

Успехи современной биологии, 2019, T. 139, № 1, стр. 84-102
Бореальные леса северо-западной пацифики в условиях глобальных изменений климата. Ландшафтно-экологический прогноз
Э. Г. Коломыц *
Институт экологии Волжского бассейна
Тольятти, Россия
* E-mail: egk2000@mail.ru
Поступила в редакцию 27.04.2018
После доработки 25.08.2018
Принята к публикации 25.08.2018
Аннотация
Представлен количественный анализ реакции лесных экосистем в различных секторах Тихоокеанского мегаэкотона Северной Евразии на прогнозируемые изменения климата по глобальным прогнозным моделям E GISS и HadCM3 на предстоящие 50–200 лет. Проведено сравнение полученных экологических сценариев с аналогичными прогнозами по внутриконтинентальным регионам Восточной Европы. Рассмотрены два прогнозных климатических тренда – холодно-аридный и термоаридный. Менделеевскому островодужному экорегиону (о-в Кунашир) свойственна исключительно высокая скорость функционального поглощения ареалов почти всех базовых растительных сообществ экстразональными объектами, что выходит на региональные масштабы климатогенной трансформации биогеоценологической структуры островодужных гео(эко)систем. В окраинно-материковом же низкогорном Нижнем Приамурье на фоне аналогичных, но гораздо более слабых, фитоценологических трансформаций широкое развитие получают буферные лесные сообщества.
ВВЕДЕНИЕ
Современный этап развития континентальной биосферы наиболее ярко выражен в пределах Тихоокеанского кольца – наиболее активного на планете подвижного тектонического пояса, который включает систему вулканических островных дуг и прилегающие материковые окраины с проявлениями активной сейсмичности и вулканизма (Мещеряков, 1981; Герасимов, 1986; Rice, 1978). Частью этого подвижного пояса является Тихоокеанский ороклиматический мегаэкотон Северной Евразии, охватывающий восточную тектонически активную окраину материка и островную сушу в пределах умеренного и субарктического биоклиматических поясов (Коломыц, 2017). Начиная с середины мезозоя, Тихоокеанский подвижной пояс оказывается одним из истоков формирования континентальной биосферы. Установлено, что окраинно-материковые и островные территории его внетропической части послужили генераторами и хранителями континентальных и океанических элементов биоты на фоне пульсационного режима абиотической среды. Здесь сосредоточены условия для развития очаговых биоценотических процессов и для формирования таксонов растений и животных высокого систематического ранга, для появления в природно-территориальных структурах устойчивых новообразований (Юрцев, 1974; Крестов, 2004; Панфилов, 2005; Hulten, 1933; Hopkins, 1972; Nakamura, Krestov, 2005).
Важным направлением ландшафтно-экологического анализа является разработка проблемы хроноорганизации, по терминологии (Солнцев, 1981), природных экосистем Тихоокеанского мегаэкотона, которая связана с внешней (в данном случае климатической) регуляцией их функциональных состояний. В данном сообщении рассмотрены климатогенные механизмы эволюционных преобразований лесных экосистем с помощью эмпирической имитации этой динамики по ландшафтно-экологическим прогнозам. Основная цель состояла в том, чтобы установить, как локальные гео(эко)системы, принадлежащие различным структурным подразделениям мегаэкотона, реагируют на вековые крупномасштабные изменения климата – ведущего экзогенного ландшафтообразующего фактора, то есть каковы закономерности взаимодействия природно-территориальных структур разной возрастной генерации с климатической системой. Научный поиск, учитывающий свойство пространственно-временного самоподобия операционной системы (Пузаченко, Скулкин, 1981), был направлен на определение основных параметров структурно-функциональной организации и устойчивости экотонных систем локального уровня в меняющемся климате.
ОБЪЕКТЫ
Прогнозный экологический анализ проведен на примере двух регионов: 1) островодужного – территории и окрестностей действующего влк. Менделеева на о-ве Кунашир Южно-Курильской гряды; 2) окраинно-материкового – Комсомольского заповедника в Нижнем Приамурье. По определению (Сочава, 1980), первый регион входит в Неопацифический сектор Тихоокеанского кольца, а второй – в сектор Субпацифики. В каждом из этих регионов на экспериментальных полигонах была проведена крупномасштабная ландшафтно-экологическая съемка с фиксацией более 40 параметров состояния лесных биогеоценозов (БГЦ) по 35–50 пробным площадям.
Южные Курилы, и в особенности о-в Кунашир, представляют собой образец высокого разнообразия
типов растительности умеренного пояса. По ботанико-географическому районированию (Атлас…,
2009), о-в Кунашир входит в Южно-Курильско-Хоккайдский округ и в Южно-Курильский район
с господством темнохвойных и хвойно-широколиственных лесов, обогащенных южными восточноазиатскими
элементами, с присутствием вечнозеленых субтропических и реликтовых видов. Вулкану
Менделеева свойственно также весьма четкое и достаточно полное проявление высотно-зональной
биоклиматической поясности, несмотря на его относительно небольшую высоту. В районе
влк. Менделеева сосредоточены весьма разнообразные во флористическом и ботанико-географическом
отношениях группировки растительности (Манько, Розенберг, 1970). Нами выделены здесь
шесть групп типов леса (в скобках даны условные обозначения и таксономические нормы
абсолютных высот): 1) смешанные темнохвойно-широколиственные леса  , 184 м; 2) темнохвойные (елово-пихтовые) леса
, 184 м; 2) темнохвойные (елово-пихтовые) леса 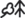 , 379 м; 3) буферные каменноберезово-еловые леса
, 379 м; 3) буферные каменноберезово-еловые леса  , 575 м; 4) каменноберезовое криволесье с кедровым стлаником
, 575 м; 4) каменноберезовое криволесье с кедровым стлаником  , 685 м; 5) заросли кедрового стланика
, 685 м; 5) заросли кедрового стланика  , 718 м; 6) буферные кедровостланиково-березово-еловые сообщества
, 718 м; 6) буферные кедровостланиково-березово-еловые сообщества  , 575 м. Эти типы относятся к соответствующим флористическим фратриям: маньчжурско-охотской,
охотской, берингийско-камчатской, берингийской и берингийско-камчатско-охотской.
, 575 м. Эти типы относятся к соответствующим флористическим фратриям: маньчжурско-охотской,
охотской, берингийско-камчатской, берингийской и берингийско-камчатско-охотской.
На территории Менделеевского полигона растительные сообщества распределены по сочетанию двух ороклиматических факторов: 1) вертикальной ландшафтной микроярусности, включающей в себя определенную ступень абсолютных высот и преобладающий тип местоположения, и 2) макроэкспозиции склонов влк. Менделеева и окружающих его равнин. Выделены четыре типа местоположений, по систематике (Глазовская, 1964): аккумулятивный А, трансаккумулятивный ТА, транзитный Т и трансэлювиальный ТЭ. В соответствии с региональными климатическими контрастами, обусловленными циркуляционной экспозицией макросклонов вулкана, проведено разделение первых трех ландшафтных ярусов на две части – охотоморскую (Ох, экспозиционный сектор ЮЗ–З–СЗ–С) и тихоокеанскую (Т/о, сектор СВ–В–ЮВ–Ю). Перечислим выделенные ороклиматические комплексы с указанием интервалов абс. высот: А – равнинно-предгорные Ох-1 (60–250 м) и Т/о-1 (60–180 м); ТА – низкогорные нижнеярусные Ох-2 (250–600 м) и Т/о-2 (180–520 м); Т – низкогорные среднеярусные Ох-3 (600–720 м) и Т/о-3 (520–670 м); ТЭ – низкогорные верхнеярусные Ох-4 и Т/о-4 (более 670–720 м).
Второй исследуемый регион входит в дальневосточную ветвь бореального биоклиматического
экотона Евразии, по определению (Базилевич и др., 1986), и охватывает подтаежные низкогорно-долинные
комплексы Нижнего Приамурья, которые выделяются как буферные зональные экосистемы
(Сочава, 1979). На территории Комсомольского заповедника сохранился один из крупных
массивов темнохвойных и хвойно-широколиственных лесов. Согласно Б.П. Колесникову (Колесников,
1956), для Нижнеамурской Субпацифики были выделены четыре базовые флористические фратрии:
маньчжурская мезофильная, маньчжурская ксерофильная, охотская и ангаридская. Процесс
миграции и региональной дифференциации третичной праманьчжурско-северояпонской формации
(Сочава, 1946), приведший к обособлению вышеупомянутых флор, сопровождался также их
интеграцией – определенным смешением в благоприятных для этого местообитаниях, с формированием
устойчивых буферных (переходных) фратрий. Нами выделены две буферные фратрии – маньчжурско-охотская
(МО) и маньчжурско-ангаридская (МА). Выделены также следующие группы типов леса: 1)
широколиственная  ; 2) кедрово-широколиственная дубовая
; 2) кедрово-широколиственная дубовая  ; 3) кедрово-широколиственные липовая
; 3) кедрово-широколиственные липовая  ; 4) елово-широколиственная
; 4) елово-широколиственная  ; 5) пихтово-еловая
; 5) пихтово-еловая  ; 6) лиственничная
; 6) лиственничная  . Отдельно выделялась лесо-болотная группа, представленная лиственничными марями.
. Отдельно выделялась лесо-болотная группа, представленная лиственничными марями.
В Комсомольском заповеднике распределение указанных фитоценологических единиц имеет свою геоморфологическую предопределенность, которая включает в себя: а) тип местоположения, б) солярную экспозицию склона (для транзитных типов местоположений) и в) степень проявления литоморфности–гидроморфности эдафтопов. Сочетания этих признаков формируют сложную низкогорно-долинную систему мезомасштабных ландшафтных сопряжений (мезокатен). Выделены следующие гидрогеоморфологические звенья катенарной системы: 1) горное элювиально-трансэлювиальное сублитоморфное (450–680 м); 2) горное транзитное и трансаккумулятивное теневых склонов лито-гидроморфное (310–680 м); 3) горное транзитное солнцепечных и нейтральных склонов литоморфное (190–450 м); 4) предгорное трансаккумулятивное солнцепечное и нейтральное сублитоморфное (110–310 м); 5) долинное элювиальное сублитоморфное (40–110 м); 6) долинное аккумулятивное субгидроморфное (20–40 м); 7) супераквальное гидроморфное (20–40 м).
МЕТОДЫ
В экологическом прогнозировании ключевое значение имеют оценки состояний экосистем и их наиболее вероятных динамических тенденций через призму конкурентных отношений их экологических ниш. В основе этого подхода лежат известные представления об экологической нише объекта в виде некоторой области распределения его состояний в данном экологическом пространстве (Odum, 1983). Вероятностный ландшафтно-экологический прогноз осуществлялся с помощью операций с современными и предполагаемыми гидротермическими нишами экосистем (Коломыц, 2005, 2008). Для этого каждая такая ниша была представлена некоторым размытым (дескриптивным) множеством в виде вектора-строки (или вектора-столбца). Составляющими вектора служили нормированные частные коэффициенты связи Cij градаций ai – i-го явления А и bj – j-го фактора В (Пузаченко, Скулкин, 1981), которые рассматривались как весовые коэффициенты. Каждый из этих коэффициентов является определенным эквивалентом вероятности (встречаемости) того или иного объекта при данной градации геофизического параметра. Вероятность эта тем выше, чем больше частный коэффициент связи (табл. 1).
Таблица 1.
Нормированные частные коэффициенты связей Cij растительных сообществ экспериментальных полигонов с климатическими характеристиками базового периода, а также их значения как весовых коэффициентов (даны в скобках)
| Фитоценологические единицы | tVIII, °С | rгод, мм | Кувл |
|---|---|---|---|
| Менделеевский островной вулканический ландшафт (о-в Кунашир) | |||
| Флористические фратрии | 0.405 (0.318) | 0.543 (0.426) | 0.327 (0.256) |
| Группы типов леса | 0.308 (0.321) | 0.381 (0.398) | 0.296 (0.281) |
| Комсомольский заповедник (Нижнее Приамурье) | |||
| Фитоценологические единицы | tVII, °С | rгод, мм | Кувл |
| Флористические фратрии | 0.088 (0.317) | 0.106 (0.381) | 0.084 (0.302) |
| Группы типов леса | 0.118 (0.358) | 0.115 (0.350) | 0.096 (0.292) |
Ландшафтно-экологический прогноз состоит в оценках вероятностей и скоростей взаимных переходов в системе данных объектов (флористических фратрий, групп типов леса или групп микроландшафтов) согласно установленным трендам изменений того или иного набора климатических параметров. Вероятности ландшафтно-экологических переходов представлены в матрицах и на графо-аналитических моделях. Под переходом нами понимается такая смена функционального, а затем и структурного состояния данного объекта, при которой объект принимает (с той или иной долей вероятности) черты других объектов-прообразов благодаря тому, что его экологическая (в данном случае частная климатическая) ниша при своем изменении все более пересекается с нишами этих объектов.
Сама процедура прогнозирования уже излагалась нами достаточно подробно (Коломыц, 2008; Залиханов и др., 2010). Схематически ее можно представить следующим образом. По каждой паре объектов А (редуцента) и В (абсорбента) рассчитывались две прогнозные характеристики: Pii ≡ К(А0 → А1) – переход объекта-редуцента А самого в себя (вероятность его устойчивости) и Pij ≡ К(А0 → В0) – вероятность поглощения его другим объектом-абсорбентом В (здесь “≡” – знак тождества). Эти характеристики выражались следующими мерами включения (∩):
С помощью последних находились: общая вероятность Pi преобразований объекта-редуцента, суммарные Mi и парциальные mij скорости его переходов во все j-е объекты-абсорбенты, а также время τ (Pii) полной трансформации редуцента, выражаемое в числах шагов или в годах. Расчетные формулы указанных характеристик имеют вид, согласно (Харбух, Бонэм-Картер, 1974; Harbaugh, Bonham-Carter, 1970):
Здесь d – число шагов. В данном случае имеем три шага: 1990–2050 гг. и 2050–2100 гг. и 2100–2200 гг. Как известно (Изменение …, 2001), начало современного глобального потепления приходится на середину–конец 1980-х гг.
Параметры Pii и Pij отображают две альтернативные категории устойчивости гео(эко)систем: ареальную (геотопическую) и миграционную (трансгрессивную), которые характеризуют соответственно состояние устойчивого равновесия системы и ее вынужденное поведение как реакции на внешнее воздействие.
По матрицам отношений включения того или иного объекта другими объектами данного ряда (флористических фратрий, групп типов леса, либо микроландшафтов) рассчитывался транзитный индекс k-го объекта Sk. Для каждого объекта определялись число входных связей (ni) и число связей выходных (nj). Подсчитывалось также N – общее число связей в системе. Исходя из этого, транзитный индекс Sk объекта определялся как:
Отметим также две главные особенности проведенных прогнозных расчетов. Во-первых, в качестве предикторов использованы три климатических показателя: средняя температура самого теплого месяца (tVII–VIII), годовое количество осадков (rгод) и коэффициент увлажнения Высоцкого–Иванова (Кувл). Во-вторых, учитывается вероятность “остаточных” переходов того или иного объекта в другие прообразы-объекты, отсутствующие в данной операционной системе (выборке), то есть в экстразональные, по определению (Спрыгин, 1986), или вневыборочные, объекты. Эти объекты могут относиться к другим зонально-региональным условиям, нередко весьма отдаленным от условий данной территории.
Для территории Нижнего Приамурья по данным метеостанций получены следующие линейные связи температур и осадков с абсолютной высотой (Hабс, в рамках низкогорно-долинного рельефа), с высокими коэффициентами корреляции R:
С другой стороны, существуют вполне определенные закономерности базового высотно-поясного распределения фитоценологических единиц по территории Комсомольского заповедника (см. далее рис. 5 и 6). Тем самым предопределена достаточно высокая чувствительность этих единиц к изменениям фоновой биоклиматической системы, что должно существенно повысить репрезентативность прогнозных ландшафтно-экологических моделей.
Третий показатель является функцией первых двух и, главным образом, среднеиюльской температуры. Так, для территории Нижнего Приамурья по данным метеостанций получена следующая линейная связь:
Тем не менее, коэффициент атмосферного увлажнения играет самостоятельную системообразующую роль, что подчеркивается значениями его весовых коэффициентов (табл. 2).
Таблица 2.
Сопоставление прогнозируемых изменений климатических параметров во внутри- и окраинно-материковых регионах Северной Евразии, а также в островодужной Неопацифике
| Регион | Экспериментальный полигон (прогнозный срок, год) |
Отклонения климатических параметров от их базовых значений | ||
|---|---|---|---|---|
| ΔtVII–VIII, °С | Δrгод, мм | ΔКувл | ||
| Прогнозная климатическая модель E GISS | ||||
| Среднее Поволжье | Заповедник Керженский (2050) | –0.8 | –67 | –0.13 |
| Приокско-Террасный заповедник (2050) | –1.9 | –67 | –0.02 | |
| Центральный Кавказ, Приэльбрусье | Урочище Жиласу, альпийский пояс (2100) | +1.1 | –24 | –0.21 |
| Нижнее Приамурье | Заповедник Комсомольский (2100) | –0.2 | –53 | +0.16 |
| Южный Кунашир | Влк. Менделеева (2100) | –0.6 | –160 | –0.33 |
| Прогнозная климатическая модель HadCM3, версия А2 | ||||
| Среднее Поволжье | Нижегородское Предволжье (2100) | +7.1 | –100 | –0.83 |
| Центральный Кунашир | Влк. Менделеева (2050) | –4.9 | –314 | +1.2 |
Излагаемому ниже ландшафтно-экологическому прогнозу необходимо предпослать следующее важное замечание. Предполагаемые функционально-структурные сдвиги гео(эко)систем, которые могут быть обусловлены изменениями климата (в данном случае его антропогенными изменениями) с периодом колебаний 50 лет и более, характеризуют не столько сами будущие природные комплексы, сколько ландшафтно-экологические условия и, соответственно, тот предел экологического равновесия, к которому будут стремиться реальные биогеосистемы в своих изменениях. Рассчитанное в количестве шагов τ (ш) или в числах лет τ (г) время достижения этого равновесия (см. далее табл. 6 и 7) относится только к функциональной релаксации, но не к полному периоду трансформации структуры. Последнее будет определяться не только интенсивностью внешнего воздействия, но и характерными временами различных природных признаков (Арманд, Таргульян, 1974). Для лесных ландшафтов, как известно, шаг прогнозирования, не превышающий продолжительности жизни одного поколения древостоев, позволяет определить потенциальные лесорастительные условия.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Прогнозируемые по глобальным климатическим трендам сценарии фитоценологических преобразований на Тихоокеанском мегаэкотоне Северной Евразии описывают весьма специфичную и достаточно сложную систему ландшафтно-экологических переходов двух иерархических уровней – регионального и локального. Для того что бы ее оценить, коснемся сначала известных положений геоэкологии об иерархических закономерностях динамики экосистем с привлечением наших эмпирических материалов по другим экорегионам Северной Евразии. В табл. 2 и 4 приведены основные характеристики климатических прогнозов на рассматриваемые экорегионы по двум глобальным моделям – умеренной E GISS (Hansen et al., 2007) и экстремальной HadCM3, версия А2 (Pope et al., 2000).
1. Внутриконтинентальные регионы Восточной Европы
На примере Центрального Кавказа (Залиханов и др., 2010) было установлено, что региональные внутриматериковые ландшафты испытывают в целом гораздо более слабые переходы друг в друга по сравнению с их внутренними видоизменениями, которые являются результатом взаимных трансформаций составляющих их гео(эко)систем топологической размерности. Как известно (Theurillat et al., 1998), региональные эффекты глобального потепления в горах прослеживаются главным образом по общему смещению границ растительных зон, а эффекты локальные – по динамике внутризональной структуры фитобиоты. Существенные различия в скорости реакции региональных и локальных экосистем на одни и те же изменения климата обусловлены их неодинаковыми временами релаксации: “… чем выше рангом подразделение природной среды, тем больше его возраст” (Сочава, 1978, с. 196) и, продолжим, тем дольше оно сохраняет прежнюю систему ландшафтных связей при одном и том же внешнем воздействии, то есть тем медленнее оно изменяется. Если рассматривать устойчивость гео(эко)системы “… как отношение между изменчивостью в среде и изменчивостью в самой системе” (Ricklefs, 1976, с. 378), то очевидно, что региональные природные комплексы должны быть более устойчивы, нежели локальные. Их внутренняя организация меньше зависит от состояния внешней среды, что снижает и скорость их реакции на те же внешние воздействия. Таковы общие черты иерархической системы климатогенной динамики внутриконтинентальных гео(эко)систем, прошедших значительную историю субаэрального развития.
Покажем эту фундаментальную геоэкологическую закономерность на примере разномасштабных природных комплексов Большого Кавказа, сравнив климатогенную динамику региональных ландшафтов и слагающих их геосистем топологического уровня. В целом региональные ландшафтные системы данной горной страны обладают сравнительно высокой устойчивостью к рассматриваемым климатическим изменениям (доминирующие значения Pii ≥ 55–75% (табл. 3)), в то время как относительные скорости их взаимных переходов весьма низки (Pij(ср) ≤ 15–30%). Очевидно, время релаксации кавказских региональных ландшафтов существенно больше полупериода данных климатических колебаний. Согласно модели E GISS, даже к концу прогнозного периода (2100 г.) внутренняя перестройка высокогорных ландшафтов окажется еще недостаточной для того, чтобы они сколько-нибудь существенно потеряли свой облик и начали целиком трансформироваться в верхне- и среднегорные ландшафты.
Таблица 3.
Обобщенные характеристики вероятностей и скоростей функционально-структурных переходов для ландшафтных групп северного макросклона Большого Кавказа на прогнозный срок 2100 г., согласно термоаридному климатическому тренду по модели E GISS (Залиханов и др., 2010)
| Высотные ярусы | Статистические параметры | ||||
|---|---|---|---|---|---|
| Pii | Pi | Pij(ср) | τ (ш) | τ (г) | |
| Предгорный | 0.33–0.92 | 0.08–0.45 | 0.13–0.33 | 3.0–25.3 | 327–2830 |
| Нижне- и среднегорный | 0.39–0.75 | 0.25–0.61 | 0.11–0.14 | 3.3–8.0 | 359–882 |
| Верхнегорный | 0.40–0.46 | 0.54–0.60 | 0.12–0.13 | 3.3–3.7 | 364–404 |
| Высокогорный | 0.55–0.91 | 0.09–0.45 | 0.07–0.14 | 4.4–22.7 | 488–2498 |
Таблица 4.
Изменения основных климатических параметров Менделеевского экорегиона (ст. Южно-Курильск), прогнозируемые по глобальным моделям E GISS и HadCM3
| Сроки | Климатические параметры | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| температура воздуха, °С | cумма активных температур (∑t ≥ 10), °C |
годовая испаряемость (Eo), мм | атмосферные осадки, мм * | Кувл | |||||
| tгод | tII | tVIII | rгод | rхп | rтп | ||||
| Базовый период | 4.8 | –6.0 | 15.8 | 1519 | 304 | 1319 | 410 | 909 | 4.33 |
| Прогнозная климатическая модель E GISS | |||||||||
| 2100 г. | 4.3 | –6.6 | 15.2 | 1459 | 292 | 1159 | 278 | 887 | 3.97 |
| 2200 г. | 4.1 | –6.8 | 14.7 | 1411 | 282 | 1069 | 231 | 852 | 3.79 |
| Прогнозная климатическая модель HadCM3, версия А2 | |||||||||
| 2050 г. | 3.4 | –3.2 | 10.9 | 1048 | 209 | 1005 | 458 | 432 | 4.81 |
| 2100 г. | 6.3 | –0.5 | 16.8 | 1615 | 323 | 1066 | 511 | 448 | 3.30 |
Совершенно иная картина свойственна высокогорным экосистемам локального уровня, ярким примером чего служит полигон Жиласу в Северном Приэльбрусье (рис. 1). Даже при том относительно слабом климатическом сигнале, который дает модель E GISS, мера стабилизации Pii большинства высокогорно-луговых БГЦ снижается до 35–50%, а вероятности их взаимных переходов оказываются в несколько раз выше, чем в целом у региональных высокогорных ландшафтов (Pij(ср) достигает 20–25% и более). Время релаксации топоэкосистем гораздо более конформно полупериоду рассматриваемых климатических изменений, поэтому их чувствительность к гидротермическим сигналам должна быть сравнительно высокой. Так, пустошные луга (Kobresia–lichen, lichen–Alchemilla–Kobresia) и альпийские ковры (Kobresia–Alchemilla–Sibbaldia) субнивального пояса (группа БГЦ 1) будут трансформироваться не только в соседние альпийские луга (группу 2), но и в относительно удаленные травяные сообщества субальп. Эти же альпийские ковры к середине XXI в. будут на 20–28% замещаться более ксерофитными сообществами, аналогичными разнотравно-злаковым и манжетковым субальпийским лугам (группами 3 и 4, Alchemilla, Kobresia–Alchemilla). И только бурьянистые разнотравно-чемерице-крапивные сообщества и осоковые луга субальп (группы БГЦ 5 и 6, Veratrum–Urtica, Trifolium–Veratrum, Veratrum–Alchemilla) могут переходить на 45–65% и 12–28% соответственно в свои экстразональные верхне- и среднегорные аналоги.
Рис. 1.
Северное Приэльбрусье. Полигон Жиласу. Термоаридный климатический тренд. Орграфы структурно-функциональных переходов на срок 2100 г., согласно прогнозно-климатической модели Е GISS, между различными ценотическими группами высокогорных лугов, находящимися в различных высотных поясах и в неодинаковых условиях эдафического увлажнения. Транзитные экотипы растительных сообществ: а – абсорбенты, б – субабсорбенты; в – мезотропы; г – субредуценты; д – редуценты. Вероятности стабилизации Pii и взаимных переходов Pij: е – 0.01–0.10; ж – 0.11– 0.20; и – 0.21–0.30; к – 0.31–0.40; л – 0.41–0.50; м – 0.51–0.60; н – 0.61–0.70; о – 0.71–0.80; п – 0.80–0.90.
Приведем далее пример прогнозируемых ландшафтно-экологических последствий холодно-аридного климатического тренда (по модели E GISS) на юге подтаежной зоны Среднего Поволжья, в Приокско-террасном заповеднике (рис. 2). Здесь к 2050 г. на фоне общей достаточно высокой устойчивости почти всех БГЦ (мера стабилизации составляет более 60%) намечается определенная тенденция (до 25%) трансформации ксероморфных трансэлювиальных песчаных зеленомошно-вейниковых сосновых боров (группа БГЦ 1) в соседние элювиальные мезогидроморфные моренные кислично-разнотравные ельники и елово-сосновые леса (группы 5 и 2). Последние же, в свою очередь, начнут преобразовываться с еще большей интенсивностью (около 30%) в гидроморфные элювиально-аккумулятивные хвойно-мелколиственные кустарничково-сфагновые сообщества междуречных понижений с супесчано-суглинистыми почвами (группа 6). В том же направлении изменятся (на 27%) и более дренированные мезоморфные трансэлювиальные и транзитные дубово-липово-березовые разнотравные и широкотравные леса на известняково-суглинистом субстрате (группа 4).
Рис. 2.
Среднее Поволжье. Прогнозно-климатическая модель E GISS на 2050 г. Экорегион Приокско-Террасного заповедника. Холодно-гумидный климатический тренд. Орграфы функциональных переходов между группами фаций (биогеоценозов), имеющих различную зональную принадлежность и находящихся на различных уровнях базового функционирования. Условные обозначения переходов те же, что и на рис. 1. Остальные обозначения см. в тексте (здесь и на рис. 3–5).

Возникнут также явные признаки трансгрессии в данный экорегион вневыборочных южнотаежных сообществ. Последние будут проникать главным образом по речным долинам, замещая более чем на 65% переувлажненные леса. Вторая менее интенсивная волна трансгрессии (до 30%) охватит умеренно дренированные водоразделы и склоны с мезоморфными сосново-липово-дубовыми широкотравными лесами на глееватых супесчаных почвах (группы БГЦ 3).
Таким образом, даже относительно небольшой холодно-аридный климатический сигнал, со сменой годового коэффициента увлажнения с 1.52 до 1.78–1.79, способен вызвать определенный, хотя и достаточно локализованный, процесс распространения лесных и лесо-болотных ассоциаций южной тайги почти на всю территорию подтаежной зоны. Тем не менее, общая ландшафтная структура данного внутриконтинентального экорегиона остается мало измененной. Ее климатогенная динамика ограничивается преимущественно взаимными переходами между ее местными элементами, и сколько-нибудь существенного регионального сдвига не просматривается.
Во внутриконтинентальных регионах лишь исключительно сильные климатические сигналы способны трансформировать локальные ландшафтно-экологические переходы в региональные. Такие трансформации могут быть связаны, например, с известной экстремальной климатической моделью HadCM3, дающей в Среднем Поволжье к 2100 г. повышение средних январских и июльских температур на 5–7°С, при сравнительно небольшом увеличении количества осадков (Коломыц, 2008, рис. 41). По нашим оценкам, в лесостепных районах Жигулей и Присурья произойдет вековое падение Кувл с 0.95–1.07 до 0.36–0.45, а в Заволжско-Керженском подтаежном районе – с 1.37 до 0.65, что будет означать резко выраженный термоаридный климатический тренд. Начнется прогрессирующее летнее иссушение почвы – всеобщее явление, предполагаемое при глобальном потеплении на юге умеренного пояса практически всеми прогнозно-климатическими моделями (Climate change…, 1996). В связи с этим по всей средней полосе Русской равнины ожидается достаточно резкий стартовый скачок преобразований лесных экосистем. Функциональная устойчивость бореальных и неморальных лесов может быть нарушена уже в первые десятилетия XXI в. При экстремальном глобальном потеплении процесс аридизации широколиственных лесов должен прогрессивно ускоряться в следующем экзодинамическом сукцессионном ряду: чистые и еловые дубравы → боровые сосняки и елово-сосновые субори → типичная хвойная и мелколиственная лесостепь → северная и средняя степь → южная (сухая) степь. Остепнение дубрав должно непременно идти через промежуточную стадию бореальных, главным образом боровых, лесных и редколесных ассоциаций.
Зона смешанных лесов Русской равнины отличается от неморальнолесной подзоны более высокой устойчивостью лесных сообществ к глобальному потеплению. Общая климатическая термоаридизация вызовет местные переходы, направленные преимущественно снизу вверх по локальным катенам – от супераквальных и аккумулятивных местоположений к трансэлювиальным и элювиальным. В условиях низменных зандров аридизация хвойнолесных сообществ вызовет их переходы в состояния типичной и южной лесостепи. Остепнению подвергнутся в первую очередь ксероморфные и мезоксероморфные елово-сосновые леса верхних звеньев катен. Согласно модели HadCM3, уже к 2050 г. здесь будут развиваться сообщества северных и средних степей.
2. Островодужный вулканический экорегион
На начальном островодужном геологическом этапе формирования континентальной биосферы реакция молодых вулканических и поствулканических ландшафтов на колебания климатической системы существенно отличается от описанной выше реакции, свойственной внутриконтинентальным равнинным и горным ландшафтам, прошедшим уже значительный и весьма сложный путь субаэрального развития. Эти отличия обусловлены двумя важнейшими чертами биоклиматической системы островодужной Неопацифики на юге бореального пояса. Сюда относятся (Коломыц, Сурова, 2018): 1) одновременное снижение и сжатие (по сравнению с соседним материком) системы высотной поясности, почти полный спектр которой (от лесных поясов до подгольцового пояса) укладывается в пределах низкогорного яруса макрорельефа; 2) охват локальными структурно-функциональными контрастами таксономических норм почвенно-биотических признаков целого ряда вышестоящих зонально-региональных единиц; такие геотопические контрасты на катенах вполне определенно имитируют высотно-поясную и циркуляционно-экспозиционную дифференциацию гидротермических условий. Таким образом, в рамках низкогорного островодужного ландшафта формируется определенный псевдорегиональный спектр высотных поясов, который правомерно рассматривать как очаг зарождения региональной высотной биоклиматической поясности, свойственной горным странам материков.
Описываемую ниже реакцию локальных гео(эко)систем островодужной Неопацифики на колебания региональной климатической системы будем называть неопацифическим типом климатогенной динамики растительного покрова на начальном островодужном этапе формирования континентальной биосферы. Рассмотрим основные черты этого типа.
В первую очередь, в Менделеевском ландшафте отчетливо выделяются две главные особенности прогнозируемых климатогенных фитоценотических трансформаций (рис. 3, 4; табл. 4, 5). 1. Чрезвычайно низкая геотопическая устойчивость практически всех рассматриваемых фитоценологических единиц даже при климатических сигналах слабой силы, когда тренд средних температур не превышает ±0.7–1.1°С, а годовой суммы осадков – ±15–20% (табл. 4); это отображается весьма малыми значениями параметра Pii (в основном не более 15–20%). 2. В противоположность этому, исключительно высокая скорость функционального поглощения ареалов почти всех базовых растительных сообществ экстразональными (вневыборочными) объектами, характеризующими уже региональные масштабы климатогенной трансформации фитоценологической структуры бореального островодужного ландшафта (Pij (вневыб.) ≥ ≥ 0.25–0.40 и нередко достигает 0.70–0.80).
Рис. 3.
Менделеевский вулканический экорегион. Орграфы структурно-функциональных переходов на срок 2100 г., согласно холодно-аридному тренду прогнозной климатической модели E GISS между группами типов леса, расположенными в различных экспозиционных секторах о-ва Кунашир.

Рис. 4.
Менделеевский вулканический экорегион. Орграфы структурно-функциональных переходов на срок 2100 г., согласно термоариденому тренду по прогнозной климатической модели HadCM3 между группами типов леса, расположенными в различных экспозиционных секторах о-ва Кунашир.

Таблица 5.
Обобщенные характеристики вероятностей и скоростей функционально-структурных переходов для растительных сообществ Менделеевского ландшафта на прогнозный срок 2100 г., согласно холодно-аридному климатическому тренду по модели E GISS
| Обозначения | Статистические параметры | Sk | ||||
|---|---|---|---|---|---|---|
| Pii | Pi | Pij (экстра-зональные) | τ (ш) | τ (г) | ||
| Группы типов леса | ||||||
 |
0.121 | 0.879 | 0.255 | 1.14 | 131 | 1.15 СА |
 |
0.376 | 0.624 | 0.439 | 1.60 | 184 | 0.86 СР |
 |
0.163 | 0.837 | 0.287 | 1.19 | 137 | 1.42 А |
 |
0.205 | 0.795 | 0.595 | 1.26 | 144 | 0.71 Р |
 |
0.281 | 0.719 | 0.719 | 1.39 | 160 | 0.84 СР |
 |
0.169 | 0.831 | 0.625 | 1.20 | 138 | 0.84 СР |
| Флористические фратрии | ||||||
| Маньчжурско-Охотская | 0.128 | 0.872 | 0.744 | 1.15 | 132 | 0.88 СР |
| Охотская | 0.288 | 0.717 | 0.457 | 1.39 | 160 | 0.62 Р |
| Берингийско-Камчатская | 0.221 | 0.779 | 0.641 | 1.28 | 148 | 1.13 СА |
| Берингийская | 0.053 | 0.949 | 0.949 | 1.05 | 121 | 1.37 А |
| Берингийско-Камчатско-Охотская | 0.154 | 0.846 | 0.602 | 1.18 | 136 | 1.00 М |
Обозначения в табл. 5–8. Транзитные экотипы: А – абсорбенты; СА – субабсорбенты; М – мезотропы; СР – субредуценты; Р – редуценты.
Таблица 6.
Обобщенные характеристики вероятностей и скоростей функционально-структурных переходов для растительных сообществ Менделеевского ландшафта на прогнозный срок 2100 г., согласно термоаридному климатическому тренду по модели HadCM3
| Обозначения | Статистические параметры | Sk | ||||
|---|---|---|---|---|---|---|
| Pii | Pi | Pij (экстразональные) | τ (ш) | τ (г) | ||
| Группы типов леса | ||||||
 |
0 | 1.000 | 1.000 | 1.00 | 115 | 1.36 А |
 |
0.053 | 0.947 | 0.812 | 1.60 | 121 | 1.09 СА |
 |
0 | 1.000 | 0.848 | 1.19 | 115 | 1.09 СА |
 |
0.090 | 0.910 | 0.609 | 1.10 | 126 | 1.00 |
 |
0 | 1.000 | 0.809 | 1.00 | 115 | 0.82 СР |
 |
0 | 1.000 | 0.519 | 1.00 | 115 | 0.64 Р |
| Флористические фратрии | ||||||
| Маньчжурско-Охотская | 0 | 1.000 | 1.000 | 1.00 | 115 | 1.60 А |
| Охотская | 0.064 | 0.936 | 0.872 | 1.07 | 123 | 1.00 М |
| Берингийско-Камчатская | 0.082 | 0.918 | 0.836 | 1.10 | 125 | 1.00 М |
| Берингийская | 0 | 1.000 | 1.000 | 1.00 | 115 | 1.00 М |
| Берингийско-Камчатско-Охотская | 0 | 1.000 | 0.772 | 1.00 | 115 | 0.40 Р |
Таблица 7.
Обобщенные характеристики вероятностей и скоростей функционально-структурных переходов для растительных сообществ Комсомольского заповедника на прогнозный срок 2050 г., согласно термоаридному климатическому тренду по модели E GISS
| Обозначения | Статистические параметры | Sk | ||||
|---|---|---|---|---|---|---|
| Pii | Pi | Pij (экстра-зональные) | τ (ш) | τ (г) | ||
| Группы типов леса | ||||||
 |
0.574 | 0.426 | 0.321 | 2.35 | 99 | 1.00 М |
 |
0.567 | 0.433 | 0.045 | 2.31 | 88 | 1.07 СА |
 |
0.064 | 0.936 | 0.703 | 1.07 | 41 | 1.00 M |
 |
0.604 | 0.396 | 0.295 | 2.23 | 85 | 1.14 А |
 |
0.184 | 0.816 | 0.151 | 1.23 | 47 | 0.71 Р |
 |
0.554 | 0.456 | 0.259 | 2.19 | 839 | 1.08 СА |
| Флористические фратрии | ||||||
| Маньчжурская мезофильная | 0.282 | 0.718 | 0.483 | 1.39 | 53 | 1.06 СА |
| Маньчжурская ксерофильная | 0.464 | 0.536 | 0.231 | 1.87 | 71 | 0.95 СР |
| Маньчжурско-Охотская | 0.375 | 0.625 | 0.233 | 1.60 | 61 | 0.94 СР |
| Охотская | 0.367 | 0.633 | 0 | 1.58 | 60 | 0.94 СР |
| Маньчжурско-ангаридская | 0.373 | 0.627 | 0.434 | 1.59 | 60 | 0.94 СР |
| Ангаридская | 0.456 | 0.544 | 0.544 | 1.84 | 70 | 1.17 А |
Итак, климатогенные преобразования природно-территориальных структур Менделеевского вулканического ландшафта выходят на региональный уровень. Смещения высотно-зональных границ в пределах лишь одного, низкогорного, геоморфологического яруса приобретают масштабы гораздо более крупных, региональных изменений. Как мы уже видели, во внутриконтинентальных экорегионах при аналогичных климатических трендах эти структурно-функциональные изменения остаются преимущественно на топологическом уровне.
2.1. Региональное похолодание
Модель E GISS дает на весь прогнозный период возрастающий холодно-аридный климатический тренд – понижение температур (как в холодный, так и в теплый периоды года) и сокращение атмосферных осадков. Еще более резко выражен данный тренд по экстремальной модели HadCM3 на ближайший срок – до 2050 г. (табл. 4). Будем рассматривать в основном первую, более умеренную, модель. Если на первом прогнозном шаге (до 2100 г.) зимнее и летнее понижение температуры одинаково (около 0.6°С), то на втором шаге (к 2200 г.) летнее похолодание почти на 0.5°С больше зимнего. На этом фоне явную сезонную диспропорцию обнаружит количество осадков. Атмосферные осадки теплого периода уменьшатся всего на 3–6%, между тем в холодный период их сокращение составит 32% на первом шаге и 25% – на втором. В результате произойдет снижение Кувл. Уменьшение снежности зимы и усиление морозов должны вызвать более глубокое промерзание почвы, что весьма неблагоприятно скажется на произрастании теплолюбивых неморальных растений (особенно субтропических реликтов маньчжурской флоры) и будет способствовать их вытеснению бореальными видами охотской и камчатской флор.
Согласно холодно-аридному тренду, широколиственно-темнохвойный экорегион центрального Кунашира сместится в северо-восточном направлении, в сторону соседнего о-ва Итуруп (ст. Курильск). Здесь прибрежно-равнинный и низкогорный биоклиматический фон создают каменно-березовые леса и криволесья, а также лиственничные леса и редколесья с каменной березой и кедровым стлаником; гольцы среднегорий заняты вересковыми тундрами, верещатниками и субальпийскими лугами (Атлас…, 2009).
Что могут представлять собой эти вневыборочные (экстразональные) фитоценотические объекты? Здесь можно наметить три обобщенные группы (рис. 3).
Группа I. В подгольцовом поясе и у верхней границы горно-лесного пояса кедровый стланик и березовое криволесье будут на 60–70% и более замещены ассоциациями вышележащих высотных поясов, поэтому экстразональными здесь можно считать горные тундры, луга и кустарники, аналоги которых распространены, например, на абс. высотах более 1200–1300 м в привершинной полосе вулканического массива Руруй (северо-восток Кунашира). Этими сообществами являются (Атлас…, 2009): верещатники (Empetrum nigrum s.t., Rododendron aureum, Ledum macrophyllum, Vaccinium uliginosum) в комплексе с разреженными стланиками (Pinus pumila, Duschekia maximoviczii) и фрагментами субальпийских лугов (Solidafo viga-aurea, Geranium erianthum, Pedicularis resupinata, Nilingia ajanensis, Parageum calihifolium), а также и с бамбучниками (Sasa spp.).
Группа II. Пихто-ельники и березово-еловые сообщества горнолесного пояса Менделеевского ландшафта будут, по-видимому, преобразовываться (на 45–55%) в средне- и высокогорные ольховые стланики (Duchekia maximovizhii) в комплексе с кедровыми стланиками, лугами и верещатниками. Их аналоги широко представлены на отметках 800–1200 м в массиве Руруй.
Группа III. Наконец, в предгорные хвойно-широколиственные леса могут трансгрессировать каменно-березняки (Betula ermanii) с кедровым стлаником из верхней полосы лесного пояса Менделеевского вулкана, трансформируясь одновременно в криволесные травяные и стланиковые ассоциации. Аналогами последних могут служить распространенные ныне в высотной полосе 600–900 м влк. Тятя ольховые стланики, высокотравные (Filipendula kamischatica, Solidago spiraeifolia, Aruncus cliacus, Cacalia hastata), папоротниковые (Dryopteris spp.) и вейниковые (Calamagrostis langsdorfii) ассоциации.
На самом же побережье широкое распространение получат, по-видимому, луговые сообщества – осоково-злаково-разнотравные луга с бамбучником и видами сухих местообитаний (Artemisia, Cichorium, Asteraceae, Caryophyllaceae). Палеоаналоги подобных ассоциаций были развиты на приморских равнинах Кунашира при похолодании в первой половине позднего голоцена – 1700–1300 тыс. л.н. (Разжигаева и др., 2011).
Сопоставление структурных схем фитоценотических переходов в 2100 г. и 2200 г. показывает, что по мере углубления холодно-аридного климатического тренда трансгрессия экстразональных объектов (верещатниково-стланиково-тундровых сообществ) из верхнего низкогорного яруса в средний и нижний ярусы будет приобретать все более обширный характер. Их наступление приостановится только в предгорном ярусе Менделеевского ландшафта, где, как уже говорилось, коренные хвойно-широколиственные леса маньчжурско-охотской флоры будут замещены преимущественно (на 40–50%) березово-еловыми лесами и березовыми криволесьями (с кедровым стлаником в подлеске), принадлежащими в значительной мере берингийско-камчатской, охотской и берингийско-камчатско-охотской фратриям.
Судя по значениям параметра Pii ≈ 0.120, хвойно-широколиственные леса буферной маньчжурско-охотской флоры, а также березово-еловые сообщества берингийско-камчатско-охотской фратрии при холодно-аридном климатическом тренде будут обладать минимальной геотопической устойчивостью, переходя в вышележащие по склону группы леса с вероятностью от 11 до 30%, при одновременном поглощении их зарождающимися в верхнем ярусе верещатниками, а также ольховыми и кедровыми стланиками (табл. 5). Гораздо более устойчивой в своих ареалах окажется группа пихтово-еловых лесов охотской флоры (Pii = 0.32–0.38). Однако и эти сообщества на 45–55% захлестнет трансгрессия новых экстразональных гольцово-тундровых сообществ. Данные сообщества на 60–80% и более охватят также ареалы современных подгольцовых кедровых стлаников и березовых криволесий. Тем не менее, последние будут проявлять определенную миграционную устойчивость, распространяясь в свою очередь на территории современных хвойно-широколиственных и березово-еловых лесов, что и придаст им качества местных абсорбентов и мезотропов.
Очевидно также, что региональная трансгрессия описанных выше горно-тундровых, луговых и верещатниково-стланиковых сообществ вызовет на горных склонах охотоморского сектора более резкую трансформацию лесных сообществ, нежели на склонах сектора тихоокеанского, который, будучи изначально более прохладным, окажется менее чувствительным к холодно-аридному сигналу. По этой же причине смешанные леса охотоморских предгорий станут на 12–13% переходить в пихто-ельники и каменноберезняки соответственно нижнего и среднего ярусов тихоокеанского сектора. Последнее будет означать определенное выравнивание известных биоклиматических контрастов между охотоморским и тихоокеанским секторами, с исчезновением в охотоморском секторе многих теплолюбивых форм и целых сообществ.
2.2. Региональное потепление
Ландшафтно-экологический прогноз для глобального потепления охарактеризован по климатической модели HadCM3 на срок 2100 г. Согласно данному сценарию, повышение температуры в рассматриваемом регионе Неопацифики будет сопровождаться сокращением атмосферных осадков (табл. 4), то есть ожидается явный термоаридный климатический тренд. В островодужном вулканическом ландшафте этот тренд должен вызвать свои специфические фитоценологические перестройки, весьма отличные от тех, которые обусловлены описанным выше холодно-аридным трендом. Прежде всего, резко снизится, вплоть до нуля, параметр Pii – показатель геотопической устойчивости локальных гео(эко)систем. Ослабнет также в целом интенсивность внутриландшафтных фитоценологических переходов и несоизмеримо возрастет трансгрессия экстразональных объектов – лесных ассоциаций из более южных широт.
При термоаридном тренде кардинально меняются структурные схемы доминирования флористических фратрий и групп типов леса в их климатогенных взаимоотношениях (рис. 4; табл. 6). Предгорно-равнинные хвойно-широколиственные леса маньчжурско-охотской флоры, в отличие от их положения при холодно-аридном тренде, приобретают максимальную миграционную устойчивость и становятся относительными абсорбентами, поглощая до 12–17% площади горных березово-еловых лесов, каменноберезового криволесья и тернарных буферных сообществ, распространяясь даже в подгольцовый пояс кедрового стланика. Вторая местная трансгрессия намечается от среднеярусных пихто-ельников субабсорбентов, которые двинутся вверх по склонам и захватят 11–12% площади каменноберезового криволесья и кедровых стлаников. Наконец, полностью исчезнут буферные стланиково-березово-еловые ассоциации берингийско-камчатско-охотской флоры у верхней границы горно-лесного пояса.
Однако все отмеченные фитоценологические переходы будут ничтожны по сравнению с феноменом внешней трансгрессии более южных растительных формаций. В течение 115–125 лет следует ожидать полную функционально-структурную трансформацию всего растительного покрова Менделеевского островного ландшафта (табл. 6). Можно представить следующую последовательную картину такой трансформации.
На предгорные равнины начнут распространяться бамбуковые пихтово-широколиственные леса (Abies sachalinensis, Quercus crispula, Acer mono var. monoratum, Sasa kurile-nesis) с соседнего о-ва Хоккайдо, обладающего по сравнению с Кунаширом гораздо более континентальным климатом. Однако эти экстразональные абсорбенты будут мало отличаться от современных хвойно-широколиственных лесов побережий и предгорий Южного Кунашира. Важнее, по-видимому, будет дальнейшее наступление этих ассоциаций вверх по склонам вулкана вплоть до верхнего яруса с поглощением пояса березового криволесья и с выходом местами в подгольцовый пояс кедрового стланика. Палинологические материалы показывают (Разжигаева и др., 2009), что в эпоху потепления в позднем голоцене (около 6100–2550 л.н.) на равнинах Кунашира господствовали темнохвойно-широколиственные леса с широким участием дуба.
В перспективе дальнейшего потепления вполне допустима трансгрессия в нижний и даже средний низкогорные ярусы влк. Менделеева лесных ассоциаций из более южных и еще более континентальных регионов островной Неопацифики и окраинно-материковой Субпацифики. Таковыми могут оказаться, например, дубово-буковые сообщества о-ва Хоккайдо (Fagus crenata, Quercus crispula), смешанные и широколиственные леса, распространенные в настоящее время в Маньчжурии и Северо-Восточном Китае (Pinus koraiensis, Quercus mongolica, Q. dentala, Fagus crenata, Ilex pedunculosas), широколиственные леса о-ва Хонсю. Не исключено также проникновение к подножьям вулкана субтропических хвойных лесов (Chamaecyparis obtusa) с о-ва Хонсю. Наконец, по долинам рек и на древних морских террасах возможно появление ясенево-ильмовых сообществ (Fraxinus mandshurica, Ulmus propinqua) с о-ва Сахалин, а при дальнейшей трансгрессирующей термоаридизации – даже ксерофитных лещинных дубняков (Quercus mongolica), аналогичных современным дубнякам Южного Приморья.
Описанные процессы термоаридного функционально-структурного преобразования растительного покрова должны проявляться в первую очередь и наиболее интенсивно в более континентальном охотоморском секторе Менделеевского экорегиона и в гораздо меньшей степени – в секторе тихоокеанском.
Современные аналоги ожидаемых сообществ при термоаридизации суббореальных лесов в нижнем и даже среднем ярусах влк. Менделеева можно найти в окрестностях соседней кальдеры Головнина на юго-западной оконечности о-ва Кунашир. Здесь расположены экструзивные купола, на периферии которых проявляется активная фумарольная деятельность. Температура термальных источников составляет около 100°С, а воды в оз. Кипящее – от 35–65°С до 90°С с прогреванием обширных окружающих областей, охватывающих и достаточно удаленное оз. Горячее (Горшков, 1967; Козлов, Жариков, 2012). Интенсивный геотермальный прогрев почво-грунтов обусловил появление в этом районе на фоне господствующих пихтово-еловых лесов двух ареалов-рефугиумов экстразональных широколиственнолесных ассоциаций. Меньший ареал расположен по северо-восточной окраине самой кальдеры Головнина, примыкающей к оз. Кипящее, а более крупный – у юго-западного подножья внешней соммы (Атлас…, 2009). Это дубово-каменноберезово-кленовые сообщества (Quercus crispula, Acer mayrii) типа колочных лесов, обогащенные широколиственными породами (Kolopanax septemlobus, Phellodendron sachalinensa, Gerasus sachalinensis, C. maximoviczii, Ulmus laciniata, U. japonica). Экстразональный (геотермальный) генезис этих суббореальных ассоциаций подчеркивается тем, что у побережья оз. Кипящее они лежат гипсометрически выше пояса бореальных пихто-ельников. Благодаря локальной геотермальной энергии эти широколиственные леса имитируют образец лесного покрова, который может распространиться в Менделеевском экорегионе при термоаридном климатическом тренде.
3. Окраинно-материковый низкогорный экорегион
Для данного региона оба рассматриваемых прогнозных экологических сценария по модели E GISS на сроки 2050 г. и 2200 г. описывают аридизацию, однако в первом случае на фоне потепления, а во втором – похолодания Cij. При этом к 2050 г. сокращение осадков предполагается в два с лишним раза большее, нежели к 2200 г., поэтому термоаридный тренд будет выражен гораздо сильнее по сравнению с трендом холодно-аридным. Соответственно, в первом случае более интенсивными должны быть и процессы фитоценологических трансформаций.
3.1. Региональное потепление
При термоаридном тренде (по модели E GISS на срок 2050 г.) в Нижнем Приамурье, по-видимому, будет развиваться биоклиматическая ситуация, аналогичная той, которая имела здесь место во второй климатический оптимум голоцена – в суббореальный период (4.5–3.2 тыс. л.н.), по схеме Блитта-Сернандера, когда среднеиюльская температура была не менее чем на 1°С выше современной, а количество осадков сократилось. Таковой была верхнеголоценовая фаза 5ж кедрово-широколиственных лесов в Амуро-Уссурийском регионе на 44–46° с. ш. (Короткий и др., 1999).
Основные смещения границ между микроландшафтами будут направлены снизу вверх по мезокатенам – от трансаккумулятивного, аккумулятивного, элювиального и даже супераквального звеньев к транзитному и горному трансэлювиальному звеньям. Намечается следующая наиболее четко выраженная цепочка ландшафтно-экологических переходов: горное трансэлювиальное → горное транзитное солнцепечных склонов → долинное элювиальное, с вероятностью 17–22% и с усилением процесса последовательного поглощения вниз по мезокатене.
На этом общем фоне достаточно разнообразно будут представлены внутрирегиональные фитоценотические перестройки (рис. 5; табл. 7). Распространенные сегодня в среднем и верхнем ярусах (в интервале высот 350–650 м) бореальные пихто-ельники охотской флоры окажутся в состоянии редуцентов, с минимальной устойчивостью (Sk = 0.77). Они будут замещаться мезо-ксерофильными дубо-липняками (с вероятностью около 0.21) и в меньшей степени – горными кедровниками и лиственничниками (соответственно на 18 и 14%). Эти два местных субабсорбента (Sk = 1.04–1.08) проявят определенную устойчивость обоих типов – как геотопическую (для широколиственной группы Pii = 0.34; для лиственничной группы Pii = 0.56), так и миграционную. Однако внутренние функционально-структурные преобразования бореальных лесов данного экорегиона будут в значительной мере подавлены внешней фитоценологической экспансией. Наступление экстразональных объектов проявится наиболее эффективно в нижних Ад- и СА-звеньях ландшафтных сопряжений. Резкой трансформации, с вероятностью 55–65%, подвергнутся мезогигрофильные кедрово-липовые леса на нижних надпойменных террасах р. Амур, а также в долинах р. Горин и ее притоков. Повышенный дренаж этих территорий будет способствовать внедрению сюда с юга полидоминантных широколиственных лесов с пихтой цельнолистной Abies holophylla и с возможным участием умеренно термофильных пород: дуба зубчатого Quercus dendata, бархата амурского Phellodendron amurensis, ореха маньчжурского Juglans manshurica, граба сердцелистного Carpinus cordata. Последние два вида на равнинах Нижнего Приамурья в настоящее время не встречаются (Ахметьева, 1977). Лиственничные сфагновые мари в западинах могут полностью исчезнуть, заменяясь гигромезофитными лесо-луговыми сообществами. Подобные широколиственные леса в суббореальный период господствовали в Южном Приморье (Родникова и др., 2012).
Трансгрессия полидоминантных широколиственных лесов на склоны низкогорий Нижнего Приамурья окажется ограниченной по высоте и весьма выборочной. Эти внешние абсорбенты могут достичь абс. отметок не более 200–300 м и внедрятся максимально (на 29–33%) в ареалы маньчжурско-охотских елово-широколиственных лесов и ангаридских лиственничников. Гораздо слабее (на 16–17%) их трансгрессия коснется лесов маньчжурской флоры. Вышележащего пояса пихто-ельников (300–680 м) данная трансгрессия не достигнет вовсе.
Следует отметить, что при термоаридном климатическом тренде обогащение буферных маньчжурско-охотских лесов широколиственными породами происходит за счет главным образом их внешней трансгрессии (описанной выше), а не внутренней. Это подтверждает известный тезис (Криштофович, 1946; Васильев, 1946) о том, что эволюционный климакс, представленный в данном случае буферными елово-широколиственными лесами (Коломыц, 2017), формируется за счет региональных миграционных процессов в растительном покрове.
3.2. Региональное похолодание
Палеогеографическим аналогом прогнозируемого холодно-аридного биоклиматического тренда (по модели E GISS на срок 2200 г.) может служить фаза 5з позднеголоценового похолодания на рубеже суббореал–субатлантика, в период 2200–1500 л.н. (Короткий и др., 1999). На равнинах Амуро-Уссурийского региона это была фаза хвойно-широколиственных лесов с широким распространением лиственничников и липняков (Родникова и др., 2012). Летние температуры были примерно на 1°С ниже современных, а очередная морская регрессия снижала атмосферное увлажнение. Это в целом соответствует климатическому сценарию по модели E GISS на срок 2200 г.
Матрицы и орграфы флористических и фитоценотических транформаций при холодно-аридном тренде (рис. 6, табл. 8) демонстрируют господство взаимных (прямых и обратных) переходов в бинарных нетранзитивных отношениях объектов (с близкими значениями их, в общем, невысокой вероятности – от 0.03–0.06 до 0.15–0.18). Множественность таких переходов свидетельствует о почти повсеместных процессах смешения флор и усложнения лесных фитоценозов, что и служит одним из механизмов формирования буферных экосистем в окраинно-материковой Субпацифике. Смешению флор при холодно-аридном тренде способствует более низкая, чем при термоаридизации, общая картина фитоценотических трансформаций, более спокойная климатогенная динамика растительных сообществ с доминированием Pij ≤ 0.10.
Рис. 5.
Комсомольский заповедник. Орграфы функционально-структурных переходов между группами типов леса на срок 2050 г., согласно термоаридному тренду по прогнозной климатической модели E GISS. Лесообразующие породы: 1 – дуб; 2 – липа; 3 – кедр; 4 – ель, пихта; 5 – лиственница.

Рис. 6.
Комсомольский заповедник. Орграфы функционально-структурных переходов между группами типов леса на срок 2200 г., согласно холодно-аридному тренду по прогнозной климатической модели E GISS. Условные обозначения те же, что и на рис. 1 и 5.

Таблица 8.
Обобщенные характеристики вероятностей и скоростей функционально-структурных переходов для растительных сообществ Комсомольского заповедника на прогнозный срок 2200 г., согласно холодно-аридному климатическому тренду по модели E GISS
| Обозначения | Статистические параметры | Sk | ||||
|---|---|---|---|---|---|---|
| Pii | Pi | Pij (экстра-зональные) | τ (ш) | τ (г) | ||
| Группы типов леса | ||||||
 |
0.594 | 0.406 | 0.375 | 4.93 | 927 | 1.17 A |
 |
0.594 | 0.406 | 0 | 4.93 | 927 | 1.00 M |
 |
0.405 | 0.595 | 0.118 | 3.36 | 632 | 1.00 M |
 |
0.497 | 0.503 | 0.389 | 3.98 | 748 | 0.91 СР |
 |
0.137 | 0.863 | 0.512 | 2.31 | 434 | 0.92 СР |
 |
0.747 | 0.253 | 0.156 | 7.94 | 149 | 1.00 М |
| Флористические фратрии | ||||||
| Маньчжурская мезофильная | 0.514 | 0.486 | 0 | 4.12 | 774 | 0.95 СР |
| Маньчжурская ксерофильная | 0.497 | 0.503 | 0.171 | 3.97 | 746 | 1.00 М |
| Маньчжурско-Охотская | 0.329 | 0.671 | 0.420 | 2.98 | 560 | 1.05 СА |
| Охотская | 0.302 | 0.698 | 0.439 | 2.87 | 539 | 1.05 СА |
| Маньчжурско-ангаридская | 0.584 | 0.416 | 0.039 | 4.81 | 904 | 0.91 СР |
| Ангаридская | 0.601 | 0.399 | 0.039 | 5.00 | 940 | 1.04 СА |
В низкогорном Нижнем Приамурье холодно-аридный тренд должен вызвать два основных потока внешней фитоценологической трансгрессии. Первый поток будет представлять собой 35–40%-ное поглощение верхнеярусных ТЭ пихто-ельников охотской флоры возникающими и расширяющимися ареалами подгольцовых кедровых стлаников флоры берингийской. Очагом зарождения второго потока станет зимнее выхолаживание речных долин с развитием долинных лиственничников, лиственничных сфагновых марей и торфяных болот с длительно-сезонной мерзлотой. Трансгрессия долинных лиственничников в дубово-липовые и кедрово-елово-широколиственные леса соседних плакоров (с вероятностью до 25%) будет означать, что зимнее инверсионное выхолаживание охватит и элювиальные местоположения, которые перейдут в разряд субредуцентов.
Одновременно горные лиственничники укрепят свои позиции, проявляя высокую гомеостатическую устойчивость (Pii = 0.774), и начнут понемногу захватывать соседние ареалы пихто-ельников, кедровников и дубо-липняков (Pij = 0.03–0.08). Этот процесс распространится преимущественно в горно-транзитных теневых местоположениях, и здесь трансформирующиеся леса будут отличаться также высокой устойчивостью (Pii = 0.632). Наряду с этим достаточно отчетливо (с вероятностью 14–15%) будет выражена тенденция замещения мезофитных неморальных пихто-ельников и гигромезофитных елово-широколиственных лесов (субредуцентов) мезоксерофитными и ксерофитными липово-дубовыми лесами (субабсорбентами) с внедрением в них лиственницы.
В целом можно ожидать общую аридизацию лесного покрова данного региона с усилением всего комплекса ангаридской флоры, переходящей в разряд субабсорбентов (Sk = 1.05) с мерой стабилизации 0.54. Таким образом, холодно-аридный тренд в Нижнем Приамурье должен усиливать один из процессов вековых смен лесообразующих древостоев (Колесников, 1956), а именно длительно устойчивую смену маньчжурской бореально-неморальной флоры флорой маньчжурско-ангаридской неморально-бореальной.
Полученные нами прогнозные ландшафтно-экологические модели по Нижнему Приамурью раскрывают конкретные механизмы отмеченного Б.П. Колесниковым (Колесников, 1956) формирования “многоформенного и динамичного” лесного покрова данного окраинно-материкового экорегиона в условиях периодически сменяемых в течение плейстоцена и голоцена волн потепления и похолодания (Короткий и др., 1999), которые обеспечили “сборность флоры” (Урусов, 1988) как “…результат ее насыщенности до предела климатических возможностей”, по образному выражению А.А. Криштофовича (Колесников, 1969, с. 218).
ЗАКЛЮЧЕНИЕ
Выполнено описание климатогенных механизмов эволюционных ландшафтно-экологических процессов в различных секторах Тихоокеанского мегаэкотона Северной Евразии с помощью имитации этих процессов по ландшафтно-экологическим прогнозам. Проведено сравнение полученных прогнозных сценариев с аналогичными прогнозами по внутриконтинентальным регионам Восточной Европы, что позволило ярче оттенить особенности глобальных климатогенных изменений природной среды в Тихоокеанском подвижном поясе.
Островодужным вулканогенным образованиям локального и субрегионального уровней в Северо-Западной Неопацифике свойственны: 1) чрезвычайно низкая геотопическая устойчивость флористических, фитоценотических и ландшафтных единиц; 2) исключительно высокая скорость функционального поглощения ареалов почти всех базовых растительных сообществ экстразональными объектами, что характеризует уже региональные масштабы климатогенной трансформации биогеоценологической структуры островодужного ландшафта. В Менделеевском бореальнолесном экорегионе при холодно-аридном тренде ожидается массовая трансгрессия кедровых и ольховых стлаников, вересковых тундр, верещатников и субальпийских лугов, а при термоаридизации – смешанных и широколиственных лесов, произрастающих сегодня на о-ве Хоккайдо и в Северо-Восточном Китае.
Реакция лесов различных формаций в Нижнеамурской окраинно-материковой Субпацифике на тот или иной климатический сигнал проявляется через призму сложных межвидовых отношений и многообразия лесообразовательных процессов, которые слагаются из возрастных, вековых и восстановительных смен лесных сообществ. Проведенному ландшафтно-экологическому прогнозу наиболее адекватны возрастные смены древостоев, проходящие, согласно Б.П. Колесникову (Колесников, 1956), по незамкнутой спиралевидной схеме продолжительностью не менее 2–2.5-векового периода и вызванные, как мы полагаем, суперпозицией (взаимным наложением) внешних климатических сигналов и автохтонной смены эдафических условий местопроизрастания.
При термоаридном климатическом тренде прогнозируемое обогащение буферных маньчжурско-охотских лесов широколиственными породами происходит за счет главным образом их внешней (экстразональной) трансгрессии, а не внутренней. Это подтверждает известный тезис (Криштофович, 1946; Васильев, 1946) о том, что эволюционный климакс, представленный в данном случае буферными елово-широколиственными лесами, формируется за счет региональных миграционных процессов в растительном покрове. Холодно-аридному же тренду будет отвечать множественность слабых фитоценотических переходов, что говорит о повсеместных процессах смешения флор и усложнения лесных сообществ. По-видимому, именно благодаря периодическим похолоданиям леса Нижнего Приамурья сформировались и продолжают формироваться “… как исключительно многоформенные и непостоянные по внешнему облику” (Колесников, 1956, с. 136). При термоаридном тренде подобные процессы выражены гораздо слабее. Таким образом, холодно-аридный экологический сценарий детализирует гипотетическую картину формирования лесного покрова бореальной Субпацифики, которое шло “…путем прибоя волн с севера и северо-востока, с отложением продуктов… флористического привноса” (Криштофович, 1946, с. 64).
Список литературы
Арманд Д.Л., Таргульян В.О. Некоторые принципиальные ограничения эксперимента и моделирования в географии // Изв. АН СССР. Сер. геогр. 1974. № 4. С. 129–138.
Атлас Курильских островов / РАН. ИГ РАН. ТИГ ДВО РАН / Ред. В.М. Котляков, П.Я. Бакланов, Н.Н. Комедчиков М., Владивосток: ДИК, 2009. 516 с.
Ахметьева Н.П. Палеогеография Нижнего Приамурья. М.: Наука, 1977. 11 с.
Базилевич Н.И., Гребенщиков О.С., Тишков А.А. Географические закономерности структуры и функционирования экосистем. М.: Наука, 1986. 297 с.
Васильев В.Н. Закономерности процесса смен растительности // Мат. по истории флоры и растительности СССР. М., Л.: АН СССР, 1946. Вып. II. С. 365–403.
Герасимов И.П. Проблемы глобальной геоморфологии. М.: Наука, 1986. 207 с.
Глазовская М.А. Геохимические основы типологии и методики исследований природных ландшафтов. М.: МГУ, 1964. 230 с.
Горшков Г.С. Вулканизм Курильской островной дуги. М.: Наука, 1967. 288 с.
Залиханов М.Ч., Коломыц Э.Г., Сурова Н.А. и др. Высокогорная геоэкология в моделях. М.: Наука, 2010. 487 с.
Изменение климата, 2001 г. Обобщенный доклад МГЭИК / Ред. Р.Т. Уотсон Geneva: World Meteorological Organization, 2003. 220 p.
Козлов Д.Н., Жариков Р.В. Тепловизионная съемка активных вулканов Курильских островов в 2009–2011 гг. // Вестн. КРАУНЦ. Науки о Земле. 2012. № 1. Вып. 19. С. 1–7.
Колесников Б.П. Кедровые леса Дальнего Востока // Тр. ДВФ АН СССР. Т. 2 (4). М., Л.: АН СССР, 1956. 263 с.
Колесников Б.П. Растительность // Южная часть Дальнего Востока / Ред. И.П. Герасимов М.: Наука, 1969. С. 206–250.
Коломыц Э.Г. Бореальный экотон и географическая зональность. Атлас-монография. М.: Наука, 2005. 390 с.
Коломыц Э.Г. Локальные механизмы глобальных изменений природных экосистем. М.: Наука, 2008. 427 с.
Коломыц Э.Г. Тихоокеанский мегаэкотон Северной Евразии. Эволюционная модель континентальной биосферы. М.: ГЕОС, 2017. 496 с.
Коломыц Э.Г., Сурова Н.А. Островные вулканические экосистемы в Тихоокеанском подвижном поясе как истоки формирования континентальной биосферы // Успехи соврем. биол. 2018. Т. 138. № 1. С. 44–69.
Короткий А.М., Волков В.Г., Гребенникова Т.А. и др. Дальний Восток // Изменение климата и ландшафтов за последние 65 млн лет (кайнозой: от палеоцена до голоцена). Глава 7 / Ред. А.А. Величко М.: ГЕОС, 1999. С. 146–164.
Крестов П.В. Растительный покров Командорских островов // Бот. журн. 2004. Т. 89. № 11. С. 1740–1762.
Криштофович А.Н. Эволюция растительного покрова в геологическом прошлом и ее основные факторы // Мат. по истории флоры и растительности СССР. М., Л.: АН СССР, 1946. Вып. II. С. 21–87.
Манько Ю.И., Розенберг В.А. Высотная поясность растительности на вулкане Менделеева // Биологические ресурсы острова Сахалин и Курильских островов. Владивосток: БПИ ДВО АН СССР, 1970. С. 65–71.
Мещеряков Ю.А. Избранные труды. Рельеф и современная геодинамика. М.: Наука, 1981. 277 с.
Панфилов В.Д. Центры эволюции и исторические миграции биоты Земного шара. Научный дневник. М.: ИГ РАН, 2005. 448 с.
Пузаченко Ю.Г., Скулкин В.С. Структура растительности лесной зоны СССР: системный анализ. М.: Наука, 1981. 275 с.
Разжигаева Н.Г., Ганзей Л.А., Гребенникова Т.А. и др. Эволюция и рубежи перестройки ландшафтов Курильских островов в голоцене // География и геоэкология на современном этапе взаимодействия природы и общества. СПб: СПбГУ, фак. геогр. геоэкол., 2009. С. 727–733.
Разжигаева Н.Г., Ганзей Л.А., Мохова Л.М., Пшеничникова Н.Ф. Луговые ландшафты Южных Курил: происхождение, возраст и развитие // Геогр. природ. ресурсы. 2011. № 3. С. 96–104.
Родникова И.М., Лящевская М.С., Киселева А.Г., Пшеничникова Н.Ф. Состояние и динамика почвенно-растительного покрова малых островов залива Петра Великого (Японское море) // Геогр. природ. ресурсы. 2012. № 1. С. 96–103.
Солнцев В.Н. Хроноорганизация географических явлений // Вопросы географии. Сб. 117. Геофизика ландшафта. М.: Мысль, 1981. С. 40–68.
Сочава В.Б. Вопросы флорогенеза и фитоценологии маньчжурского смешанного леса // Мат. по истории флоры и растительности СССР. Вып. II. М., Л.: АН СССР, 1946. С. 283–302.
Сочава В.Б. Введение в учение о геосистемах. Новосибирск: Наука, 1978. 319 с.
Сочава В.Б. Растительный покров на тематических картах. Новосибирск: Наука, 1979. 189 с.
Сочава В.Б. Географические аспекты сибирской тайги. Новосибирск: Наука, 1980. 256 с.
Спрыгин И.И. Материалы к познанию растительности Среднего Поволжья // Научное наследство. Т. 11. М.: Наука, 1986 (1926). 512 с.
Урусов В.М. Генезис растительности и рациональное природопользование на Дальнем Востоке. Владивосток: ТИГ ДВО АН СССР, 1988. 356 с.
Харбух Дж., Бонэм-Картер Г. Моделирование на ЭВМ в геологии. М.: Мир, 1974. 319 с.
Юрцев Б.А. Проблемы ботанической географии Северо-Восточной Азии. Л.: Наука, 1974. 159 с.
Climate change 1995. The science of climatic change / Eds J.T. Houghton, L.G. Meira Filho, B.A. Callander Cambridge, UK: Cambridge Univ. Press, 1996. 572 p.
Hansen J., Sato M., Ruedy R. et al. Climate simulations for 1880–2003 with GISS model E // Climate Dynamics. 2007. V. 29. P. 661–696.
Harbaugh J.W., Bonham-Carter G. Computer simulation in geology. N.Y., L., Sydnye, Toronto: Wiley-Interscience, 1970. 319 p.
Hopkins D.M. The paleogeography and climatic history of Beringia during late Cenozoic Time // Inter-Nord. 1972. V. 12. P. 121–150.
Hulten E. Studies on the origin and distribution of the flora in Kurile Islands // Bot. Not. Lund. 1933. P. 325–343.
Nakamura Y., Krestov P.V. Coniferous forests of the temperate zone of Asia // Coniferous forests. Ser. Ecosystems of the World. 2005. V. 6. P. 163–220.
Odum Eu.P. Basic ecology. Philadelphia: CBS College Publishing, 1983. V. 1. 328 p. V. 2. 376 p.
Pope V.D., Gallani M.L., Rowntree P.R., Stratton R.A. The impact of new physical parametrizations in Hadley Centre climate model – HadCM3 // Climate Dynamics. 2000. V. 16. P. 123–146.
Rice R.J. Fundamentals of geomorphology. London: Longman, 1978. 574 p.
Ricklefs R.E. The Economy of nature. Portland, Oregon: Chiron Press Inc., 1976. 424 p.
Theurillat J.-P., Felber F., Geissler P. et al. Sensitivity of plant and soil ecosystems of the Alps to climate change // Views from the Alps: regional perspectives on climate change. Cambridge, Massachusetts: MIT Press, 1998. P. 225–308.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии