

Успехи современной биологии, 2019, T. 139, № 4, стр. 315-325
Ассортативное скрещивание в гибридных зонах обыкновенной бурозубки, Sorex araneus L. (Soricidae, Soricomorpha)
В. Н. Орлов 1, Д. М. Кривоногов 2, Е. В. Черепанова 1, А. В. Щегольков 2, О. О. Григорьева 1, *
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
Москва, Россия
2 Нижегородский государственный университет им. Н.И. Лобачевского, Арзамасский филиал
Арзамас, Россия
* E-mail: grig@sevin.ru
Поступила в редакцию 30.01.2019
После доработки 15.03.2019
Принята к публикации 15.03.2019
Аннотация
Приводится количественная оценка ассортативности скрещиваний (R) как показателя “reinforcement” в 12 гибридных зонах между хромосомными расами обыкновенной бурозубки Sorex araneus L. Рассматривается эффект Валунда в гибридной популяции и возможности его уменьшения. В гибридных зонах с гетерозиготами CIV, CV и RIV индекс R изменяется в пределах 0.23–0.59, а в гибридных зонах с более сложными гетерозиготами (RVI, СIХ–СХI) превышает 0.82. Поток генов в гибридной зоне прерывается при R > 0.99. Ассортативное скрещивание не только уменьшает поток генов, но и поддерживает стабильность генотипов в гибридных популяциях.
ВВЕДЕНИЕ
В концепции географического видообразования необходимым условием считают длительную изоляцию популяций, а географические изоляты рассматривают в качестве новых зарождающихся видов. В случае вторичного контакта ранее изолированных популяций между ними возникают разнообразные зоны аллопатрической гибридизации (Mayr, 1963). Использование этого термина полезно, когда надо подчеркнуть, что гибридная зона возникла в результате подобного вторичного контакта. Современные географические изоляты известны, как правило, на периферии видового ареала, в то время как зоны аллопатрической гибридизации могут обнаруживаться в любой части ареала вида. Их расположение определяется прошлой рефугиальной структурой вида.
В зонах аллопатрической гибридизации возможен reinforcement – усиление постзиготической изоляции в результате появления презиготических барьеров (Coyne, Orr, 2004). Поэтому именно зоны аллопатрической гибридизации рассматривают в качестве ключевых объектов для исследования процессов видообразования (Jiggins, Mallet, 2000; Shurtliff, 2013). Но инструментов для изучения презиготической изоляции в гибридных зонах млекопитающих немного. Это традиционные методы оценки поведенческой изоляции, когда сравнивают контактирующие формы in vitro на особях, взятых из зон аллопатрии и гибридизации. Например, так была показана частичная этологическая изоляция в зонах аллопатрической гибридизации домовых мышей Mus m. musculus и M. m. domesticus (Bímová et al., 2011) и филогрупп обыкновенной полевки Microtus arvalis (Beysard et al., 2015). Подобные прямые методы оценки этологической изоляции контактирующих форм ограничены видами, с которыми удобно работать in vitro.
Хромосомный полиморфизм обыкновенной бурозубки Sorex araneus L. привлекает внимание цитогенетиков с 1970-х гг. и активно исследуется последние 30 лет. На всем огромном протяжении ареала этого палеарктического вида в настоящее время описано более 70 хромосомных рас, участков ареала с одинаковым набором Робертсоновских (Rb) транслокаций или без них (Щипанов, Павлова, 2016а). Гибридные зоны между хромосомными расами обыкновенной бурозубки начали изучать с 1980-х гг., им посвящено много работ и несколько обзоров (Щипанов и др., 2009; Щипанов, Павлова, 2016б; Wόjcik et al., 2002). При этом совершенно неизученной остается презиготическая изоляция из-за трудностей прямой оценки этологической изоляции бурозубок in vitro. Кроме того, презиготическая изоляция у обыкновенной бурозубки может быть следствием конкуренции гамет при множественных копуляциях, хорошо известных у этого вида (Dehnel, 1952).
Неоднократно отмечалось, что для количественной оценки презиготической изоляции in vivo в зонах вторичного контакта и гибридизации популяций можно использовать дефицит гетерозигот, то есть отклонения найденных частот аллозимов или аллелей от ожидаемых в предположении случайного скрещивания по Харди–Вайнбергу (Jiggins, Mallet, 2000). В гибридных зонах обыкновенной бурозубки и многих других видов индекс инбридинга (FIS) может быть одним из немногих доступных в настоящее время показателей презиготической изоляции. На дефицит гетерозигот по гаплоидным наборам хромосом в гибридных зонах обыкновенной бурозубки неоднократно обращали внимание (Щипанов и др., 2008; Орлов и др., 2012, 2013а,б; Черепанова и др., 2018; Bulatova et al., 2011), но приложение этого показателя к сравнительным исследованиям гибридных зон вызывает много вопросов. Прежде всего, остается неясным, как влияет подразделенность гибридной популяции на дефицит гетерозигот (эффект Валунда). Случаи использования ассортативного скрещивания в микроэволюционных исследованиях единичны (Bearhop et al., 2005).
Статья посвящена применению количественных показателей ассортативности скрещиваний и дефицита гетерозигот с учетом подразделенности популяций для оценки презиготической изоляции контактирующих хромосомных рас обыкновенной бурозубки.
МАТЕРИАЛЫ И МЕТОДЫ
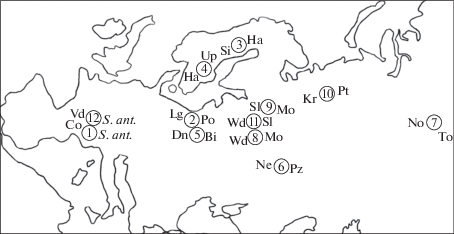
Материалом для статьи послужили опубликованные данные о распределении бурозубок гомозиготных и гетерозиготных по гаплоидным наборам хромосом с различными Rb-транслокациям в 12 гибридных зонах обыкновенной бурозубки, в том числе трех ранее описанных с участием авторов этой статьи (рис. 1).
Рис. 1.
Карта зон контакта между хромосомными расами обыкновенной бурозубки. Номера гибридных зон соответствуют таблице.
В статистических расчетах мы исходили из однолокусной модели с кодоминантным типом наследования. Расчеты основывались на матрицах генотипов особей. Из численности особей с различными генотипами были рассчитаны основные параметры генетической изменчивости выборок, в том числе: наблюдаемые частоты генотипов (Р0, H0, Q0), ожидаемые частоты генотипов (РЕ, HЕ, QЕ) в предположении случайного скрещивания по Харди–Вайнбергу, достоверность их различий (критерий χ2), частоты гаплоидных наборов хромосом (p и q), индексы: ассортативности скрещиваний R и инбридинга FIS = 1 – H0/HE (Weir, Cockerham, 1984). Чтобы наглядно показать зависимость найденных и ожидаемых генотипических частот, мы используем вариансу частот аллеля Vq. Наблюдаемые частоты гомозигот, по сравнению с ожидаемыми, повышены на величину вариансы Vq (Р0 = = p2 + Vq; Q0 = q2 + Vq), а наблюдаемые частоты гетерозигот понижены на удвоенную величину вариансы (H0 = 2pq – 2Vq).
Степень положительной ассортативности скрещиваний описывают пропорциями ассортативных (R) и случайных (Т) скрещиваний, при этом R + T = 1 (Crow, Felsenstein, 1968; Crow, Kimura, 1970; Hedrick, 1983). Индекс R рассчитан по потомству трех типов скрещиваний (Р × Р, Р × Н и Н × Н), которые определяют величину доли гомозигот Р1 в следующем поколении:
P1 = Rp2/(1 – Q0) + Тр2 (Hedrick, 1983: формула 5.24). Эта формула была разработана для случаев частичного ассортативного скрещивания между доминантными и рецессивными генотипами. Мы использовали эту формулу, поскольку при кодоминантном наследовании доля гомозигот Р1 в следующем поколении также определяется величиной потомства в приведенных выше трех типах скрещиваний. При этом мы исходили из предположения, что длительно существующие гибридные популяции находятся в равновесии и P1 = P0. Индекс R рассчитан с точностью до 0.01 (в отдельных случаях до 0.001) при наименьшей сумме квадратов отклонений долей найденных и ожидаемых генотипов в следующем поколении Р1.
Обработка генетических данных проведена в пакете GenAlEx (Peakall, Smouse, 2006), FSTAT 2.9.3.2, корреляционный анализ проведен в пакете Statistica 6.0. Для оценки вероятности отличия FIS от 0 было применено стандартное отклонение методом складного ножа, и вычислен 95%-ный доверительный интервал путем проведения бутстреп-анализа (15 000 реплик). Генетические показатели, характеризующие выборки из гибридных популяций, приведены в таблице.
Таблица
Частоты генотипов в гибридных зонах обыкновенной бурозубки. Выборки взяты в краевой (+), центральной (++) или по всей ширине гибридной зоны (+++)
| № гиб-ридной зоны | Гибридные зоны, субпопуляции, выборки, регион | Длина исследо-ванных трансект, км | Диагностические хромосомы (мультиваленты) | Численность генотипов, найденная |
Частоты генотипов, найденные (P0,H0,Q0) |
Частоты генотипов, ожидаемые (PE, HE, QE) |
χ2 | Варианса частот аллеля Vq |
q | FIS | R | Источник |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Cordon (Со)/Sorex antinorii Bon. (S. ant.) пункт 5, Les Gens, Франция | 0.1 + |
jl, h, o/hj, lo (CV) |
Сo – 44 H – 10 S. ant.– 1 |
0.800 0.182 0.018 |
0.794 0.194 0.012 |
0.23 NS |
0.006 | 0.109 | 0.062 NS |
0.42 | Brünner, Hausser, 1996 |
| 2 | Popielno (Ро)/Łȩgucki Młyn (Łg), пункты KII, ZL, Z, Польша | 5.0 + |
ik, h/hk, i (CIV), ik, o/io, k (CIV) ik, hq/hk, i, q (CV) |
Po – 22 H – 20 Łg – 9 |
0.431 0.392 0.177 |
0.393 0.468 0.139 |
1.33 NS |
0.038 | 0.373 | 0.162 * |
0.45 | Moska, 2003 |
| 3 | Hattsujö (Ha) (=Abisco)/ Sidensjö (Si), пункты 5–6, 8–11, Швеция |
10.0 + |
hn, i/hi, n (CIV) | Ha – 47 H – 28 Si – 10 |
0.553 0.329 0.118 |
0.515 0.405 0.080 |
2.98 NS |
0.038 | 0.282 | 0.188 * |
0.55 | Fredga, Narain, 2000; Fredga, 2007 |
| 4а | Hӓllefors (Hӓ)/Uppsala (Up), субпопуляция Hӓllefors, пункты 8–10, 12–14, зап. реки, Швеция |
2.0 + |
ko, pq/kp, oq (RIV) | Hӓ – 46 H – 28 Up – 7 |
0.568 0.346 0.086 |
0.549 0.384 0.067 |
0.81 NS |
0.019 | 0.259 | 0.099 * |
0.37 | Narain, Fredga, 1996: табл.1 |
| 4б | Hӓllefors (Hӓ)/Uppsala (Up), субпопуляция Uppsala, пункты 11, 15, вост. реки, Швеция |
<1.0 + |
ko, pq/kp, oq (RIV) | Hӓ – 5 H – 21 Up – 35 |
0.082 0.344 0.574 |
0.065 0.379 0.556 |
0.51 NS |
0.017 | 0.254 | 0.092 * |
0.20 | Narain, Fredga, 1996: табл.1 |
| 5а | Drnholec (Dn)/Białowieża (Bi), субпопуляция Drnholec, пункты P1, Р5, Р8, Р9, R2, Польша | 0.3 + |
gm, hi, ko, nr, p/gr, hn, ik, mp, o (CX), gm, p/mp, g (CIV) | Dn – 2 H – 20 Bi – 48 |
0.028 0.286 0.686 |
0.029 0.283 0.687 |
0.002 NS |
0.001 | 0.171 | 0.01 NS |
0.00 | Szałaj et al., 1996 |
| 5б | Drnholec (Dn)/Białowieża (Bi), субпопуляция Białowieża, пункты P2, Р4, Р6, Р7, Р10,
R1, R3–R5, Польша |
0.3 + |
gm, hi, ko, nr, p/gr, hn, ik, mp, o (CX), gm, p/mp, g (CIV) | Dn – 75 H – 15 Bi – 6 |
0.781 0.156 0.063 |
0.738 0.242 0.020 |
12.00 *** |
0.043 | 0.141 | 0.355 ** |
0.87 | Szałaj et al., 1996 |
| 6 | Пенза (Pz) / Нерусса (Ne), Рязанская обл. | 0.9 + |
gm, hn, io/go, hi, mn (RVI) | Pz – 46 H – 11 Ne – 4 |
0.754 0.180 0.066 |
0.713 0.263 0.024 |
6.02 * |
0.042 | 0.156 | 0.316 * |
0.83 | Черепанова и др., 2018 |
| 7а | Новосибирск (No)/Томск (To), субпопуляция Новосибирск, 5.5–10.06 км трансекты, Западная Сибирь |
4.1 + |
go, hn, ik, mp/gk, hi, mn, o, p (CIX) | No – 56 H – 20 To – 10 |
0.651 0.233 0.116 |
0.588 0.358 0.054 |
12.92 *** |
0.063 | 0.233 | 0.349 *** |
0.82 | Polyakov et al., 2011: табл. S2 |
| 7б | Новосибирск (No)/Томск (To), субпопуляция Томск, 10.2–12.8 км трансекты, Западная Сибирь | 2.8 + |
go, hn, ik, mp/gk, hi, mn, o, p (CIX) | No – 18 H – 14 To – 41 |
0.246 0.192 0.562 |
0.117 0.450 0.433 |
24.07 | 0.154 | 0.342 | 0.573 *** |
0.86 | Polyakov et al., 2011: табл. S2 |
| 8 | Западная Двина (Wd)/ Москва (Mo), Валдай | 0.5 ++ |
hk, ip, qr/hi, kr, pq (RVI) | Wd – 15 H – 9 Mo – 13 |
0.406 0.243 0.351 |
0.278 0.499 0.223 |
9.70 ** |
0.126 | 0.473 | 0.513 ** |
0.85 | Орлов и др., 2012 |
| 9а | Селигер (Sl)/Москва (Mo), субпопуляция Москва, 4–5-й км трансекты, Валдай |
2.0 + |
g/o, hn, ik, m/q, pr/gm, hi, kr, no pq (CXI) | Sl – 14 H – 9 Mo – 43 |
0.212 0.136 0.652 |
0.078 0.404 0.518 |
28.93 *** |
0.134 | 0.280 | 0.663 *** |
0.96 | Bulatova et al., 2011: табл. 3 |
| 9б | Селигер (Sl)/Москва (Mo), субпопуляция Селигер, 6–8-й км трансекты, Валдай |
3.0 + |
g/o, hn, ik, m/q, pr/gm, hi, kr, no pq (CXI) | Sl –100 H – 25 Mo – 27 |
0.658 0.164 0.178 |
0.547 0.385 0.068 |
49.81 | 0.110 | 0.280 | 0.574 *** |
0.85 | Bulatova et al., 2011: табл. 3 |
| 10 | Печора (Pt) / Кириллов (Kr), пункты 2–13, р. Мезень | 2.4 ++ |
gi, hn, mo/gm, hi, no (RVI) | Pt – 46 H – 21 Kr – 37 |
0.442 0.202 0.356 |
0.295 0.496 0.209 |
36.59 *** |
0.147 | 0.457 | 0.593 *** |
0.90 | Pavlova, Shchipanov, 2014 |
| 11 | Селигер (Sl)/Западная Двина (Wd), Валдай | 2.5 + |
g/o, hn, ik, m/q, pr/gm, hk, ip, no, qr (CXI, CIX) | Sl – 78 H – 13 Wd – 12 |
0.757 0.126 0.117 |
0.673 0.295 0.032 |
33.67 *** |
0.085 | 0.180 | 0.573 *** |
0.95 | Орлов и др., 2013 |
| 12 | Sorex antinorii Bon. (S. ant.)/ Vaud (Vd), пункты 4–6, Швейцария |
3.0 +++ |
gi, hj, lo, kn, m, r/gm, hi, jl, no, kr (CXI), gi, hj, m, i/gm, hi, jl (CVII), kn, o, r/no, kr (CV) | S. ant. – 81 H – 12 Vd – 43 |
0.596 0.088 0.316 |
0.409 0.461 0.130 |
88.92 *** |
0.186 | 0.360 | 0.809 *** |
0.995 | Brünner et al., 2002 |
ПОЛОЖИТЕЛЬНОЕ АССОРТАТИВНОЕ СКРЕЩИВАНИЕ В ГИБРИДНЫХ ЗОНАХ
У всех изученных самцов обыкновенной бурозубки – комплексных гетерозигот, в мейозе I которых образуются мультиваленты (цепи из 4–11 хромосом, СIV–CХI, или кольца из 4, 6 хромосом, RIV, RVI), отмечались развитые семенники и сперматозоиды в семенных канальцах. Описаны особи с рекомбинантными кариотипами, что свидетельствует о плодовитости гибридов первого поколения (F1) и возвратных скрещиваниях (Bulatova et al., 2011). Тем не менее, по данным исследования мейоза гибридов, частота образования унивалентов в диакинезе и нерасхождений в метафазе I растет по мере усложнения конфигураций спаривания хромосом от простых гетерозигот (тривалентов, CIII) к тетравалентам, CIV, и мультивалентам (Jadwiszczak, Banaszek, 2006; Borodin, 2008). Иногда у комплексных гетерозигот отмечается повышенная смертность половых клеток в гаметогенезе и уменьшение веса семенников, что не связано с нарушениями в метафазе I мейоза (Narain, Fredga, 1998; Jadwiszczak, Banaszek, 2006). Таким образом, пониженная плодовитость комплексных гетерозигот, в отличие от простых гетерозигот, весьма вероятна (Borodin, 2008), хотя оценить ее количественно в настоящее время невозможно.
Естественно ожидать, что, в случае пониженной плодовитости гибридов в зонах контакта ранее изолированных форм, отбор будет формировать ассортативное скрещивание и reinforcement вплоть до полной репродуктивной изоляции контактирующих форм (Noor, 1995; Coyne, Orr, 2004). По этой причине гибриды с явными нарушениями плодовитости не известны в природных популяциях млекопитающих. Показатель ассортативности скрещиваний или оплодотворений может рассматриваться как важный компонент reinforcement. В гибридных зонах между хромосомными расами будет уменьшаться частота скрещиваний между расами и, соответственно, увеличиваться внутри рас.
Как следствие ассортативности скрещиваний в гибридной популяции возникает дефицит гетерозигот и избыток гомозигот по сравнению со случайным скрещиванием по Харди–Вайнбергу. Ассортативное скрещивание сходно с инбридингом в том отношении, что изменяет частоты генотипов, но не аллелей (Hedrick, 1983). Подобно индексу инбридинга, ассортативное скрещивание характеризует степень инбредности популяции и не тождественно с другими показателями reinforcement, например с показателем этологической изоляции. Параметры R и FIS в исследованных гибридных популяциях (таблица) по нашим данным связаны очень сильной прямой связью, r = 0.909 (уровень значимости p < 0.001).
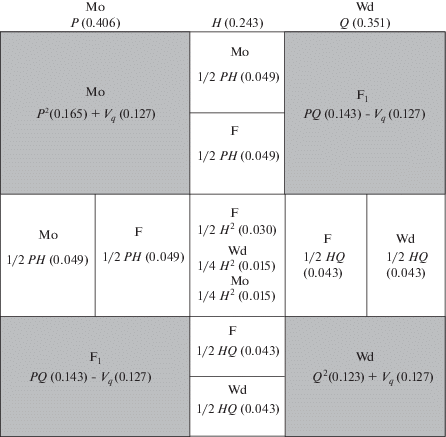
Поскольку у бурозубок происходит ежегодная смена поколений, то наблюдаемые в данном поколении неравновесные частоты генотипов (P0, H0, Q0) должны повториться на следующий год в следующем поколении (P1, H1, Q1). В качестве примера рассмотрим возможное влияние ассортативного скрещивания на частоты генотипов в гибридной зоне хромосомных рас Западная Двина/Москва (таблица, зона 8). Мы выбрали эту исследованную нами ранее гибридную зону (Орлов и др., 2012), чтобы показать, что для такого анализа пригодны даже небольшие выборки (менее 50 особей), но собранные на малых участках гибридной популяции, около 0.25 км2, в наибольшей степени приближенной к панмиксным. Ожидаемые при случайном скрещивании доли генотипов можно наглядно показать на решетке Пеннета (рис. 2). При случайных скрещиваниях в этой гибридной популяции частоты генотипов значительно изменятся в следующем поколении и останутся такими в последующих поколениях (закон Харди–Вайнберга). В данной гибридной зоне неравновесные частоты генотипов будут сохраняться из поколения в поколение только в том случае, если 84.9% случайных скрещиваний или оплодотворений Мо × Wd будут замещены неслучайными Мо × Мо и Wd × Wd (R = 0.849). Тогда частоты каждой гомозиготы увеличатся на вариансу частоты аллеля Vq = 0.127 (Р1 = РE + Vq и Q1 = QE + + Vq), а частоты гетерозигот уменьшатся на 2Vq (H1 = = HE – 2Vq) до значений близких к Р0 и H0 в исходном поколении родителей.
Рис. 2.
Найденная частота генотипов в гибридной популяции хромосомных рас Западная Двина (Wd)/Москва (Мо) (таблица, зона 8) и ожидаемые частоты при условии случайного скрещивания и их коррекция ассортативным скрещиванием (Vq) при R = 0.85. Окрашены области скрещиваний между исходными расами, светлыми оставлены области возвратных скрещиваний и скрещиваний между гибридами.
Расчеты частот генотипов в панмиксных популяциях при положительном ассортативном скрещивании исходят из уравнения последовательности гармонического ряда. Общее выражение для Hn после n поколений ассортативного скрещивания имеет вид: Hn = H0/(1 + nq) (Li, 1976). Следовательно, предельная величина Hn при неограниченном возрастании числа поколений ассортативного скрещивания стремится к 0 (полная изоляция контактирующих форм), однако скорость снижения доли гетерозигот замедляется после нескольких начальных поколений, и доля гетерозигот может стабилизироваться при определенных значениях R (Crow, Kimura, 1970; Hedrick, 1983). Таким образом, количественные показатели ассортативности скрещиваний могут использоваться для оценки презиготической изоляции.
В случае частичных ассортативных скрещиваний в популяции с двухаллельными генотипами и примерно равными долями Р и Q гетерозиготность достигает равновесных значений Н = 0.39 при R = 0.5 и Н = 0.2 при R = 0.9 всего через 20 поколений (Hedrick, 1983). Лишь в трех исследованных гибридных зонах значения q наиболее близки к 0.5: в зоне контакта рас Popielno/Łȩgucki Młyn, Западная Двина/Москва и Печора/Кириллов, и значения Н и R достаточно хорошо совпадают с приведенными теоретическими расчетами (таблица, зоны 2, 8, 10). Стабильность гибридных зон обыкновенной бурозубки на протяжении десятилетий показана для трех гибридных зон с комплексными гетерозиготами (Yannic et al., 2009; Bulatova et al., 2011; Polyakov et al., 2011) и одной – с простыми гетерозиготами (Jones, Searle, 2003). О вероятности равновесных значений частот генотипов можно судить по длительности существования зон контакта хромосомных рас на протяжении всего голоцена – 10 тыс. лет.
ВЛИЯНИЕ ПОДРАЗДЕЛЕННОСТИ ГИБРИДНЫХ ЗОН НА ДЕФИЦИТ ГЕТЕРОЗИГОТ
В использовании показателей ассортативности скрещивания наиболее сложной остается задача минимизации эффекта Валунда. Анализ подразделенности гибридных популяций обыкновенной бурозубки возможен, поскольку этот вид уже много десятилетий служит модельным объектом для популяционно-экологических исследований млекопитающих. Структура гибридных зон, глубина проникновения одной хромосомной расы на территорию другой, зависит не только от успешности скрещиваний и плодовитости гибридов, но и от изменения типов расселения на границе локальных популяций, соотношения в популяциях оседлых особей и нерезидентов, особенностей социальных взаимодействий рас, например частоты дружелюбных тактильных контактов у нерезидентов, и др. (Щипанов и др., 2008а,б).
В гибридной популяции сохраняется граница первоначального контакта расселявшихся хромосомных рас, граница гибридных субпопуляций. Эта граница заметна по преобладанию в выборках одной из контактирующих рас. Гибридные субпопуляции отличаются частотами гомозигот и поддерживаются уменьшением и изменением типа миграционного потока на границе локальных популяций (Щипанов и др., 2008а). Субпопуляции также могут оказаться подразделенными. С учетом размеров индивидуальных участков самцов в период размножения, средних и максимальных (1579 м) известных дистанций при повторных отловах бурозубок, можно полагать, что особь способна передавать свои гены на участке радиусом не менее 2000 м от места рождения (Щипанов и др., 2008б). Поэтому можно считать частоту аллеля постоянной на 3–4 км трансекты в пределах гибридной субпопуляции. Рассмотрим подразделенность гибридной популяции на примере нескольких гибридных зон.
Из 12 проанализированных гибридных зон с комплексными гетерозиготами только для четырех имеются данные по всей ширине гибридной зоны (таблица, зоны 4а,б, 7а,б, 9а,б, 12). Еще в трех гибридных зонах исследованы, вероятнее всего, центральные участки (таблица, зоны 5а,б, 8, 10), и два из них крайне узкие, 0.65 и 0.5 км. В остальных пяти гибридных зонах исследованы краевые участки, во всяком случае, в них резко преобладает одна раса.
Ширина гибридной зоны рас Hällefors/Uppsala с гетерозиготами RIV достигает 21 км. Эту гибридную зону разделяет р. Kolbäcksan шириной 50 м (Narain, Fredga, 1996). Такая река не может служить существенной преградой при расселении бурозубок, но на ее восточном берегу преобладают в выборках бурозубки расы Uppsala, а на западном – бурозубки расы Hällefors. Бурозубки расы Uppsala обнаружены на западном берегу не более чем в 2 км, а гетерозиготы встречаются и в 11–15 км от реки. На восточном берегу бурозубки расы Hällefors проникают в популяцию Uppsala меньше чем на 1 км, а гетерозиготы встречаются на удалении и в 6–10 км от реки.
Участки гибридных зон с двумя гомозиготами и гетерозиготами можно выделить как гибридные субпопуляции Hällefors и Uppsala. Их небольшая ширина указывает на отсутствие подразделенности и эффекта Валунда, и в обеих субпопуляциях отмечен небольшой дефицит гетерозигот (таблица, зоны 4а,б). Гетерозиготы проникают дальше гомозигот, вероятно, потому что более сходны по социальным взаимодействиям с обеими расами. Те участки гибридной зоны, где встречается только одна раса и гетерозиготы, также можно выделить в особые субпопуляции, но в них индекс инбридинга не отличается от нуля. Если объединить все выборки этой гибридной зоны, то индекс инбридинга неожиданно увеличится в 5 раз. Столь сильный эффект Валунда связан со значительной шириной и гетерогенностью этой гибридной зоны.
В гибридной зоне Селигер/Москва с максимально сложными гетерозиготами (CXI) глубина проникновения гомозиготных и гетерозиготных особей не отличается, обе расы и гетерозиготы встречаются с 4-го по 8-й км трансекты, причем на 4–5-м км преобладает раса Москва, а на 6–8-м – раса Селигер (Bulatova et al., 2011: табл. 3). Следовательно, область совместного распространения рас этой гибридной зоны даже несколько шире, чем в зоне с более простыми гетерозиготами Hällefors/Uppsala. Последняя расположена в области Скандинавского ледникового щита, поэтому могла сформироваться позже.
Дефицит гетерозигот в гибридной субпопуляции с преобладанием расы Москва (субпопуляция Москва) больше, чем на участке с преобладанием расы Селигер (субпопуляция Селигер) (таблица, зоны 9а,б). Обобщенный показатель индекса инбридинга по всей гибридной зоне близок к субпопуляции Москва, следовательно, эффект Валунда невелик.
Ширина зоны, в которой совместно встречаются хромосомные расы Новосибирск и Томск и сложные гетерозиготы CIX, достигает 7 км. На отрезке трансекты длиной 4 км преобладает по численности раса Новосибирск, а на отрезке 3 км – раса Томск (Polyakov et al., 2011: табл. S2). Индекс инбридинга заметно больше в той части гибридной зоны, где преобладает раса Томск (субпопуляция Томск) (таблица, зоны 7а,б). Обобщенный индекс инбридинга по всей гибридной зоне близок к субпопуляции Томск, следовательно, и в этой гибридной зоне эффект Валунда невелик.
В крайне узкой гибридной зоне рас Drnholec/Białowieża шириной всего 0.65 км имеется небольшая физическая преграда – железнодорожная насыпь (Szałaj et al., 1996). С одной стороны насыпи в популяции преобладает раса Białowieża, а с другой – раса Drnholec. В субпопуляции с преобладанием расы Białowieża дефицит гетерозигот несомненен (таблица, зона 5б). Напротив, в той части гибридной зоны, где преобладает раса Drnholec (субпопуляция Drnholec), дефицита гетерозигот нет, что удивительно при столь сложных комплексных гетерозиготах CX. При объединении в расчетах всех выборок этой гибридной зоны индекс инбридинга резко возрастает на 60% по сравнению с субпопуляцией Białowieża.
Приведенные данные показывают, что для минимизации эффекта Валунда достаточно рассчитать индекс инбридинга отдельно в каждой гибридной субпопуляции. Как в широких, так и в узких гибридных зонах даже небольшие физические преграды могут служить границей субпопуляций. Гибридные зоны не симметричны, субпопуляции отличаются по размерам и дефициту гетерозигот. Хотя структура гибридных зон влияет на расчеты дефицита гетерозигот, но причиной его возникновения может быть только ассортативное скрещивание.
ОЦЕНКА ПРЕЗИГОТИЧЕСКОЙ ИЗОЛЯЦИИ ХРОМОСОМНЫХ РАС
Ширина гибридных зон с простыми гетерозиготами (CIII) достигает десятков и сотен километров, в них не отмечается дефицита гетерозигот (Borisov et al., 2014), следовательно, между такими расами нет и презиготической изоляции.
В сравнении гибридных зон с комплексными гетерозиготами надо исходить из сходства и различия их субпопуляций. Различия субпопуляций отмечены во всех трех гибридных популяциях, для которых имеются данные по всей ширине гибридной зоны (таблица, рис. 3, зоны 4а,б, 7а,б, 9а,б). Природа этих различий еще не изучена, но может объясняться разными возможностями расселения гетерозигот и гомозигот в гибридной популяции и различиями реципрокных скрещиваний. В двух гибридных зонах, Hällefors/Uppsala и Пенза/Нерусса, показаны участки гибридной популяции с одной гомозиготой и гетерозиготами, что может объясняться только более успешным расселением гетерозигот. Вероятнее всего, с этим связаны и значительные различия субпопуляций в гибридной зоне Drnholec/Białowieża (рис. 3, зона 5).
Рис. 3.
Коэффициенты ассортативного скрещивания (R) в двух группах гибридных популяций, отличающихся типами гетерозигот: 1–4 с гетерозиготами RIV, CIV, CV; 5–12 c гетерозиготами RVI, CIX–CXI. Линиями соединены значения R в субпопуляциях одной гибридной зоны. Номера гибридных зон соответствуют таблице.

С учетом высказанных выше соображений о влиянии подразделенности гибридной зоны на величину дефицита гетерозигот можно оценить ассортативное скрещивание различных хромосомных рас. Из данных таблицы заметна связь между величиной ассортативного скрещивания (R) и усложнением комплексных гетерозигот. В четырех гибридных зонах с более простыми гетерозиготами CIV, CV и RIV индекс R изменяется в пределах 0.23–0.59 (таблица, рис. 3, зоны 1–4), а в остальных гибридных зонах с более сложными гетерозиготами (RVI, СIХ–СХI) – превышает 0.82.
При ассортативном скрещивании сокращение доли гетерозигот (2Vq) происходит за счет гибридов первого поколения, F1 (рис. 2), что должно вызывать значительную презиготическую изоляцию контактирующих хромосомных рас. Сокращение доли F1 (2Vq) при ассортативном скрещивании наглядно показано на рис. 4 (таблица, зоны 8, 12). В гибридных зонах хромосомных рас Западная Двина/Москва и Западная Двина/Селигер с индексами ассортативного скрещивания (R) 0.85 и 0.95 соответственно (таблица, зоны 8, 11) поток генов не прерывается (Григорьева и др., 2015), но достоверно сокращается, судя по морфометрическим различиям челюстного аппарата контактирующих рас (Орлов и др., 2013б).
Рис. 4.
Частоты генотипов в двух гибридных популяциях обыкновенной бурозубки: Западная Двина (Wd)/Москва (Мо) (таблица, зона 8) и S. antinorii/Vaud (таблица, зона 12). Черные столбики – найденные частоты, белые столбики – ожидаемые частоты в следующем поколении при условии случайного скрещивания хромосомных рас и их коррекция ассортативным скрещиванием (Vq).
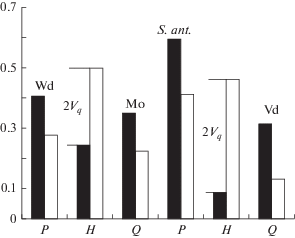
Полная генетическая изоляция хромосомной расы Vaud и S. antinorii достигается при таких же гетерозиготах (CXI, CVII), как и в других генетически не изолированных хромосомных расах (Yannic et al., 2009). Рассчитанная нами доля ассортативных скрещиваний 0.995 оказывается достаточной для прекращения потока генов между хромосомной расой Vaud и S. antinorii (таблица, зона 12). В этой гибридной зоне известным максимальным значениям ассортативности скрещиваний соответствует и максимальный дефицит гетерозигот, показателем чего является высокий FIS. Таким образом, нам впервые удалось показать, что поток генов в гибридной зоне прерывает именно ассортативное скрещивание, а не какой-то особый тип генетической изоляции (Baker, Bradley, 2006), отличающийся от репродуктивной изоляции.
ВОЗМОЖНА ЛИ СТАБИЛИЗАЦИЯ ГИБРИДНЫХ ЗОН БЕЗ АССОРТАТИВНОГО СКРЕЩИВАНИЯ?
В качестве одного из объяснений феномена узких гибридных зон c дефицитом гетерозигот часто упоминают гипотезу напряженной гибридной зоны (Barton, Hewitt, 1985), согласно которой гибридная зона может оставаться стабильной сколь угодно долгое время в результате баланса между пониженной плодовитостью гибридов и миграцией особей из ареалов исходных форм.
В настоящее время приходится сомневаться в том, что формирование гибридной популяции происходит без влияния отбора. Хорошо известно, что в классической модели с постоянной приспособленностью двухаллельных генотипов их устойчивое равновесие возможно только при повышенной приспособленности гетерозигот и невозможно – при пониженной (на коротких временных интервалах без вмешательства отбора) (Hedrick, 1983). Поэтому и в открытых гибридных популяциях в природе миграция исходных форм может поддерживать только нестабильное равновесие, тем более при известных резких колебаниях численности популяций в природе. При пониженной плодовитости гетерозигот и случайности скрещиваний уменьшается не только доля Н (H0 < HЕ), но и доля Q (Q0 < QЕ). Поэтому без вмешательства отбора, ассортативного скрещивания, неизбежно исчезновение одной из контактирующих форм. Похожие случаи описывают как поглощение в процессе гибридизации. Напротив, во всех исследованных гибридных зонах обыкновенной бурозубки с заметным дефицитом гетерозигот найденные доли Р и Q всегда больше ожидаемых при условии случайности скрещиваний (таблица). Поэтому с полной уверенностью можно утверждать, что стабильность генотипов в гибридных популяциях (в том числе стабильный дефицит гетерозигот) и сохранение в гибридной зоне обеих рас невозможно объяснить исключительно пониженной приспособленностью гетерозигот в сочетании с миграцией гомозигот в гибридную зону. Именно ассортативное скрещивание может быть фактором, способным не только вызвать дефицит гетерозигот, но в то же время увеличивать и сохранять в гибридной популяции обе доли гомозигот. В гипотезе напряженной гибридной зоны не учитывались хорошо известное в настоящее время усиление презиготической изоляции – reinforcement – и прекращение гибридизации на заключительном этапе видообразования.
ЗАКЛЮЧЕНИЕ
В анализе гибридных популяций мы исходили из того, что ассортативное скрещивание формируется отбором, направленным против гетерозигот с пониженной приспособленностью. Дефицит гетерозигот стабилизируется в соответствии с достигнутым уровнем ассортативного скрещивания, поэтому оба показателя (R и FIS) связаны высоким уровнем корреляции. Мы показали, что для минимизации эффекта Валунда при сравнении гибридных зон с комплексными гетерозиготами необходимо исходить из сходства и различия их субпопуляций.
Нам впервые удалось показать связь между величиной ассортативного скрещивания и усложнением комплексных гетерозигот, а также то, что поток генов в гибридной зоне прерывает именно ассортативное скрещивание. Ассортативное скрещивание не только уменьшает поток генов, но и поддерживает стабильность генотипов в гибридных популяциях.
Многие вопросы использования ассортативного скрещивания для анализа reinforcement в гибридных зонах остаются неизученными. Обыкновенная бурозубка может стать удобным объектом для исследования reinforcement с использованием количественной оценки ассортативности скрещиваний в десятках зон контакта хромосомных рас, находящихся на разных этапах процесса видообразования.
Список литературы
Григорьева О.О., Борисов Ю.М., Стахеев В.В. и др. Генетическая структура популяций обыкновенной бурозубки Sorex araneus L. 1758 (Mammalia, Lipotyphla) на сплошных и фрагментированных участках ареала // Генетика. 2015. Т. 51. № 6. С. 711–723. (Grigoryeva O.O., Borisov Yu.M., Stakheev V.V. et al. Genetic structure of the common shrew Sorex araneus L. 1758 (Mammalia, Lipotyphla) in continuous and fragmented areas // Rus. J. Genet. 2015. V. 51 (6). P. 607–618.)
Орлов В.Н., Борисов Ю.М., Черепанова Е.В. и др. Узкая гибридная зона хромосомных рас Москва и Западная Двина и особенности изоляции популяций обыкновенной бурозубки Sorex araneus (Mammalia) // Генетика. 2012. Т. 48. № 1. С. 80–88.
Орлов В.Н., Борисов Ю.М., Черепанова Е.В., Милишников А.Н. Аcсортативное скрещивание в гибридных зонах хромосомной расы Западная Двина обыкновенной бурозубки Sorex araneus (Mammalia) // Докл. РАН. 2013а. Т. 451. № 1. С. 110–113.
Орлов В.Н., Сычева В.Б., Черепанова Е.В., Борисов Ю.М. Краниометрические различия контактирующих хромосомных рас обыкновенной бурозубки Sorex araneus (Mammalia) как следствие их ограниченной гибридизации // Генетика. 2013б. Т. 49. № 4. С. 479–490.
Черепанова Е.В., Кривоногов Д.М., Щегольков А.В. и др. Формирование зон контакта хромосомных рас обыкновенной бурозубки (Sorex araneus, Soricomorpha) в бассейне Волги // Зоол. журн. 2018. Т. 97. № 4. С. 723–734.
Щипанов Н.А., Павлова С.В. Многоуровневая подразделенность в группе видов “araneus” рода Sorex. 1. Хромосомная дифференциация // Зоол. журн. 2016а. Т. 95. № 2. С. 216–233.
Щипанов Н.А., Павлова С.В. Многоуровневая подразделенность в группе видов “araneus” рода Sorex. 2. Иерархическая структура вида S. araneus // Зоол. журн. 2016б. Т. 95. № 3. С. 353–365.
Щипанов Н.А., Булатова Н.Ш., Павлова С.В. Распределение обыкновенных бурозубок (Sorex araneus L.) двух хромосомных рас в зоне интерградации. Может ли изменение типа расселения поддерживать независимость генных частот? // Генетика. 2008а. Т. 44. № 6. С. 734–745.
Щипанов Н.А., Купцов А.В., Демидова Т.Б. и др. Нерезидентность и расселение у обыкновенных бурозубок (Sorex araneus, Insectivora) // Зоол. журн. 2008б. Т. 87. № 3. С. 331–343.
Щипанов Н.А., Булатова Н.Ш., Павлова С.В., Щипанов А.Н. Обыкновенная бурозубка (Sorex araneus) – модельный вид эколого-эволюционных исследований // Зоол. журн. 2009. Т. 88. № 8. С. 975–989.
Baker R.J., Bradley R.D. Speciation in mammals and the genetic species concept // J. Mammal. 2006. V. 87 (4). P. 643–662.
Barton N.H., Hewitt G.M. Analysis of hybrid zones // Ann. Rev. Ecol. System. 1985. V. 16. P. 113–148.
Bearhop S., Fiedler W., Furness R.W. et al. Assortative mating as a mechanism for rapid evolution of a migratory divide // Science. 2005. V. 310. P. 501–504. https://doi.org/10.1126/science.1115661
Beysard M., Krebs-Wheaton R.K., Heckel G. Tracing reinforcement through asymmetrical partner preference in the European common vole Microtus arvalis // BMC Evol. Biol. 2015. V. 15. P. 170–181.
Bímová B.V., Macholán M., Baird S.J.E. et al. Reinforcement selection acting on the European house mouse hybrid zone // Mol. Ecol. 2011. V. 20. P. 2403–2424.
Borisov Yu.M., Kriscshuk I.A., Cherepanova E.V. et al. Chromosomal polymorphism of common shrew, Sorex araneus L., in Belarus // Acta Theriol. 2014. V. 59. № 2. P. 243–249.
Borodin P.M. Chromosome and speciation // Biosphere Origin and Evolution / Ed. N. Dobretsov, N. Kolchanov, A. Rozanov. N.Y.: Springer, 2008. P. 315–325.
Brünner H., Hausser J. Genetic and karyotypic structure of a hybrid zone between the chromosomal races Cordon and Valais in the common shrew, Sorex araneus // Hereditas. 1996. V. 125. P. 147–158.
Brünner H., Lugon-Moulin N., Hausser J. Alps, genes, and chromosomes: their role in the formation of species in the Sorex group (Mammalia, Insectivora), as inferred from two hybrid zones // Cytogenet. Gen. Res. 2002. V. 96. P. 85–96.
Bulatova N., Jones R.M., White T.A. et al. Natural hybridization between extremely divergent chromosomal races of the common shrew (Sorex araneus, Soricidae, Soricomorpha): hybrid zone in European Russia // J. Evol. Biol. 2011. V. 24. № 3. P. 573–586.
Coyne J.A., Orr H.A. Speciation. Sunderland: Sinauer Associates, Inc., 2004. 480 p.
Crow J.F., Felsenstein J. The effect of assortative mating on the genetic composition of a population // Eug. Quart. 1968. V. 15. № 2. P. 85–97. https://doi.org/10.1080/19485565.1968.9987760
Crow J.F., Kimura M. An introduction to population genetics theory. N.Y.: Harpers and Row, 1970. 591 p.
Dehnel A. The biology of breeding of common shrew S. araneus L. in laboratory conditions // Ann. Univ. Curie-Skłodovska. 1952. V. 6. P. 359–376.
Fredga K. Reconstruction of the postglacial colonization of Sorex araneus into northern Scandinavia based on karyotype studies, and the subdivision of the Abisko race into three // Russ. J. Theriol. 2007. V. 6. № 1. P. 85–96.
Fredga K., Narain Y. The complex hybrid zone between the Abisko and Sidensjö chromosome races of Sorex araneus in Sweden // Biol. J. Linn. Soc. 2000. V. 70. P. 285–307.
Hedrick P.W. Genetics of populations. Boston: Science Books International, 1983. 629 p.
Jadwiszczak K.A., Banaszek A. Fertility in the male common shrews, Sorex araneus, from the extremely narrow hybrid zone between chromosome races // Mamm. Biol. 2006. V. 71. 5. P. 257–267.
Jiggins C.D., Mallet J. Bimodal hybrid zones and speciation // Tr. Ecol. Evol. 2000. V. 15. P. 250–255.
Jones R.M., Searle J.B. Mapping of course of the Oxford – Hermitage chromosomal hybrid zone in the common shrew Sorex araneus – a GIS approach // Mammalia. 2003. V. 87. № 2. P. 193–200.
Li C.C. First course in population genetics. Pacific Grove: Boxwood Press, 1976. 555 p.
Mayr E. Animal species and evolution. Harvard: Harvard Univ. Press, 1963. 659 p.
Moska M. A hybrid zone between the chromosomal races Łȩgucki Młyn and Popielno of the common shrew in north-eastern Poland: preliminary results // Acta Theriol. 2003. V. 48. P. 441–455.
Narain Y., Fredga K. A hybrid zone between the Hällefors and Uppsala chromosome races of Sorex araneus in central Sweden // Hereditas. 1996. V. 125. P. 137–145.
Narain Y., Fredga K. Spermatogenesis in common shrews, Sorex araneus, from a hybrid zone with extensive Robertsonian polymorphism // Cytogen. Cell Genet. 1998. V. 80. P. 158–164.
Noor M.A. Speciation driven by natural selection in Drosophila // Nature. 1995. V. 375. P. 674–675.
Pavlova S.V., Shchipanov N.A. A hybrid zone between the Kirillov and Petchora chromosomal races of the common shrew (Sorex araneus L., 1758) in northeastern European Russia: a preliminary description // Acta Theriol. 2014. V. 59. P. 415–426.
Peakall R., Smouse P.E. GenAIEx V6: genetic analysis in Excel. Population genetic software for teaching and research // Mol. Ecol. Notes. 2006. V. 6. № 1. P. 288–295.
Polyakov A., White Th., Jones R. et al. Natural hybridization between extremely divergent chromosomal races of the common shrew (Sorex araneus, Soricidae, Soricomorpha): hybrid zone in Siberia // J. Evol. Biol. 2011. V. 24. P. 1393–1402.
Shurtliff Q.R. Mammalian hybrid zones: a review // Mammal. Rev. 2013. V. 43. P. 1–29.
Szałaj K.A., Fedyk S., Banaszek A. et al. A hybrid zone between two chromosome races of the common shrew, Sorex araneus, in eastern Poland. Preliminary results // Hereditas. 1996. V. 125. P. 169–176.
Weir B.S., Cockerham C.C. Estimating F-statistics for the analysis of population structure // Evolution. 1984. V. 38. P. 1358–1370.
Wόjcik J.M., Ratkiewicz M., Searle J.B. Evolution of the common shrew Sorex araneus: chromosomal and molecular aspects // Acta Theriol. 2002. V. 47. Suppl. 1. P. 139–167.
Yannic G., Basset P., Hausser J. Chromosomal rearrangements and gene flow over time in an inter-specific hybrid zone of the Sorex araneus group // Heredity. 2009. V. 93. P. 1–10.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии