

Стратиграфия. Геологическая корреляция, 2022, T. 30, № 3, стр. 101-116
Диатомовые водоросли и силикофлагеллаты раннего эоцена Преддонецкой моноклинали (Русская плита): биостратиграфические и палеогеографические аспекты
Геологический институт РАН
Москва, Россия
* E-mail: tanya.oreshkina@gmail.com
Поступила в редакцию 27.10.2021
После доработки 08.12.2021
Принята к публикации 22.12.2021
- EDN: RRQSSJ
- DOI: 10.31857/S0869592X22030036
Аннотация
Комплекс диатомей вёшенской свиты, изученный в керне скв. Р-321 Преддонецкой моноклинали (юг Русской плиты), отнесен к зоне Moisseevia uralensis нижнего ипра. По имеющимся данным зона Moisseevia uralensis соответствует зонам по диноцистам Deflandrea oebisfeldensis и Dracodinium (=Wetzeliella) astra и, соответственно, зоне NP10 по наннопланктону. Присутствие зонального вида силикофлагеллат Naviculopsis foliacea подтверждает раннеэоценовый возраст вёшенской свиты. Таксономическое сходство диатомовых водорослей и силикофлагеллат рассматриваемого региона с ассоциациями бассейнов Северной Европы предполагает устойчивую связь этих акваторий в начале раннего эоцена.
ВВЕДЕНИЕ
Особенностью палеогеновых трансгрессий на эпиконтинентальных пространствах Северного Перитетиса является присутствие терригенно-кремнистых фаций в краевых частях морских бассейнов (Radionova et al., 2003). Одним из широко распространенных стратиграфических интервалов, обогащенных остатками кремневого планктона, является переход от палеоцена к эоцену. В этом интервале последовательности ассоциаций диатомей и силикофлагеллат прослежены в Среднем Поволжье (Стрельникова, 1992; Глезер, 1995; Oreshkina, Oberhansli, 2003; Орешкина, Александрова, 2007, 2017; Oreshkina, Radionova, 2014; Aleksandrova, Radionova, 2006), Воронежской антеклизе (Стрельникова, 1992), в Калининградской области (Стрельникова и др., 1978), Полярном Предуралье (Орешкина и др., 1998; Oreshkina, 2000, 2012; Iakovleva et al., 2000) Зауралье (Oreshkina et al., 2004; Орешкина и др., 2008; Александрова и др., 2012) и Тургайском прогибе (Radionova et al., 2001). На юге Русской плиты в Преддонецкой структурно-фациальной зоне (СФЗ) восточного обрамления Донбасса (Постановления…, 2001; Ахметьев, Беньямовский, 2003) верхнепалеоценовый–нижнеэоценовый цикл осадконакопления включает бузиновские, вёшенские, суровикинские и осиновские слои (Беньямовский, 2016; Александрова и др., 2020), выделенные Г.П. Леоновым (1961) и затем переведенные в ранг свит (за исключением осиновских слоев) В.П. Семеновым (1965). Бузиновская свита представлена в базальной части фосфорито-галечниковыми отложениями с окатанными ядрами моллюсков, которые перекрываются толщей зеленых глауконитовых песков (мощность до 20 м). Вёшенская свита сложена песчаными опоковидными или трепеловидными глинами (мощность до 16 м), фациально замещающимися неоднородными глинистыми рыхлыми кварцево-глауконитовыми песками, иногда с прослоями и линзами кремнистых песчаников. Суровикинская свита, местами отделенная от нижележащих отложений слоем плотного грубозернистого песчаника с галькой кремнистой глины, представлена светлыми мелкозернистыми кварцевыми слабоглауконитовыми песками с прослоями конкреционных кремнистых и сливных песчаников (мощность до 25 м). К осиновским слоям, с размывом перекрывающим нижележащие толщи, отнесена однообразная толща кварцевых песков и песчаников мощностью до 25 м.
Комплексное литолого-биостратиграфическое изучение этого стратиграфического интервала в опорных разрезах Преддонецкой СФЗ не выявило присутствия микрофоссилий в большей части опробованных разрезов. Исключением является скв. 1238, пробуренная около станицы Базковская (Шолоховский район Ростовской области), где были обнаружены комплексы диноцист танета–нижнего ипра (Александрова и др., 2020; Яковлева, Александрова, 2021). Силикофоссилии в скв. 1238 обнаружены не были. Противоположная ситуация выявлена в скв. Р-321 той же структурно-фациальной зоны, где палиноморфы отсутствуют, а из бузиновской и вёшенской свит выделен представительный комплекс диатомовых и силикофлагеллат. Цель настоящей статьи – рассмотреть таксономический состав и стратиграфическое положение этого комплекса, провести сравнение с другими местонахождениями силикофоссилий окраинных бассейнов Северного Перитетиса.
МАТЕРИАЛ И МЕТОДЫ
Скважина Р-321 пробурена в 2.2 км западнее пос. Григорьевский Урюпинского района Волгоградской области (50°52′ с.ш., 41°37′ в.д.) с абс. высотой устья скважины 185 м (рис. 1). Образцы керна и описание разреза скважины были предоставлены нам сотрудником Южно-Воронежской геологической партии В.М. Рыбораком, под руководством которого проходили работы по геологическому, гидрогеологическому и инженерно-геологическому доизучению масштаба 1 : 200 000 на площади листа М-37-XII (Новохоперск) (Рыборак, Шокуров, 1999). Скважиной (рис. 2) вскрыты (снизу вверх):
Рис. 1.
(а) Палеогеографическая схема Северной Евразии (по Палеогеографический…, 1998) для интервала перехода от палеоцена к эоцену и (б) местоположение скв. Р-321 (настоящая работа) и других разрезов, упоминаемых в тексте.

Рис. 2.
Распределение реперных видов диатомей зоны Moisseevia uralensis в разрезе скв. Р-321 и соотношение доминирующих групп диатомовых водорослей. 1 – пески; 2 – песчаники; 3 – алевритистые глины; 4 – “бурый железняк”; 5 – конгломерат.

Верхний мел (даний?).
Слой 1, инт. 18.45–15.95 м. Песок ржаво-желтый, мелкозернистый, алевритистый, полевошпатово-кварцевый.
Бузиновская свита.
Слой 2, инт. 15.95–15.45 м. “Бурый железняк”.
Слой 3, инт. 15.45–15.3 м. Конгломерат.
Слой 4, инт. 15.3–14.8 м. Песчаник опоковидный.
Вёшенская свита.
Слой 5, инт. 14.8–6.6 м. Глина зеленовато-серая, алевритистая.
Обоянская свита.
Слои 6–13, инт. 6.6–0.6 м. Пески и песчаники мелко-среднезернистые, кварцевые.
Пробы были обработаны с использованием стандартной процедуры выделения биокремнистых микрофоссилий и органикостенных палиноморф, принятой в лабораториях палеофлористики и микропалеонтологии ГИН РАН (Александрова и др., 2012). Морские палиноморфы в изученных образцах не обнаружены. Полуколичественная оценка общего содержания диатомей и отдельных видов проводилась по балльной системе при увеличении ×400: A (Abundant, в массе) = 1 или более экземпляров в каждом поле зрения (ПЗ); C (Common, часто) = 1 экземпляр в 2 ПЗ; F (Few, нередко) = 1–2 экземпляра в каждом ряду покровного стекла; R (Rare, единично) = несколько экземпляров в препарате; B (Barren) = полное отсутствие диатомей или их фрагментов; T (Trace) = = редкие фрагменты створок. Сохранность диатомей оценивается как G (Good, хорошая) = минимальное количество обломков; M (moderate, средняя) = примерно равное количество обломков и целых створок; P (poor, плохая) = целые створки встречаются редко. Микросъемка проводилась в световом микроскопе Motic BA310 и сканирующем микроскопе Vega ©Tescan MV-2300. При таксономической идентификации диатомей были учтены изменения в названии родов, признанных преоккупированными (Blanco, Wetzel, 2016), и другие таксономические новации. Для биостратиграфического расчленения используется модифицированная зональная шкала по диатомовым водорослям палеогена внетропической области Северного полушария (Стрельникова, 1992), сопоставленная с зонами по диноцистам (Oreshkina, 2000, 2012; Oreshkina et al., 2004; Орешкина, Александрова, 2007, 2017; Iakovleva et al., 2000). Стратиграфическое распределение и таксономический состав диатомовых водорослей и силикофлагеллат, а также их изображения приведены на рис. 2, в табл. 1, табл. I–III .
Таблица 1.
Распределение диатомей и силикофлагеллат в скв. Р-321
| № образцов | 3 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
|---|---|---|---|---|---|---|---|---|---|
| Обилие | A | A | F | A | F | A | F | A | F |
| Сохранность | G | G | G | G | G | G | G | G | G |
| Диатомовые водоросли | |||||||||
| Actinoptychus pericavatus Brun | F | F | F | C | C | C | F | F | |
| Actinoptychus seductilis A. Schmidt | F | F | F | C | F | C | F | F | F |
| Actinoptychus simbirskiana A. Schmidt | F | F | F | F | C | C | F | F | |
| Anuloplicata concentrica (A. Schmidt) Gleser | C | C | C | C | F | C | C | C | F |
| Anuloplicata ornata (Grunow) Gleser | C | C | C | C | F | C | C | C | F |
| Biddulphia tuomeyi (Bailey) Roper | R | R | R | R | R | ||||
| Chaetoceros sp. 1 sensu Homann, 1991, pl. 9, fig. 3, 4 | R | ||||||||
| Coscinodiscus argus Ehrenberg | F | F | F | C | R | A | C | C | R |
| Costopyxis broschii (Grunow) Strelnikova et Nikolaev | F | F | F | F | F | F | F | R | |
| Costopyxis reticulata (Long, Fuge et Smith) Gleser | R | F | R | ||||||
| Cymatosira spp. | R | R | |||||||
| Diploneis sp. | R | ||||||||
| Eunotogramma variabile Grunow | R | F | R | F | F | F | |||
| Eupyxidicula moelleri (Grunow) Blanco et Wetzel | C | F | C | A | R | A | A | A | F |
| Eupyxidicula turris (Greville et Arnott) Blanco et Wetzel | F | F | F | ||||||
| Goniothecium wittianus Pantocsek | R | R | R | R | R | ||||
| Grunowiella gemmata(Grunow) Van Hearck | F | F | F | F | F | ||||
| Hemiaulus arcticus var. bornholmensis Cleve-Euler | R | R | F | ||||||
| Hemialus febriatus (Grunow) Fenner | F | F | F | F | F | ||||
| Hemialus frigidus (Grunow) Fenner | F | F | F | R | |||||
| Hemiaulus aff. incurvus Shibkova | R | ||||||||
| Hemiaulus curvatulus Strelnikova | R | R | |||||||
| Hemiaulus morsianus (Grunow) Fenner | R | R | R | F | F | F | |||
| Hyalodiscus radiatus (O’Meara) Grunow | F | R | F | F | R | C | F | C | |
| Jousea elliptica (Jousé) Gleser | F | R | |||||||
| Moisseevia probabilis (A. Schmidt) Strelnikova | R | R | R | ||||||
| Moisseevia uralensis (Jousé) Strelnikova | R | R | R | F | |||||
| Paralia crenulata (Grunow) Gleser | F | R | F | ||||||
| Paralia grunowii Gleser | A | C | A | A | F | A | A | A | F |
| Paralia selecta (A. Schmidt) Gleser | R | ||||||||
| Porotheca danica (Grunow) Fenner | R | ||||||||
| Proboscia sp. 1 = Rhizosolenia clavigera sensu Fenner, 1994 pl. 9, fig. 3, 4 | F | А | |||||||
| Pseudopodosira hyalina Jousé | R | R | R | C | |||||
| Pseudopodosira pileiformis Jousé | R | R | F | F | F | C | C | F | |
| Pseudopodisira westii (W. Smith) Sheshukova et Gleser | R | R | R | R | F | F | F | ||
| Pterotheca spada Tempère et Peragalli | R | R | |||||||
| Radiaplicata clavigera (Grunow) Gleser | F | F | F | F | |||||
| Rattrayella sp. | R | R | |||||||
| Rhaphoneis lancetulla Grunow | F | F | F | F | |||||
| Sheshukovia fenestra (Witt) Fenner | F | R | |||||||
| Sheshukovia flos (Ehrenberg) Fenner | R | R | F | F | F | F | |||
| Soleum exsculptum Heiberg | F | F | |||||||
| Stellarima microtrias (Ehrenberg) Hasle et Sims | R | R | R | R | R | R | |||
| Stephanogonia polygona Ehrenberg | R | ||||||||
| Trinacria excavata Heiberg | F | R | R | R | |||||
| Trinacria subcoronata Sheshukova et Gleser | R | ||||||||
| Силикофлагеллаты | |||||||||
| Corbisema gleserae Bukry | R | R | |||||||
| Corbisema hastata globulata Bukry | R | R | F | F | F | ||||
| Dictyocha brevispina (Lemmermann) Bukry | R | R | R | ||||||
| Dictyocha deflandrei Frenguelli ex Glezer | R | F | F | F | F | ||||
| Dictyocha precarentis Bukry | F | F | |||||||
| Naviculopsis eobiapiculata Bukry | R | ||||||||
| Naviculopsis foliacea Deflandre | R | R | |||||||
| Naviculopsis minor (Schulz) Frenguelli | R | R | R |
Таблица I . Диатомеи из палеогеновых отложений скв. Р-321. Длина масштабной линейки 20 мкм для фиг. 1–3 и 10 мкм для фиг. 4–24 . 1, 2, 5 – Eupyxidicula moelleri (Grunow) Blanco et Wetzel: 1, 2 – обр. 14; 5 – обр. 10; 3 – Costopyxis broschii (Grunow) Strelnikova et Nikolaev, обр. 14; 4 – Moisseevia uralensis (Jousé) Strelnikova, обр. 10; 6 – Hemialus febriatus (Grunow) Fenner, обр. 10; 7 – Hemiaulus aff. incurvus Schibkova, обр. 3; 8 – Hemiaulus frigidus (Grunow) Fenner, обр. 14; 9 – Hemiaulus sp., обр. 14; 10, 11 – Hemiaulus morsianus (Grunow) Fenner: 10 – обр. 14; 11 – обр. 10; 12 – Hemiaulus curvatulus Strelnikova, обр. 14; 13 – Hemiaulus arcticus var. bornholmensis Cleve-Euler, обр. 12; 14, 23, 24 – Coscinodiscus argus Ehrenberg, обр. 10; 15 – Grunowiella sp., обр. 13; 16 – Grunowiella gemmata (Grunow) Van Hearck, обр. 14; 17, 18, 19, 22 – Cymatosira spp.: 17, 18, 19 – обр. 13; 22 – обр. 14; 20 – Jousea elliptica (Jousé) Gleser, обр. 13; 21 – Soleum exsculptum Heiberg, обр. 13.

Таблица II. Диатомеи из палеогеновых отложений скв. Р-321. Длина масштабной линейки 20 мкм для фиг. 1 и 10 мкм для фиг. 2–24 . 1, 2, 7 – Paralia grunowii Gleser: 1 – обр. 14; 2 – обр. 10; 7 – обр. 8; 3 – Paralia selecta (A. Schmidt) Gleser, обр. 10; 4 – Anuloplicata concentrica (A. Schmidt) Gleser, обр. 14; 5 – Paralia crenulata (Grunow) Gleser, обр. 8; 6 – Anuloplicata ornata (Grunow) Gleser, обр. 8; 8 – Actinoptychus pericavatus Brun, обр. 10; 9 – Actinoptychus simbiskianua A. Schmidt, обр. 8; 10 – Radiaplicata clavigera (Grunow) Gleser, обр. 14; 11 – Hyalodiscus radiatus (O’Meara) Grunow, обр. 10; 12, 13 – Actinoptychus sp., обр. 14; 14 – Actinoptychus seductilis A. Schmidt, обр. 14; 15 – Moisseevia probabilis (A. Schmidt) Strelnikova, обр. 12; 16 – Biddulphia tuomeyi (Bailey) Roper, обр. 12; 17 – Rhaphoneis lancetulla Grunow, обр. 10; 18 – Stephanogonia polygona Ehrenberg, обр. 3; 19 – Pterotheca spada Tempère et Peragallo, обр. 10; 20 – Biddulphia sp., обр. 9; 21 – Eunotogramma variabile Grunow, обр. 12; 22 – Goniothecium wittianus Pantocsek, обр. 14; 23 – Diploneis sp., обр. 13; 24 – Porotheca danica (Grunow) Fenner, обр. 14.

Таблица III. Диатомеи и силикофлагеллаты из палеогеновых отложений скв. Р-321. Длина масштабной линейки 20 мкм для фиг. 1, 23, 24 и 10 мкм для фиг. 2–22 . 1 – Sheshukovia sp., обр. 14; 2, 6 – Sheshukovia flos (Ehrenberg) Fenner, обр. 13; 3, 4, 8, 9 – Sheshukovia fenestra (Witt) Fenner, обр. 10; 5, 7 – Trinacria excavata Heiberg, обр. 10; 10 – Naviculopsis foliacea Deflandre, обр. 10; 11 – Naviculopsis eobiapiculata Bukry, обр.12; 12 –Corbisema hastata globulata Bukry, обр. 13; 13 – Corbisema gleserae Bukry, обр. 13; 14 –Dictyocha precarentis Bukry, обр. 10; 15, 20 – Dictyocha deflandrei Frenguelli ex Glezer: 15 – обр. 14; 20 – обр. 13; 16, 17 – Dictyocha brevispina (Lemmermann) Bukry: 16 – обр. 10; 17 – обр. 12; 18, 19 – Naviculopsis minor (Schulz) Frenguelli: 18 – обр. 10; 19 – обр. 14; 21 – Rattrayella sp., обр. 14; 22 – Stellarima microtrias (Ehrenberg) Hasle et Sims, обр. 3; 23 –Pseudopodosira pileiformis Jousé, обр. 14; 24 – Rhizosolenia clavigera (Grunow) Homann, обр. 9.

РЕЗУЛЬТАТЫ
При изучении керна в нижней части разреза в слое “бурого железняка” – ожелезненного песчаника, отнесенного к бузиновской свите, установлено массовое присутствие створок диатомей с признаками ожелезнения. Основной доминант – группа тихопелагических родов Paralia/Anuloplicata.
Выше по всему разрезу вёшенской свиты доминирующая позиция группы Paralia/Anuloplicata сохраняется, колеблясь в пределах 82–96% (рис. 2). Интервалы массового присутствия диатомей (обр. 3, 8, 10, 12, 14, 15) чередуются с интервалами их пониженного содержания и обедненного таксономического состава (обр. 9, 11, 13), что характерно для нестабильной обстановки краевой зоны морского бассейна. Ассоциация представлена несколькими группами диатомей (табл. I–III ). Первая – группа тихопелагических и бентосных родов, которая помимо рода Paralia включает роды Anuloplicata, Radiaplicata, Pseudopodosira, Hyalodiscus, Actinoptychus, Rhaphoneis, Eunotogramma, Diploneis. Характерны редкие виды рода Actinoptychus – A. seductilis, A. simbiskiana, A. pericavatus. Вторая группа, так называемые роды-споры Сhaetoceros, Goniothecium, Pterotheca, Stephanogonia, Jousea, также тяготеет к прибрежным обстановкам. Неритические виды с широким диапазоном стратиграфического распространения представлены Eupyxidicula turris, Stellarima microtrias, Trinacria excavata, T. subcoronata, Sheshukovia fenestra, S. flos, Coscinodiscus argus. К экзотам можно отнести род Rattrayella, представленный мелкими, не идентифицированными до вида створками. Также следует отметить присутствие нескольких морфотипов рода Cymatosira, которые в настоящее время известны в отложениях Норвежского моря, начиная с позднего эоцена (Schrader, Fenner, 1976).
Стратиграфически важные таксоны – Moisseevia uralensis, Soleum exsculptum, Eupyxidicula moelleri, Grunowiella gemmata, Hemialus febriatus, H. frigidus, H. arcticus var. bornholmensis, H. curvatulus, H. morsianus, Costopyxis broschii, C. reticulata, Biddulphia tuomeyi, Coscinodiscus decrescenoides – характерны для зоны Moisseevia uralensis нижнего эоцена диатомовой шкалы внетропической области. По-видимому, региональной особенностью выделенных ассоциаций является присутствие Jousea elliptica, Rhaphoneis lancetulla, Cymatosira spp., Proboscia spp. и упомянутых выше видов рода Actinoptychus.
По имеющимся данным по Полярному Предуралью (Iakovleva et al., 2000; Oreshkina, 2000, 2012) и Западной Сибири (Oreshkina et al., 2004; Орешкина и др., 2008; Александрова и др., 2012), зона Moisseevia uralensis соответствует диноцистовым зонам Deflandrea oebisfeldensis и Dracodinium (=Wetzeliella) astra, которые, в свою очередь, отвечают зоне NP10 по наннопланктону и датируются 55.6–54.8 млн лет (Яковлева, 2017). Для этой зоны характерно снижение таксономического разнообразия по сравнению с комплексами диатомовых зон Trinacria mirabile (верхний танет) и Hemiaulus proteus (нижний ипр) Поволжья и Зауралья (Орешкина, Александрова, 2007, 2017).
Комплекс силикофлагеллат (табл. III) представлен тремя родами – известным с мела родом Corbisema, а также родами Dictyocha и Naviculopsis, появляющимися соответственно в среднем и позднем палеоцене (Perch-Nielsen, 1976, 1985; McCartney et al., 2020). Характерны Corbisema gleserae, C. hastata globulata, D. deflandrei, D. precarentis. В составе рода Naviculopsis появляются N. foliacea, N. aff. minor. Последний таксон отличается от типовой формы более мелкими размерами, ромбовидной формой базальной части скелета и узкой апикальной перекладиной. Присутствие зонального вида-индекса Naviculopsis foliacea, первое появление которого по разным данным фиксируется с основания эоцена (Perch-Nielsen, 1976, 1985; McCartney et al., 2020), является дополнительным аргументом в пользу раннеэоценового возраста вёшенской свиты.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Полученные данные по силикофоссилиям скв. Р-321, как и результаты изучения диноцист из керна скв. 1238 и скв. 5/93 (Монастырщина) той же СФЗ Преддонецкой моноклинали (Орешкина и др., 2021; Яковлева, Александрова, 2021), дают возможность уточнить стратиграфический объем бузиновской и вёшенской свит.
В скв. 1238 в бузиновской свите установлена диноцистовая зона Alisocysta margarita нижнего танета, в вёшенской свите – зоны Apectodinium hyperacanthum и Axiodinium augustum верхнего танета–нижнего ипра. Следует отметить, что зона Axiodinium augustum по диноцистам является маркером глобального события климатического оптимума на границе палеоцена–эоцена (РЕТМ; ~55.8–55.6 млн лет назад). В скв. 5/93 (Монастырщина) комплекс диноцист присутствует только в верхней части вёшенской свиты и отнесен к зоне Stenodinium meckelfeldensis, сопоставляемой с верхней частью зоны NP10 по наннопланктону (Орешкина и др., 2021). В скв. Р-321 в отложениях, отнесенных при первичном описании керна к бузиновской и вёшенской свитам, установлена диатомовая зона Moisseevia uralensis, которая по имеющимся данным соотносится с диноцистовыми зонами Deflandrea oebisfeldensis и Dracodinium (=Wetzeliella) astra (NP 10) (рис. 3). Исходя из полученных микропалеонтологических данных, слои “бурого железняка”, конгломерата и опоковидного песчаника общей мощностью 1.15 м следует относить не к бузиновской, а к вёшенской свите. Динамика ассоциаций силикофоссилий и литологическое строение разреза отражают два этапа развития мелководного прибрежного бассейна: (1) относительно застойные условия с признаками ожелезнения донных осадков (слои 2–4); (2) развитие трансгрессии с массовым присутствием спикул губок в начале трансгрессивного этапа, появлением отдельных открыто-морских видов по мере нарастания трансгрессии (слой 5). Массовое присутствие тихопелагических видов по всему разрезу скважины говорит о незначительных глубинах этого бассейна. Таким образом, в изученных нами разрезах Преддонецкой СФЗ вёшенская свита имеет неодинаковый стратиграфический объем и различия в литологическом составе.
Рис. 3.
Корреляция разрезов Преддонецкой и Ульяновско-Сызранской структурно-фациальных зон (СФЗ) с международной и региональной стратиграфическими шкалами, зональными шкалами по диатомовым водорослям и диноцистам. Сокращения: зоны по диноцистам: Och.rom./S.chl. – Ochetodinium romanum/Samlandia chlamydophora, E. – Eatonicysta, Ch. – Charlesdownea, Dr. – Dracodinium, D. – Deflandrea, A. – Axioidinium; зоны по диатомеям: M. – Moisseevia, H. – Hemiaulus.
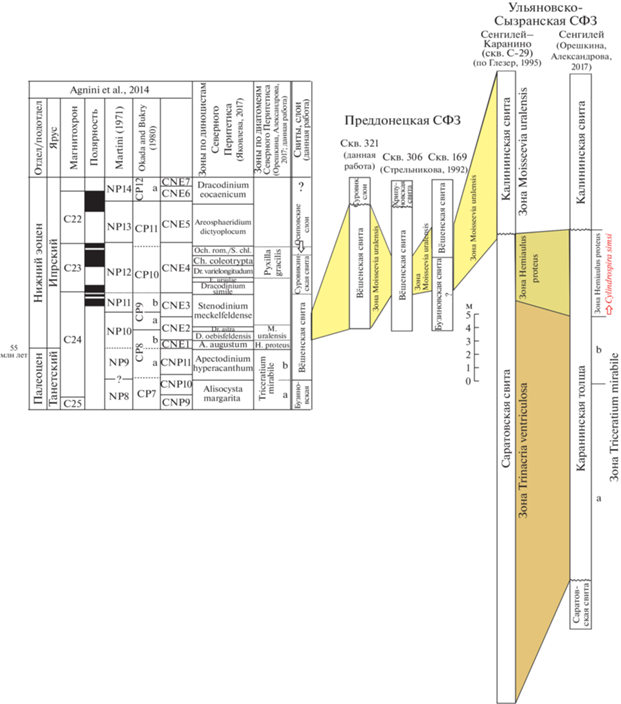
По данным Н.И. Стрельниковой (1992), таксономически близкие комплексы диатомей выделены из разрезов скв. 169 у станции Новохопёрск и скв. 306 (район Новохопёрска) (рис. 1, 3). Н.И. Стрельникова относила эти комплексы к слоям с Aulacodiscus tener (Witt) Hustedt и считала их переходными между палеоценом и эоценом. Для них характерны те же таксоны, что и в комплексе скв. Р-321, включая фоновую ассоциацию с доминированием группы Paralia/Anuloplicata и реперные виды Coscinodiscus (=Moisseevia) uralensis, Jousea elliptica, Pyxidicula (=Eupyxidicula) moelleri, Coscinodiscus decrescenoides, C. argus, Actinoptychus pericavatus.
Комплекс силикофоссилий с близким составом диатомей и силикофлагеллат описан из калининской свиты в скв. С-29 у с. Каранино Ульяновской области (рис. 1, 3), расположенной к северо-западу от г. Сенгилей в верховьях р. Атцы (Глезер, 1995). В подстилающей калининскую свиту толще диатомитов разреза Сенгилей (“Граное Ухо”), отнесенной к каранинской толще, установлена непрерывная последовательность зон по диатомовым водорослям в интервале перехода от палеоцена к эоцену: зона Trinacria mirabile с двумя подзонами (верхний танет) и зона Hemiaulus proteus (нижний ипр) (Унифицированная…, 2015; Орешкина, Александрова, 2017). Комплекс калининской свиты характеризуется обеднением таксономического состава по сравнению с комплексом зоны Hemiaulus proteus, хотя отдельные таксоны, такие как Hemiaulus proteus, H. polymorphus var. morsianus, H. febriatus, Soleum exsculptum, Grunowiella gemmata, сохраняют свое присутствие. Отмечается появление Pseudotriceratium exornatum (Meinster) Gleser, Actinopthychus pericavatus, силикофлагеллаты Naviculopsis foliacea.
Интервалы, обогащенные остатками кремневых микрофоссилий в интервале перехода от палеоцена к эоцену, широко распространены в палеогеновых бассейнах Северной Европы и донных отложениях Северного моря (рис. 1, 4). На Самбийском полуострове (Калининградская область) в скв. 2-я Пионерская (Стрельникова и др., 1978; Стрельникова, 1992) в темно-серых глинах и алевролитах самбийской свиты (интервал 36.6–33.3 м) присутствует комплекс диатомей с зональным видом-индексом нижнего ипра Hemiaulus proteus и характерными представителями позднепалеоценовой диатомовой флоры: Triceratium sundbyense Hustedt, Trinacria ventriculosa (A. Schmidt) Gleser, Anaulus weyprechtii Grunow, Pseudopodosira anissimovae (Gleser et Rubina) Strelnikova. Состав комплекса типичен для зоны Hemiaulus proteus. Более молодая датировка самбийской свиты получена по диноцистам из близко расположенной скв. 1 Янтарный (Александрова, Запорожец, 2008; Kasinski et al., 2020), где были установлены зона Deflandrea oebisfeldensis нижнего ипра и ее аналоги. Можно предположить, что разница в оценке возраста может быть связана с различным объемом самбийской свиты в изученных разрезах или влиянием факторов размыва и переотложения, характерных для зоны ледниковых дислокаций.
Рис. 4.
Корреляция диатомовых комплексов палеогена бассейна Северного моря в интервале перехода от палеоцена к эоцену.

Близкие по составу комплексы диатомей и силикофлагеллат из пограничных отложений палеоцена–эоцена известны из многочисленных публикаций по Северному морю (рис. 5), что отмечалось Н.И. Стрельниковой (1992), З.И. Глезер (1995) и др. Из наиболее широко известных наземных местонахождений – формация Фур Северной Дании (Heiberg, 1863; Grunow, 1866; Kitton, 1870; Benda, 1972; Sims, 1989; Homann, 1991; Tsutsui et al., 2018) и разрезы Северной Германии (Schulz, 1927; Benda, 1965), которые изучались в основном в таксономическом аспекте. Схема зонального расчленения формации Фур по силикофлагеллатам из 5 зон (рис. 4) была предложена K. Перч-Ниелсен (Perch-Nielsen, 1976). Выделенные зоны были привязаны к тефрохронологической шкале с корреляционным потенциалом, охватывающим практически всю Северную Европу и сопредельные акватории Северной Атлантики. Позже A. Митленер (Mitlehner, 1996) при изучении ограниченной выборки образцов из формации Фур выявил на уровне пеплового слоя –21 биотическое событие, выраженное в увеличении количества открыто-морских видов, свидетельствующем об усилении связей с бассейнами Северной Атлантики и Тетиса. На значительную перестройку комплексов диноцист в формации Фур на этом уровне же указывают данные П. Виллумсен (Willumsen, 2004). Показательно, что на этом уровне появляется монотипный таксон с ультракоротким стратиграфическим диапазоном и атипичной морфологией, описанный (Mitlehner, 1995) как новый род Cylindrospira. Практически одновременно Н.И. Стрельниковой и В. А. Николаевым (1995) этот таксон был найден и описан как новый род Gyrocylindrus в верхней части разреза Сенгилей Ульяновской области. В настоящее время эта часть разреза относится к каранинской толще и к зоне Hemiaulus proteus, которая отвечает диноцистовой зоне Apectodinium augustum (Орешкина, Александрова, 2007, 2017).
Сходные по составу комплексы диатомей нижнего ипра описаны из отложений скв. Харре, пробуренной в заливе Лимфиорд, примерно в 20 км от береговых обнажений формации Фур (Fenner, 1994). В интервале между пепловыми слоями от ‒34 до +130 (примерно 25 м по мощности) доминируют меропланктонные виды диатомовых и хризофиты. Среди диатомовых пресноводные формы и бентос играют подчиненную роль, что предполагает осадконакопление в условиях мелководного шельфа с незначительным влиянием речного и эолового факторов. Для определения возраста выделенных комплексов Дж. Феннер (Fenner, 1994) использует как анализ стратиграфического распределения диатомей в океанических осадках по данным глубоководного бурения, так и совместные данные по тефрохронологии и карбонатному планктону в Северной Атлантике. Исходя из этих корреляций, скв. Харре в интервале между пеплами –30 и +130 отнесена к зоне NP10 (=СP9a) по наннопланктону.
Реконструкции палеообстановок (Mitlehner, 1996) для формации Фур говорят о глубинах не более 50 м. Близость береговой линии подтверждается обилием ископаемых остатков насекомых, наземных растений и птиц. Механизм накопления диатомитов связывается с локальным апвеллингом, обусловленным сезонными сгонными ветрами. Находки глендонитов подтверждают относительно низкие придонные температуры (ниже 5°C; Vickers et al., 2020).
В донных осадках Северного моря (Malm et al., 1984; Thomsen, Danielsen, 1995; Mitlehner, 1995; Richardt, Sheldon, 2014) и в ряде наземных разрезов Парижского и Бельгийского бассейнов установлены интервалы, содержащие пиритизированные створки диатомей, что связывают с аноксическими условиями в придонных слоях. Начиная с работы по лондонским глинам (Shrubsole, Kitton, 1881), пиритизированные комплексы диатомей пытались использовать в качестве реперов для стратиграфического расчленения и корреляции донных осадков Северного моря. А. Митленер (Mitlehner, 1996) детализировал зональную схему по микропланктону для осадков Северного моря (King, 1983), опираясь на смену комплексов доминирующих видов среди пиритизированных комплексов диатомей (рис. 4), что, по его мнению, маркировало границу палеоцена и эоцена. Позже в серии работ по североморским бассейнам (Van Eetvelde, Cornet, 2002; Van Eetvelde et al., 2004; Van Eetvelde, 2005) было показано, что граница палеоцена–эоцена (основание изотопного события CIE) находится ниже пика численности диатомеи Fenestrella antiqua (Grunow) Swatman и вблизи пика численности Coscinodiscus morsianus var. morsianus (Sims) Mitlehner. С учетом исторически сложившихся различий в номенклатуре диатомовых водорослей в отечественной и зарубежной литературе по Северной Европе, приведенный в этих работах список видов практически идентичен составу диатомей нижнего ипра Преддонецкой и Ульяновско-Сызранской СФЗ. Тем не менее сопоставление последовательной смены диатомовых комплексов палеогена Североморского бассейна и эпиконтинентальных бассейнов Восточно-Европейской платформы затрудняется отсутствием единого подхода к биостратиграфическому расчленению на зональной основе, прямых корреляций с другими группами микропланктона, в частности с диноцистами.
ЗАКЛЮЧЕНИЕ
В Преддонецкой СФЗ терригенно-кремнистые фации приурочены к верхней части вёшенской свиты, развитой в центральной части рассматриваемого региона. Комплекс диатомей отнесен к диатомовой зоне Moisseevia uralensis, отвечающей диноцистовым зонам Deflandrea oebisfeldensis и Dracodinium (=Wetzeliella) astra (Oreshkina et al., 2004; Орешкина, Александрова, 2007; Oreshkina, 2012), которые, в свою очередь, соотносятся с зоной NP10 по наннопланктону (55.6–54.8 млн лет) (Яковлева, 2017). Комплекс силикофлагеллат отнесен к зоне Naviculopsis foliacea нижнего ипра (Perch-Nielsen, 1976, 1985; McCartney et al., 2020). Таким образом, в Преддонецкой СФЗ развитие биокремнистых фаций приурочено к нижнему ипру и к интервалу, охватывающему конец лютета–начало приабона (Орешкина и др., 2021).
Комплекс диатомовой зоны Moisseevia uralensis с доминированием группы Paralia/Anuloplicata и спорадическим присутствием зонального вида-индекса Moisseevia uralensis, а также видов Jousea elliptica, Eupyxidicula moelleri, Coscinodiscus decrescenoides, C. argus, Soleum exsculptum, Grunowiella gemmata, Rhaphoneis lancetulla, Actinopthychus pericavatus, A. seductilis, A. simbirskiana отражает нестабильные палеообстановки мелководного бассейна, близость береговой линии, возможное опреснение. Нижняя часть вёшенской свиты, представленная слоем “бурого железняка”, прослоем конгломерата и слоем опоковидного песчаника общей мощностью 1.15 м, отражает ингрессивный эпизод с условиями застойного бассейна со слабой гидродинамикой и пониженным содержанием кислорода в придонных слоях. Последующая стадия развивавшейся трансгрессии в прибрежной нестабильной обстановке (кремнистые глины мощностью 8.2 м) охарактеризована таксономически более разнообразными диатомовыми ассоциациями без следов пиритизации.
Изученный интервал вёшенской свиты, обогащенный кремневым микропланктоном, является частью регионального события биогенного кремненакопления конца палеоцена–начала раннего эоцена, известного в Среднем Поволжье, на побережье Балтийского моря и в североморских бассейнах. Близкий таксономический состав диатомей и силикофлагеллат этих регионов подразумевает устойчивый водообмен и миграционные связи между этими регионами Северного Перитетиса.
Благодарности. Автор выражает благодарность С.В. Гришину за помощь в лабораторной обработке проб, Н.В. Горьковой за фотосъемку в сканирующем электронном микроскопе. Критические замечания и полезные рекомендации Г.Н. Александровой и рецензентов В.С. Вишневской, И.Б. Цой и А.Ю. Гладенкова помогли значительно улучшить содержание статьи.
Источники финансирования. Исследования проведены по теме государственного задания ГИН РАН и при поддержке гранта РФФИ № 18-05-00505.
Список литературы
Александрова Г.Н., Запорожец Н.И. Палинологическая характеристика верхнемеловых и палеогеновых отложений запада Самбийского полуострова (Калининградская область). Статья 2 // Стратиграфия. Геол. корреляция. 2008. Т. 16. № 5. С. 75–86.
Александрова Г.Н., Орешкина Т.В., Яковлева А.И., Радионова Э.П. Диатомеи и диноцисты в интервале позднего палеоцена–раннего эоцена в биокремнистых фациях Среднего Зауралья // Стратиграфия. Геол. корреляция. 2012. Т. 20. № 4. С. 68–94.
Александрова Г.Н., Орешкина Т.В., Яковлева А.И. Актуализация региональной стратиграфической схемы палеогена юга Русской плиты: проблемы и пути решения // Проблемы региональной геологии Северной Евразии. Материалы конференции “XII Научные чтения, посвященные памяти проф. М.В. Муратова”. М.: МГРИ-РГГРУ, 2020. С. 3–5.
Ахметьев М.А., Беньямовский В.Н. Стратиграфическая схема морского палеогена юга Европейской России // Бюлл. МОИП. Отд. геол. 2003. Вып. 5. С. 40–51.
Беньямовский В.Н. Предложения по структурно-фациальному районированию и субрегиональным горизонтам палеогена Воронежско-Придонецкого субрегиона // ПАЛЕОСТРАТ-2016. Москва, 26–28 января 2016 г. Программа и тезисы докладов. Ред. Алексеев А.С. М. : ПИН РАН, 1916. С. 15–16.
Глезер З.И. Зональное расчленение палеогеновых отложений по диатомовым водорослям // Сов. геология. 1979. № 11. С. 19–30.
Глезер З.И. Комплексы диатомей и силикофлaгеллат – показатели синхронности диатомитов Среднего Поволжья, Дании и океанических осадков нижнего палеогена // Современный и ископаемый планктон Мирового океана. М.: Наука, 1995. С. 57–63.
Леонов Г.П. Основные вопросы региональной стратиграфии палеогеновых отложений Русской плиты. М.: Изд-во МГУ, 1961. 552 с.
Орешкина Т.В., Александрова Г.Н. Терминальный палеоцен Среднего Поволжья: биостратиграфия и палеообстановки // Стратиграфия. Геол. корреляция. 2007. Т. 15. № 2. С. 93–118.
Орешкина Т.В., Александрова Г.Н. Палеонтологическая характеристика палеоцена–нижнего эоцена Ульяновско-Сызранской структурно-фациальной зоны Поволжско-Прикаспийского субрегиона // Стратиграфия. Геол. корреляция. 2017. Т. 25. № 3. С. 73–98.
Орешкина Т.В., Алексеев А.С., Смирнова С.Б. Мел-палеогеновые отложения Полярного Предуралья: биостратиграфические и палеогеографические аспекты // Урал: фундаментальные проблемы геодинамики и стратиграфии. М.: Наука, 1998. С. 183–192 (Тр. ГИН РАН. Вып. 500).
Орешкина Т.В., Яковлева А.И., Александрова Г.Н. Прямая корреляция зональных шкал бореального палеогена по диатомеям и диноцистам (по материалам скв. 19-У, пос. Усть-Манья, восточный склон Северного Урала) // Геология и геофизика. 2008. Т. 49. Вып. 10–11. С. 347–350.
Орешкина Т.В., Яковлева А.И., Александрова Г.Н. Силикофоссилии и диноцисты из терригенно-кремнистых отложений нижнего палеогена юга Русской плиты и их значение для обоснования возраста свит // Стратиграфия. Геол. корреляция. 2021. Т. 29. № 3. С. 45–71. https://doi.org/10.31857/S0869592X21030042
Палеогеографический атлас Северной Евразии. М.: Институт тектоники литосферных плит РАЕН, 1998. CDROM.
Постановления Межведомственного стратиграфического комитета и его постоянных комиссий. Вып. 32. СПб.: Изд-во ВСЕГЕИ, 2001. С. 18–20.
Рыборак В.М., Шокуров В.П. Отчет о проведении геологического, гидрогеологического и инженерно-геологического доизучения масштаба 1: 200 000 с эколого-геологическими исследованиями на площади листа M-37-XII (Новохоперск). Воронеж: ВоронежГеология, 1999.
Семенов В.П. Палеоген Воронежской антеклизы. Воронеж: Изд-во Воронежского ун-та, 1965. 279 с.
Стрельникова Н.И. Палеогеновые диатомовые водоросли. СПб.: Изд-во Санкт-Петербургского ун-та, 1992. 312 с.
Стрельникова Н.И., Каплан А.А., Травина М.А. Палеогеновые диатомеи, силикофлагелляты и эбриидеи Калининградской области // Морская микропалеонтология. М.: Наука, 1978. С. 57–67.
Стрельникова Н.И., Николаев В.А. Новый род Gyrocylindrus (Bacillariophyta) из отложений России и Дании // Ботан. журн. 1995. Т. 80. № 8. С. 93–95.
Унифицированная стратиграфическая схема палеогеновых отложений Поволжско-Прикаспийского субрегиона. Объяснительная записка. М.: ВНИГНИ, 2015. 96 с.
Яковлева А.И. Детализация эоценовой диноцистовой шкалы для Восточного Перитетиса // Бюлл. МОИП. Отд. геол. 2017. Т. 92. Вып. 2. С. 32–48.
Яковлева А.И., Александрова Г.Н. Палеоген Преддонецкой моноклинали и его палинологическая характеристика // Стратиграфия. Геол. корреляция. 2021. Т. 29. № 1. С. 59–98.
Agnini C., Fornaciari E., Raffi I., Catanzariti R., Palike H., Backman J., Rio D. Biozonation and biochronology of Paleogene calcareous nannofossils from low and middle latitudes // Newsl. Stratigr. 2014. V. 47. P. 131–181.
Aleksandrova G.N., Radionova E.P. On the Late Paleocene stratigraphy of the Saratov Volga Region: micropaleontological characteristics of the Kamyshin Formation, Dyupa Gully Section // Paleontol. J. 2006. V. 40. Suppl. 5. P. 543–557.
Benda L. Diatomeen aus dem Eozän Norddeutschlands // Paläont. Zeitschrift. 1965. № 39. P. 165–187.
Benda L. The diatoms of the Moler Formation of Denmark (Lower Eocene). A preliminary report // Beih. Nowa Hedwigia. 1972. № 39. P. 251–266.
Blanco S., Wetzel C.E. Replacement names for botanical taxa involving algal genera // Phytotaxa. 2016. V. 266. № 3. P. 195–205. https://doi.org/10.11646/phytotaxa.266.3.3
Fenner J. Diatoms of the Fur Formation, their taxonomy and biostratigraphic interpretation – results from the Harre borehole, Denmark // Aarhus Geosci. 1994. № 1. P. 99–163.
Grunow A. Moler aus Jütland, eingesandt von Th. Jensen, analisiert von A. Grunow // Hedwigia. 1866. № 5. P. 145–146.
Heiberg P.A.C. Conspectus criticus diatomacearum danicarum: Kritisk Oversigt over De Danske Diatomeer. Copenhagen: William Priors Forlag, 1863. 135 p.
Heilmann-Clausen C. The Danisch subbasin. Paleogene dinoflagellates// Geol. Jb. 1988. V. A 100. P. 339–343.
Heilmann-Clausen C., Thomsen E. Early Paleogene warm climates and biosphere dynamics // Guide to field excursion in Denmark, June 12–13, 1999. University of Aarhus, Department of Earth Sciences, 1999. 43 p.
Homann M. Die Diatomeen der Fur-Formation (Alt-tertiär) aus dem Limfjord-Gebeit, Nordjutland/Dänemark // Geologisches Jahrbuch. 1991. Reihe A. 123. 285 p.
Iakovleva A.I., Oreshkina T.V., Alekseev A.S., Rousseau D.-D.A. New Paleogene micropaleontological and palaeogeographical data in the Petchora Depression, northern European Russia // Earth Planet. Sci. 2000. V. 330. P. 485–491.
Kasinski J.R., Kramarska R., Slodkovska B., Sivkov V., Piwocki M. Paleocene and Eocene deposits on the eastern margin of the Gulf of Gdañsk (Yantarny P-1 bore hole, Kaliningrad region, Russia) // Geol. Quarterly. 2020. V. 64. № 1. P. 29–53. https://doi.org/10.7306/gq.1513
King C. Cainozoic Micropalaeontological Biostratigraphy of the North Sea // Inst. Geol. Sci. Rep. 1983. V. 82. № 7. 40 p.
Kitton F. Diatomaceous deposits from Jutland// Journal of the Quekett Microscopical Club. 1870. № 2. P. 99–102, 168–171.
Malm O.A., Christensen O.B., Furnes H., Løvlie R., Rueslåtten H., Østby K.L. The Lower Tertiary Balder Formation: an organogenic and tuffaceous deposit in the North Sea region // Petroleum Geology of the North European. London: Graham & Trotman, 1984. P. 149–170.
Martini E. Standard Tertiary and Quaternary calcareous nannoplankton zonation // Proc. II Planktonic Conf. Roma, 1970. 1971. P. 739–785.
McCartney K., Witkowski J., Nowakowski R., Wrobel R., Zgłobicka I. Evolution of the silicoflagellate naviculopsid skeletal morphology in the Cenozoic // Mar. Micropaleontol. 2020. V. 156. https://doi.org/10.1016/j.marmicro.2019.101820
Mitlehner A.G. Cylindrospira, a new diatom genus from the palaeogene of Denmark with palaeoecological significance // Diatom Res. 1995. V. 10. № 2. P. 321–331. https://doi.org/10.1080/0269249X.1995.9705352
Mitlehner A.G. Paleoenvironments in the North Sea Basin around the Paleocene–Eocene boundary: evidence from diatoms and other siliceous microfossils // Correlation of the Early Paleogene in Northwest Europe. Geol. Soc. Spec. Publ. 1996. № 101. P. 255–273.
Okada H., Bukry D. Supplementary modification and introduction of code numbers to the low-coccolith biostratigraphic zonation (Bukry, 1973; 1975) // Mar. Micropaleontol. 1980. V. 5. № 3. P. 321–325. https://doi.org/10.1016/0377-8398(80)90016-X
Oreshkina T.V. New data on Early Eocene diatom successions of the West Polar Urals margin: biostratigraphic and paleogeographic implications // GFF. 2000. V. 122. Pt. 1. P. 124–126.
Oreshkina T.V. Evidence of Late Paleocene–Early Eocene hyperthermal events in biosiliceous sediments of Western Siberia and adjacent areas // Austrian J. Earth Sci. 2012. V. 105. № 1. P. 145–153.
Oreshkina T.V., Oberhansli H. Diatom turnover in the early Paleogene diatomite of the Sengiley section, Middle Povolzhie, Russia: a response to the Initial Eocene Thermal Maximum? // Causes and Consequences of Globally Warm Climates in the Early Paleogene. Geol. Soc. Am. Spec. Pap. 2003. V. 369. P. 169–179.
Oreshkina T.V., Radionova E.P. Diatom record of the Paleocene–Eocene Thermal Maximum in marine paleobasins of Central Russia, Transuralia and adjacent regions // Nova Hedwigia. 2014. B. 143. P. 307–336.
Oreshkina T.V., Aleksandrova G.N., Kozlova G.N. Early E-ocene marine planktonic record of the East Urals margin (Sverdlovsk region): biostratigraphy and paleoenvironments // Neues Jahrb. Geol. Paläontol. Abhandlungen. 2004. V. 234. № 1. P. 201–222.
Perch-Nielsen K. New silicoflagellates and a silicoflagellate zonation in north European Palaeocene and Eocene diatomites // Bull. Geol. Soc. Denmark. 1976. V. 25. P. 27–40.
Perch-Nielsen K. Cenozoic calcareous nannofossils // Plankton Stratigraphy. Cambridge: Cambridge University Press, 1985. P. 427–554.
Radionova E.P., Khokhlova I.E., Beniamovskii V.N., Shcherbinina E.A., Iakovleva A.I., Sadchikova T.A. Paleocene/Eocene transition in the northeastern Peri-Tethys area: Sokolovskii key section of the Turgay Passage (Kazakhstan)// Bull. Soc. Géol. France. 2001. V. 172. № 2. P. 245–256.
Radionova E.P., Beniamovski V.N., Iakovleva A.I., Muzylöv N.G., Oreshkina T.V., Shcherbinina E.A., Kozlova G.E. Early Paleogene transgressions: stratigraphical and sedimentological evidence from the northern Peri-Tethys // Causes and Consequences of Globally Warm Climates in the Early Paleogene. Geol. Soc. Am. Spec. Pap. 2003. V. 369. P. 239–261.
Richardt P.A., Sheldon E. Palynological and microfossil biostratigraphy and palaeoecology over the Paleocene–E-ocene transition, Femern Bælt, Northern Germany // Geol. Surv. Denmark Greenland Bull. 2014. V. 31. P. 35–38.
Schrader H.-J., Fenner J. Norwegian Sea Cenozoic diatom biostratigraphy and taxonomy // Init. Rep. DSDP. 1976. V. 38. P. 921–1099.
Schulz P. Diatomeen aus norddeutschen basalt-tuffen und -tuffgeschieben // Zeitschrift fur Geschiebeforschung. 1927. № 3. P. 113–126.
Shrubsole W.H., Kitton F. The diatoms of the London Clay. With a list of species and remarks // J. Roy. Microscopical Soc. 1881. № 11. P. 381–387.
Sims P.A. Some Cretaceous and Palaeocene species of Coscinodiscus: a micromorphological and systematic study // Diatom Res. 1989. V. 4. № 2. P. 351–371.
Thomsen E., Danielsen M. Transitional Paleocene–Eocene ash-bearing diatomite in the eastern North Sea // Tertiary Res. 1995. № 15. P. 11–120.
Tsutsui H., Jordan R.W., Nishiwaki N., Nishida S. Morphometric analysis of early Eocene Corbisema skeletons (Silicoflagellata) in Mors, Denmark // J. Micropalaeontol. 2018. № 37. P. 283–293. https://doi.org/10.5194/jm-37-283-2018
Van Eetvelde Y. Biostratigraphical study of the pyritized diatom assemblages from the upper Paleocene and lower Eocene sediments of the Danish, Dieppe-Hampshire and Belgian Basins – paleoenvironmental contributions. Ph.D. thesis in Geology. Facultes universitaires Notre-Dame de la paix Namur. Département de Géologie, 2005. 241 p.
Van Eetvelde Y., Cornet C. Some Eocene species of marine pyritized diatoms in the core of Saint-Josse (France) // Diatom Res. 2002. № 17. P. 423–435.
Van Eetvelde Y., Dupuis C., Cornet C. Pyritized diatoms: a good fossil marker in the Upper Paleocene-Lower Eocene sediments from the Belgian and Dieppe-Hampshire basins // Netherlands J. Geosci. 2004. V. 83. № 3. P. 173–178.
Vickers M.L., Lengger S.K., Bernasconi S.M., Thibault N., Schultz B.P., Fernandez A., Ullmann C.V., McCormack P., Bjerrum Ch.J., Rasmussen J.A., Hougård I.W., Korte Ch. Cold spells in the Nordic seas during the Early Eocene Greenhouse // Nature Communications. 2020. V. 11 (4713). https://doi.org/10.1038/s41467-020-18558-7
Willumsen P.S. Palynology of the Lower Eocene deposits of northwest Jutland, Denmark // Bull. Geol. Soc. Denmark. 2004. V. 52. P. 141–157.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция