

Стратиграфия. Геологическая корреляция, 2022, T. 30, № 3, стр. 62-89
Верхнемеловые отложения Вольской структурной зоны Восточно-Европейской платформы: турон–нижний кампан разреза Коммунар. Статья 2. Макрофаунистическая характеристика, выводы
Е. М. Первушов 1, *, И. П. Рябов 1, В. Б. Сельцер 1, И. Валащик 2, Е. А. Калякин 1, А. А. Гужикова 1, Е. И. Ильинский 1, Д. В. Худяков 1
1 Саратовский национальный исследовательский государственный университет
им. Н.Г. Чернышевского
Саратов, Россия
2 Варшавский университет
Варшава, Польша
* E-mail: pervushovem@mail.ru
Поступила в редакцию 28.03.2021
После доработки 31.08.2021
Принята к публикации 15.09.2021
- EDN: TXHQDI
- DOI: 10.31857/S0869592X22030048
Аннотация
В разрезе Коммунар в литологически однородном комплексе карбонатных пород прослежено вертикальное распространение иноцерамов, белемнитов и аммонитов, иглокожих, губок и брахиопод. Результаты исследований представителей макрофоссилий подтверждают и дополняют характеристику выделенных по бентосным фораминиферам биостратиграфических подразделений среднего–верхнего турона, коньяка, нижнего и верхнего сантона, базального горизонта нижнего кампана. В разрезе Коммунар, впервые на территории Поволжья, прослежен биостратиграфически непрерывный пограничный интервал верхнего коньяка–нижнего сантона. Представлено соотношение биостратиграфических подразделений, выделенных по установленным в разрезе Коммунар представителям микро- и макрофоссилий. Результаты бореально-тетической магнитохронологической корреляции позволяют заключить, что геохронологическое положение нижней границы сантона в разных регионах различается на величину порядка миллиона лет, что сопоставимо с продолжительностью этого века.
ВВЕДЕНИЕ
Расчленение туронско-нижнекампанских карбонатных пород в карьере Коммунар основано на изучении бентосных фораминифер и сопровождается результатами магнитостратиграфических исследований, что позволяет представить сопоставления регионального и межрегионального уровня. Эти данные, освещенные в первой статье, посвященной рассмотрению верхнемеловых отложений разреза Коммунар (Первушов и др., в печати), дополняются согласованием положения границ биостратиграфических подразделений по данным распределения макрофоссилий (рис. 1), характеристика комплексов которых представлена в данной работе. Обзорная схема района исследований, с указанием положения изученных разрезов, схема взаимоотношений стратиграфических комплексов верхнего мела Вольско-Хвалынской впадины и структурно-тектоническая схема Правобережного Поволжья представлены в первой статье (Первушов и др., в печати, рис. 1–3).
Рис. 1.
Распределение ископаемых беспозвоночных в разрезе Коммунар. Сокращения: зона БФ и подзона БФ – зона и подзона по бентосным фораминиферам, выделенные по схеме В.Н. Беньямовского (2008а, 2008б), МО – можжевелоовражная свита, МЛ – мезинолапшиновская свита.

Рис. 1.
Окончание
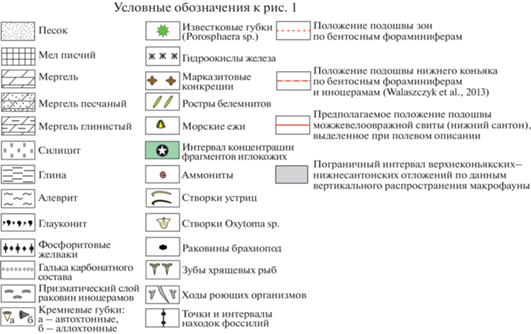
Рис. 2.
Соотношение биостратиграфических подразделений в разрезе Коммунар, установленных в нижней части верхнемеловых отложений.

Рис. 3.
Сопоставление магнитостратиграфического разреза турона–нижнего кампана карьера Коммунар с одновозрастными отложениями других регионов и Шкалой геологического времени (GTS) (Gradstein et al., 2020). Сокращения: зона по БФ и подзона по БФ – зона и подзона по бентосным фораминиферам, выделенные по схеме В.Н. Беньямовского (2008а, 2008б); зона по ПФ – зона по планктонным фораминиферам; МО – можжевелоовражная свита, МЛ – мезинолапшиновская свита. Условные обозначения см. рис. 1.
БИОСТРАТИГРАФИЧЕСКОЕ РАСЧЛЕНЕНИЕ РАЗРЕЗА КОММУНАР
Ранее в разрезе Коммунар на основании находок иноцерамов и кремневых губок были выделены маломощные интервалы верхнего, а позже и нижнего сантона (Первушов и др., 2020а, 2020б). При полевом описании подошва выделяемых слоев соотносилась с интервалами насыщения карбонатных пород турона–коньяка псаммитовым или пелитовым материалом (рис. 1). Выделение зон и подзон по бентосным фораминиферам (БФ) в соответствии со шкалой В.Н. Беньямовского (2008а, 2008б) и последующими ее корректировками (Walaszczyk et al., 2013; Vishnevskaya et al., 2018), сопряженное с представлениями о вертикальном распространении цефалопод, иноцерамов, иглокожих и губок, подтвердило обоснованность полевого выделения слоев 3, 4, 6, 8, 9 и 10. Подошва слоя 5 была выбрана условно, по поверхности одной из рабочих ступеней карьера. По результатам изучения бентосных фораминифер слои 4 и 5 соответствуют зоне БФ LC6 (нижний коньяк), при этом большая часть слоя 4 отвечает подзоне LC6а (верхняя часть верхнего турона–нижняя часть нижнего коньяка), а верхи слоя 4 и слой 5 соответствуют подзоне LC6b (верхняя часть нижнего коньяка).
В карьерах Большевик и Красный Октябрь породы переходного коньякского–сантонского интервала не были выявлены (Олферьев и др., 2009а, 2009б, 2014), поэтому в разрезе Коммунар особое внимание уделено рассмотрению положения подзоны БФ LC8b, подошва которой определена в верхней части слоя 6, а кровля – в средней части слоя 7 (рис. 1). Подошва и кровля этого биостратона литологически не выражены, а по своему объему подзона объединяет терминальную часть верхнего коньяка и базальные интервалы нижнего сантона (Беньямовский, 2008б). В средней части подзоны LC8b отмечен прослой галек карбонатного состава, марказитовых стяжений и фоссилий, который принят за подошву слоя 7. Над этим прослоем найдены скелеты кремневых губок и раковины иноцерамов кардиссоидного облика, что может свидетельствовать о раннесантонском возрасте вмещающих образований в стратотипическом разрезе вольской свиты (коньяк Поволжья).
Соотношение биостратиграфических подразделений, которые установлены в разрезе Коммунар, представлено на рис. 2.
Иноцерамы (рис. 1). В нижней части разреза Коммунар, до уровня обр. 3 (микрофаунистические пробы) иноцерамы представлены одним таксоном Inoceramus apicalis Woods. Этот вид указывает на нижнюю часть, но не на основание среднего турона (Tröger, 1987). Inoceramus lamarcki lamarcki Parkinson установлен в интервале между обр. 3 и 4, известен из верхней части среднего турона. Выше, с уровня обр. 5/1–6, формы Inoceramus lamarcki stuemckei Heinz, Inoceramus inaequivalvis Schlüter и Mytiloides striatoconcentricus (Gümbel) доказывают присутствие верхнего турона. Граница среднего и верхнего турона не выявлена, поскольку не встречен вид Inoceramus perplexus Whitfield, являющийся маркером основания верхнего турона.
В верхней части слоя 3, в интервале между обр. 7 и 8, и в базальной части слоя 4 установлен вид Inoceramus lusatiae Andert (табл. I , фиг. 5 ). В основании слоя 4 впервые появляется Cremnoceramus waltersdorfensis waltersdorfensis (Andert) (табл. II, фиг. 5, 6 ) – ранний представитель группы C. waltersdorfensis (Walaszczyk, Wood, 1998). В разрезах Зальцгиттер-Зальдер (Wood et al., 1984; Walaszczyk, Wood, 1998; Walaszczyk et al., 2010), Пуэбло (Walaszczyk, Cobban, 2000) и в обнажениях на р. Висла (Walaszczyk, 1992), вскрывающих пограничный интервал турона и коньяка, в терминальной части турона вид C. waltersdorfensis waltersdorfensis образует моноспецифичную группировку. Этот акме-интервал называется событием Cremnoceramus waltersdorfensis (Walaszczyk, Wood, 1998; Walaszczyk, 2000). Вид I. lusatiae появляется несколько раньше, в самой верхней части турона. Это означает, что пограничный интервал турона и коньяка в изучаемом разрезе полный (рис. 2).
Таблица I . Двустворчатые моллюски турона–коньяка из разреза Коммунар. Длина масштабной линейки 1 см. 1 – Inoceramus lamarcki lamarcki Parkinson, экз. SSU SVB № 58/131, левая створка; верхний турон, слой 3; 2 – Inoceramus undulatus Mantell, экз. SSU SVB № 58/138, левая створка; верхний турон, верхняя часть слоя 3; 3 – Mytiloides striatoconcentricus (Gümbel), экз. SSU SVB № 58/141, левая створка; верхний турон, верхняя часть слоя 3; 4 – Inoceramus inaequivalvis Schlüter, экз. SSU SVB № 58/143, левая створка; верхний турон, верхняя часть слоя 3; 5 – Inoceramus lusatiae Andert, экз. SSU SVB № 58/145, правая створка; верхи верхнего турона, верхняя часть слоя 3; 6 – Neithea sexcostata (Woodward), экз. SSU SVB № 58/147, правая створка; верхний турон, средняя часть слоя 3; 7 – Platyceramus sp., экз. SSU SVB № 58/152, левая створка; средний коньяк, верхняя часть слоя 6; 8 – Inoceramus lamarcki stuemckei Heinz, экз. SSU SVB № 58/114, правая створка; верхний турон, слой 3.

Таблица II. Иноцерамы турона–коньяка из разреза Коммунар. Длина масштабной линейки 1 см. 1 – Cremnoceramus deformis erectus (Meek), экз. SSU SVB № 58/115, левая створка; нижний коньяк, слой 4; 2 – Cremnoceramus crassus crassus (Petrascheck), экз. SSU SVB № 58/116, левая створка; нижний коньяк, верхняя часть слоя 4; 3 – Cremnoceramus deformis erectus (Meek), экз. SSU SVB № 58/117, левая створка; нижний коньяк, слой 4; 4 – Cremnoceramus deformis erectus (Meek), экз. SSU SVB № 58/120, левая створка; нижний коньяк, слой 4; 5 – Cremnoceramus waltersdorfensis waltersdorfensis (Andert), экз. SSU SVB № 58/127, правая створка и отпечаток левой створки; верхи верхнего турона, кровля слоя 3; 6 – Cremnoceramus waltersdorfensis waltersdorfensis (Andert), экз. SSU SVB № 58/129, правая створка; верхи верхнего турона, кровля слоя 3; 7 – Inoceramus gibbosus Schlüter, экз. SSU SVB № 58/152, правая створка; средний коньяк, нижняя часть слоя 6.

Таблица III. Белемниты и аммониты турона, иноцерамы коньяка из разреза Коммунар. Длина масштабной линейки 1 см. 1 – Sphenoceramus subcardissoides (Schlüter), экз. SSU SVB № 58/70, левая створка; верхний коньяк, вольская свита, слой 6; 2 – Goniocamax sp. cf. intermedius (Arkhangelsky), экз. SSU SVB № 58/100, неполный ростр: 2а – спинная сторона, 2б – сбоку, 2в – сечение в передней части; верхний турон, слой 3; 3 – Hyphantoceras reussianum (d’Orbigny), экз. SSU SVB № 58/07, фрагмент оборота спирали фрагмокона: 3а – сбоку, 3б – со стороны начальных оборотов; верхний турон, слой 3 (Сельцер, Иванов, 2010; табл. 20 , фиг. 7 а-в); 4 – Volviceramus involutus (J. de C. Sowerby), экз. SSU SVB № 58/77, левая створка; средний коньяк, слой 6; 5 – Inoceramus digitatus Heine (non J. de C. Sowerby), экз. SSU SVB № 58/67, правая створка; верхний коньяк, слой 6; 6 – Scaphites geinitzi (d’Orbigny), экз. SSU SVB № 58/66, сбоку; верхний турон, слой 3; 7 – Sciponoceras bohemicum bohemicum (Fritsch), экз. SSU SVB № 58/44, сбоку; верхний турон, слой 3.

Таблица IV. Иноцерам Volviceramus sp., экз. SSU SVB № 58/153, правая и неполная левая створки; разрез Коммунар, средний коньяк, слой 6. Длина масштабной линейки 1 см.

Таблица V. Иноцерамы и белемниты турона–сантона из разреза Коммунар. Длина масштабной линейки 1 см, кроме особо отмеченных случаев. 1 – Inoceramus lusatiae Andert, экз. SSU SVB № 58/111, левая створка; верхи верхнего турона, верхняя часть слоя 3; 2 – Inoceramus lusatiae Andert, экз. SSU SVB № 58/19, левая створка; нижний коньяк, нижняя часть слоя 4; 3 – Actinocamax verus fragilis Arkhangelsky, экз. SSU SVB № 58/137: 3a – спинная сторона, 3б – сбоку, 3в – альвеолярный излом; нижний сантон, слой 7; 4 – Actinocamax verus fragilis Arkhangelsky, экз. SSU SVB № 58/124: 4a – спинная сторона, 4б – сбоку, 4в – альвеолярный излом; нижний–верхний сантон, основание слоя 8; 5 – Belemnitella schmidi Christensen et Schulz, экз. SSU SVB № 58/130: 5a – спинная сторона, 5б– брюшная сторона, 5в – неполный спинно-брюшной раскол, 5г – спереди; нижний сантон, верхняя часть слоя 7; 6 – Sphenoceramus pachti (Arkhangelsky), экз. SSU SVB № 58/157; правая створка, нижний сантон, верхняя часть слоя 7.

Таблица VI. Аммониты турона–коньяка из разреза Коммунар. Длина масштабной линейки 1 см. 1 – Lewesiceras mantelli Wright et Wright, экз. SSU SVB № 58/123: 1а – со стороны устья, 1б – сбоку; верхний турон, слой 3; 2 – Puzosia muelleri de Grossouvre, экз. SSU SVB № 58/121, левая створка; нижний коньяк, нижняя часть слоя 4: 2а – с вентральной стороны, 2б – сбоку, 2в – со стороны устья.
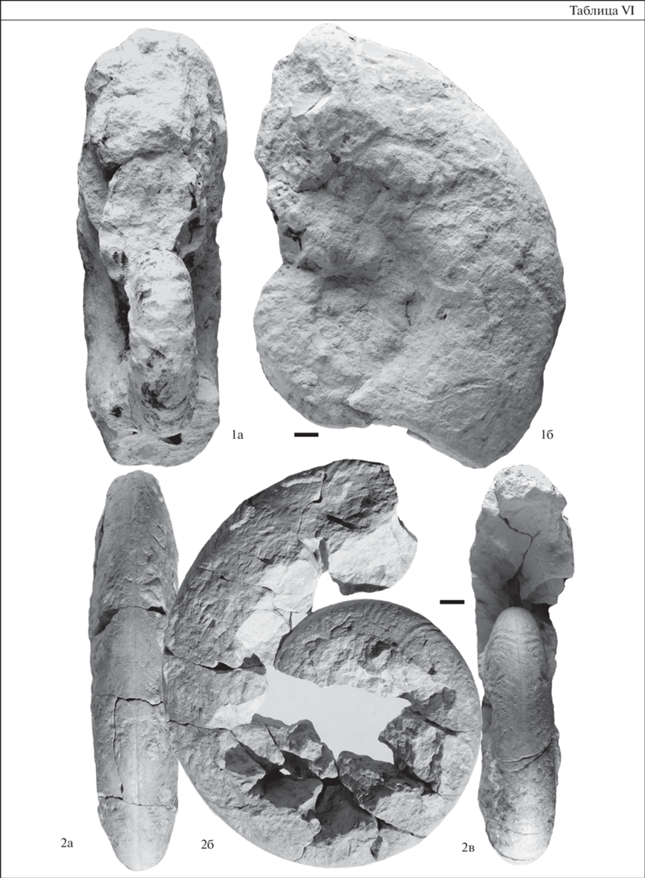
Таблица VII. Морские ежи турона–коньяка из разреза Коммунар. Длина масштабной линейки 1 см. 1 – Conulus subrotundus Mantell, экз. SSU EAK № 201/34: 1a – сверху, 1б – снизу, 1в – сбоку, 1г – сзади; средний–верхний турон, слой 3; 2 – Conulus subrotundus Mantell, экз. SSU EAK № 201/35: 2a – сверху, 2б – сбоку; средний–верхний турон, слой 3; 3 – Micraster corbovis Forbes, экз. SSU EAK № 203/91: 3a – сверху, 3б – снизу, 3в – сбоку, 3г – сзади; средний–верхний турон, слой 3; 4 – Micraster cortestudinarium (Goldfuss), экз. SSU EAK № 203/50: 4a – сверху, 4б – снизу, 4в – сбоку, 4г – сзади; средний–верхний турон, слой 3.
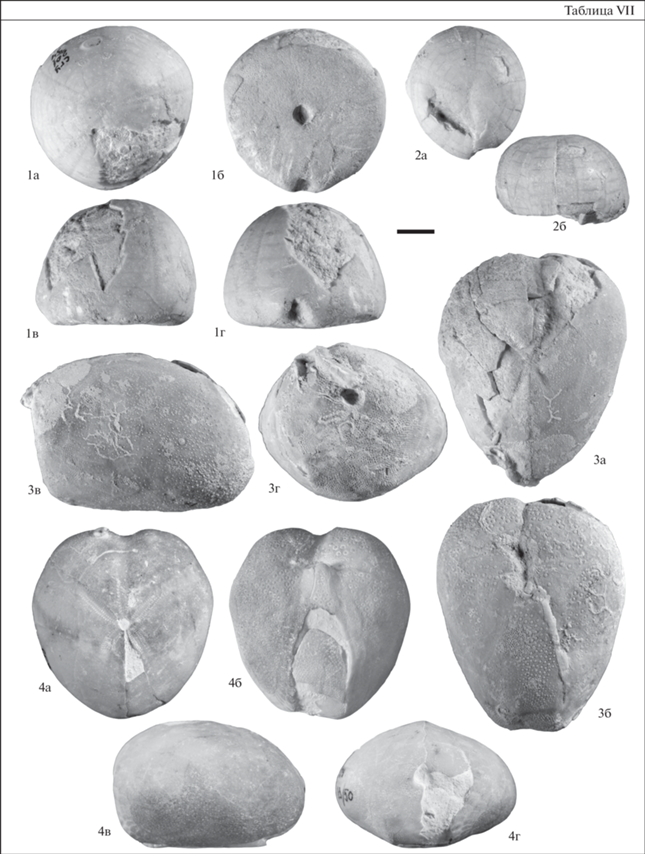
Таблица VIII. Морские ежи коньяка–сантона из разреза Коммунар. Длина масштабной линейки 1 см; а – вид сверху; б – вид снизу; в – вид сбоку; г – вид сзади. 1 – Micraster coranguinum (Leske), экз. SSU EAK № 203/76; 2 – Micraster rogalae Nowak, экз. SSU EAK № 203/77; верхний коньяк–нижний сантон, слой 7; 3 – Galeola ex gr. senonensis (d’Orbigny), экз. SSU EAK № 204.1/2; нижний кампан, слой 9; 4 – Galeola ex gr. senonensis (d’Orbigny), экз. SSU EAK № 204.1/4; нижний кампан, слой 9; 5 – Offaster pilula (Lamarck), экз. SSU EAK № 204.2/3; нижний кампан, слой 9.
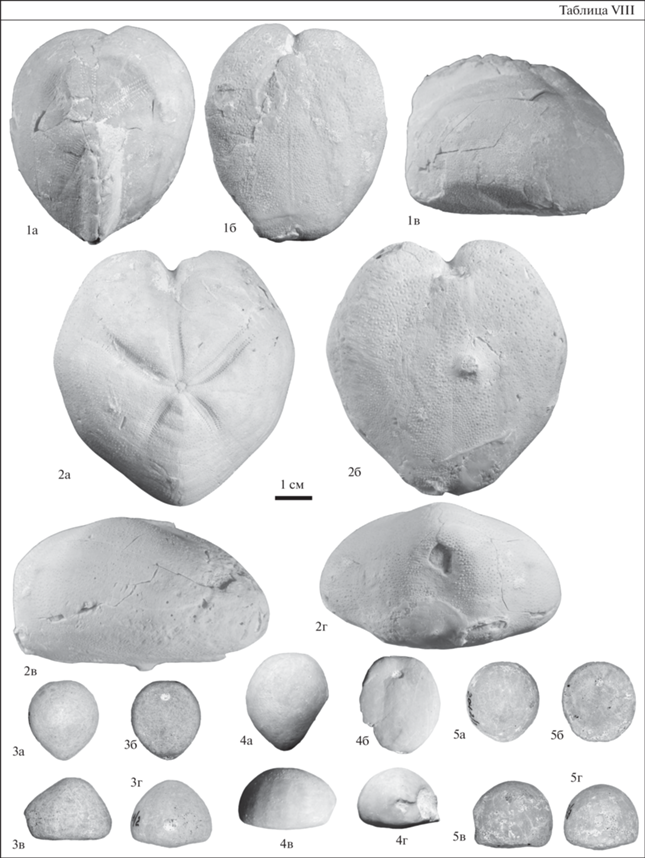
Таблица IХ. Губки сантона и брахиоподы турона–коньяка из разреза Коммунар. Длина масштабной линейки 1 см, кроме особо отмеченных случаев. 1 – Hexactinosa gen., экз. SSU PEM № 122/6027, скелет выполнен марказитом: 1а – сбоку, 1б – снизу; нижний сантон, слой 7; 2 – Coeloptychium sp. ind., экз. SSU PEM № 58/03, скелет в карбонатной породе, вид снизу; нижний сантон, верхняя часть слоя 7; 3 – Botryosella sp. ind., экз. SSU PEM № 122/6644, скелет выполнен марказитом, сбоку; нижний сантон, слой 7; 4 – Porosphaera sp., экз. SSU KDV № 311/1/3-2, морфология поверхности скелета изображена: 4a – в обратно отраженных электронах, 4б – во вторичных электронах; нижний коньяк, средняя часть слоя 4; 5 – Concinnithyris protobesa Sahni, экз. SSU IEI № 251/2/7: 5a – брахиальная створка, 5б – педальная створка, 5в – сбоку, 5г – лобный край; средний–верхний турон, слой 3; 6 – Cretirhynchia subplicata (Mantell), экз. SSU IEI № 251/2/31: 6a – брахиальная створка, 6б ‒ сбоку, 6в – педальная створка, 6г – лобный край; нижний коньяк, слой 4; 7 – Orbinrhynchia cuvieri (d’Orbigny), экз. SSU IEI № 251/2/22: 7a – брахиальная створка, 7б – педальная створка, 7в – сбоку, 7г – лобный край; средний–верхний турон, слой 3; 8 – Gemmarcula sp., экз. SSU IEI № 251/2/13: 8a – брахиальная створка, 8б – педальная створка, 8в – сбоку, 8г – лобный край; средний–верхний турон, слой 3; 9 – Orbirhynchia dispansa Pettitt, экз. SSU IEI № 251/2/23: 9a – брахиальная створка, 9б – педальная створка, 9в – сбоку, 9г – лобный край; средний–верхний турон, слой 3; 10 – Orbirhynchia sp. 1, экз. SSU IEI № 251/2/15: 10a – брахиальная створка, 10б – педальная створка, 10в – сбоку, 10г – лобный край; средний–верхний турон, слой 3.

Иноцерамы из слоя 4, кроме Inoceramus lusatiae, являются членами клады Cremnoceramus. В нижней части слоя это C. deformis erectus (Meek) (табл. II, фиг. 1, 4, 5 ), а в верхней части переходные формы между Cremnoceramus crassus inconstans (Woods) и C. crassus crassus (Petrascheck) (табл. II, фиг. 2). Все кремноцерамы указывают на то, что слой 4 представляет собой нижнюю часть нижнего коньяка, вплоть до уровня с C. crassus inconstans, который относится к низам выше расположенной иноцерамовой зоны C. crassus crassus (Walaszczyk, Wood, 1998).
Слой 6, несмотря на то что только верхняя его треть охарактеризована последовательностью иноцерамов, относится к среднему и верхнему коньяку. Положение подошвы среднего коньяка не может быть точно установлено, поскольку находка Inoceramus gibossus Schlüter (табл. II, фиг. 7 ) указывает на пограничные отложения нижнего и среднего коньяка. Этот вид появляется в конце раннего коньяка и распространен выше, во всем среднем коньяке (Walaszczyk, Wood, 2018). В самой верхней части слоя 6 найдены Sphenoceramus subcardissoides (Schlüter) (табл. III, фиг. 1) и Sphenoceramus digitatus Heine (non J. de C. Sowerby) (табл. III, фиг. 5 ). Оба таксона характеризуют верхний коньяк северных регионов Европы (Walaszczyk et al., 2017). На южной периферии этого региона они постепенно замещаются представителями родов Magadiceramus и Platyceramus. Таким образом, интервал, охватывающий уровни обр. 22–23, рассматривается как верхнеконьякский. Часть слоя 6, в интервале между обр. 16 и 21 и ниже первого появления Sph. subcardissoides, не может быть точно датирована, так как Platyceramus sp. (табл. I , фиг. 7 ) и вольвицерамиды (табл. III, фиг. 4 ; табл. IV) распространены как в среднем, так и верхнем коньяке (Walaszczyk et al., 2017).
Один экземпляр Sphenoceramus pachti (Arkhangelsky) встречен в верхней части слоя 7 (табл. V, фиг. 6 ). Несмотря на то что вид распространен на протяжении всего раннего сантона (Seitz, 1965), его находки редки в верхах нижнесантонских отложений. Последовательность в распространении иноцерамов дает возможность судить о присутствии в изучаемом разрезе верхней части среднего турона и большей части верхнего турона, включая его терминальные интервалы. Коньякские отложения, с учетом затруднений в определении границы среднего и верхнего коньяка, представлены всеми тремя подъярусами.
Аммониты (рис. 1). Находки аммоноидей Lewesiceras mantelli Wright et Wright, Scaphites geinitzii d’Orbigny, Hyphantoceras reussianum (d’Orbigny) и Sciponoceras bohemicum bohemicum (Fritsch) указывают на позднетуронский возраст отложений средней и верхней части третьего слоя (табл. III, фиг. 3, 6, 7 ; табл. VI, фиг. 1). В частности, вид L. mantelli известен из верхнего турона Европы и южной периферии Европейской России (Houša, 1967; Wiese et al., 2004; Сельцер, Иванов, 2010; Сельцер, 2018а; Kennedy, Gale, 2015; Kennedy, Kaplan, 2019). Вид S. geinitzii встречается в одновозрастных отложениях Западной Европы, Крыма, Северного Кавказа, Прикаспийского региона и Поволжья (Kaplan, Schmid, 1988; Сельцер, Иванов, 2010; Kennedy, Kaplan, 2019). Отметим находку фрагмента оборота H. reussianum (d’Orbigny) с характерной скульптурой из чередующихся тонких высоких ребер с шиповидными выступами (Kaplan, Schmid, 1988). В Поволжье эта форма установлена из одновозрастных отложений других структурно-фациальных зон (Первушов и др., 2017; Сельцер, 2020). В западноевропейских разрезах выделяется секвенция “Hyphantoceras Sequence”, отражающая высокое стояние моря позднего турона, способствовавшее расселению этих моллюсков (Wiese et al., 2004), что, вероятно, проявилось и в юго-восточных районах Восточно-Европейской палеобиогеографической провинции.
В слое 4 найден аммонит Puzosia muelleri de Grossouvre (табл. VI, фиг. 2) (Сельцер, 2018б). В Вестфалии эта форма прослеживается не выше нижнеконьякской аммонитовой зоны Peroniceras tridorsatum или иноцерамовой зоны Cremnoceramus deformis (Kaplan, Kennedy, 1994). Это позволяет утверждать, что верхняя часть слоя 3 может быть датирована поздним туроном, а нижняя часть слоя 4 – ранним коньяком.
Белемниты (рис. 1). В интервале слоев 2 и 3 встречены единичные Goniocamax sp. cf. intermedius (Arkhangelsky) (табл. III, фиг. 2) – типичные формы среднего и верхнего турона востока Европейской палеобиогеографической области (Найдин, 1964; Christensen, 1997; Košťák, 2004).
Ростры белемнитов Actinocamax verus fragilis Arkhangelsky и первые Belemnitella shmidi Christensen et Schulz (табл. V, фиг. 3, 5 ) встречены в верхней части слоя 7 (Первушов и др., 2020б; Сельцер и др., 2020). Отметим, что с началом сантона связывается появление представителей рода Belemnitella (Christensen, 1986, 1991, 1997; Christensen, Schulz, 1997), поэтому находка ростра типичной белемнителлы указывает на принадлежность верхней части слоя 7 к основанию сантона.
Из слоя 8 происходят ростры Belemnitella praecursor praepraecursor Naidin и редкие Actinocamax verus fragilis Arkhangelsky (рис. 3; табл. V, фиг. 4 ). Из верхней части слоя 8 отобраны ростры позднесантонского Actinocamax verus cylindricus Makhlin. Находка B. praecursor praepraecursor Naidin (Первушов и др., 2020а, 2020б) заслуживает отдельного рассмотрения. По мнению В.К. Христенсена, этот подвид должен рассматриваться в объеме Belemnitella praecursor s.s. Stolley, поскольку Д.П. Найдин не дал его подробного описания (Christensen, 1986, 1991). В разрезах Донбасса, откуда приведено описание подвида B. praecursor praepraecursor, он встречен ниже типичных praecursor, как бы предваряя их (Найдин, 1974). Представители подвида praepraecursor отличаются малой длиной ростров и малыми значениями щелевого угла, что, на наш взгляд, подчеркивает самостоятельность данного подвида. В действующей стратиграфической схеме верхнемеловых отложений Восточно-Европейской платформы подвид B. praecursor praepraecursor Naidin является одним из маркеров верхнего сантона (лона patootensis/praecursor praepraecursor) (Олферьев, Алексеев, 2005).
В кровле слоя 8 и в слое 9 встречены раннекампанские ростры Belemnitella mucronatiformis Jeletzky (Первушов и др., 2020а, 2020б). Можно предположить, что верхняя часть конденсированных образований слоя 8 формировалась уже в раннекампанское время, что подтверждается находками бентосных фораминифер.
Иглокожие (рис. 1). Среди иглокожих наиболее представителен комплекс морских ежей. В подошве слоя 3 найден единственный экземпляр Phymosoma sp. В нижней части разреза, в 1.8 м выше кровли альба, установлено первое появление представителей Micraster sp., совместно с Conulus subrotundus Mantell (табл. VII, фиг. 1, 2), который характерен для всего средне-верхнетуронского интервала рассматриваемого разреза. Наиболее значимыми являются данные о вертикальном распространении представителей Micraster. Первые находки M. corbovis Forbes (табл. VII, фиг. 3) приурочены к средней части слоя 3, отвечающего подзоне LC5b верхнего турона по БФ. Распространение вида прослежено вплоть до границы турона–коньяка, последние находки этих форм приурочены к верхней части слоя 3, отвечающей подзоне LC5с верхнего турона по БФ. Здесь же отмечены первые находки M. cortestudinarium (Goldfuss), встречающиеся и выше по разрезу, в мергелях нижнего коньяка. На данный момент совместное нахождение M. corbovis Forbes и M. cortestudinarium (Goldfuss) в терминальной части верхнетуронских отложений вольских разрезов не установлено, что позволяет сделать вывод о последовательной смене этих позднетуронских представителей Micraster. Подобная ситуация характерна и для сопредельных территорий, а последовательность видов M. corbovis Forb.–M. cortestudinarium (Goldfuss) отражает позднетуронский–раннеконьякский этап развития рода (Калякин, 2019).
Комплекс иглокожих в интервале слоев 4 и 5, в нижней и средней частях слоя 6 значительно беднее предшествующего туронского. В нижней части слоя 4 достоверно установлены панцири M. cortestudinarium (Goldfuss) (табл. VII, фиг. 4 ). Вверх по разрезу, в слоях 4, 5 и в нижней части слоя 6, встречены разрозненные неопределимые фрагменты панцирей эхиноидей. В нижних 1.5 метрах слоя 4, в обр. 8–10 установлены отдельные членики морских лилий Bourgueticrinus sp. В обр. 8–9 встречены остатки офиур. Маргинальные таблички морских звезд семейства Goniasteridae выделены в микрофаунистических обр. 8–10, 12, 14, 18–19. Количество иглокожих в микрофаунистических пробах значительно уменьшается по сравнению с ранее рассмотренным интервалом туронских образований. Особенности вертикального распространения криноидей, астероидей и офиуроидей в туроне–коньяке Вольской структурной зоны прямо соотносятся с установленными ранее закономерностями в разрезах Чухонастовка, Каменный Брод, Мирошники и Сплавнуха. Максимальная концентрация остатков иглокожих отмечена в средне-верхнетуронской части разрезов, а в пробах из пород коньяка их содержание значительно снижается. Этот уровень концентрации иглокожих рассматривается как маркирующий, который прослеживается в составе средне-верхнетуронских отложений Нижнего Поволжья (Калякин и др., 2018; Первушов и др., 2019; Pervushov et al., 2019).
Слой 6 и нижняя часть слоя 7 охарактеризованы совместным распространением M. coranguinum (Leske) и M. rogalae Nowak (табл. VIII, фиг. 1, 2), а в обр. 24 определены гониастериды. Данный комплекс иглокожих наименее представителен в разрезе Коммунар, но при этом он является одним из наиболее информативных. Распространение двух видов Micraster в относительно маломощном интервале разреза, соотнесенном с подзонами по БФ LC8a и LC8b, отражает коньякский и, возможно, раннесантонский этап развития рода (Калякин, 2019).
В обр. 27 (верхняя часть слоя 7) установлены редкие разрозненные членики Bourgueticrinus sp. и таблички морских звезд семейства Goniasteridae, последние выделены и в обр. 29.
Морские ежи в слое 8 представлены единичным фрагментированным панцирем Micraster sp. В обр. 30 определены отдельные членики Bourgueticrinus sp. и таблички морских звезд семейства Goniasteridae. В базальной части нижнекампанских отложений (слой 9) первыми появляются Galeola ex gr. senonensis (d’Orbigny) и Offaster pilula (Lamarck) (табл. VIII, фиг. 3–5 ). На этот интервал приходятся наибольшие частоты их встречаемости, при этом G. ex gr. senonensis встречен только в базальной части кампанских отложений, а O. pilula распространен вплоть до границы нижнего–верхнего кампана. В обр. 31 встречены членики Bourgueticrinus sp. и таблички неопределимых гониастерид.
В туронском–сантонском интервале разреза Коммунар отмечена последовательная, без существенных лакун, смена представителей филогенетического ряда Micraster. На этом основании предлагается выделение следующих биостратиграфических зон по морским ежам, которые прослеживаются в Поволжье и соседних регионах (рис. 2).
Средний–верхний турон. Филозона Micraster corbovis. Характерными таксонами являются Micraster corbovis Forbes, Echinocorys gravesi (Desor), Conulus subrotundus Mantell. Фаунистическая характеристика зоны приведена на рис. 1. Стратотип зоны – разрез Коммунар, г. Вольск. Распространение зоны прослежено на большей части Среднего и Нижнего Поволжья от разреза близ г. Новоульяновска (Ульяновская область) до разреза Каменный Брод (Волгоградская область). Сопоставление с биостратиграфическими подразделениями по другим группам беспозвоночных приведено на рис. 2. Micraster corbovis известен из среднего–верхнего турона Англии (зоны Terebratulina lata и Sternotaxis plana), верхнего турона Польши, окрестности г. Ополе (зона Inoceramus perplexus), Франции, Германии, Донбасса, Волыно-Подольской плиты, Северного Кавказа, Мангышлака (верхнетуронские–коньякские отложения разреза Бесакты), Копетдага, турона Воронежской антеклизы (разрез Копанище, Павловский свод) и из разреза на р. Тузлов Ростовской области (Москвин, Эндельман, 1972; Найдин, 2004; Olszewska-Nejbert, 2007; Smith, Wright, 2012; Калякин, 2019).
Верхний турон–средний коньяк. Филозона Micraster cortestudinarium. Характерным таксоном является Micraster cortestudinarium (Goldfuss). Фаунистическая характеристика зоны приведена на рис. 1. Стратотип зоны – разрез Коммунар, г. Вольск. Распространение зоны прослежено в пределах Вольской впадины, а также южнее г. Саратова (разрез Нижняя Банновка). Находки Micraster cortestudinarium известны из верхнего турона–коньяка Англии (зона M. cortestudinarium), Копетдага, Донбасса, Северного Кавказа, нижнего коньяка (зоны Cremnoceramus deformis deformis–C. crassus crassus) Мангышлака, Польши (зоны C. deformis erectus и C. waltersdorfensis hannovrensis, C. crassus inconstans и C. deformis deformis/C. crassus crassus), коньяка Франции, Германии, Северной Испании (Москвин, Эндельман, 1972; Olszewska-Nejbert, 2007; Smith, Wright, 2012; Калякин, 2019).
Верхний коньяк–нижний сантон. Филозона Micraster coranguinum–Micraster rogalae. Характерными таксонами являются Micraster coranguinum (Leske), Micraster rogalae Nowak. Фаунистическая характеристика зоны приведена на рис. 1. Стратотип зоны – разрез Коммунар, г. Вольск. Распространение зоны прослежено в пределах Вольской впадины и южнее г. Саратова (разрезы Широкий Буерак, Пудовкино). Находки Micraster coranguinum известны из среднего коньяка–среднего сантона Англии (вид-индекс зоны Micraster coranguinum), Франции, Германии, верхнего коньяка Мангышлака (зона Magadiceramus subquadratus), Северного Кавказа, Копетдага, Грузии, коньяка Восточной Украины и Донбасса, сантона Польши. Micraster rogalae встречается в среднем–верхнем коньяке Галиции (Западная Украина), верхнем коньяке–нижнем сантоне Мангышлака, среднем сантоне Южной Польши, верхней части нижнего сантона–нижней части верхнего сантона Северной Германии (Lügerdorf), сантоне Бельгии (Москвин, Эндельман, 1972; Olszewska-Nejbert, 2007; Smith, Wright, 2012; Калякин, 2019).
В основании нижнекампанской толщи находки Micraster удовлетворительной сохранности отсутствуют, что не позволяет обосновать выделение филозоны. Более того, в подстилающих отложениях верхнего сантона также отсутствуют находки морских ежей приемлемой сохранности. Таким образом, в данном случае при выделении зон по морским ежам в нижнем кампане нарушается принцип их смыкаемости. Однако уже с основания кампанской толщи в породе встречены Galeola ex gr. senonensis (d’Orbigny) и Offaster pilula (Lamarck). Первое появление и наибольшая частота встречаемости этих форм отмечены именно в слое 9, в базальной части нижнекампанских отложений. O. pilula (Lamarck) является видом-индексом одноименной нижнекампанской зоны в Западной Европе. В некоторых регионах рассматриваемый интервал разработан более детально. В частности, в Германии выделены последовательные зоны по морским ежам: O. pilula, O. pilula/G. senonensis и G. senonensis (Neumann et al., 2002). В начале кампана в Западной Европе проявился трансгрессивный эпизод, известный как pilula-трансгрессия, за которой последовала еще одна senonensis-трансгрессия. Д.П. Найдин считал, что на территории Восточно-Европейской платформы одновременно с pilula-трансгрессией развивалась так называемая “птеривая” трансгрессия (Найдин, 1995; Волков, Найдин, 1998; Беньямовский, 2005). Многочисленные находки O. pilula (Lamarck) в кампанской толще в вольских разрезах являются отражением масштабного раннекампанского биособытия – pilula-трансгрессии. В то же время находки G. ex gr. senonensis (d’Orbigny) в базальной части нижнекампанских отложений этих же разрезов позволяют предположить, что так называемая senonensis-трансгрессия также имела место в пределах региона. Однако временной интервал предполагаемых событий и объем каждого из них требуют детализации. Учитывая эти данные, авторы предлагают выделить в нижнем кампане биозону Offaster pilula (Первушов и др., 2020б).
Нижний кампан. Провинциальная зона распространения вида Offaster pilula. Характерными таксонами являются Offaster pilula (Lamarck), Galeola ex gr. senonensis (d’Orbigny), Echinocorys ovata (Leske), E. marginata (Goldfuss), E. turrita Lambert, Isomicraster gibbus (Lam.), I. faasi Rouchadze, C. matesovi Moskvin. Фаунистическая характеристика зоны приведена на рис. 1. Стратотип зоны – разрез Коммунар, г. Вольск. Распространение зоны прослежено в пределах Вольской и Хвалынской (Апалиха) впадин. Виды Offaster pilula (Lamarck) и Galeola senonensis (d’Orbigny) известны из нижнего кампана Европы (Англии, Франции, Германии, Польши), где установлены одноименные зоны, а также из нижнего кампана Северного Кавказа. Другие характерные виды морских ежей также широко распространены в нижнекампанских отложениях Европы (Англии, Франции, Германии, Бельгии, Польши), а также Северного Кавказа и Мангышлака (Калякин, 2017). Установленная в Западной Европе последовательность нижнекампанских биостратиграфических зон (O. pilula–O. pilula/G. senonensis–G. senonensis), событийно отражающая смену pilula-трансгрессии senonensis-трансгрессией, в пределах Поволжья не выявлена. Более того, O. pilula (Lamarck) имеет более широкое стратиграфическое распространение в вольских разрезах, чем G. ex gr. senonensis (d’Orbigny).
Большинство одноименных зон ранее были установлены на территории Западно-Европейской плиты (Neumann et al., 2002; Olszewska-Nejbert, 2007; Smith, Wright, 2012). В пределах же Русской плиты зоны по морским ежам не описывались и полноценно не охарактеризованы. Соотношение зон по морским ежам, установленных в Западной Европе и предлагаемых для европейской части России, требует последующей детализации и уточнения.
Анализ крупной фракции (более 0.4 мм) микрофаунистических проб позволяет проследить распространение микроразмерных элементов представителей разных групп иглокожих в разрезе (Калякин и др., 2018; Pervushov et al., 2019). Для вольских разрезов подобный подход использован впервые, а полученные здесь результаты в значительной мере дополняют данные о площадном и стратиграфическом распространении иглокожих. В слое 3 отдельные членики морских лилий Bourgueticrinus sp. отмечены в обр. 3–7. Разрозненные маргинальные таблички неопределимых морских звезд (гониастерид) установлены в обр. 2–7; с небольшими перерывами они встречаются почти по всему разрезу, вплоть до обр. 31 (рис. 1). Интересным является наличие в обр. 3–7 фрагментов офиур. Установленный в разрезе Коммунар комплекс морских лилий и морских звезд сопоставим с сообществом иглокожих, выделенным ранее из расположенных южнее разрезов Чухонастовка и Каменный Брод (Калякин и др., 2018; Pervushov et al., 2019). Отличительной чертой комплекса разреза Коммунар является полное отсутствие представителей пентакринид, в частности Nielsenicrinus carinatus (Roemer), характерных для синхронных интервалов во многих разрезах юго-западной части Ульяновско-Саратовского прогиба (Первушов и др., 2019; Pervushov et al., 2019).
Брахиоподы (рис. 1, табл. IX ). В результате обработки объемных проб, отобранных из нижней части слоя 3 (подзона LC4a, средний–верхний турон), было получено большое количество раковин брахиопод, преимущественно деформированных и частично разрушенных. Среди них определены Orbirhynchia dispansa Pettitt, O. cf. dispansa Pettitt, O. cuvieri (d’Orbigny), Orbirhynchia sp., Gibbithyris grandis Sahni, Concinnithyris protobesa Sahni, C. cf. subundata J. Sowerby, Gemmarcula sp., Cretirhynchia sp., Gyrosoria lata (Etheridge), Terebratulina striatula (Mantell). В количественном отношении и по таксономическому разнообразию наиболее представительны ринхонеллидные орбиринхии. Представители Orbirhynchia dispansa Pettitt, Orbirhynchia sp., Gibbithyris grandis Sahni, Gyrosoria lata (Etheridge) и Terebratulina striatula (Mantell) ранее встречены в средне-верхнетуронских отложениях разрезов Каменный Брод, Чухонастовка-2 и Нижняя Банновка.
По количеству и таксономическому разнообразию комплекс брахиопод нижней части слоя 4 (подзона LC6a, верхний турон–нижний коньяк) заметно беднее ранее рассмотренного. Определены брахиоподы Orbirhynchia dispansa Pettitt, Orbirhynchia sp., Cretirhynchia subplicata (Mantell), Cretirhynchia sp., Gyrosoria lata (Etheridge), Gibbithyris ellipsoidalis Sahni.
В турон-коньякское время в пределах Европейской палеобиогеографической области были широко распространены представители родов Gibbithyris Sahni, Concinnithyris Sahni, Najdinothyris Katz, Kingena Davidson, Orbirhynchia Pettitt и Cretirhynchia Pettitt. Предполагаем, что таксономическое разнообразие туронских–коньякских сообществ брахиопод восточных районов этой палеобиохории значительно менее представительно. На территории современного Поволжья в отложениях турона и коньяка не отмечены находки представителей надсемейств Megathyridoidea и Platidioidea, подсемейств Capillithyridinae, Trigonoseminae и Dalligadinae. На западе Европейской палеобиогеографической области представители этих групп брахиопод в отложениях турона и коньяка нередки. В работе (Gaspard, 1997) упоминается зона Terebratulina lata, характерная для среднего турона Англии. Впоследствии (Sklenář, Simon, 2009) была отмечена условность использования этого вида в качестве биостратиграфического маркера для указанного стратиграфического диапазона, в связи с установленным более значительным его вертикальным распространением. На территории Западной Европы комплексы брахиопод из отложений турона менее изучены, чем из более поздних образований. В частности, в работах (Surlyk, 1972, 1984) описаны комплексы маастрихтских брахиопод Дании, которые получены при использовании методики извлечения фоссилий из массовых проб породы.
В верхней части слоя 7 брахиоподы редки: несколько экземпляров Terebratulina cf. striatula (Mantell), Cretirhynchia sp., Orbirhynchia sp., Concinnithyris sp. Представители Terebratulina cf. striatula (Mantell) и Cretirhynchia sp. ранее были установлены из нижнесантонского “губкового” горизонта разреза Пудовкино.
Спонгиофауна (рис. 1, табл. IX ). В песчанистых мергелях слоев 2 и 3 (банновская свита) найдены единичные мелкорослые гексактинеллиды Plocoscyphia sp. и Guettardiscyphia sp. Это типичные элементы туронского спонгиокомплекса, который распространен в более представительном составе в южной части Поволжья (местонахождения Красный Яр-1, Каменный Брод, Чухонастовка и Мирошники-1) (Первушов и др., 2019; Pervushov et al., 2019). Фоссилии в разной степени фосфатизированы и обычно представлены фрагментами.
В карьере Коммунар в мергелях банновской свиты установлены известковые спикульные губки Porosphaera. Очень мелкие сферические скелеты выделены в микрофаунистических пробах по всему интервалу туронских отложений, в обр. 2–14. В нижней части рассматриваемого интервала (подзона LC4a) большая часть фоссилий отличается плохой или частичной сохранностью, что обычно для мелких скелетов Porosphaera. Несколько губок сферической формы определены как Porosphaera cf. globularis. В верхней части туронского интервала (подзоны LC4b и LC5b) выделено более 10 скелетов Porosphaera sp. более крупных размеров, до 0.5–3.5 мм. Ранее находки известковых спикульных губок из карбонатных пород среднего турона были сделаны в разрезах юго-западной части Ульяновско-Саратовского прогиба (Первушов, Худяков, 2020), где на разрезах Чухонастовка-2 и Филин собрано большое количество крупных скелетов Porosphaera из интервалов зоны БФ LC5с (верхний турон).
Ранее находки губок из “немых” интервалов карбонатных пород коньяка не были известны. В юго-западной части Ульяновско-Саратовского прогиба в верхах среднего коньяка прослежены уровни равномерно рассеянного и конденсированного распространения скелетов кремневых губок. Субавтохтонные захоронения гексактинеллид прослежены в разрезах Каменный Брод (Pervushov et al., 2019), Меловатка-1, Мирошники и Вишневое (Олферьев и др., 2008; Первушов и др., 2019). На остальной территории Поволжья в результате предсантонского размыва отложения среднего и верхнего коньяка отсутствуют.
В основании слоя 4 (обр. 8, подзона LC6a) и в средней части слоя 5 (обр. 14, подзона LC6a) обнаружены известковые губки Porosphaera sp. (табл. IX , фиг. 4 ). Шаровидные скелеты белого цвета, размером 1.0–1.5 мм выделены из порошков микрофаунистических проб. В кровле слоя 6 (зона LC7, средний коньяк) найден единственный сильно окатанный скелет Porosphaera sp. диаметром 10–13 мм. Из мергелей подзоны LC6a (обр. 8, 9) выделены скелеты Porosphaera sp. диаметром 1.0–1.5 мм. В интервале среднего–верхнего коньяка найден сильно окатанный скелет Porosphaera sp. диаметром 10–13 мм.
В верхах слоя 6 и в нижней части слоя 7 кремневые губки равномерно распространены и представлены отпечатками тонкостенных форм или крупными дырчатыми желто-коричневыми стяжениями фосфатного и марказитового состава. Плохая первичная сохранность скелетов не позволяет определить таксономическую принадлежность фоссилий. Установлено много крупных форм Plocoscyphia sp., Camerospongia sp., Microblastium sp., Sororistirps sp., Sporadoscinia sp., Etheridgia sp., мелких фрагментов толстостенных Cephalites sp. и менее разнообразных представителей Hexactinosa: Guettardiscyphia sp., ? Labyrintholites sp., Eurete sp.
Скелеты гексактинеллид распространены и в верхней части слоя 7, в интервале 1.0–1.5 м ниже его кровли. Фоссилии окатаны и часто представлены в виде плотных окатышей со следами биоэрозии. Наряду с крупными формами полной сохранности на карбонатно-фосфатном цементе, распространены фрагменты фосфатизированных скелетов. Губки захоронены в пределах биотопа обитания, но окатаны и до некоторой степени сцементированы вмещающим их скелеты осадком. По видовому разнообразию и в количественном отношении преобладают представители Lychniscosa: Cephalites sp., Ventriculites sp., Microblastium sp., Sporadoscinia sp., Sororistirps sp., Camerospongia sp., Etheridgia sp., Plocoscyphia sp., а среди Hexactinosa определены Leptophragmidae gen. ind., Guettardiscyphia sp., Botryosella sp., Eurete sp., Hexactinosa gen. nov.
В Вольской структурной зоне “губковый” горизонт (слой 8) полно представлен только в карьере Коммунар. Спонгиокомплекс, в составе которого доминировали гексактинеллиды, формировался на протяжении нескольких фаз, о чем можно суть по фоссилизации и степени сохранности скелетов, совместном захоронении форм, обитавших в разных гидродинамических обстановках водной среды (Первушов и др., 2020а, 2020б). В составе спонгиокомплекса установлены формы, которые рассматриваются как новые, в том числе и как переходные виды между известными сантонскими и кампанскими представителями родов. Здесь многочисленны губки Sororistirps, занимающие в филогенетической линии промежуточное положение между раннесантонскими (S. radiatum Mantell) и кампанскими (S. tubiforme Schrammen [= Ventriculites infundibuliforme]) представителями рода.
В базальном горизонте сенгилеевской свиты (слой 9) встречены мелкие фосфатизированные и окатанные фрагменты скелетов гексактинеллид, которые переотложены из подстилающих пород. Здесь же установлены три экземпляра известковых губок Porosphaera cf. nuciformis (Hagenow) диаметром 10–12 мм (Первушов, Худяков, 2020). В подошве слоя 10 найдено несколько экземпляров известковых губок со следами биоэрозии. Двояковыпуклые формы диаметром до 12–15 мм рассматриваются как Porosphaera sp. 1, а сферические скелеты диаметром до 12 мм, со многими бугорками крепления опорных спикул выделены как Porosphaera sp. 2.
Кремневые и тем более известковые губки остаются наименее изученными группами позднемеловых беспозвоночных. Лимитирующими факторами их использования при биостратиграфических построениях являются стенофациальный характер кремневых губок, приуроченность их поселений к участкам придонных течений и неполная сохранность скелетных форм или спикульной решетки. Представления о “губковых” слоях как маркирующих образованиях следует считать несостоятельными, поскольку эти диахронные образования установлены в отложениях всех ярусов верхнего мела Поволжья.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Результаты работ предшествующих исследователей верхнемеловых отложений Поволжья (Морозов, 1962; Морозов, Орехова, 1969; Морозов, Бондарева, 1970) способствовали формированию устойчивого представления о карбонатных породах турона–коньяка как о едином геологическом теле. На рубеже веков в субрегиональной стратиграфической схеме верхнемеловых отложений Среднего и Нижнего Поволжья (Стратиграфическая…, 2004; Олферьев, Алексеев, 2005) выделен губкинский надгоризонт, соответствующий стратиграфическому диапазону турона–коньяка. Но выделение банновской и вольской свит (горизонтов) на основании литологически различных интервалов или явно прослеживаемых поверхностей стратиграфических перерывов невозможно во многих изученных разрезах. Установление в верхней части губкинского горизонта собственно коньякских образований оказалось возможным при изучении фораминифер (Барышникова, 1955, 1956, 1959; Барышникова и др., 1961): был определен новый вид их бентосных форм – Protostensioeina emscherica (Baryshnikova), который в настоящее время является зональной формой нижнего коньяка (Беньямовский, 2008а).
В Вольской структурной зоне, в карьере Коммунар, доступен для изучения наиболее стратиграфически полный и палеонтологически охарактеризованный на территории Поволжья интервал коньякских–нижнесантонских отложений. Здесь по литологии пород и их палеонтологической характеристике выделено несколько последовательных интервалов. Слои 2 и 3 отнесены к банновской свите (средний–верхний турон), слои 4–6 – к вольской свите (все три подъяруса коньяка). В качестве пограничного интервала терминального коньяка–нижней части нижнего сантона рассматривается нижняя часть слоя 7. Верхняя часть слоя 7 отнесена к нижнему (можжевелоовражная свита), а слой 8 – к верхнему сантону (мезинолапшиновская свита). Слой 9 – базальный горизонт, а слой 10 – нижняя часть сенгилеевской свиты (нижний кампан).
Материалы биостратиграфического расчленения нижней части карбонатных пород разреза Коммунар сопоставлены с результатами изучения разрезов губкинского горизонта, расположенных в юго-западной части Ульяновско-Саратовского прогиба: стратотипа банновской свиты (турон, Нижняя Банновка) и предполагаемого гипостратотипа вольской свиты (коньяк, Каменный Брод). При корреляции этих разрезов прослеживается увеличение мощности отложений почти всех биостратонов в южном направлении. Но губкинский горизонт в каждом из этих разрезов охарактеризован разными подзонами БФ, что обусловлено развитием в туронское–сантонское время локальных структур, в пределах которых были расположены рассматриваемые разрезы. Почти во всех разрезах прослеживаются подзоны БФ LC4a, LC4b, LC5a (за исключением разреза Коммунар) и зоны LC6 и LC7.
На основании комплексной палеонтологической характеристики интервала вольской свиты в разрезе Каменный Брод, отличающегося максимальной (около 35 м) мощностью в регионе (мощность вольской свиты в разрезе Коммунар – около 10 м, в разрезе Большевике – 6 м), этот разрез было предложено рассматривать в качестве гипостратотипа вольской свиты (Pervushov et al., 2019). Но в разрезе Каменный Брод установлены только образования нижнего (зона LC6) и среднего (зона LC7) коньяка, а более поздние образования уничтожены эрозией, которая предшествовала сантонскому осадконакоплению.
Детальное биостратиграфическое расчленение разреза Коммунар основано на распределении бентосных фораминифер. Возраст выделенных здесь биостратиграфических подразделений по БФ согласуется с региональными шкалами по БФ, составленными для территорий Европейской палеобиогеографической области в целом (Беньямовский, 2008а, 2008б; Vishnevskaya et al., 2018) и для Предкарпатского прогиба в частности (Walaszczyk et al., 2016). На рисунке 1, где проведено сопоставление литологической колонки разреза Коммунар с биостратиграфическими подразделениями по БФ, красная (в электронной версии) штрихпунктирная линия отображает представления авторов настоящей работы о положении границы верхнего турона–нижнего коньяка по данным иноцерамов и бентосных фораминифер (Walaszczyk et al., 2013). Это мнение основано на результатах комплексного изучения разрезов верхнемеловых отложений Мангышлака и дополняет информацию по БФ, представленную в работах В.Н. Беньямовского (2008а, 2008б).
В разрезе Коммунар, несмотря на существенно сокращенную мощность пород турона–нижнего сантона, прослежена значительная последовательность подзон по бентосным фораминиферам, в частности установлены подзоны БФ LC5b и LC5c верхнего турона, которые неизвестны в разрезах юго-западной части Ульяновско-Саратовского прогиба. Комплексы бентосных фораминифер подзон LC8a и LC8b (верхний коньяк–нижний сантон) впервые установлены на территории Поволжья. Предположение о стратиграфическом перерыве между подзоной LC6b и зоной LC7 по БФ (нижний–средний коньяк) основано на литологических признаках в строении этого разреза.
Тем не менее, в туронских–нижнекампанских отложениях рассматриваемого разреза на основе наиболее детальных зональных схем по бентосным фораминиферам, белемнитам и иноцерамам установлены неоднократные хиатусы. Лишь переходные интервалы верхнего турона–нижнего коньяка и верхнего коньяка–нижнего сантона охарактеризованы непрерывной последовательностью зон по бентосным фораминиферам, иноцерамам и морским ежам. В разрезе Коммунар комплекс пород терминального коньяка–нижнего сантона (подзона LC8b по БФ) рассматривается как непрерывный, стратиграфически наиболее полный среди известных разрезов на территории Поволжья.
В разрезе Коммунар прослежены изменения в составе палеобиоценозов на протяжении туронского–раннекампанского времени, что позволило сравнить состав синхронных бентосных поселений и проследить особенности расселения их представителей в пределах современного Поволжья. В частности, при анализе микрофаунистических проб из разреза Коммунар в мергелях среднего–верхнего турона выделен уровень концентрации скелетных мелкоразмерных элементов иглокожих. Ранее предполагалось, что северная граница ареала морских лилий, морских звезд и офиур в туронское время находилась южнее широты разреза Сплавнуха (Калякин и др., 2018). Но сообщество иглокожих, обитавших в позднетуронское время на юго-западе Ульяновско-Саратовского прогиба, отличалось бóльшим разнообразием и количеством.
Туронская трансгрессия способствовала проникновению в акваторию региона разнообразных представителей бентоса. Предполагается, что максимум трансгрессии совпал с завершением туронского века (Найдин и др., 1986; Sahagian et al., 1996; Haq, 2014) или началом раннего коньяка (Pervushov et al., 2019) и сопровождался возрастанием биоразнообразия морской биоты в целом (Невесская, 1999). С началом коньякского времени связывается устойчивая тенденция к углублению бассейна, обусловленная проявлением региональной тектоники в юго-западной части Ульяновско-Саратовского прогиба. При этом разнообразие и численность моллюскового сообщества заметно сократились. Регрессивные тенденции в развитии бассейна в конце среднего коньяка–начале раннего сантона способствовали широкому площадному распространению губково-моллюсковых поселений в условиях более прохладных вод с периодическим проявлением активной гидродинамики.
В разрезе Коммунар наиболее полно представлен спонгиокомплекс позднего сантона. Положение этого “губкового” горизонта биостратиграфически достоверно обосновано, он разнообразен в таксономическом отношении и многочислен. В составе сообщества кремневых губок установлены ранее неизвестные в Поволжье формы на уровне вида и рода. Изучение новых видов в составе родов Sororistirps, Botryosella, Sporadopyle, семейства Coeloptychiidae и отряда Hexactinosa, дополняющих филогенетические линии родственных групп, создает предпосылки для разработки биостратиграфической шкалы сантона–маастрихта по гексактинеллидам. Материалы по расселению губок в среднем коньяке (Каменный Брод), в раннем и позднем (Коммунар) сантоне позволяют проследить сукцессию спонгий на протяжении коньякского–раннекампанского времени.
Анализ комплекса фоссилий и литологического состава пород туронского–раннекампанского возраста в разрезах Красный Октябрь, Большевик и Коммунар позволяет предположить, что Вольская структурная зона в туронское–раннекампанское время развивалась обособленно от юго-западной части Ульяновско-Саратовского прогиба. Об этом свидетельствуют сокращенные мощности отложений турона, коньяка и особенно нижнего и верхнего сантона в разрезах Вольской впадины. В структуре верхнемеловых пород юго-западной части Ульяновско-Саратовского прогиба значительные интервалы нижнего сантона и нижнего кампана отсутствуют, в то время как в разрезе Коммунар они литологически и палеонтологически охарактеризованы.
Магнитостратиграфическая характеристика разреза Коммунар согласуется с полученными ранее данными об обратной полярности сантонских отложений в сводном разрезе Озерки–Липовка (Guzhikova et al., 2019) и прямой полярности туронских–коньякских пород в разрезе Нижняя Банновка (Guzhikova et al., 2019). При этом в разрезе Коммунар отсутствуют аналоги магнитозоны обратной полярности, обнаруженной в туроне разрезов Озерки-1, 2, 3 (Guzhikova et al., 2019), Большой Каменный овраг (Гужикова и др., 2020б), и интервала с аномальными направлениями намагниченности, задокументированного в коньяке разреза Каменный Брод (Pervushov et al., 2019). Био- и магнитостратиграфические материалы по разрезу Коммунар и другим разрезам можно увязать между собой, исходя из клиноформного строения турона–коньяка, свойственного территории Нижнего и Среднего Поволжья (коньяк и особенно начало сантона – это время тектонической активизации, в результате которой формировались структурные элементы Саратовских и Доно-Медведицких дислокаций). Такое предположение подтверждается присутствием интервала с аномальными направлениями намагниченности только в разрезе Каменный Брод, который является наиболее полным разрезом турона–коньяка в правобережном Поволжье (Pervushov et al., 2019).
В Шкале геологического времени (GTS) (Gradstein et al., 2020) туронский, коньякский и сантонский ярусы характеризуются нормальной полярностью, в то время как в разрезе Коммунар магнитозона прямой полярности соответствует только турону–коньяку, а сантонские отложения охвачены магнитозоной обратного знака (рис. 3).
Однако магнитополярная характеристика сантона в GTS не может рассматриваться в качестве эталона. Во-первых, разрезы сантона, в которых фиксируется исключительно прямая полярность, сосредоточены только в одном регионе – в Северном Средиземноморье, преимущественно на Апеннинском полуострове (Coccioni, Premoli Silva, 2015). Их стратификация базируется на микрофаунистических данных, не позволяющих надежно определить положения границ сантонского яруса по аналогии с лимитотипом нижней границы (GSSP) сантона в Северной Испании (Lamolda et al., 2014) и разрезами, претендующими на роль GSSP кампана в Южной Англии (Montgomery et al., 1998), Техасе (Gale et al., 2008) и Крыму (Гужиков и др., 2021а, 2021б). Во-вторых, магнитостратиграфическая интерпретация линейных аномалий, результаты которой также учтены в GTS, неоднозначна по ряду причин, свойственных мезозойской океанской коре (Гужиков и др., 2007). В то же время магнитостратиграфические материалы по разным регионам, например по Поволжью (Guzhikova et al., 2019; Гужикова и др., 2021; Pervushov et al., 2019), Туаркыру (Гужиков и др., 2003), Западной Сибири (Гнибиденко и др., 2014) и Южной Англии (Montgomery et al., 1998), фиксируют либо знакопеременную, либо обратную полярность в сантонском ярусе (рис. 3).
Обратная намагниченность сантонских отложений не препятствует обоснованию положения нижней границы сантона в разрезе Коммунар в пограничном интервале между слоями 6 и 7 (в пределах подзоны LC8b по БФ). Скорее всего, сантонская–кампанская R-зона соответствует низам магнитного хрона C33r, а разное положение основания хрона 33r по отношению к границам сантонского яруса в удаленных разрезах обусловлено проблемами биостратиграфических корреляций, возникающих при сопоставлении разобщенных палеобиогеографических областей (рис. 3). Продолжительность сантонского века невелика, ~2 млн лет (Gradstein et al., 2020), и сопоставима с величиной диахронности других ярусных границ, определяемых в разных регионах по разным палеонтологическим группам (Гужиков, Барабошкин, 2006; Гужиков и др., 2007). С этой точки зрения сантонские отложения в разрезах Коммунар и Липовка являются аналогами сантона на Туаркыре и верхов сантона в Южной Англии, но соответствуют нижнему кампану Апеннин (рис. 3). Если принять предложение ряда исследователей, в том числе авторов GTS (Gradstein et al., 2020), об использовании подошвы хрона 33r в качестве ведущего признака для обоснования подошвы кампана в Международной стратиграфической шкале (МСШ), то те из ныне относимых к сантону отложений Поволжья, Туаркыра и Южной Англии, которые охвачены обратной полярностью, будут соответствовать кампанскому ярусу МСШ.
Проблема палеомагнитной зональности туронского–коньякского интервала палеомагнитной шкалы выходит за рамки обсуждения результатов исследований. Отметим только, что магнитостратиграфические данные по верхнему мелу Поволжья (Guzhikova et al., 2019; Гужикова и др., 2020б; Pervushov et al., 2019), Крыма (Гужикова и др., 2020а), Туаркыра (Гужиков и др., 2003), Западной Сибири (Гнибиденко и др., 2014), Южной Англии (Montgomery et al., 1998) дают основания для предположения об аномальном (недипольном) характере туронского–коньякского поля (Гужикова и др., 2021). Однако для доказательства этой гипотезы необходимо получение новых магнитостратиграфических данных по одновозрастным отложениям других регионов.
ВЫВОДЫ
Представлено описание стратотипа вольской свиты (коньяк Среднего и Нижнего Поволжья), выделенного ранее в карьере Коммунар, который расположен в северной части г. Вольска. Прослежена последовательность всех зон/подзон бентосных фораминифер, установленных в структуре трех подъярусов коньяка.
По результатам палеонтологических, палео- и петромагнитных исследований в разрезе Коммунар охарактеризованы отложения среднего–верхнего турона, коньяка, нижнего и верхнего сантона, нижнего кампана. Выделенные здесь зоны и подзоны по бентосным фораминиферам сопоставлены с биостратиграфическими подразделениями, установленными по иноцерамам, белемнитам, аммонитам, иглокожим, брахиоподам и губкам. Отличительными чертами изученного разреза являются: биостратиграфически непрерывный пограничный интервал верхнего коньяка–нижнего сантона и аномально сокращенный по мощности интервал сантонского яруса.
Вольская структурная зона, рассматриваемая в составе наложенного Ульяновско-Саратовского прогиба, в течение туронского–раннекампанского времени развивалась обособленно. На фоне общего погружения юго-западной и северо-восточной частей Ульяновско-Саратовского прогиба Вольский сегмент этой отрицательной структуры в это же время испытывал инверсионные движения. Особенности подобного развития Вольской структурной зоны проявились в сокращенных значениях мощности и неоднократных хиатусах в рассматриваемом интервале отложений.
По результатам палеомагнитных исследований разреза Коммунар установлено, что сантон Вольско-Хвалынской впадины характеризуется магнитозоной обратной полярности, соответствующей нижней части хрона 33r. Аналогичная магнитозона обратного знака была выявлена ранее в сантонских отложениях Карамышской депрессии (Guzhikova et al., 2019).
Проведено магнитостратиграфическое сопоставление сантона разных регионов, из которого следует, что сантонские отложения Нижнего Поволжья являются возрастными аналогами сантона Туаркыра, верхов сантона Южной Англии и нижнего кампана Апеннинского полуострова.
Результаты межрегионального прослеживания уровней палеонтологически обоснованной границы сантона–кампана и основания хрона 33r фиксируют временной сдвиг подошвы кампана относительно изохронной геомагнитной инверсии, сопоставимый с длительностью сантонского века (~2 млн лет). Это обстоятельство ставит под сомнение целесообразность выделения весьма непродолжительных геохронологических интервалов (веков) в качестве подразделений Международной геохронологической (стратиграфической) шкалы, потому что временной интервал стратона планетарного распространения должен существенно превышать диахронность его границ (Гужиков, Барабошкин, 2006).
Благодарности. Авторы благодарны А.Г. Маникину, В.А. Грищенко и В.А. Фомину (СГУ) за участие в полевом изучении разреза, В.А. Мусатову (НВНИИГГ) за предоставление условий для проведения обработки микрофаунистических проб, А.Ю. Гужикову (СГУ) за консультации при интерпретации полученных результатов. Авторы выражают особую благодарность администрации ООО “Холцим (РУС) в Вольске” и лично маркшейдеру А.А. Грониной, АО “ХайдельбергЦемент Волга”, руководству Вольского краеведческого музея за содействие в организации полевых работ. Авторы благодарят Е.Ю. Барабошкина (МГУ) за сделанные замечания и предложения, способствовавшие существенному улучшению публикации.
Источники финансирования. Полевые работы и магнитостратиграфические исследования выполнены за счет гранта Российского научного фонда (проект № 20-77-00028) “Проверка гипотезы о существовании эпох обратной полярности в туронском, коньякском и сантонском веках (поздний мел)”. Исследование бентосных фораминифер выполнено при финансовой поддержке РФФИ в рамках научного проекта № 20-35-90077/20 “Бентосные фораминиферы, как ключевой фактор детального расчленения и стратиграфической корреляции турон-коньякских отложений Поволжья”.
Список литературы
Барышникова В.И. Стратиграфия верхнемеловых отложений бассейна среднего течения Дона по фауне фораминифер // Научная конференция по стратиграфии мезозоя и палеогена Нижнего Поволжья. Тезисы докладов. Саратов: Изд-во Саратовского ун-та, 1955. С. 62–64.
Барышникова В.И. Стратиграфическое расчленение верхнемеловых отложений бассейна среднего течения р. Дона по распределению фораминифер // Труды Всесоюзного совещания по разработке унифицированной схемы стратиграфии мезозойских отложений Русской платформы. Л.: ГНТИ–НГТЛ, 1956. С. 269–275.
Барышникова В.И. О расчленении турон-коньякских отложений бассейна среднего течения р. Дона // Ученые записки СГУ им. Н.Г. Чернышевского. Саратов: Изд-во Саратовского ун-та, 1959. Т. 65. С. 65–77.
Барышникова В.И., Иванова А.Н., Морозов Н.С., Хабарова Т.Н. Стратиграфия верхнемеловых отложений Саратовского и Сталинградского Поволжья // Материалы Всесоюзного совещания по уточнению унифицированной схемы стратиграфии мезозойских отложений Русской платформы. Труды ВНИГНИ. 1961. Вып. 29. Т. 3. С. 110–119.
Беньямовский В.Н. Палеобиогеографические сценарии позднемелового–раннепалеогенового времени в пределах северной периферии Тетиса // Биосфера–экосистема–биота в прошлом Земли: палеобиогеографические аспекты. Труды ГИН РАН. 2005. Вып. 516. С. 267–308.
Беньямовский В.Н. Схема инфразонального биостратиграфического расчленения верхнего мела Восточно-Европейской провинции по бентосным фораминиферам. Статья 1. Сеноман–коньяк // Стратиграфия. Геол. корреляция. 2008а. Т. 16. № 3. С. 36–46.
Беньямовский В.Н. Схема инфразонального биостратиграфического расчленения верхнего мела Восточно-Европейской провинции по бентосным фораминиферам. Статья 2. Сантон–маастрихт // Стратиграфия. Геол. корреляция. 2008б. Т. 16. № 5. С. 62–74.
Волков Ю.В., Найдин Д.П. Пассатные течения и меридиональное расселение некоторых морских организмов позднего мела // Докл. АН. 1998. Т. 58. № 3. С. 367–370.
Гнибиденко З.Н., Лебедева Н.К., Шурыгин Б.Н. Региональный магнитостратиграфический разрез верхнемеловых отложений юга Западной Сибири (Омская впадина) // Докл. АН. 2014. Т. 458. № 1. С. 83–87.
Гужиков А.Ю., Барабошкин Е.Ю. Оценка диахронности биостратиграфических границ путем магнитохронологической калибровки зональных шкал нижнего мела Тетического и Бореального поясов // Докл. АН. 2006. Т. 409. № 3. С. 365–368.
Гужиков А.Ю., Молостовский Э.А., Назаров Х., Фомин В.А., Барабошкин Е.Ю., Копаевич Л.Ф. Магнитостратиграфические данные по верхнему мелу Туаркыра (Туркменистан) и их значение для общей палеомагнитной шкалы // Физика Земли. 2003. № 9. С. 31–44.
Гужиков А.Ю., Барабошкин Е.Ю., Фомин В.А. Магнитостратиграфическая шкала меловой системы: современное состояние, проблемы построения и перспективы развития // Меловая система России и ближнего зарубежья: проблемы стратиграфии и палеогеографии. Ред. Первушов Е.М. Саратов: Изд-во Саратовского ун-та, 2007. С. 69–86.
Гужиков А.Ю., Барабошкин Е.Ю., Александрова Г.Н., Рябов И.П., Устинова М.А., Копаевич Л.Ф., Миранцев Г.В., Кузнецов А.Б., Фокин П.А., Косоруков В.Л. Био-, хемо- и магнитостратиграфия пограничного интервала сантона–кампана разрезов Кудрино и Аксу-Дере (Юго-Западный Крым): проблемы глобальной корреляции и выбора лимитотипа нижней границы кампанского яруса. Статья 1. Геологическое описание, седиментология, биостратиграфия // Стратиграфия. Геол. корреляция. 2021а. Т. 29. № 4. С. 71–117.
Гужиков А.Ю., Барабошкин Е.Ю., Александрова Г.Н., Рябов И.П., Устинова М.А., Копаевич Л.Ф., Миранцев Г.В., Кузнецов А.Б., Фокин П.А., Косоруков В.Л. Био-, хемо- и магнитостратиграфия пограничного интервала сантона–кампана разрезов Кудрино и Аксу-Дере (Юго-Западный Крым): проблемы глобальной корреляции и выбора лимитотипа нижней границы кампанского яруса. Статья 2. Магнито- и хемостратиграфия, обсуждение данных // Стратиграфия. Геол. корреляция. 2021б. Т. 29. № 5. С. 27–58.
Гужикова А.А., Рябов И.П., Копаевич Л.Ф. Новые палеомагнитные и микрофаунистические данные по турону–сантону разреза “Аксу-Дере” (ЮЗ Крым) // Меловая система России и ближнего зарубежья: проблемы стратиграфии и палеогеографии. Материалы X Всероссийского совещания, Магадан, 20–25 сентября 2020 г. Ред. Барабошкин Е.Ю., Гужиков А.Ю. Магадан: ОАО “МАОБТИ”, 2020а. С. 81–84.
Гужикова А.А., Первушов Е.М., Рябов И.П., Фомин В.А. Магнитозона обратной полярности в туроне–коньяке северного окончания Доно-Медведицких дислокаций // Изв. Сарат. ун-та. Нов. сер. Сер. Науки о Земле. 2020б. Т. 20. Вып. 4. С. 262–277.
Гужикова А.А., Рябов И.П., Грищенко В.А., Фомин В.А., Гужиков А.Ю., Первушов Е.М. Магнитостратиграфия турона–сантона Нижнего и Среднего Поволжья // Палеонтология, стратиграфия и палеогеография мезозоя и кайнозоя бореальных районов. Материалы научн. онлайн-сессии, 19–22 апреля 2021 г. [электронный ресурс]. Ред. Лебедева Н.К., Горячева А.А., Дзюба О.С., Шурыгин Б.Н. Новосибирск: ИНГГ СО РАН, 2021. С. 54–58.
Калякин Е.А. Позднемеловые морские ежи Центральнорусской палеобиогеографической провинции // Проблемы палеоэкологии и исторической геоэкологии. Сборник трудов Всеросс. научн. конф., посвященной памяти проф. В.Г. Очева. Саратов: Изд-во СГТУ, 2017. С. 90–96.
Калякин Е.А. Морские ежи Micraster Центральнорусской палеобиогеографической провинции // Труды Всеросс. палеонтол. общ-ва. Том II. М.: ПИН РАН, 2019. С. 69–85.
Калякин Е.А., Миранцев Г.В., Первушов Е.М., Рябов И.П. Туронский–коньякский комплекс иглокожих из разрезов “Чухонастовка” и “Каменный Брод” (южная часть Ульяновско-Саратовского прогиба) // Меловая система России и ближнего зарубежья: проблемы стратиграфии и палеогеографии. Материалы IX Всероссийского совещания. Ред. Барабошкин Е.Ю., Липницкая Т.А., Гужиков А.Ю. Белгород: Политерра, 2018. С. 151–154.
Морозов Н.С. Верхнемеловые отложения междуречья Дона и Северного Донца и южной части Волго-Донского водораздела. Саратов: Изд-во Сарат. ун-та, 1962.
Морозов Н.С., Бондарева М.В. Верхнемеловые отложения междуречья Медведицы и Волги в пределах Саратовской области // Вопросы геологии Южного Урала и Поволжья. 1970. Вып. 7. Ч. 1. С. 116–141.
Морозов Н.С., Орехова В.М. Меловая·система. Верхний отдел // Геология СССР. Т. 46. Ростовская, Волгоградская, Астраханская области и Калмыцкая АССР. Геологическое описание. М.: Недра, 1969. С. 318–361.
Москвин М.М., Эндельман Л.Г. Позднемеловые морские ежи Мангышлака и их стратиграфическое значение // Биостратиграфия мезозойских отложений нефтегазоносных областей СССР. М.: ИГиРГИ, 1972. С. 3–10.
Найдин Д.П. Верхнемеловые белемниты Русской платформы и сопредельных областей. М.: Изд-во Моск. ун-та, 1964.
Найдин Д.П. Подкласс Endocochlia – внутрираковинные // Атлас верхнемеловой фауны Донбасса. М.: Недра, 1974. С. 197–240.
Найдин Д.П. Эвстазия и эпиконтинентальные моря Восточно-Европейской платформы. Ст. 2. Верхнемеловые секвенции платформы // Бюлл. МОИП. Отд. геол. 1995. Т. 70. Вып. 5. С. 49–65.
Найдин Д.П. Разрез туронских и коньякских отложений на р. Тузлов (Ростовская область) // Труды НИИ геологии СГУ. Нов. сер. 2004. Т. XVI. С. 172–178.
Найдин Д.П., Похиалайнен В.П., Кац Ю.И., Красилов В.А. Меловой период. М.: Наука, 1986.
Невесская Л.А. Этапы развития бентоса фанерозойских морей. Мезозой. Кайнозой. М.: Наука, 1999. 503 с. (Труды ПИН РАН. Т. 274.)
Олферьев А.Г., Алексеев А.С. Стратиграфическая схема верхнемеловых отложений Восточно-Европейской платформы. Объяснительная записка. М.: ПИН РАН, 2005.
Олферьев А.Г., Беньямовский В.Н., Вишневская В.С., Иванов А.В., Копаевич Л.Ф., Первушов Е.М., Сельцер В.Б., Тесакова Е.М., Харитонов В.М., Щербинина Е.А. Верхнемеловые отложения северо-запада Саратовской области. Статья 2. Проблемы хроностратиграфической корреляции и геологической истории региона // Стратиграфия. Геол. корреляция. 2008. Т. 16. № 3. С. 47–74.
Олферьев А.Г., Беньямовский В.Н., Иванов А.В., Овечкина М.Н., Сельцер В.Б., Харитонов В.М. Верхнемеловые отложения севера Саратовской области. Статья 1. Разрез карьера “Большевик” в окрестностях Вольска // Бюлл. МОИП. Отд. геол. 2009а. Т. 84. Вып. 2. С. 5–22.
Олферьев А.Г., Беньямовскuй В.Н., Иванов А.В., Овечкина М.Н., Сельцер В.Б., Харитонов В.М. Верхнемеловые отложения севера Саратовской области. Ст. 2. Биостратиграфическое расчленение разреза карьера “Большевик” в окрестностях Вольска // Бюлл. МОИП. Отд. геол. 2009б. Т. 84. Вып. 4. С. 29–46.
Олферьев А.Г., Сельцер В.Б., Алексеев А.С., Амон Э.О., Беньямовский В.Н., Иванов А.В., Овечкина М.Н., Харитонов В.М. Верхнемеловые отложения севера Саратовской области. Статья 3. Биостратиграфическое расчленение разреза карьера “Красный Октябрь” на южной окраине г. Вольска // Бюлл. МОИП. Отд. гeол. 2014. Т. 89. Вып. 6. С. 45–76.
Первушов Е.М., Худяков Д.В. Позднемеловые известковые губки юго-востока Восточно-Европейской платформы // Изв. Сарат. ун-та. Нов. сер. Сер. Науки о Земле. 2020. Т. 20. Вып. 3. С. 184–191.
Первушов Е.М., Сельцер В.Б., Калякин Е.А., Фомин В.А., Рябов И.П., Ильинский Е.И., Гужикова А.А., Бирюков А.В., Суринский А.М. Комплексное био- и магнитостратиграфическое изучение разрезов “Озерки” (верхний мел, Саратовское правобережье). Статья 2. Характеристика ориктокомплексов и биостратиграфия // Изв. Сарат. ун-та. Нов. сер. Сер. Науки о Земле. 2017. Т. 17. Вып. 3. С. 182–199.
Первушов Е.М., Сельцер В.Б., Калякин Е.А., Ильинский Е.И., Рябов И.П. Туронские–коньякские отложения юго-западной части Ульяновско-Саратовского прогиба // Изв. вузов. Геология и разведка. 2019. № 5. С. 10–27. https://doi.org/10.32454/0016-7762-2019-5-10-27
Первушов Е.М., Сельцер В.Б., Калякин Е.А., Рябов И.П. Сантон Вольско-Хвалынской структурной зоны. Статья 1. История исследования и строение разрезов // Изв. Сарат. ун-та. Нов. сер. Сер. Науки о Земле. 2020а. Т. 20. Вып. 3. С. 192–203. https://doi.org/10.18500/1819-7663-2020-20-3-192-203
Первушов Е.М., Сельцер В.Б., Калякин Е.А., Рябов И.П. Сантон Вольско-Хвалынской структурной зоны. Статья 2. Биостратиграфия и современное структурное положение // Изв. Сарат. ун-та. Нов. сер. Сер. Науки о Земле. 2020б. Т. 20. Вып. 3. С. 204–221. https://doi.org/10.18500/1819-7663-2020-20-3-204-221
Первушов Е.М., Рябов И.П., Сельцер В.Б., Валащик И., Калякин Е.А., Гужикова А.А., Ильинский Е.И., Худяков Д.В. Верхнемеловые отложения Вольской структурной зоны Восточно-Европейской платформы: турон–нижний кампан разреза Коммунар. Статья 1. Описание разреза, бентосные фораминиферы, магнитостратиграфия // Стратиграфия. Геол. корреляция (в печати).
Сельцер В.Б. Верхнетуронские аммониты из центральной части Саратовского правобережья // Меловая система России и ближнего зарубежья: проблемы стратиграфии и палеогеографии. Материалы IX Всеросс. совещания. Ред. Барабошкин Е.Ю., Липницкая Т.А., Гужиков А.Ю. Белгород: Политерра, 2018а. С. 249–252.
Сельцер В.Б. О находках коньякских (верхний мел) аммонитов на территории Воронежской и Саратовской областей // Современные проблемы изучения головоногих моллюсков. Морфология, систематика, экология, эволюция, биостратиграфия. Материалы совещания. Ред. Леонова Т.Б., Барсков И.С., Митта В.В. М.: ПИН РАН, 2018б. С. 90–93.
Сельцер В.Б. Гетероморфные аммониты Hyphantoceras и Eubostrychoceras из туронских отложений Саратовского Поволжья // Биогеография и эволюционные процессы. Материалы LXVI сессии Палеонтол. общ-ва РАН. СПб.: ВСЕГЕИ, 2020. С. 151–153.
Сельцер В.Б., Иванов А.В. Атлас позднемеловых аммонитов Саратовского Поволжья. М.: Университет, 2010. 152 с.
Сельцер В.Б., Первушов Е.М., Калякин Е.А. О сантонских отложениях вольских меловых карьеров // Недра Поволжья и Прикаспия. 2020. Вып. 102. С. 61–81.
Стратиграфическая схема верхнемеловых отложений Восточно-Европейской платформы. СПб.: Картографическая фабрика ВСЕГЕИ, 2004.
Christensen W.K. Upper Cretaceous belemnites from the Vomb Trough in Scania Sweden // Sveriges Geologiska Undersokning. Ser. Ca. 1986. № 57. P. 3–39.
Christensen W.K. Belemnites from the Coniacian to Lower Campanian chalks of Norfolk and Southern England // Palaeontology. 1991. V. 34. Pt. 3. P. 695–749.
Christensen W.K. The Late Cretaceous belemnite family Belemnitellidae: taxonomy and evolutionary history // Bull. Geol. Soc. Denmark. 1997. V. 44. P. 59–88.
Christensen W.K., Schulz M.-G. Coniacian and Santonian belemnite faunas from Bornholm, Denmark // Fossils and Strata. 1997. № 44. P. 1–73.
Coccioni R., Premoli Silva I. Revised Upper Albian–Maastrichtian planktonic foraminiferal biostratigraphy and magnetostratigraphy of the classical Tethyan Gubbio section (Italy) // Newslett. Stratigr. 2015. V. 48. P. 47–90.
Gale A.S., Hancock J.M., Kennedy J.W., Petrizzo M.R., Lees J., Walaszczyk I., Wray D. An integrated study (geochemistry, stable oxygen and carbon isotopes, nannofossils, planktonic foraminifera, inoceramid bivalves, ammonites and crinoids) of the Waxahachie Dam Spillway section, north Texas: a possible boundary stratotype for the base of the Campanian Stage // Cretaceous Res. 2008. V. 29. P. 131–167.
Gaspard D. Distribution and recognition of phases in the Aptian–Turonian (Cretaceous) brachiopod development in NW Europe // Geol. Carpatica. 1997. V. 48. P. 145–161.
Gradstein F.M., Ogg J.G., Schmitz M.B., Ogg G.M. Geologic Time Scale 2020. Elsevier, 2020.
Guzhikova A.A., Guzhikov A.Yu., Pervushov E.M., Ryabov I.P., Surinskiy A.M. Existence of the reversal polarity zones in Turonian–Coniacian from the Lower Volga (Russia): new data // Recent Advances in Rock Magnetism, Environmental Magnetism and Paleomagnetism. Eds. Nurgaliev D., Shcherbakov V., Kosterov A., Spassov S. Springer Geophysics. Springer, Cham, 2019. P. 353–369.
Haq B.U. Cretaceous eustasy revisited // Global Planet. Change. 2014. V. 113. P. 44–58.
Houša V. Lewesiceras Spath (Pachydiscidae, Ammonoidea) from Turonian of Bohemia // Sbornik Geol. Ved. Paleontologie. 1967. V. 9. P. 7–49.
Kaplan U., Kennedy W.J. Ammoniten des westfalischen Coniac // Geol. und Palaontol. in Westfalen. 1994. H. 31. P. 155.
Kaplan U., Schmid F. Die heteromorphen Ammoniten der Gattungen Eubostrychoceras und Hyphantoceras aus dem Turon NW-Deutschland // Geol. und Palaontol. in Westfalen. 1988. H. 12. P. 47–87.
Kennedy W.J., Gale A.S. Late Turonian ammonites Haute-Normandie France // Acta Geol. Polon. 2015. V. 65. № 4. P. 507–524.
Kennedy W.J., Kaplan U. Ammoniten aus dem Turonium des Munsterlander Kreidebeckens // Geol. und Palaontol. in Westfalen. 2019. V. 92. P. 3–223.
Košťák M. Cenomanian through the lowermost Coniacian Belemnitellidae Pavlov (Belemnitida, Coleoidea) of the East European Province // Geolines. 2004. V. 18. P. 59–103.
Lamolda M.A., Paul C.R.C., Peryt D., Pons J.M. The Global Boundary Stratotype and Section Point (GSSP) for the base of the Santonian Stage, “Cantera de Margas”, Olazagutia, northern Spain // Episodes. 2014. V. 37. № 1. P. 2–13.
Montgomery P., Hailwood E.A., Gale A.S., Burnett J.A. The magnetostratigraphy of Coniacian–Late Campanian chalk sequences in southern England // Earth Planet. Sci. Lett. 1998. V. 156. P. 209–224.
Neumann C., Jagt J.W.M., van der Ham R.W.J.M. Rare Campanian echinoids from Höver and Misburg (Hannover Area, Lower Saxony, Germany) // Mitteilungen aus dem Museum für Naturkunde in Berlin. Geowissenschaftliche Reihe. 2002. № 5. S. 121–139.
Olszewska-Nejbert D. Late Cretaceous (Turonian–Coniacian) irregular echinoids of western Kazakhstan (Mangyshlak) and southern Poland (Opole) // Acta Geol. Polon. 2007. V. 57. № 1. P. 1–87.
Pervushov E.M., Ryabov I.P., Guzhikov A.Yu., Vishnevskaya V.S., Kopaevich L.F., Guzhikova A.A., Kalyakin E.A., Fomin V.A., Sel’tser V.B., Il’inskii E.I., Mirantsev G.V., Proshina P.A. Turonian–Coniacian deposits of the Kamennyi Brod-1 Section (Southern Ulyanovsk-Saratov Trough) // Stratigr. Geol. Correl. 2019. V. 27. № 7. P. 804–839. https://doi.org/10.1134/S0869593819070025
Sahagian D., Pinous O., Olferiev A., Zakharov V. Eustatic curve for the Middle Jurassic–Cretaceous based on Russian Platform and Siberian stratigraphy: zonal resolution // AAPG Bull. 1996. V. 80. № 9. P. 1433–1458.
Seitz O. Die Inoceramen des Santon und Unter-Campan von Nordwestdeutschland. II. Teil (Biometrie, Dimorphismus und Stratigraphie der Untergattung Sphenoceramus (J. Böhm) // Beihefte zum Geologischen Jahrbuch. 1965. V. 69. P. 1–194.
Sklenář J., Simon E. Brachiopod Gyrosoria Cooper, 1973 – a comparative palaeoecological, stratigraphical and taxonomical study // Bull. Geosci. Czech Geol. Surv. 2009. V. 84. № 3. P. 437–464.
Smith A.B., Wright C.W. British Cretaceous echinoids. Part 9. Atelostomata 2, Spatangoida (2) // Monogr. Palaeontograph. Soc. London. 2012. V. 166(639). P. 635–754.
Surlyk F. Morphological adaptations and population structures of the Danish Chalk brachiopods (Maastrichtian, Upper Cretaceous) // Det Kongelige Danske Videnskabernes Selskab, Biologiske Skrifter. 1972. V. 19. № 2. P. 1–57.
Surlyk F. The Maastrichtian Stage in NW Europe, and its brachiopod zonation // Bull. Geol. Soc. Denmark. 1984. V. 33. P. 217–223.
Thibault N., Jarvis I., Voigt S., Gale A.S., Attree K., Jenkyns H.C. Astronomical calibration and global correlation of the Santonian (Cretaceous) based on the marine carbon isotope record // Paleoceanography. 2016. V. 31. P. 847–865.
Tröger K.-A. Problems of Upper Cretaceous inoceramid biostratigraphy and paleobiogeography in Europe and western Asia // Cretaceous of the Western Tethys. Proc. 3rd Int. Cretaceous Symp. Ed. Wiedmann J. Tubingen, 1987. P. 911–930.
Vishnevskaya V.S., Kopayevich L.F., Benyamovsky V.N., Ovechkina M.N. Correlation of the Upper Cretaceous zonal schemes of the Eastern European Platform based on foraminifera, radiolarian, and nannoplankton // Moscow Univ. Geol. Bull. 2018. V. 73(2). P. 131–140.
Walaszczyk I. Turonian through Santonian deposits of the Central Polish Uplands; their facies development, inoceramid paleontology and stratigraphy // Acta Geol. Polon. 1992. V. 42. P. 1–122.
Walaszczyk I. Inoceramid bivalves at the Turonian/Coniacian boundary: biostratigraphy, events, and diversity trend // Acta Geol. Polon. 2000. V. 50. P. 421–430.
Walaszczyk I., Cobban W.A. Inoceramid faunas and biostratigraphy of the Upper Turonian–Lower Coniacian of the Western Interior of the United States // Spec. Pap. Palaeontol. 2000. V. 64. P. 1–118.
Walaszczyk I., Wood C.J. Inoceramids and biostratigraphy at the Turonian/Coniacian boundary; based on the Salzgitter-Salder Quarry, Lower Saxony, Germany, and the Slupia Nadbrzezna section, Central Poland // Acta Geol. Polon. 1998. V. 48. P. 395–434.
Walaszczyk I., Wood C.J. Inoceramid bivalves from the Coniacian (Upper Cretaceous) of the Staffhorst shaft (Lower Saxony, Germany) – Stratigraphical significance of a unique succession // Cretaceous Res. 2018. V. 87. P. 226–240.
Walaszczyk I., Wood C.J., Lees J.A., Danuta P., Voigt S., Wiese F. Salzgitter Salder Quarry (Lower Saxony, Germany) and Slupia Nadbrzeïna river cliff section (central Poland): a proposed candidate composite Global Boundary Stratotype Section and Point for the Coniacian Stage (Upper Cretaceous) // Acta Geol. Polon. 2010. V. 60. P. 445–477.
Walaszczyk I., Kopaevich L.F., Beniamovski V.N. Inoceramid and foraminiferal record and bio-zonation of the Turonian and Coniacian (Upper Cretaceous) of the Mangyshlak Mts., western Kazakhstan // Acta Geol. Polon. 2013. V. 63. № 4. P. 469–487.
Walaszczyk I., Dubicka Z., Olszewska-Nejbert D., Remin Z. Integrated biostratigraphy of the Santonian through Maastrichtian (Upper Cretaceous) of extra-Carpathian Poland // Acta Geol. Polon. 2016. V. 66. № 3. P. 313–350.
Walaszczyk I., Plint A.G., Landman N.H. Inoceramid bivalves from the Coniacian and basal Santonian (Upper Cretaceous) of the Western Canada Foreland basin // Bull. Am. Mus. Nat. Hist. 2017. V. 414. P. 53–103.
Wiese F., Čech S., Ekrt B., Košťák M., Mazuch M., Voigt S. The Upper Turonian of the Bohemian Cretaceous Basin (Czech Republic) exemplified by the Úpohlavy working quarry: integrated stratigraphy and palaeoceanography of a gateway to the Tethys // Cretaceous Res. 2004. V. 25. P. 329–352.
Wood C.J., Ernst G., Rasemann G. The Turonian–Coniacian stage boundary in Lower Saxony (Germany) and adjacent areas: the Saltzgitter-Salder Quarry as a proposed international standard section // Bull. Geol. Soc. Denmark. 1984. V. 33. P. 225–238.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция