

Успехи современной биологии, 2019, T. 139, № 5, стр. 516-520
Вариабельность наполнения и изменения микроструктуры стенки медового зобика пчелы Apis mellifera L.
Е. К. Еськов 1, *, М. Д. Еськова 1, А. С. Роженков 1
1 Российский государственный аграрный заочный университет
Балашиха, Московская обл., Россия
* E-mail: ekeskov@yandex.ru
Поступила в редакцию 09.04.2019
После доработки 02.05.2019
Принята к публикации 07.05.2019
Аннотация
Установлена зависимость наполнения зобиков от физиологического состояния пчел и качества углеводного корма. Полупроницаемостью стенок зобика обеспечивается регуляция содержания воды в углеводном корме. Стенку зобика формирует мышечный слой, под которым располагается слизистая оболочка. Образующие ее продолговатые клетки по мере наполнения зобика вытягиваются вдоль мышечного слоя. Отравления пчел свинцом или кадмием порождают необратимые дегенеративно-дистрофические и некробиотические изменения стенки зобика.
ВВЕДЕНИЕ
Медовый зобик медоносной пчелы представляет собой эластичный тонкостенный резервуар, используемый для транспортировки и хранения индивидуальных запасов углеводного корма или воды (Snodgrass, 1956). От емкости зобика зависит эффективность использования пчелой кормовых ресурсов и обеспечения пчелиной семьи углеводным кормом. В зобике формируется углеводно-белковый трофический субстрат, используемый для кормления личинок.
В годичном цикле жизни пчелиной семьи рабочие пчелы потребляют углеводный корм, различающийся по содержанию воды, эссенциальных и токсичных элементов. С возрастающим техногенным загрязнением природной среды связано загрязнение кормовых участков пчел и потребляемого пчелами углеводного (нектара и меда) и белкового корма (преимущественно цветочной пыльцы) тяжелыми металлами. Накопление в корме токсикантов может выступать в качестве одной из причин массовой гибели пчелиных семей, наблюдаемой в последние годы (Ильясов и др., 2017; Еськов, Еськова, 2018а,б; Dainat et al., 2012).
Настоящей работой предпринято изучение изменчивости наполнения и микроструктуры медового зобика. Изучали также изменение микроструктуры стенок зобиков при отравлениях свинцом или кадмием, что актуально для выявления причин гибели пчел при потреблении загрязненного корма.
МАТЕРИАЛЫ И МЕТОДЫ
Исследование выполнено на рабочих пчелах Apis mellifera L., живших в своих семьях или содержавшихся по 400–500 особей в энтомологических садках. Пчелы, содержавшиеся в садках, потребляли чистые растворы сахарозы в дистиллированной воде или с солями токсикантов, в качестве которых использовали трехводный уксусно-кислый свинец или двухводный ацетат кадмия. На пчелах, живших в своих семьях, изучали сезонную и возрастную изменчивость наполнения зобиков растворами углеводов и водой.
Зобики взвешивали с точностью 0.5 мг на электронных весах AF 224RCE (Shinko Denshi, Япония). Для определения воды в субстратах, наполнявших зобики, их высушивали до постоянной массы при 102°С в терморегулируемом шкафу СНОЛ (Россия). Гипертермии пчел подвергали в суховоздушном термостате ТС-80 (Россия).
Изменения микроструктуры стенок зобиков под влиянием отравления свинцом и кадмием определяли гистологическим методом. Для этого ампутированные зобики пчел, потреблявших корм, загрязненный солями свинца или кадмия, а также чистые растворы сахарозы, фиксировали в 10%-ном нейтральном формалине. После промывания в дистиллированной воде зобики пропитывали парафином в рабочей камере типа АТ-4м. В качестве обезвоживающей среды использовался изопропиловый спирт. Для получения гистологических срезов использовали роторный микротом. Срезы (их толщина составляла около 4 мкм) помещали на предметное стекло и окрашивали гематоксилин-эозином (Меркулов, 1961). Для визуального анализа и фотографирования препаратов использовали светооптический микроскоп.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Вариабельность наполнения зобика
У пчел, завершающих постэмбриональное развитие, масса зобиков варьирует в широких пределах – от 0.7 до 61 мг, что зависит от количества содержащейся в них ювенильной жидкости. Она расходуется в начале имагинальной стадии. Некоторая часть пчел, достигнув стадии имаго, потребляет эту жидкость, находясь в расплодных ячейках сот. Этим обусловливается высокая вариабельность наполнения зобиков у пчел, выходящих из ячеек.
После выхода из ячеек ювенильная жидкость в зобиках расходуется за 22 ± 10 ч. У пчел, не имеющих возможности потребления углеводного корма и погибающих от голода, масса зобиков уменьшается в среднем до 0.63 мг (мин. – 0.49, макс. – 0.85 мг). Подобно этому у пчел среднего и старшего возрастов, погибающих от голода, масса зобиков составляет в среднем 0.68 мг (мин. – 0.52, макс. – 0.85 мг). Очевидно, в указанных пределах варьирует масса пустых зобиков рабочих пчел.
В начале имагинальной стадии пчелы, имеющие неограниченный доступ к углеводному корму, слабо наполняют им зобики. Через 5–12 ч после выхода из ячеек масса зобиков составляет в среднем 2.4 ± 1.6 мг. Максимальное их наполнение примерно у 5% пчел достигает 10–15 мг. К 2-суточному возрасту наполнение зобиков достигает в среднем 5.3 ± 3.9 мг, а в дальнейшем существенно варьирует в зависимости от экологической ситуации и физиологического состояния пчел.
В течение весенне-летнего сезона зобики пчел, находящихся в гнезде, имеют относительно небольшое наполнение, зависящее от локализации на сотах. Обычно по наполнению зобиков различаются пчелы, находящиеся на расплодных и кормовых ячейках сот. При низкой продуктивности кормового участка в конце весны–начале лета у пчел, находящихся на расплодных сотах, масса зобиков составляет в среднем 1.6 мг (макс. – 7 мг), возрастая к началу осени до 5.1 (макс. – 32 мг), а на кормовых – 2.7 (макс. – 19) и 23.6 мг (макс. – 55 мг) соответственно. В периоды, характеризующиеся высокой продуктивностью кормового участка, наполнение зобиков пчел, независимо от локализации в гнезде, возрастает.
У пчел, вылетающих из улья, наполнение зобиков зависит от мотивации вылетов. С большими запасами корма покидают гнездо пчелы переселяющихся пчелиных семей. Эти пчелы, вылетая из улья, набирают в зобики в среднем по 20.2 мг (мин. – 9, макс. – 48 мг) углеводного корма. Значительное наполнение зобиков углеводным кормом стимулирует дым. Под его воздействием перед вылетом из улья пчела может набирать в зобик до 88 мг меда.
Относительно небольшое количество корма содержат зобики пчел, впервые вылетающих из гнезда после завершения зимовки. Эти пчелы, вылетая на очистительный облет, набирают в зобики в среднем по 1.4 мг (макс. – 6 мг) корма. Подобно этому, небольшие его запасы содержат зобики пчел, вылетающих к источникам корма или воды, находящимся на небольшом расстоянии от улья. Но вылетая к известным источникам корма, находящимся на большом расстоянии, пчелы увеличивают наполнение зобиков. Их нагрузка возрастает также при понижении продуктивности кормового участка. Например, в то время, когда суточная доставка корма трехкратно превышала его суточное потребление семьей, масса зобиков у большинства вылетающих пчел не превышала 2 мг (макс. – 16 мг). При уменьшении продуктивности кормового участка до суточной потребности семьи у пчел-фуражиров, вылетающих из улья, наполнение зобиков возрастало в среднем до 5.3 мг (макс. – 31 мг).
От продуктивности кормового участка зависит наполнение зобиков пчел-фуражиров, возвращающихся в улей. У пчел, посещающих цветки, обильно выделяющие нектар или кормушки с растворами углеводов, находящихся на небольшом расстоянии от улья, максимальная масса зобиков достигает 70–74 мг. У пчел-водоносов она обычно не превышает 58 мг. Это объясняется различиями по удельному весу воды и нектара.
От начала к завершению зимовки наполнение зобиков уменьшается, что связано с наполнением ректумов. У пчел, зимовавших при 3 ± 1°С, в ноябре–декабре масса зобиков составляла в среднем 8.4 мг (макс. – 38 мг), в январе–феврале – 3.3 (макс. – 37 мг) и в марте – 2.4 мг (макс. – 29 мг).
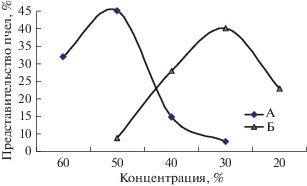
У пчел, содержащихся в энтомологических садках (изолированных от семьи), масса зобиков зависит от качества углеводного корма. С понижением концентрации раствора сахарозы или другого углевода, потребляемого пчелами, масса зобиков уменьшается. У разновозрастных пчел через 3 ± 1 сут жизни в садках при потреблении меда масса зобиков устанавливается в среднем на уровне 27.4 мг. Она существенно различалась у пчел, потреблявших 60%-ную сахарозу, составляя в среднем 23.3 мг и уменьшалась до 7.5 мг при потреблении 20%-ной сахарозы (рис. 1).
Рис. 1.
Наполнение зобиков в зависимости от концентрации раствора сахарозы: А – 20%-ная, Б – 60%-ная.

Зобики используются пчелами для регуляции концентрации содержащегося в них углеводного корма. Низкоконцентрированные растворы углеводов в зобиках обезвоживаются, а высококонцентрированные – разбавляются (рис. 2). Поэтому у разновозрастных пчел, содержавшихся в садках и потреблявших 20%-ную сахарозу, через 10 сут ее концентрация в зобиках возрастала в среднем до 28.4% (максимум до 46%). В противоположность этому при потреблении 60%-ной сахарозы ее концентрация в зобиках уменьшалась в среднем до 54%, варьируя у разных пчел от 34 до 58%.
Рис. 2.
Распределение пчел по концентрациям содержимого зобиков (ось абсцисс) при потреблении 60%-го (А) или 20%-го (Б) растворов сахарозы в течение 10 сут жизни в энтомологических садках.
Гипертермия интенсифицирует изменение концентрации сахарозы в зобиках пчел. При потреблении 20%-ной сахарозы под влиянием содержания пчел в течение 5 ч при 40°С ее концентрация в зобиках повышалась в 1.9 раза (Р ≥ 0.99). В случае потребления 60%-ной сахарозы концентрация ее раствора в зобиках возрастала в 1.2 раза (Р ≥ 0.95). При гипертермии вода в содержимом зобиков расходуется на охлаждение пчел, что способствует повышению их выживания и соответственно понижению концентрации углеводного корма (Еськов, Бабкина, 1990).
Изменчивость микроструктуры стенки зобика
Наружную часть стенки зобика образует мышечный слой, под которым располагается слизистая оболочка, представленная однослойным эпителием. Ее образуют клетки вытянутой формы, со светлой цитоплазмой и овальными базофильными ядрами. Эти клетки имеют неровную мембрану со множеством коротких отростков (микроворсинок). Они обеспечивают увеличение поверхности клеточных мембран. На границе мышечного слоя и слизи выявляются единичные трахейные трубочки и тонкие соединительнотканные перемычки.
Локализация клеток слизистой оболочки зависит от наполнения зобика. У пустого или слабо заполненного зобика клетки располагаются почти под прямым углом к мышечному слою. По мере наполнения зобика происходит распрямление складок и увеличение наклона клеток (рис. 3). У наполненного зобика клетки слизистой оболочки растягиваются вдоль мышечного слоя.
Рис. 3.
Фрагменты стенок медового зобика, изменяющихся при его наполнении ((а) – пустой, (б), (в) – фазы наполнения): 1 – мышечный слой, 2 – эпителиальная клетка, 3 – ядро эпителиальной клетки, 4 – ворсинки, 5 – светлая цитоплазма.

Отравление пчелы свинцом порождает дегенеративно-дистрофические и некробиотические изменения стенки зобика. Этому сопутствует сморщивание отдельных групп эпителиальных клеток и частичная утрата ими межклеточных соединений. В мышечных волокнах образуются очаги зернисто-глыбчатого распада, а местами исчезает поперечно-полосатая исчерченность (рис. 4).
Рис. 4.
Микроструктура медового зобика пчелы, отравленной свинцом: 1 – мышечный слой с признаками дегенерации, выражающейся в исчезновении поперечно-полосатой исчерченности и некробиотических изменениях; 2 – цитоплазма сморщенных клеток.

Сходные изменения микроструктуры стенки зобика происходят при отравлении пчел солями кадмия. В мышечном слое дегенеративно-дистрофические и некробиотические изменения выражаются в возникновении очагов зернисто-глыбчатого распада. В цитоплазме образуются эозинофильные включения, и происходит сморщивание эпителиальных клеток. При этом ядра приобретают признаки гиперхромотоза и пикноза.
ЗАКЛЮЧЕНИЕ
Наполнение зобика углеводным кормом варьирует в широких пределах, изменяясь в зависимости от возраста, физиологического состояния пчелы и качества потребляемого трофического субстрата. При одинаковом наполнении масса зобиков зависит от удельной массы наполняющих субстратов. Поэтому с увеличением концентрации углеводного корма масса зобика возрастает. Но наполнение зобиков лимитируется объемом брюшного отдела и находится в обратной зависимости от наполнения ректумов.
Зобик пчелы наряду с функцией хранения и транспортировки корма участвует в регуляции содержания воды в углеводном корме (Еськов, 2018). Вероятно, это достигается полупроницаемостью стенок зобика. Нектар и низкоконцентрированные растворы углеводов обезвоживаются, что необходимо для заготовки меда. Но в годичном цикле жизни взрослые пчелы не потребляют мед в чистом виде. В весенне-летний период жизни пчелы растворяют мед водой, доставляемой в гнездо пчелами-водоносами. В течение зимовки для растворения меда в зобиках используется эндогенная вода (Еськов, 1995).
Специфические изменения микроструктуры стенок зобика происходят при его наполнении, что выражается в изменении ориентации клеток, образующих слизистую оболочку. Они вытягиваются вдоль мышечного слоя при растяжении стенки зобика в процессе его наполнения.
Отравления пчел свинцом или кадмием порождают необратимые дегенеративно-дистрофические и некробиотические изменения стенки зобика. Эпителиальные клетки сморщиваются, а в мышечном слое возникают очаги зернисто-глыбчатого распада. Эти изменения микроструктуры стенок зобика с высокой вероятностью позволяют диагностировать причину гибели пчел, содержащихся в техногенно-загрязненных зонах.
Список литературы
Еськов Е.К. Экология медоносной пчелы. Рязань: Русское слово, 1995. 396 с.
Еськов Е.К. Экзо- и эндогенная вода в теле медоносной пчелы (Apis mellifera, Hymenoptera, Apidae) // Зоол. журн. 2018. Т. 97. С. 151–160.
Еськов Е.К., Бабкина Н.Г. Возрастная и сезонная изменчивость индивидуальной устойчивости медоносной пчелы Apis mellifera L. к экстремальным температурам // Энтомол. обозрение. 1990. Т. 69. № 1. С. 481–485.
Еськов Е.К., Еськова М.Д. Этолого-физиологическое реагирование пчел Apis mellifera L. на загрязнение свинцом углеводного корма // Изв. РАН. Серия биол. 2018а. № 3. С. 322–325.
Еськов Е.К., Еськова М.Д. Миграционное поведение и слеты пчел // Пчеловодство. 2018б. № 7. С. 20–23.
Ильясов Р.А., Поскряков А.В., Николенко А.Г. Семь причин смертности семей пчелы Apis mellifera в России // Пчеловодство. 2017. № 9. С. 10–14.
Меркулов Г.А. Курс патологогистологической техники. Четвертое изд. Л.: МЕДГИ3, 1961. 343 с.
Dainat B., Vanengelsdorp D., Neumann P. Colony collapse disorder in Europe // Environ. Microbiol. Rep. 2012. V. 4. P. 123–125.
Snodgrass R. E. The anatomy of the honey-bee. N.Y.: Univ. Press, 1956. 334 p.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии