

Успехи современной биологии, 2019, T. 139, № 3, стр. 302-309
О гетерохронии покровительственной окраски кабарги Moschus moschiferus L. (Moschidae, Cetartiodactyla)
В. И. Приходько *
Институт проблем экологии и эволюции им. А.Н. Северцова РАН
Москва, Россия
* E-mail: pvi-1949@mail.ru
Поступила в редакцию 15.01.2019
После доработки 15.01.2019
Принята к публикации 15.01.2019
Аннотация
Приведены результаты собственных 38-летних исследований изменчивости покровительственной окраски кабарги Moschus moschiferus L. в природных популяциях и при разведении в условиях неволи. Выделено три типа окраски: темный, промежуточный и светлый. Все типы окраски кабарги из северной части ареала закладываются в эмбриогенезе. На ранней стадии онтогенеза происходит трансформация окраски: светлый и промежуточный типы замещаются на более темные тона, вплоть до исходного темного типа. Темпы и направление преобразований окрасочных признаков обусловлены численностью популяций и экспериментальных групп, в которых возрастает роль инбридинга как важного фактора их перекомбинации в онтогенезе. Наличие у животных целого комплекса окрасочных признаков, обусловленных их сменой на ранних стадиях онтогенеза, представляет собой широко распространенную форму гетерохронии у кабарги. Обсуждается роль покровительственной окраски как критерия в таксономических исследованиях и при мониторинге за флюктуирующими популяциями для оценки уровней снижения численности и уязвимости вида.
ВВЕДЕНИЕ
Гетерохрония трактуется как изменение сроков и темпов перестройки органов и признаков в ходе филогенеза или на стадиях онтогенеза. Геккель (Haeckel, 1866), первым исследовавший этот феномен, выделял два направления закладки и индивидуального развития структур организма, которые выявляются только при сравнительном анализе (Матвеев, 1945). А.Н. Северцов (Северцов, 1949) полагал, что при гетерохронии не создаются новые признаки или органы, а происходит лишь перекомбинация уже существующих. Однако роль изменчивости эпигенетического характера в микроэволюции остается во многом неясной. Причем одна из главных нерешенных проблем состоит в отсутствии выраженных корреляций между фенотипической изменчивостью и генетической (Шмальгаузен, 1939; Шварц, 1980).
Как было показано в отечественных работах, наиболее быстрые наследственные изменения признаков, включая окраску, происходят при доместикации диких животных. Процесс одомашнивания сопровождается возникновением наследственных перестроек, обусловленных изменениями поведения при направленной селекции среди животных. На серебристо-черной лисице и представителях грызунов выявлено влияние доместикации не только на окраску, репродукцию, но и на процессы индивидуального развития (Беляев, 1972, 1987; Трут, 2000; Васильев и др., 2004). Результаты многолетних исследований легли в основу разработки концепции дестабилизирующего отбора (Беляев, 1983). Темпы и направление фенотипических преобразований при доместикации обусловлены, как полагает ряд исследователей, гетерохронией (Васильев и др., 2004).
Изменчивость окраски следует рассматривать как отдельную форму гетерохронии, если ее вариации проявляются в разное время на стадиях онтогенеза. Работ о влиянии доместикации, приручения или средовых факторов на процесс смены окрасочных признаков волосяного покрова у копытных до сих пор нет. Ранее было показано наличие широкой изменчивости окраски у подвидовых форм кабарги (Соколов, Приходько, 1998). Цель работы – исследование закономерностей реализации и преобразования покровительственной окраски у животных этого вида в природе и при разведении в условиях неволи.
МАТЕРИАЛЫ И МЕТОДЫ
В работе использовали музейные коллекции (Зоологический музей МГУ, Москва; Зоологический институт РАН, Санкт-Петербург) и собственные сборы шкур кабарги из шести географических популяций. Материалом для представленной работы послужили также дикие животные, встреченные при работе в природе (на территориях бывшего СССР, Монголии, Индии) и литературные источники (Цалкин, 1947; Егоров, 1965; Gao, 1963). Общая выборка составляет 197 экземпляров (табл. 1). В результате обработки материала выделено три типа покровительственной окраски у исследованного вида.
Таблица 1.
Соотношение типов окраски волосяного покрова осмотренных шкур (экз.) кабарги из популяций разных регионов
| Тип окраски | Регион | |||||||
|---|---|---|---|---|---|---|---|---|
| Алтай, Саяны | Забайкалье, Бурятия |
Якутия, Северо-Восток Сибири |
Приморье | о. Сахалин | Монголия | Непал, Индия |
Китай, Бирма, Вьетнам |
|
| Темный | 16 (53)* | 23 (49) | 9 (32) | 14 (61) | 1 (11) | 3 (21) | 7 (27) | 5 (20) |
| Светлый | 6 (20) | 15 (32) | 3 (11) | 3 (13) | 3 (33) | 5 (36) | 14 (54) | 15 (60) |
| Промежуточный | 7 (23) | 9 (19) | 16 (57) | 6 (26) | 4 (45) | 6 (43) | 5 (19) | 5 (20) |
| Белый (альбинос) | 1 (3) | 1 (11) | ||||||
1. Темный тип – голова окрашена светлее туловища, кончики ушей черные, хорошо выражена пятнистость, общий фон окраски буровато-коричневый, кончики кроющих остевых волос окрашены в черный или темно-коричневый цвет, зонарное кольцо светлое или белое шириной 1–3 мм.
2. Светлый тип – голова окрашена темнее туловища, общий фон окраски светло-коричневый с легкой штриховатостью, пятнистость отсутствует или слабо выражена, кончики ушей светлые. Основная масса волос имеет желтоватое зонарное кольцо шириной 3–5 мм, кончики остевых волос светло-коричневые.
3. Промежуточный тип – шея и передняя часть туловища окрашены светлее задней части тела, кончики ушей темные, пятнистость выражена слабо, зонарное кольцо широкое 3–8 мм, кончики остевых волос коричневые.
В ходе экспериментальной работы на научно-экспериментальной базе “Черноголовка” ИПЭЭ РАН исследованы окрасочные признаки потомков от диких животных, основателями которых были четыре самца и пять самок репродуктивного возраста. Указанное количество особей соответствует среднему числу и половому составу внутрипопуляционной группировки (дема) вида в природе (Приходько, 2003). Всего за период с 1977 г. по 2015 г. получено 227 телят, включая пять особей седьмого поколения. Подбор пар осуществлялся лишь в целях исключения и минимизации инбридинга при длительном разведении животных в неволе. Отбор путем селекции по признакам оборонительного поведения и приручения не проводился.
Темпы и направление окрасочных преобразований в ходе онтогенеза исследовали на 19 потомках первого поколения от пяти диких особей-основателей двух экспериментальных групп, стоки которых принадлежат к разным популяциям (табл. 2). Для статистической обработки данных использовали кластерный анализ в программе STATISTICA v. 6.0.
Таблица 2.
Распределение типов окраски кабарги в природе и при скрещивании животных в неволе
| Тип окраски |
Дикие животные Забайкалье, Бурятия |
Результаты экспериментального скрещивания: потомки первого поколения |
|
|---|---|---|---|
| при скрещивании животных из иркутского дема (группа 1) |
при скрещивании животных из бурятской и иркутской популяций (группа 2) |
||
| Темный | 23 (49)* | 0 | 4 (31) |
| Промежуточный | 9 (19) | 2 (33) | 6 (46) |
| Светлый | 15 (32) | 4 (67) | 3 (23) |
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Гетерохрония окраски животных из природных популяций
Приспособительный характер окраски волосяного покрова известен давно и свойствен всем животным. Назначение окраски часто объясняется ее покровительственной функцией. Разнообразие окрасочных структур, как было показано на огромном фактическом материале, всегда связано с экологией видов, особенно со средой их обитания (Котт, 1950). Как свидетельствуют литературные данные, во многих сибирских популяциях кабарги встречаются отдельные особи как с темной, так и с относительно светлой окраской волосяного покрова (Цалкин, 1947; Егоров, 1965). Животные с разным типом окраски обнаружены нами практически во всех природных популяциях и изменчивость по этому признаку обусловлена ранними сроками ее закладки.
Частота встречаемости крайних вариантов фенотипа у подвидов кабарги в природных популяциях различна. Цифровой материал, приведенный в табл. 1, показывает, что в северной части ареала животные с темным и промежуточными типами окраски преобладают над животными со светлым типом. У кабарги Сибири прослеживается также тенденция к усилению темных тонов в окраске волосяного покрова в направлении с запада на восток, а также с северо-востока на юг к Приморью. Кабарга, заселяющая обширный Китай-Индо-Тибетский регион, также неоднородна по окраске: в природе встречаются как темноокрашенные, так и светлоокрашенные особи. К темному типу исследователи часто относят животных с промежуточным типом окраски, как трудно распознаваемым и связанным с серией постепенных переходов (Groves, 1975; Grubb, 1982).
При осмотре коллекций шкур кабарги установлено, что темный тип окраски свойствен взрослым животным, а светлый в большинстве случаев – полувзрослым особям. Различия в окраске волосяного покрова между северными и южными подвидами кабарги определяются в основном разным типом окраски остевых волос и характером их распределения по туловищу животного (рис. 1а,б). Пятнистость, штриховатость, темная или рыжая окраска верхушек остевых волос – полиморфные фенотипические признаки. Различия в покровительственной окраске между группами подвидов кабарги связаны с ее изменчивостью, обусловленной снижением численности вида в границах всего ареала и возможным инбридингом в фрагментированных и изолированных демах (Лю, Шэн, 2002; Приходько, 2018а).

Гетерохрония окраски при скрещивании животных в неволе
Межпопуляционные различия по фенетическим признакам обусловлены главным образом их генетическим своеобразием (Тимофеев-Ресовский и др., 1973). Между тем характер фенотипических реализаций наследственной программы играет решающую роль при изменении условий среды (Шварц, 1980). Ранее нами было показано, что изменения в покровительственной окраске могут быть вызваны также инбридингом (Соколов, Приходько, 1998; Приходько, 2003). Особенности наследования и становления окраски волосяного покрова кабарги исследованы при разведении животных в неволе. Родительские особи были представлены из популяций с генетически разными стоками. Животные из иркутской популяции были отловлены в пространственно изолированном деме, численность которого не превышала 5–7 особей. В Бурятии, где также проводился отлов животных, экологическая плотность достигала высоких показателей.
При скрещивании в неволе особей светлого и темного типа, а также при скрещивании потомков со сходным окрасочным нарядом были выделены, как и в природе, три группы животных, различающихся фенотипически: темные, промежуточные и светлые (табл. 2). Телята с темным типом окраски в группе из стока иркутского дема в пометах не отмечены. Подавляющее большинство животных первого поколения было окрашено по светлому типу.
При ассортативном скрещивании особей из разных популяций было получено наиболее пестрое по фенотипу потомство, но с преобладанием промежуточных форм. По-видимому, генетические процессы, идущие в симпатрических и аллопатрических популяциях, принципиально различны. Животные с крайними типами окраски оказались ближе к средним нормам родительских особей. Соотношение этих форм, полученных при скрещивании, составляет 4 : 8 : 7 и носит закономерный характер, то есть в целом соответствует соотношению 1.3 : 1. Такое соотношение характерно для дигибридного расщепления при наличии комплементарных генов. Приняв это допущение, можно предположить, что окраска кабарги контролируется двумя парами аллелей: Аа и Bb. Темный тип окраски определяется присутствием доминантных аллелей, то есть темноокрашенные особи имеют генотип [A–, B–]. При любых других сочетаниях генов: [A–, bb], [aa, B–] или [aa, bb], окраска животных имеет светлый (штриховатый) тип. Варианты дигибридного сочетания генов объясняют, почему при скрещивании между собой особей с разными фенотипами происходит выщепление всех типов окраски.
Как показали длительные визуальные наблюдения, светлый тип окраски нестабилен. Как в природе, так и в неволе светло окрашенные животные с возрастом темнеют и могут быть отнесены к группе с промежуточным типом окраски. Потемнение отмечено также у животных с промежуточным окрасочным типом. Более того, все типы окраски кабарги из северной части ареала закладываются в эмбриогенезе. Смена светлого и промежуточного на более темный тип окраски происходит быстрыми темпами – спустя три года после отела, то есть на начальной стадии онтогенеза. Вероятно, перекомбинация окраски и низкие частоты рецессивных аллелей объясняют малочисленность особей со светлым типом окраски в северной части ареала.
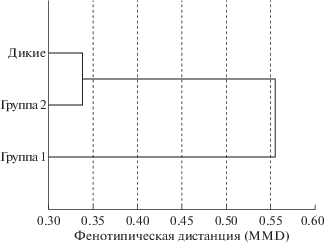
Особый интерес представляют данные, полученные при сравнении уровней окрасочных признаков животных, разводимых в экспериментальных условиях, при сопоставлении с фенотипической изменчивостью, наблюдаемой у диких особей из бурятско-забайкальской популяции. Все исходные животные и образцы шкур от диких особей относятся к сибирскому подвиду M. m. moschiferus L. С этой целью были вычислены фенотипические дистанции между шкурами природной выборки и шкурами животных, обозначенных как дикие, и двумя группами потомков, полученных в экспериментальных условиях. На рис. 2 показаны результаты кластерного анализа, проведенного методом UPGMA. Выделяются два кластера, один из которых обособленный, объединяет природную выборку диких животных и экспериментальную гибридную группу. Другой, представленный внешней ветвью кластера, включает группу потомков экспериментальных животных из иркутского дема, имеет высокий уровень фенотипического обособления, который составляет 0.555. Такие значения фенотипических дистанций характерны при сравнении данных, полученных для хорошо дифференцированных географических популяций, принадлежащих к разным подвидам. Таким образом, проявление комплекса окрасочных признаков в природных популяциях и при гибридном скрещивании оказалось сходным. Из этого следует, что для поддержания баланса гетерохронии окраски в природе требуется межпопуляционный обмен особями. Такой обмен осуществляется регулярно и возможен только при достижении определенной плотности населения в пространственных стратах вида (Приходько, 2018б). Механизмом выселения лишних особей, в основном молодых, с заселенного пространства выступает территориальность, свойственная кабарге.
Рис. 2.
Кластерный анализ фенотипических дистанций (MMD) между выборками природной популяции и экспериментальных групп (табл. 2). Данные обработаны с применением программы STATISTICA v. 6.0.
Уже со второго поколения инбридинг, избежать которого не удалось, приводил к снижению доли новорожденных с доминантным темным типом окраски, свойственной диким животным. В каждом поколении инбридинг действовал как крайне неблагоприятный фактор. По мере возрастания инбредной депрессии в поколениях кабарги наблюдается также снижение воспроизводства через увеличение смертности и отчасти за счет потери репродуктивного потенциала. В нашем случае снижение жизнеспособности (выживаемости) телят и плодовитости самок сопровождалось сдвигом соотношения полов в приплоде в сторону самцов. От одной самки седьмого поколения нам не удалось получить приплод в течение 15 лет ее жизни.
При разведении животных в неволе, было также установлено, что уровни воспроизводства и проявления “дикой” окраски наиболее сильно снижаются между вторым и четвертым поколениями. Существенные изменения наблюдаются и в смертности телят, которая составляет 25–30% в каждом поколении вплоть до четвертого. В последующих поколениях потеря остающейся “дикой” окраски происходит с темпами 8–10%, а жизнеспособность телят падает по 7–10% в каждой генерации. С четвертого поколения почти все телята рождаются со светлым типом окраски, что было не свойственно особям-сеголеткам до второго поколения. Следовательно, жизнеспособность и другие аспекты приспособленности к условиям содержания снижаются пропорционально уменьшению доли животных с темной окраской, которая, в свою очередь, связана обратной зависимостью с размером групп в неволе и возрастанием в них роли инбридинга.
Результаты исследований скорости и направления гетерохронии окраски кабарги имеют научное и природоохранное значение. Окраска волосяного покрова часто используется как диагностический признак для уточнения систематического ранга отдельных групп млекопитающих (Соколов, 1973; Чернова, 2003; Чернова, Целикова, 2004; Hausman, 1924; Noback, 1951). Для понимания таксономических связей внутри рода Moschus необходимо учитывать особенности окраски волосяного покрова у животных северного и южного подвидов. У китайского подвида M. m. chrysogaster географическая и индивидуальная изменчивость в основном тоне окраски прослеживается достаточно четко. Среди животных встречаются как светлые, так и относительно темные особи (Wang et al., 1993). По этому признаку темноокрашенных особей нередко относят в отдельный вид Moschus berezovskii F. (Flerov, 1928; Gao, 1963; Grubb, 1982; Wang et al., 1993; Groves et al., 1995). Кабарга М. Березовского M. berezovskii F., ранее выделенная (Flerov, 1928) как самостоятельный вид, является лишь цветовой морфой M. m. chrysogaster H., у которой наблюдается процесс возвращения к исходному типу окраски (Соколов, Приходько, 1998). Однако подобный возврат к окрасочным чертам прошлого весьма длителен, если вообще возможен. Некоторые аллели, отвечающие за наследование целого ряда признаков окраски кабарги, могли быть утрачены в прошлом.
Природные изменения на рубеже плиоцена–плейстоцена могли привести к разрыву в недалеком прошлом обширного ареала и установлению между северными и южными подвидами пространственной изоляции. Вероятно, именно изоляция, а также широкая изменчивость окраски волосяного покрова позволили исследователям выделить в пределах ареала от трех до шести видов (Флеров, 1930; Flerov, 1928; Hodgson, 1839; Gao, 1963; Groves, 1975; Grubb, 1982). Однако при этом южные и северные кабарги кариологически однотипны (Соколов, Приходько, 1998). Некоторые гетерохронии, например расчленяющая окраска шеи, достигли различий, выходящих за рамки подвидового уровня. Размеры волос у обеих форм однотипны и достоверно не различаются. Не обнаружено различий и в микроструктуре волос (Sheng et al., 1993). В то же время различия в расчленяющей окраске шеи хорошо выражены даже у телят в ювенильном наряде, что свидетельствует о далеко зашедшей дивергенции генных окрасочных комплексов у сравниваемых форм. Расчленяющая шею окраска кабарги – древнейший признак, унаследованнный, по-видимому, от представителя рода Archaeomeryx как исходной группы для некоторых жвачных, в том числе и для кабарговых.
Окрасочные признаки необходимо использовать при мониторинге за флюктуирующими популяциями для оценки уязвимости вида. Появление в природных группировках особей со светлым типом окраски волосяного покрова свидетельствует о снижении численности подвида и даже вида. В последние годы в природе происходит осветление окраски волосяного покрова сибирской кабарги, что свидетельствует о низкой численности внутрипопуляционных группировок в условиях их возрастающей мозаичности и изоляции (Приходько, 2018а). В северной части ареала, по мере ежегодного сокращения численности вида, например в Иркутской области, Забайкалье и Горном Алтае, растет количество случаев появления в группировках животных со светлой окраской, проявляющейся при инбредной депрессии. Группировки, преимущественно с низкой численностью, сохранились на юге ареала лишь в локальных и изолированных зонах обитания, в основном на территориях заповедников и национальных парков. Их генетическое разнообразие, по-видимому, достигло низкого уровня. Последствия отбора в нежизнеспособных группировках, скорее всего, приведут к постепенному исчезновению животных с покровительственной окраской, к преобладанию в поколениях особей со светлым типом волосяного покрова. В последние годы это явление приобретает общепопуляционный характер, являясь самым заметным микроэволюционным направлением в изменении покровительственной окраски современной кабарги.
ЗАКЛЮЧЕНИЕ
Выявленная смена окрасочных признаков в ходе онтогенеза подтверждает точку зрения А.Н. Северцова (Северцов, 1949) об их перекомбинации, при которой светлый признак окраски замещается более темным тоном, причем быстрее, чем предполагалось ранее. Как свидетельствуют полученные данные, развивающийся у телят светлый тип окраски в малочисленных демах и популяциях оказывается функционально не адаптивным и даже губительным для животных из-за демаскирующих особенностей. Дальнейшее развитие этого признака, проявляющегося на ранней стадии онтогенеза, приостанавливается вплоть до тех стадий, когда его замещает более адаптивный темный тип, который эволюционно адекватен горно-таежной среде обитания в северной части ареала. Однотонная темная окраска волосяного покрова является исходной для всех современных форм кабарги. Сочетание разных типов окраски, когда-то существовавших у предковых и сформированных у современных южных форм, следует рассматривать как признак, появляющийся еще при эмбриональном развитии. Следовательно, накопление целого комплекса окрасочных признаков, обусловленных их сменой на ранних стадиях онтогенеза, представляет собой широко распространенную форму гетерохронии у кабарги.
Список литературы
Беляев Д.К. Генетические аспекты доместикации животных // Проблемы доместикации животных и растений / Ред. Б.С. Матвеев. М.: Наука, 1972. С. 39–45.
Беляев Д.К. Дестабилизирующий отбор // Развитие эволюционной теории в СССР (1917–1970-е годы)/ Ред. С.Р. Микулинский, Ю.И. Полянский. Л.: Наука, 1983. С. 266–277.
Беляев Д.К. Проблемы и перспективы исследований по генетике и селекции животных // Генетика. 1987. Т. 23. № 6. С. 937–946.
Егоров О.В. Дикие копытные Якутии. М.: Наука, 1965. 259 с.
Васильев А.Г., Фалеев В.И., Галактионов Ю.Л. и др. Реализация морфологического разнообразия в природных популяциях млекопитающих. Новосибирск: СО РАН, 2004. 232 с.
Котт Х. Приспособительная окраска животных. М.: Иностранная литература, 1950. 543 с.
Лю Чжисяо, Шэн Хэлинь. Влияние изоляции и фрагментации местообитаний на популяцию альпийской кабарги // Экология. 2002. № 2. С. 132–136.
Матвеев Б.С. О системе соотносительных изменений формы, функции и среды в эволюции животных // Зоол. журн. 1945. Т. 24. № 1. С. 3–22.
Приходько В.И. Кабарга: происхождение, систематика, экология, поведение и коммуникация. М.: ГЕОС, 2003. 443 с.
Приходько В.И. Динамика численности кабарги (Moschus moschiferus L.) в России // Вест. охотовед. 2018а. Т. 15. № 1. С. 26–32.
Приходько В.И. Межподвидовая симпатрия у кабарги, Moschus moschiferus L. (Moschidae, Cetartiodactyla) // Бюл. МОИП. Отд. биол. 2018б. Т. 123. Вып. 5. С. 3–9.
Северцов А.Н. Морфологические закономерности эволюции. М.: АН СССР, 1949. Т. 5. 610 с.
Соколов В.Е. Кожный покров млекопитающих. М.: Наука, 1973. 487 с.
Соколов В.Е., Приходько В.И. Систематика кабарги (Artiodactyla, Mammalia). Сообщение 2 // Изв. АН. Сер. биол. 1998. № 1. С. 37–46.
Тимофеев-Ресовский Н.В., Яблоков А.В., Глотов Н.В. Очерк учения о популяциях. М.: Наука, 1973. 278 с.
Трут Л.Н. Проблема дестабилизирующего отбора в развитии // Современные концепции эволюционной генетики / Ред. А.Л. Маркель, В.К. Шумный. Новосибирск: ИЦиГ СО РАН, 2000. С. 7–21.
Флеров К.К. К систематике и географическому распространению кабарги (Moschus, Cervidae) // Ежегод. Зоол. музея АН СССР. 1930. Т. ХХХI. Вып. 1. С. 1–20.
Цалкин В.И. Систематика кабарги // Гепнер В.Г., Цалкин В.И. Олени СССР. М.: МОИП, 1947. С. 120–175.
Чернова О.Ф. Архитектоника и диагностическое значение коры и сердцевины волос // Изв. РАН. Сер. биол. 2003. № 1. С. 73–83.
Чернова О.Ф., Целикова Т.Н. Атлас волос млекопитающих. Тонкая структура остевых волос и игл в сканирующем электронном микроскопе. М.: КМК, 2004. 429 с.
Шварц С.С. Экологические закономерности эволюции. М.: Наука, 1980. 278 с.
Шмальгаузен И.И. Пути и закономерности эволюционного процесса. М.-Л.: АН СССР, 1939. 231 с.
Flerov C.C. Preliminary note on the diagnostic characters of the genus Moschus Linne (Mammalia, Cervidae) // C. R. Acad. Sci. USSR (A). 1928. V. 24. P. 515–519.
Gao Y.-T. Taxonomic notes on the Chinese musk deer // Acta Zoologica Sinica. 1963. № 15. P. 479–488.
Groves C.P. The taxonomy of the genus Moschus (Mammalia, Artiodactyla), with particular reference to the Indian region // J. Bombay Nat. His. Soc. 1975. V. 72. P. 662–676.
Groves C.P., Wang Y.-X., Grubb P. Taxonomy of musk deer, genus Moschus (Moschidae, Mammalia) // Acta Theriol. Sinica. 1995. V. 15. № 3. P. 181–197.
Grubb P. The systematics of Sino-Himalayan musk deer (Moschus), with particular reference to the species described by Hodgson B.N. // Saugetierkundliche Mitteilungen. 1982. B. 30. S. 127–135.
Haeckel E. Generelle Morphologie der Organismen: allgemene Grundzüge der organismdien Formen-Wissenschaft mechanisch begründet durch die von Charles Darwin reformirte Descendenz-Theorie. Berlin: Verlag von Georg Reimer, 1866. Bd. 1–2. 652 s.
Hausman L.A. Further studies in the relationships of the structural characteristics of mammalian hair // Am. Nat. 1924. V. 58. P. 544–557.
Hodgson B.H. On three new species of musk deer (Moschus) inhabiting the Himalayan districts // J. As. Soc. Bengal. 1839. V. 8. P. 202–203.
Noback Ch.R. Morphology and phylogeny of hair // Ann. N.Y. Acad. Sci. 1951. V. 53. P. 476–491.
Sheng H., Zhang E., Cher G., Ni B. A comparative study on morphology of deer hair // Deer of China – biology and management / Eds N. Ohtaishi, H.L. Sheng. Amsterdam: Elsevier Science Publishers, 1993. P. 73–79.
Wang Y.-X., Ma S., Li C. The taxonomy, distribution and status of forest musk deer in China // Deer of China – biology and management / Eds N. Ohtaishi, H.L. Sheng. Amsterdam: Elsevier Science Publishers, 1993. P. 22–30.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии