry, southwest France) // The Campanian–Maastrichtian stage boundary: characterisation at Tercis les Bains (France): correlation with Europe and other Continents. Ed. Odin G.S. IUGS Spec. Publ. (monograph) Ser., 36; Developments in Palaeontology and Stratigraphy Ser., 19. Amsterdam: Elsevier Science Publ., 2001. P. 221–234.
Schiøler P., Brinkhuis H., Roncaglia L., Wilson G.J. Dinoflagellate biostratigraphy and sequence stratigraphy of the type Maastrichtien (Upper Cretaceous), ENCI Quarry, the Netherland // Marine Micropaleontology. 1997. V. 31. P. 65–95.
Schulz M.-G. Morphometrisch-variationsstatistische Untersuchungen zur Phylogenie der Belemniten-Gattung Belemnella im Untermaastricht NW-Europas. (Die Maastricht-Stufe in NW-Deutschland, Teil 1) // Geologisches Jahrbuch. Reihe A. 1979. V. 47. P. 3–157.
Schulz M.-G., Schmid F. Das Ober-Maastricht von Hemmoor (N-Deutschland): Faunezonen-Gliederung und Korrelsation mit dem Ober-Maastricht von Dänemark und Limburg // Newsl. Stratigr. 1983. V. 13. P. 21–39.
Slimani H. Nouvelle zonation aux kystes de dinoflagellés du Campanien au Danien dans le nord et l’est de la Belgique et dans le sud-est des Pays-Bas // Mem. Geol. Surv. Belgium. 2000. V. 46. P. 1–88.
Slimani H. Les kystes de dinoflagellés du Campanien au Danien dans la région de Maastricht (Belgique et Pays-Bas) et de Turnhout (Belgique): biozonation et corrélation avec d’autres régions en Europe occidentale // Geologica et Palaeontologica. 2001. V. 35. P. 161–201.
Slimani H., Louwye S., Dusar M., Lagrou D. Connecting the Chalk Group of the Campine Basin to the dinoflagellate cyst biostratigraphy of the Campanian to Danian in borehole Meer (northern Belgium) // Geologie en Mijnbouw/Netherlands Journal of Geosciences. 2011. V. 90. № 2–3. P. 129–164.
Slimani H., Guédé K.É., Williams G.L., Asebriy L., Ahmamou M’Fedal. Campanian to Eocene dinoflagellate cyst biostratigraphy from the Tahar and Sekada sections at Arba Ayacha, western External Rif, Morocco // Rev. Palaeobot. Palynol. 2016. V. 228. P. 26–46.
Sluijs A., Pross J., Brinkhuis H. From greenhouse to icehouse; organic-walled dinoflagellate cysts as paleoenvironmental indicators in the Paleogene // Earth Sci. Rev. 2005. V. 68. P. 281–315.
Sluijs A., Brinkhuis H., Crouch E.M., John C.M., Handley L., Munsterman D., Bohaty S.M., Zachos J.C., Reichart G.-J., Schouten S., Pancost R.D., Sinninghe Damste J.S., Welters N.L.D., Lotter A.F., Dickens G.R. Eustatic variations during the Paleocene–Eocene greenhouse world // Paleoceanography. 2008. V. 23. PA4216. DOI: 42https://doi.org/10.1029/2008PA001615
Surlyk F., Rasmussen S.L., Boussaha M., Schiøler P., Schovsbo N.H., Sheldon E., Stemmerik L., Thibault N. Upper Campanian–Maastrichtian holostratigraphy of the eastern Danish Basin // Cretaceous Res. 2013. V. 46. P. 232–256.
Thibault N. Calcareous nannofossil biostratigraphy and turnover dynamics in the late Campanian–Maastrichtian of the tropical South Atlantic // Rev. Micropaléontol. 2016. V. 59. № 1. P. 57–69.
Thibault N., Gardin S. The late Maastrichtian nannofossil record of climate change in the South Atlantic DSDP Hole 525A // Marine Micropaleontology. 2007. V. 65. P. 163–184.
Thibault N., Harlou R., Schovsbo N., Schiøler P., Minoletti F., Galbrun B., Lauridsen B.W., Sheldon E., Stemmerik L., Surlyk F. Upper Campanian–Maastrichtian nannofossil biostratigraphy and high-resolution carbon-isotope stratigraphy of the Danish Basin: towards a standard δ13C curve fot the Boreal Realm // Cretaceous Res. 2012a. V. 33. P. 72–90.
Thibault N., Husson D., Harlou R., Gardin S., Galbrun B., Huret E., Minoletti F. Astronomical calibration of upper Campanian–Maastrichtian carbon isotope events and calcareous plankton biostratigraphy in the Indian Ocean (ODP Hole 762C): Implication for the age of the Campanian–Maastrichtian boundary // Palaeogeogr. Palaeoclimat. Palaeoecol. 2012b. V. 337–338. P. 52–71.
Voigt S., Gale A., Jung C., Jenkyns H. Global correlation of Upper Campanian–Maastrichtian successions using carbon isotope stratigraphy: development of a new Maastrichtian timescale // Newsl. Stratigr. 2012. V. 45. P. 25–53.
Wiedmann J., Schmidt W. Upper Cretaceous ammonites from Jamaica and their stratigraphic and paleogeographic implications // Biostratigraphy of Jamaica. Eds. Wright R.M., Robinson E. Geol. Soc. Am. Mem. 1993. № 182. P. 77–92.
Williams G.L., Brinkhuis H., Pearce M.A., Fensome R.A., Weegink J.W. Southern ocean and global dinoflagellate cyst events compared: index events for the Late Cretaceous–Neogene // Proceedings of the Ocean Drilling Program, Scientific Results. 2004. V. 189. P. 1–98.
Zaklinskaya E.D. Phylogeny and classification of the Normapolles // Rev. Palaeobot. Palynol. 1981. V. 33. P. 139–147.
Дополнительные материалы
- скачать ESM.tiff
- Приложение 1. Распределение палиноморф в разрезе Бешкош.
Инструменты
Стратиграфия. Геологическая корреляция