

Успехи современной биологии, 2021, T. 141, № 2, стр. 149-163
Современные представления о токсинах высших грибов: высокомолекулярные белки
А. А. Ховпачев 1, *, В. А. Башарин 1, С. В. Чепур 2, **, С. В. Волобуев 3, ***, М. А. Юдин 2, А. С. Гоголевский 2, А. С. Никифоров 2, Л. Б. Калинина 3, М. А. Тюнин 2
1 Военно-медицинская академия им. С.М. Кирова
Санкт-Петербург, Россия
2 Государственный научно-исследовательский испытательный институт военной медицины
Санкт-Петербург, Россия
3 Ботанический институт им. В.Л. Комарова РАН
Санкт-Петербург, Россия
* E-mail: vtmz@vmeda.org
** E-mail: gniiivm_2@mil.ru
*** E-mail: rbo.fungi@binran.ru
Поступила в редакцию 27.07.2020
После доработки 09.08.2020
Принята к публикации 10.08.2020
Аннотация
В продолжение обсуждения токсинов высших грибов рассмотрены их ядовитые высокомолекулярные белки. Токсины подобного строения актуальны для исследований как соединения, способные приводить к интоксикациям у человека. Их используют как инструменты в биотехнологии, а также изучают в качестве перспективных лекарственных средств. Однако, с точки зрения токсинологии, ядовитые белки макромицетов охарактеризованы в недостаточной степени. Согласно общим представлениям о токсинах, рассмотрены их функции и основные принципы строения. Проанализированы механизмы действия, реализуемые перфоринами, аминооксидазами, рибонуклеазами, активаторами вторичных мессенджеров, иммунотоксинами и ядовитыми протеазами макромицетов. Обобщены некоторые данные об их каталитической активности. На примерах эгеролизинов, перфоринов мухоморов, вольватоксина, лектина VVL, болесатина раскрыты современные представления о реализации ими токсического процесса на уровнях от молекулярного до организменного. Сделан вывод о том, что дальнейшее изучение распределения высокомолекулярных белковых токсинов позволит приблизить осмысление их биологической роли в плодовых телах макромицетов, а исследование более детальных механизмов токсического процесса – их систематизировать.
ВВЕДЕНИЕ
Высшие грибы – значимый источник биологически активных высокомолекулярных белков, изучение которых актуально по причине возможного применения в качестве новых медицинских препаратов и мощных исследовательских инструментов (Вассер, 2015). В течение последнего десятилетия их исследования продвинулись за счет попыток упорядочить геномы важных, с точки зрения биотехнологии, фармакологии и медицины, видов макромицетов. Применительно к человеку на клеточном и тканевом уровне эффекты белков рассматривают в рамках антипролиферативного, цитостатического, иммуномодулирующего и тромболитического действия, в отношении других организмов – антибактериального, антипротозойного, нематоцидного, антифунгинального и инсектицидного (Кудрявцева и др., 2008; Белова, 2018; Xu et al., 2011; Hassan et al., 2015; Sabotič et al., 2015; Olombrada et al., 2017; Novak et al., 2020). Исследование механизмов их действия представляет существенный элемент оценки рисков для здоровья человека в ходе фармакологического применения препаратов на их основе (Белова, 2018; Магазов и др., 2019). В то же время часть из них, ввиду выраженности биологического эффекта, подлежит непосредственному изучению в рамках токсинологии.
Высокомолекулярные белки макромицетов могут обусловливать пищевые интоксикации, или мицетизмы, поэтому они значимы еще и с клинической точки зрения (Kretz et al., 1991b; Landi et al., 2017). Мицетизмы подобного рода сопряжены с трудной дифференциальной диагностикой и невысокой частотой лабораторно подтвержденных случаев (White et al., 2019). Это связано как с неустойчивостью белковых токсинов, так и низкой корреляцией между биологической активностью и клиническими проявлениями интоксикаций в условиях их алиментарного попадания в организм. Подобная закономерность характерна для токсического процесса, оказываемого многими пассивно-ядовитыми животными и растениями, у которых отсутствует аппарат доставки токсинов (Guidebook to protein…, 1997; Yao et al., 1998; Berry, Crickmore, 2017). Кроме этого, способность к продукции ядовитых протеинов также встречают и за пределами токсикологически значимых грибов (Odenthal et al., 1982; Žužek et al., 2006). Так, среди культивируемых с пищевыми целями макромицетов встречают белки, эквивалентные по уровню внутривенной токсичности для млекопитающих аманитотоксинам (Hassan et al., 2015).
В целом, белковые токсины высших грибов как регуляторные элементы, способные в малых концентрациях приводить к критическому сдвигу регуляции и гибели биологических систем вне их контроля, часто подпадают под узкое медицинское толкование (Магазов и др., 2019; Nayak et al., 2013). Вместе с тем, в сравнении с подобными соединениями других природных источников, им уделяют гораздо меньшее внимание (Белова, 2018; Landi et al., 2017). Несмотря на довольно длительный период, истекший с начала изучения белковых токсинов макромицетов – первые исследования агглютинирующих и лизирующих эритроциты фракций в составе плодовых тел, причисленных к родам Amanita Pers., Entoloma P. Kumm., Lactarius Pers. и Inocybe (Fr.) Fr., относят к 1907 и 1911 гг. – по настоящий момент нет их единого понимания, а вследствие этого и классификации (Ford, 1911). Это требует от биологов, фармакологов и врачей дальнейшего углубления знаний о них (Вассер, 2015; Wong et al., 2020). В связи с этим в продолжение обсуждения токсинов высших грибов предпринята попытка обобщения современных сведений об их структурной организации, механизмах действия и биологическом разнообразии продуцентов.
ХАРАКТЕРИСТИКА ВЫСОКОМОЛЕКУЛЯРНЫХ БЕЛКОВЫХ ТОКСИНОВ МАКРОМИЦЕТОВ
Функции и общие принципы молекулярной организации
В настоящий момент классификация токсинов белковой структуры макромицетов отсутствует. Тем не менее, представляя лишь часть общего в рамках токсинологии, ядовитые свойства их белков не уникальны в концепции своего развития. В ходе обсуждения это позволяет сравнивать рассматриваемые соединения с аналогичными среди других царств живых организмов, тем самым восполняя пробелы их понимания и осмысления в качестве индивидуальных представителей различных классов токсинов.
С точки зрения фитофармакологии, по соотношениям структура–активность среди белков макромицетов принято выделять лектины, перфорины, грибные иммуномодулирующие белки, ингибиторы синтеза белка и лакказы (Xu et al., 2011). Применительно к токсинологии эти соотношения во многом дублируют представления о бактериальных токсинах (Schmitt et al., 1999), среди которых выделяют по механизмам действия пять основных типов: повреждающие мембрану, ингибиторы синтеза белка, активаторы вторичных мессенджеров, активаторы иммунной системы и протеазы.
Помимо предложенных к рассмотрению, также известно большое количество токсинов, которые способны рецептор-опосредованно влиять на ЦНС млекопитающих, тем самым детерминируя нарушения функций организма или его гибель (Guidebook to protein…, 1997). В данный момент продуценты подобных белков среди макромицетов не известны. Вместе с тем, среди токсинов высших грибов эту нишу заполняют вещества, способные к имитации эндогенных нейромедиаторов, со структурой простых азотсодержащих соединений и ациклических терпенов, которые, по всей видимости, влияют на свойства биомембран нейронов (White et al., 2019).
Касательно биологических функций, на выполнение которых направлены белковые токсины макромицетов, существует несколько гипотез. С одной стороны, они могут действовать как классические защитные соединения в отношении конкурирующих организмов, а с другой – участвовать в регуляции эндогенных процессов, например в реакциях на абиотический стресс или на процессы развития: активный рост, плодоношение и старение (Кудрявцева и др., 2008; Berry, Crickmore, 2017). Кроме того, рассматривают их роль в качестве антиметаболических и запасных белков, а также факторов ауторегуляции (Lapadula, Ayub, 2017). Вместе с тем известны случаи опосредованных макромицетами инфекционных заболеваний, что позволяет предположить роль их токсинов в качестве патогенетических факторов (Mori et al., 1998).
Структурно токсины макромицетов, как и ядовитые белки микроорганизмов, животных и растений, представляют либо отдельные соединения, либо олигомерные комплексы с несколькими видами активности (Hassan et al., 2015). Считают, что эволюционно образование таких агрегатов возникло благодаря объединению двух или более функциональных белков. Это принцип организации двух доменов: один участок обладает каталитической активностью, а другой – отвечает за распознавание и доставку в клетку-мишень (Schmitt et al., 1999).
Тропность белковых токсинов определена их связывающим доменом. Среди токсинов грибов эта функция опосредована лектинными участками, а также аффинностью, обусловленной их молекулярными химическими свойствами (Xu et al., 2011). В качестве лектинов рассматривают неиммунные белки или гликопротеины, которые обладают способностью связывать углеводы с высокой степенью селективности, в результате чего in vitro возникает явление клеточной агглютинации. Следует отметить, что макромицеты представляют один из основных источников лектинов без прочей биологической активности. В течение последних лет был обнаружен ряд грибных лектинов, биологическую активность которых предстоит изучить (Hassan et al., 2015).
К характерным свойствам высокомолекулярных токсинов белковой структуры в качестве сигнальных молекул относят наличие экранирующего элемента, исключающего их случайное взаимодействие с субстратом в собственной клетке и за пределами мишени. Этот механизм также способствует передаче сигнала на большом расстоянии без ослабления его силы (Guidebook to protein…, 1997). Среди токсинов макромицетов известны механизмы протеолитического расщепления субъединицы-прекурсора и инициирования процесса сборки мономеров в полноценный функциональный комплекс только после их взаимодействия со структурными элементами мишеней.
Токсины, повреждающие мембраны
Среди ядовитых белков высших грибов встречают два класса токсинов, повреждающих биологические мембраны: перфорины и прооксиданты.
Перфорины PFTs, или порообразующие токсины (pore-forming toxins), оказывают повреждающее действие на плазматические мембраны посредством циклизации в полимерные комплексы, образующие ионные каналы с различной пропускной способностью. На клеточном уровне образование пор приводит к развитию коллоидно-осмотического лизиса, в то время как на уровне целостного организма они оказывают выраженные кардио-, нейро- и пульмонотоксические действия (Современные методы изучения …, 2020; Gouaux, 1997; Dal Peraro, van der Goot, 2015).
Среди грибов токсичность изолированных перфоринов впервые была показана на примере аспергиллгемолизина оппортунистического патогена Aspergillus fumigatus Fresen., среднесмертельные дозы (ЛД50) которого при внутривенном введении крысам и курицам составляют 0.75 и 0.35 мг/кг соответственно (Henrici, 1939). Начало исследований ядовитых свойств перфоринов среди макромицетов связано с изучением фаллолизинов Мухомора зеленого, или бледной поганки (Amanita phalloides), рубесценслизина М. серо-розового (A. rubescens) и фасцикуларелизина Ложноопенка серно-желтого (Hypholoma fasciculare). Внутривенная токсичность фаллолизинов для мышей составляет ЛД50 = 0.22 мг/кг, а для крыс – ЛД50 = 0.1 мг/кг, ЛД99 = 0.66 мг/кг (Odenthal et al., 1975; Faulstich et al., 1983). Сравнимым уровнем токсичности обладают рубесценслизин, ЛД50 которого при внутривенном введении 0.15 мг/кг (крысы) и 0.31 мг/кг (мыши), и фасцикуларелизин – 0.5 мг/кг (крысы) (Seeger, 1975; Seeger et al., 1981).
На сегодня перфорины идентифицированы, выделены и в различной степени охарактеризованы для ряда макромицетов (табл. 1). Интерес вызывает наличие токсинов подобного рода среди съедобных и культивируемых в пищевых целях видов. Объяснением этого считают относительную термолабильность рассматриваемых перфоринов. Вместе с тем известны эпизодические случаи летальных интоксикаций после приема в пищу в большом количестве или после недостаточной обработки их съедобных продуцентов (Lin et al., 1974; Al-Deen et al., 1987; Jin et al., 2014; White et al., 2019).
Таблица 1.
Перфорины макромицетов (адаптировано по: Nayak et al., 2013)
| Перфорин | Продуцент |
|---|---|
| Эгеролизин | Cyclocybe aegerita (V. Brig.) Vizzini |
| Фаллолизины А, В | Amanita phalloides (Vaill. ex Fr.) Link |
| Рубесценслизин | Amanita rubescens Pers. |
| Фламмутоксины А, В | Flammulina velutipes (Curtis) Singer |
| Фасцикуларелизин | Hypholoma fasciculare (Huds.) P. Kumm. |
| Лектин LSL | Laetiporus sulphureus (Bull.) Murrill |
| Дейтеролизин | Moniliophthora perniciosa (Stahel) Aime et Phillips-Morab |
| MpPRIA 1, 2 | |
| Эрилизины А, B1, B2 | Pleurotus eryngii (DC.) Quél. |
| Эрингеолизин | |
| Небродеолизин | Pleurotus nebrodensis (Inzenga) Quél. |
| Остреолизин А | Pleurotus ostreatus (Jacq.) P. Kumm., Pleurotus pulmonarius (Fr.) Quél. |
| Плевротолизины А, В | |
| Токсин 40 кДа | Entoloma rhodopolium (Fr.) P. Kumm. |
| Шизолизин | Schizophyllum commune Fr. |
| Вольватоксин А2 | Volvariella volvacea (Bull.) Singer |
Токсины высших грибов с порообразующей активностью принадлежат надсемейству мембран-атакующих перфоринов MACPF (membrane attack complex/perforin). Они представляют мономерные протеины, которые образуют каналы при совместном участии и для которых характерна либо одно-, либо двухкомпонентная система полимеризации (Gouaux, 1997; Nayak et al., 2013). Наличие дополнительной субъединицы в ходе реакции полимеризации для некоторых обязательно, а для других – нет. Последние могут реализовывать свой эффект либо потенцируя друг от друга, либо наоборот – ингибируя. Для ряда перфоринов грибов показано, что их максимальная токсичность возникает при фиксированном соотношении субъединиц разных типов (Faulstich et al., 1983; Lukoyanova et al., 2015). Ингибирующие процесс полимеризации субъединицы, как правило, играют роль неактивного предшественника токсина (Weng et al., 2003; Lin et al., 2004).
Структурно среди известных мембран-атакующих перфоринов грибов в наибольшей степени охарактеризованы белки с эгеролизиновым доменом (Pfam: PF06355) (Ota et al., 2013; Frangež et al., 2017). В составе MACPF они входят в семейство холестерин-зависимых цитолизинов CDC (cholesterol-dependent cytolysin), также известных как тиол-активируемые перфорины (Gouaux, 1997; Ota et al., 2013). Несмотря на то, что эталонным в составе эгеролизинов считают ген Аа-PRI1 Опенка тополиного (C. aegerita ≡ Agrocybe aegerita (V. Brig.) Singer), впервые подобная последовательность была секвенирована из мицелия A. fumigatus (Fernandez Espinar, Labarère, 1997; Jin et al., 2014). В настоящий момент к числу мембран-атакующих перфоринов макромицетов, гомологичных эгеролизину (16.0 кДа), относят небродеолизин (27.0 кДа) (Lv et al., 2009), эрилизины (Ery A = 14.9 кДа, Ery B1 = 14.5 кДа, Ery B2 = 37.4 кДа) (Ngai, Ng, 2006), остреолизин А (Oly A = 12.5кДа) и плевротолизины А, В (Ply A = 14.5 кДа, Ply В = 59.0 кДа) (Ota et al., 2013; Lukoyanova et al., 2015), MpPRIA 1, 2 тропического фитопатогена какао Монилиофторы опасной (M. perniciosa) (Mondego et al., 2008).
Сходными с эгеролизинами по массе, по электрохимическим и токсикологическим свойствам являются фаллолизины А, В (Pha А = Pha B = 34.0 кДа), рубесценслизин (35.0 кДа), фасцикуларелизин (35.5 кДа), у которых наличие домена PF06355 не описано. Также, несмотря на то, что N-концевые последовательности эрингеолизина (17.0 кДа), присутствующего наравне с эрилизинами в базидиомах Вешенки степной (P. eryngii), и дейтеролизина, который вместе с MpPRIA 1, 2 кодирует геном M. perniciosa, гомологичны PF06355, их не причисляют к белкам семейства эгеролизинов (Mondego et al., 2008; Nayak et al., 2013).
Эгеролизины представляют двухкомпонентные системы, для последовательностей которых характерны частые повторы цистеина, ароматических и кислых остатков. Индивидуальные белки в составе эгеролизинов стабильны в широком диапазоне рН, однако термолабильны без возможности рефолдинга при температурах более 39°С, их активность подвергнута значительному снижению в присутствии Hg2+ в микромолярном диапазоне концентраций (Žužek et al., 2006).
По всей видимости, функциональное значение аминокислотных повторов в составе эгеролизинов макромицетов отведено реализации аффинитета к клеточным структурам, который наблюдают в пределах семейства бактериальных токсинов с повторяющимися последовательностями RTX (repeats in toxin) (Guidebook to protein…, 1997). Тем не менее, в отличие от грибных эгеролизинов, для RTX-токсинов: гемолизина кишечной палочки и аденилатциклазы коклюшной палочки, а также семейства инсектицидных токсинов Vip3B Bacillus thuringiensis Berliner, – характерны повторы, богатые глицином и аспартатом. Есть мнение, что их аминокислотные последовательности проявляют высокую степень сходства с участками некоторых эквинатоксинов (Ota et al., 2013) и инсектицидных токсинов семейства Cry34/Cry35 B. thuringiensis (Berry, Crickmore, 2017; Novak et al., 2020).
Кроме структурного сходства эгеролизинов с токсинами представителей других царств, определенные параллели присутствуют и в механизмах их токсического действия (Frangež et al., 2017). Так, остреолизин Oly А и плевротолизины Ply А, В, сопутствующие друг другу в плодовых телах Вешенок обыкновенной (P. ostreatus) и легочной (P. pulmonarius), разрабатывают в рамках экологически чистых инсектицидов, но вместе с тем, при внутривенном введении млекопитающим они проявляют выраженные кардиотоксические свойства (Novak et al., 2020).
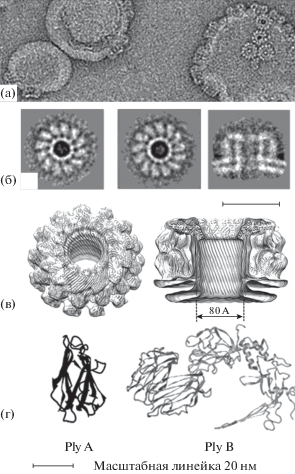
Самопроизвольная олигомеризация в растворе субъединиц типа А Ply и Oly, которые идентичны на 90%, возможна только в присутствии Ply B. К молекулам-мишеням, запускающим процесс сборки, относят специфический для насекомых сфинголипид церамид-фосфоэтаноламин, а также сфингомиелин и холестерин мембран млекопитающих. Методами комбинированной кристаллографии и криоэлектронной микроскопии показано, что сборка 12–13-кратной поры с внутренним диаметром 8 нм происходит посредством присоединения одной молекулы Ply B к димеру Ply A или Oly A, отражая оптимальное соотношение субъединиц 1 : 2 (рис. 1) (Lukoyanova et al., 2015). При данном соотношении их токсичность составляет: для мышей ЛД50 = 1.14 мг/кг внутрибрюшинно, для крыс – ЛД50 = 1.17 мг/кг, ЛД99/15 мин = 1.4 мг/кг внутривенно, в то время как per os достигает ЛД50 = 319 мг/кг (Al-Deen et al., 1987; Žužek et al., 2006; Rebolj et al., 2007).
Рис. 1.
Плевротолизины: поры на липосомах и структуры мономеров (по: Lukoyanova et al., 2015). (а) – криоэлектронная микрофотография пор на липидных монослоях; (б) – усредненные 12- и 13-мерные поры и их сагиттальный вид; (в) – трехмерная реконструкция поры; (г) – трехмерные структуры плевротолизинов А и В.
Несмотря на высокую внутривенную токсичность перфоринов в составе базидиом, экстраполированная пероральная ЛД50 свежих плодовых тел P. ostreatus для человека массой 70 кг составляет около 2.8 кг. При исследовании подострой per os токсичности P. ostreatus у грызунов наблюдают микроабсцедирование печени, а также других паренхиматозных органов, которое сопровождает инфильтрация воспалительными клеточными элементами. Отмечают амилоидные изменения селезенки, лимфоидную инфильтрацию кишки и фокальные геморрагические включения в легких (Al-Deen et al., 1987).
При парентеральном введении млекопитающим плевротолизины приводят к нарушению координации движений, клонико-тоническим судорогам, далее переходящим в атоническую кому. Подобные наблюдения были сделаны уже в ходе ранних исследований фаллолизина, рубесценслизина и фасцикуларелизина, внесение которых в среду препарата Лангендорфа угнетает сократимость миокарда, в дальнейшем приводя к его остановке в фазе систолы (Lin et al., 1974; Odenthal et al., 1975, 1982; Seeger, Bunsen, 1980). В совокупности с электрокардиографическими данными, эти признаки позволили предположить, что гибель вследствие токсического действия перфоринов наступает вследствие сердечного механизма танатогенеза (Odenthal et al., 1975).
Эгеролизины P. ostreatus дозозависимо приводят к гибели эритроцитов, моноцитов, лимфоцитов, тромбоцитов, эндотелия и нейронов человека. Вызываемый токсинами гемолиз сопровождает длительная лаг-фаза, после окончания которой скорость разрушения эритроцитов быстро нарастает, характеризуясь крутой кривой концентрация–отклик (Žužek et al., 2006). Клеточные и субклеточные эффекты оптимальных соотношений перфоринов P. ostreatus возникают in vitro при следующих концентрациях: 7–14 нмоль/л – порог клеточной утечки Ca2+, 64 нмоль/л – IC50 (концентрация полумаксимального ингибирования) гемолиза эритроцитов, 81.2 нмоль/л – IC50 апоптоза фибробластов, 137–250 нмоль/л – IC50 апоптоза эндотелиоцитов пуповины, 450 нмоль/л – IC50 дегрануляции нейтрофилов, при 700 нмоль/л наблюдают блеббинг нейронов (Rebolj et al., 2007; Juntes et al., 2009).
Чтобы пережить инициируемый утечкой ионов стресс, пораженные клетки-мишени активируют ряд защитных мер, в том числе провоспалительный каскад (Frangež et al., 2017). Снижение концентрации клеточного калия вызывает активацию стресс- и митоген-активируемых протеинкиназ MAPK (mitogen-activated protein kinase): р38 и внеклеточных сигнально-регулируемых киназ ERK (extracellular signal-regulated kinases). Активация MAPK приводит к высвобождению запасов кальция из эндоплазматического ретикулума, который в дальнейшем также подвержен выходу из клеток через образуемые токсинами поры. Цепь событий воспалительного комплекса включает мобилизацию каспазы 1 и активацию ИЛ-1 (Dal Peraro, van der Goot, 2015); выясняется роль дополнительных путей воспалительного ответа (Bonfini, Buchon, 2016).
Способность к восстановлению целостности плазматической мембраны после нанесенного повреждения и скорость, с которой это происходит, варьируют между типами клеток. Репарация мембран в условиях действия перфоринов может быть опосредована либо эндоцитозом пораженных участков мембраны, либо наоборот, их отпочковыванием по механизму генерации мультивезикулярных тел по пути ESCRT (endosomal sorting complexes required for transport) (Rebolj et al., 2007). Считают, что ESCRT-опосредованная цитоплазматическая экструзия выступает в данных условиях одним из основных механизмов клеточной защиты, который не требует дополнительного синтеза белка, тем самым позволяя своевременно изолировать вызванные перфоринами повреждения в режиме низкого расхода энергии (Ota et al., 2013). Так, методами электронной микроскопии показано, что, воздействуя на клетки почечного эпителия линии Madin-Darby, Ply A/Oly A и Ply B индуцируют выработку внутри- и внеклеточных мембранных пузырьков, которые включают в себя как микровезикулы диаметром от 100 нм до 1 мкм, так и более крупные – до 10 мкм. В ходе процесса экструзии наблюдают проникновение за пределы клеток белков, задействованных в реализации пути ESCRT, а также кластерную сортировку холестерина, сфингомиелина и их аддуктов с перфоринами P. ostreatus на поверхности мембран (Skočaj et al., 2016).
Клеточные эффекты плевротолизинов опосредуют ряд явлений, изученных на изолированных органах животных. Показано, что остреолизин и плевротолизины на изолированных препаратах кишки и сердца вызывают мышечный спазм, иногда купируемый папаверином. На электрически стимулированном левом предсердии и на препарате Лангендорфа токсины вызывают сначала кратковременный положительный, а затем отрицательный инотропные эффекты (Juntes et al., 2009; Sabotič et al., 2015). В крови, оттекающей от искусственно перфузируемой печени грызунов после введения в систему перфоринов P. ostreatus возникает быстрое повышение концентрации К+. Далее, отражая динамику развития цитолиза гепатоцитов, в крови, которая оттекает от органа, нарастает содержание цитоплазматических и митохондриальных ферментов. В перфузируемой почке крыс перфорины также индуцируют массивную протеинурию и снижение ионной реабсорбции (Žužek et al., 2006; Juntes et al., 2009).
Плевротолизины приводят к специфическим изменениям на электроэнцефалограмме, которые связывают с генерацией пор на мембранных структурах гематоэнцефалического барьера (Frangež et al., 2017). В связи с этим считают, что наблюдаемая в ряде случаев остановка дыхательных движений может быть опосредована их прямым влиянием на дыхательный центр (Žužek et al., 2006). Вместе с тем показано, что у ваготомированных свиней в условиях общей анестезии с искусственной вентиляцией вследствие продленного введения Ply A/Oly A и PlyB возникают спазм и периваскулярный отек коронарных артерий, ведущие к выраженной брадикардии с желудочковыми экстрасистолами. Изменения периферического сосудистого сопротивления в условиях интоксикации носят двухфазный характер, возрастая на начальных этапах и далее переходя в α1-резистентную вазоплегию (Juntes et al., 2009). В связи с этим полагают, что дыхательные расстройства по отношению к оказываемому ими ишемическому повреждению миокарда также вторичны и носят рефлекторный характер (Lukoyanova et al., 2015; Vrecl et al., 2015).
Большинство исследователей сходятся во мнении, что непосредственная причина гибели животных вследствие внутривенного введения плевротолизинов кроется в коронарном вазоспазме, более детальные механизмы которого все еще подлежат уточнению (Frangež et al., 2017). В остром эксперименте на грызунах плевротолизины приводят к нарастанию концентрации К+, значения которой могут превышать референсные показатели в два раза, достигая 10 ммоль/л еще до развития гемолиза (Sabotič et al., 2015). Исследуя реактивность сосудов на плевротолизины в условиях обработки различными фармакологическими препаратами, приходят к выводу (Vrecl et al., 2015), что опосредующая коронарный вазоспазм ионная утечка Ca2+ может быть дополнительно обусловлена прямой активацией Ca2+-каналов L‑типа. ЛД50 плевротолизинов также вызывают эффект снижения натяжения сосудов эластического типа (аорта, артерии среднего калибра) на 7–30% (Rebolj et al., 2007). Вместе с тем наблюдаемая в ходе изменений сосудистого комплаенса депрессия диастолического артериального давления ведет к снижению перфузионного давления в коронарном круге кровообращения (Juntes et al., 2009).
За пределами эгеролизинов также известны перфорины грибов, которые содержат гомологичные Cry- и Cyt-токсинам B. thuringiensis участки (Bonfini, Buchon, 2016). Например, трехдоменные вольватоксины Vol A1 (44.8 кДа) и А2 (22.4 кДа) Вольвариеллы вольвовой (V. volvacea) включают аминокислотную последовательность, на 48% соответствующую δ-эндотоксину CytB (Pfam: PF01338). Токсичность Vol А2 для грызунов составляет ЛД50 = 1.23 мг/кг при внутривенном и ЛД50 = 40 мг/кг при внутрижелудочном введении, характеризуя его относительную устойчивость к протеолизу в просвете кишки (Lin et al., 2004; Sze et al., 2004). Как и эгеролизины, помимо гемолитической активности, Vol А2 обладает цито-, кардио- и нейротоксическими эффектами, выраженность которых отличается в связи с различными уровнями аффинности токсинов и особенностями генерируемых ими пор (Nayak et al., 2013).
Процесс образования проходимой для 70 кДа декстрана 18-мерной поры происходит при участии только Vol А2. Присутствие Vol А1 ингибирует процесс полимеризации, выполняя роль активируемого после протеолиза предшественника. Активная форма токсина Vol А2 состоит из С-концевого участка 1–127, задействованного в порообразовании, и N-концевого гепарин-связывающего фрагмента 165–199, который гомологичен лектиновому участку токсина индийской кобры Naja najа (L., 1758). Адгезивные свойства гепарин-связывающего фрагмента обеспечивают амфифильные β-нити, а также обнажаемая после протеолиза Vol А1 α-спираль. Помимо отмены процесса полимеризации посредством гепарина, Vol А2 характеризует тропность к сульфатированным гликоаминогликанам кардиомиоцитов (Weng et al., 2003; Lin et al., 2004).
Из числа многодоменных перфоринов известен лектин LSL Трутовика серно-желтого (L. sulphureus) с молекулярной массой 35 кДа, образующий комплекс из трех связанных между собой каналов-гексамеров с внутренним диаметром 4 нм. Лектин LSL обладает С-концевым порообразующим участком и N-концевым модулем с двумя дисахарид-связывающими сайтами на каждую субъединицу белка (Nayak et al., 2013). В связи с этим токсин характеризуют различные pН-оптимумы по эффектам связывания с мембранами и сборки канала: максимальная скорость взаимодействия с биомембранами возникает при pH = 5–7, в то время как оптимальную активность по гемолитическому эффекту наблюдают при рН = 7–8. В водном растворе рН = 7.4 он находится в динамическом равновесии между мономером и полимером – константа диссоциации Kd = 10–6 моль/л. Помимо агглютинации N-концевой лектиновый домен LSL оказывает цитотоксическое действие путем связывания поверхностных гликопротеинов и ингибирования их активности. Он обладает гомологическим сходством с подобными сайтами амарантина, абрина, участка HA1 ботулотоксина А и лектина CEL-III морского огурца Pseudocnus echinatus von Marenzeller, 1881 (Mancheño et al., 2005; Hassan et al., 2015).
Ко второму классу грибных токсинов, проявляющих мембранотоксические свойства, относят каталитические белки с L-аминооксидазной активностью в отношении аминокислот. Выявлено, что в составе плодовых тел Мухомора зеленого (A. phalloides) свойствами аминооксидазы обладает токсофаллин – токсин с массой 55 кДа, состоящий из 503 аминокислотных остатков, из числа которых приблизительно 7% составляет пролин (Stasyk et al., 2010). Подобный белок, фигурирующий под названием токсовирин, выделен из М. вонючего (A. virosa (Fr.) Bertill.) (Antonyuk et al., 2010). Гомологичным токсофаллину считают лектин в составе плодовых тел Лаковицы двухцветной (Laccaria bicolor (Maire) P.D. Orton) (Martin et al., 2008). Токсины обладают сродством к широкому спектру простых азотсодержащих соединений, продукты окисления которых представляют стабильные радикалы, приводящие к генерации процессов пероксидации липидов. Клеточная гибель вследствие действия токсофаллина и токсовирина возникает по независимому от активации каскада каспаз пути. Соединения с известной антиоксидантной активностью: аскорбиновая кислота и глютатион – in vitro могут полностью отменять опосредованную токсинами конденсацию хроматина и ядерную фрагментацию (Antonyuk et al., 2010; Stasyk et al., 2010).
Содержание токсофаллина в базидиомах A. phalloides составляет около 60 мг/кг свежего веса, в то время как концентрация токсовирина в A. virosa менее 10 мг/кг. Оксидативный стресс, оказываемый аминооксидазами в тех количествах, в которых они присутствуют в плодовых телах, носит отсроченный характер и совпадает во временных промежутках с аналогичным действием аманитинов, чей прооксидантный эффект связывают с атомом серы переходной валентности (Zheleva et al., 2007). Вместе с тем уровень биодоступности токсовирина и токсофаллина в условиях функционирования энтерального барьера, нарушенного основными бициклопептидными токсинами мухоморов секции Phalloideae, не изучен. В связи с этим полагают (Antonyuk et al., 2010), что при потреблении термически необработанных A. phalloides и A. virosa аминооксидазы могут играть весомую роль в проявлениях фаллоидинового синдрома, в течении которого ведущее значение отводят аманитинам.
Ингибиторы синтеза белка
Среди макромицетов описаны продуценты двух классов рибонуклеаз, которые приводят к нарушению белкового анаболизма посредством необратимой модификации сарцин-рициновой петли 28S рибосомальной РНК (рРНК) большой 60S субъединицы рибосом (Ennamany et al., 1995b; Olombrada et al., 2017). Непосредственная мишень цитотоксических рибонуклеаз – сарцин-рициновая петля SRL (sarcin-ricin loop) – состоит из 14 нуклеотидных оснований, последовательность которых высоко консервативна среди рибосом как эукариот, так некоторых видов прокариот. Она принадлежит к гуанозинтрифосфатазному центру GAC (GTPase associated center) – элементу трансляционного аппарата рибосом, ответственному за управление их непрерывным движением вдоль транслируемого рибонуклеинового полимера. Нарушение работы GAC также ингибирует связывание с рибосомами факторов элонгации EF (elongation factor). Суммарно нарушение этих процессов останавливает биосинтез белка, дефицит продукции которого в конечном итоге приводит к клеточной гибели (Guidebook to protein…, 1997).
Петля получила свое название от риботоксина α-сарцина и рибосом-инактивирующего белка рицина – первых и, как следствие, типовых токсинов среди своих классов. К рибосом-инактивирующим протеинам RIP (ribosome-inactivating protein) относят высокоспецифичные N-β-гликозидазы (КФ 3.2.2.22), нарушающие синтез белка посредством расщепления аминогликозидной связи рибозы с аденином, который локализован в рибосомах гепатоцитов крыс в положении А4324 (Wong et al., 2020). Ферментативное разрушение связи при адениновом основании SRL препятствует функционированию трансляционного механизма посредством блока гидролиза гуанозинтрифосфата.
Каталитический домен RIP-токсинов характеризует консервативная вторичная структура, которая формирует активный сайт, называемый РНК-связывающей расщелиной. К важным аспектам, определяющим уровень цитотоксичности RIP, относят их способность к клеточной пенетрации. В связи с этим выделяют три типа их архитектуры. RIP типа I обычно считают нетоксичными, поскольку они не обладают эффективным доменом связывания клеток или попадания в них. В то же время, наиболее ядовитые RIP II, помимо каталитического центра, содержат лектин-подобный участок. Несмотря на высокое структурное внутригрупповое сходство, биологическая активность токсинов II типа существенно варьирует – в их составе известны нетоксичные рибонуклеазы. В составе растительных токсинов также выделяют III тип RIP, среди которых одна подгруппа содержит участок с неизвестной ролью, а другая похожа на тип I, но содержит сайт, посредством которого токсин подвержен протеолитической активации (Lapadula, Ayub, 2017).
Селективная депуринизация RIP-токсинами основания А4324 делает смежную с ним фосфодиэфирную связь между гуанином и аденином в положениях G4325 и А4326 более восприимчивой к кислотно-основному и ферментативному гидролизу (Barbieri et al., 1997). Вместе с тем, риботоксины также расщепляют эту фосфодиэфирную связь, однако, уже напрямую. По причине обладания схожей ферментативной активностью, были высказаны предложения об объединении риботоксинов и RIP-токсинов в один класс (Schmitt et al., 1999). Тем не менее, в современном понимании остается представление, что составы классов разграничены по критериям механизмов действия токсинов и консервативных последовательностей их каталитических центров. Эти особенности определяют различную степень их способности к ингибированию синтеза белка и некоторые специфические проявления токсического процесса (Olombrada et al., 2017).
Риботоксины в природе выявлены среди 73 видов живых организмов (Lapadula, Ayub, 2017). Тем не менее, до недавнего времени среди грибов они были известны исключительно для микромицетов отдела Ascomycota (Landi et al., 2017; Ruggiero et al., 2019). К наиболее изученным из их числа относят риботоксины в пределах семейства Aspergillaceae Link: α-сарцин (ЛД50 = 12.0 мг/кг, мыши, внутрибрюшинно) и митогиллин (ЛД50 = 0.06 мг/кг, обезьяны, внутривенно) (Olombrada et al., 2017).
Поскольку как мишень данное образование универсально, способность к продукции нарушающих ее работу токсинов получила широкое распространение среди живых организмов (Lapadula, Ayub, 2017). Среди шляпочных грибов способность к ингибированию рибосомального синтеза описана относительно недавно. Необходимо заметить, что уровень изученности многих рибонуклеаз макромицетов, оказывающих цитотоксическое действие, недостаточен для их строгого отнесения к той или иной группе токсинов.
Активность подобного рода выявлена и охарактеризована среди белков в составе плодовых тел Вольвариеллы вольвовой (V. volvacea), Опенка зимнего (F. velutipes), Головача круглого (Calvatia caelata (Bull.) Morgan), Лиофиллюма симедзи (Lyophyllum shimeji (Kawam.) Hongo), Гипсизигуса мраморного (Hypsizygus marmoreus (Peck) H.E. Bigelow), Оксипоруса коркового (Oxyporus corticola (Fr.) Ryvarden) и Вешенки клубнеплодной (Pleurotus tuber-regium (Fr.) Singer) (Белова, 2018; Wong et al., 2020). Рибонуклеазы в пределах этих видов лишены способности к клеточной пенетрации, однако они обладают высокой активностью на модели ингибирования синтеза протеинов в бесклеточной среде. Например, IC50 рибосом калкаелина (C. caelata), плевтурегина (P. tuber-regium) и фламмулина (F. velutipes) составляют 4.0, 0.5 и 0.25 нмоль/л соответственно (Wong et al., 2020).
Несмотря на то, что перечисленные белки относят (Wong et al., 2020) к I типу RIP-токсинов, среди них способность к депуринизации, отличающая RIP от риботоксинов, была отмечена только у вольварина (V. volvaceae), марморина и гипсина (H. marmoreus), а также порицина (O. corticola), токсичность которого для крыс при внутрибрюшинном введении составляет ЛД50 = 791 мг/кг (Lapadula, Ayub, 2017). Вольварин, как и многие токсины RIP I типа, проявляет абортивную – ЕД50 (полуэффективная доза) = 0.15 мг/особь (мыши, внутрибрюшинно) и дезоксирибонуклеазную in vitro активности – IC50 = 0.42 мкмоль/л. Для гипсина и порицина, помимо абортивного, показан тератогенный эффект. Известно, что в ходе цитотоксического процесса они действуют по внешнему апоптотическому пути, активируя каспазу 8 (Ng et al., 2010). Тем не менее, в составе геномов V. volvacea и H. marmoreus консервативные последовательности, ответственные за биосинтез каталитического центра рибосом-инактивирующих токсинов, выявлены не были (Yao et al., 1998).
Таким образом, распределение большинства известных грибных белков, инактивирующих рибосом-опосредованный синтез, между RIP- и риботоксинами остается предметом дискуссий. Изначально ошибочно к RIP I типа был причислен один из первых известных белков с рибонуклеазной активностью среди базидиомицетов – эгеритин (17.0 кДа) (Landi et al., 2017). Ранее с эгеритином связывали пищевые интоксикации после неумеренного потребления культивируемой в пищевых целях A. aegerita, в данный момент эту роль отводят рассмотренному выше эгеролизину (Jin et al., 2014). Активное состояние риботоксина требует присутствия двухвалентных катионов: in vitro оптимальный эффект подавления белкового синтеза IC50 = 0.15 нмоль/л наблюдают в условиях насыщении раствора Zn2+ (Jin et al., 2014; Ruggiero et al., 2019). При крайне высокой активности в отношении насекомых, он не активен против нематод и слаботоксичен в отношении млекопитающих. Введение грызунам лиофиллизата эгеритина в течение недели приводит к развитию жирового гепатоза и почечного повреждения. Тем не менее, его острая внутрижелудочная токсичность невелика и составляет ЛД50 = 8.8 г/кг, в то время как количество эгеритина, экстрагируемого из свежих плодовых тел, едва достигает 12 мг/кг (Landi et al., 2017).
Большее, в сравнении с эгеритином, токсикологическое значение представляет ряд рибосом-инактивирующих токсинов среди болетовых грибов, которые, помимо нарушения работы рибосомального комплекса, вызывают массивный тромбоз (Gachet et al., 1996; Matsuura et al., 2007). В 1986 г. после двух случаев интоксикаций в результате потребления в сыром виде Ксантокониума родственного (Xanthoconium affine (Peck) Singer ≡ Boletus affinis Peck) были выявлены ядовитые свойства термолабильного протеина массой 22.0 кДа болаффинина – ЛД50 при внутрибрюшинном введении мышам составляет 61 мг/кг (Ennamany et al., 1995a). Следует отметить, что ранее не известный для наших территорий X. affine был обнаружен на о. Кунашир в 2016 г. Более устойчив к внешним воздействиям болесатин Руброболета сатанинского (Rubroboletus satanas (Lenz) Kuan Zhao et Zhu L. Yang ≡ Boletus satanas Lenz). В связи с этим интоксикации с летальным исходом встречают при потреблении R. satanas даже после предварительной обработки (Ennamany et al., 1995b; Merlet et al., 2012). Из плодовых тел Болета ядовитого (Boletus venenatus Nagas) выделен дублирующий его N-концевую последовательность болевенин. Болевенин с массой 33.0 кДа обладает per os токсичностью для мышей ЛД50/24 ч = 10 мг/кг (Matsuura et al., 2007).
В отличие от болаффинина, болесатин – мономерный токсин из 567 аминокислотных остатков массой 63.0 кДа, термостабилен при температурах до 70°C и относительно устойчив к протеолизу в просвете кишки (Kretz et al., 1991а,b). Гликопротеин проявляет двухфазную активность, ингибируя биосинтез белка у грызунов при внутрибрюшинном введении в дозах более 0.8 мг/кг, вместе с тем выступая в качестве активатора иммунного ответа и фактора пролиферации лимфоцитов при 0.05 мг/кг (Licastro et al., 1993). Для крыс суточная ЛД50 болесатина при внутривенном введении составляет 0.15 мг/кг, при внутрибрюшинном – 1.0 мг/кг, в то время как пероральная токсичность для мышей составляет ЛД50/24 ч = 3.3 мг/кг (Kretz et al., 1991а; Gachet et al., 1996). Лектиновый домен токсина детально не охарактеризован, однако известно, что он опосредует образование внутрисосудистого тромбоза, а галактоза в концентрации 23 ммоль/л снижает уровень его способности к агглютинации на 50% от исходного уровня (Ennamany et al., 1995b).
При внутрижелудочном поступлении 14C-болесатин наполовину проникает в кровь в течение 2 ч. После фазы перераспределения, его находят в эпителии кишки, почках, печени и в меньшей степени в тимусе, селезенке и легких. Так, внутрижелудочное введение крысам 1.65 мг/кг токсина приводит к ингибированию белкового анаболизма в почках на 26% и в печени на 48%. Тканевая концентрация подавления белкового синтеза токсина варьирует мало и составляют IC50 = 9 мкг/мл. В течение 24 ч с момента начала интоксикации всосавшийся токсин на 80% элиминирует через почки – скорость экскреции у грызунов составляет в среднем 0.67 мл/мин (Ennamany et al., 1995b).
При попадании среднесмертельных доз болесатина в желудок, смерть животных наступает в промежуток между 16 и 24 ч от начала интоксикации (Kretz et al., 1991а). Гибель в большинстве случаев возникает в результате массивного тромбоза. Вместе с этим наблюдают умеренно выраженный гастроэнтерит, значимые дистрофические изменения в легких, печени и почках (Kretz et al., 1991b; Gachet et al., 1996). Болесатин in vitro безызбирательно по группам крови человека агглютинирует эритроциты в диапазоне концентраций от 20 до 40 нмоль/л и тромбоциты при 320 нмоль/л. Введение аспирина, гепарина и клопидогреля предотвращает внутрисосудистое свертывание у грызунов, но не оказывает защитного эффекта на уровне среднесмертельных доз (Ennamany et al., 1995a).
Показано, что болесатин также способен к гидролизу макроэргических нуклеозидтрифосфатов и ингибированию репликации ДНК (IC50 = 20 мкг/мл) (Ennamany et al., 1995а,b). В этой связи авторами сделан поспешный вывод, что цитотоксичность болесатина опосредована в большей степени гидролизом оснований вне рибосом, а не недостатком функциональных белков. Не преуменьшая значимости серии исследований, которые носят пионерский характер и были выполнены до начала активного обсуждения этих механизмов в качестве отличительной черты RIP от риботоксинов, следует отметить, что данное ошибочное заключение до сих пор широко цитируют как в любительских справочниках, так и в некоторых медицинских руководствах по лечению отравлений.
В настоящее время внерибосомальные рибонуклеазная и нуклеозидтрифосфатазная активности обнаружены для подавляющего большинства растительных RIP-токсинов (Wong et al., 2020). В то же время, посредством изучения 52 различных RIP показано (Barbieri et al., 1997), что каждый из рассмотренных токсинов был способен катализировать in vitro разрушение связей аденина в составе ДНК. Известно, что внерибосомальные мишени характеризуются случайной встречаемостью последовательности GAGA и еще более низкой вероятностью возникновения специфической конфигурации, характерной для SRL, которые необходимы для высокого уровня аффинитета к RIP-токсинам. Поэтому считают, что внерибосомальные поли- и мононуклеотидные субстраты играют роль медленных и, как следствие, не основных биологических мишеней для данного класса токсинов (Guidebook to protein…, 1997). Таким образом, несмотря на то, что нарушаемые болесатином структуры внерибосомальных нуклеотидов могут быть задействованы как в обмене макроэргов, так и в развитии стохастических эффектов, рРНК, по логике вещей, сохраняет для данного рибосом-инактивирующего протеина статус предпочтительного субстрата, опосредующего его первостепенные токсикогенные эффекты.
Активаторы иммунной системы
Несмотря на то, что наличие антигенных свойств характерно для высокомолекулярных белков, к специфическим активаторам иммунной системы и иммунотоксикантам относят ксенобиотики, которые вызывают иммуносупрессию, усиление иммунитета или аутоиммунитета, либо индуцируют аллергические реакции. Отдельные бактериальные токсины могут воздействовать непосредственно на Т-клетки и антигенпрезентирующие эффекторы. Например, пирогенные токсины-суперантигены PTSAg (pyrogenic toxin-superantigens): стафилококковые энтеротоксин и токсин синдрома токсического шока TSST-1, пирогенные экзотоксины стрептококков, способны связывать главный комплекс гистосовместимости II типа и вариабельные β-субъединицы Т-клеточного рецептора. К следствиям Т-клеточного связывания с токсинами подобного рода относят пролиферацию лимфоцитов, а также массивное высвобождение активных форм ИЛ-1, -2 и -6, γ‑IFN и ФНО. Совместно эти цитокины приводят к развитию симптомов, напоминающих системную воспалительную реакцию (Schmitt et al., 1999; Matsuyama et al., 2014).
Аналогичным образом могут действовать некоторые продуцируемые макромицетами лектины, для которых известна митогенная активность в отношении Т-хелперов 1-го типа (табл. 2). Вместе с тем среди макромицетов известно около 30 иммуномодулирующих белков FIP (fungal immunomodulatory proteins). Представляя одно из направлений разработки новых лекарственных средств лектины FIP характеризуются разнонаправленными механизмами взаимодействия с иммунитетом (Xu et al., 2011).
Таблица 2.
Митогенный потенциал некоторых лектинов макромицетов в отношении лимфоцитов человека (адаптировано по: Xu et al., 2011; Hassan et al., 2015)
| Лектин | IC50, мкмоль/л | Продуцент |
|---|---|---|
| VVL | 0.12 | Volvariella volvacea (Bull.) Singer |
| HRL | 0.15 | Hygrophorus russula (Schaeff. ex Fr.) Kauffman |
| ALL | 1 | Floccularia luteovirens (Alb. & Schwein.) Pouzar ≡ Agaricus luteovirens Alb. & Schwein. |
| BEL | 1 | Boletus edulis Bull. |
| ACG | 2 | Cyclocybe parasitica (G. Stev.) Vizzini ≡ Agrocybe parasitica G. Stev. |
| PCL | 2 | Pleurotus citrinopileatus Singer |
| SCL | 4 | Schizophyllum commune Fr. |
| GCL | 1.5 | Ganoderma capense (Lloyd) Teng |
| HEA | 20 | Hericium erinaceus (Bull.) Pers. |
| CML | 26 | Cordyceps militaryaris (L.) Fr. |
| XSL | 31.2 | Boletus ferrugineus (Schaeff.) Alessio ≡ Xerocomus spadiceus (Schaeff. Ex Fr.) Quél. |
Среди лектинов съедобных грибов наиболее выраженное иммуностимулирующее действие выявлено для лектина VVL в составе базидиом Вольвариеллы вольвовой (V. volvacea). Так, по эффектам индукции выброса ИЛ-2 и ФНО и клеточного прироста Т-хелперов 1-го типа лектин VVL в девять раз превосходит эталонную активность конковалина А. Известно, что активация и пролиферация клеток VVL опосредована Ca2+-зависимым путем, о чем свидетельствует наблюдаемое увеличение внутриклеточного притока иона и ингибирование вызванных им эффектов посредством блокатора кальциневрина циклоспорина A (Sze et al., 2004). Аналогичные эффекты лектина ALL Флоккуларии желто-зеленоватой (F. luteovirens), которые в сравнении с VVL устойчивы к термическим, кислотно-основным изменениям и дегидратации в широком диапазоне, используют в качестве терапевтического средства (Hassan et al., 2015).
Избирательная митогенная активность в отношении Т-лимфоцитов человека, которая приводит к развитию выраженного воспалительного ответа, выявлена для рассмотренного выше рибосом-инактивирующего болесатина. В диапазоне от 1 до 20 нг/мл (0.016–0.3 нмоль/л) токсин R. satanas вызывает избирательный прирост Т-лимфоцитов, по силе превышающий действие селективного митогена фитогемагглютинина PHA в концентрации 20 мкг/мл. Тем не менее, при инкубации иммунокомпетентных клеток в среде, содержащей 10 мкг/мл токсина, его митогенный потенциал полностью нивелируется, что связывают с развитием эффекта инактивации белкового синтеза (Ennamany et al., 1998). В мононуклеарных клеточных культурах оптимальные митогенные дозы болесатина вызывают высвобождение ИЛ-1α и -1β, ИЛ-2, концентрации которых превышают контрольные показатели к 72 ч наблюдений в среднем в 10 раз (Licastro et al., 1993). В рамках митогенного эффекта, помимо ИЛ-2, болесатин напрямую и опосредованно, через высвобождение инозитолтрифосфата, активирует протеинкиназу С – PKC (protein kinase C). Внутрижелудочное введение токсина грызунам в дозах 100 и 200 мкг/кг приводит к приросту активности цитозольных киназ в 5.9 и 6.2 раза соответственно. На этом фоне у животных наблюдают гиперергическую реакцию со стороны маркеров системного воспалительного ответа (Licastro et al., 1993; Ennamany et al., 1995а,b). Плазменным концентрациям ИЛ-1β, которые аналогичны полученным в ходе серии исследований (Licastro et al., 1993), у кроликов сопутствует состояние перераспределительного шока, сопровождаемого нарушением доставки кислорода и утилизации его тканями, а также гипоперфузией последних (Matsuyama et al., 2014).
В подтверждение перечисленным экспериментальным исследованиям описаны (Merlet et al., 2012) 7 нелетальных случаев иммунореактивного состояния после потребления Руброболета сатанинского (R. satanas). В течение 24 ч от начала интоксикации концентрация прокальцитонина в плазме крови пораженных составляла в среднем 25.4 нг/мл, достигая у одного пострадавшего 49 нг/мл. Прокальцитонин представляет собой специфический, чувствительный и широко применяемый маркер клинической диагностики сепсиса, а его концентрации в плазме крови более 10 нг/мл, как правило, указывают на тяжелое течение множественной органной дисфункции, сопровождаемой шоком. В связи с наблюдением 2011 г. гиперпрокальцитонинемический синдром выделяют (White et al., 2019) в отдельную нозологическую форму среди мицетизмов, которая представляет одну из редких причин повышения данного маркера неинфекционной этиологии.
Известны лектины макромицетов, которые могут оказывать иммуносупрессивное действие. Так, лектин 42.0 кДа Шампиньона двуспорового (Agaricus bisporus (J.E. Lange) Imbach) подавляет активацию Т- и В-лимфоцитов, а TML-1 и TML-2 – лектины Рядовки монгольской (Tricholoma mongolicum S. Imai) – снижают численность их популяции (Wang et al., 1995). Вместе с тем среди перфоринов описан зависимый от каспазы 1 эффект активации инфламмасом, которые через комплекс промежуточных путей индуцируют гибель макрофагов и нейтрофилов (Bonfini, Buchon, 2016).
Протеазы
Протеолитические ферменты, также известные как протеазы, протеиназы и пептидазы, присутствуют во всех организмах. В плодовых телах макромицетов они выполняют как эндогенные, так и экзогенные функции. Для макромицетов показана тесная связь между видом секретируемых ими протеиназ и трофическим типом этих грибов (Денисова, 2009). Считают, что наиболее совершенные протеолитические механизмы характерны для древоразрушающих макромицетов, из которых наибольшего внимания с точки зрения фармакологической биотехнологии были удостоены металло- и сериновые протеазы (Кудрявцева и др., 2008).
Как показывают многочисленные исследования, внеклеточные протеазы ксилотрофов и сапрофитов лигнотрофной группы направлены на разрушение древесины и представляют с эволюционной точки зрения довольно развитый ферментативный аппарат (Sabotič et al., 2015). Некоторые из них, например субтилизино- и трипсиноподобные протеазы, рассматривают как факторы патогенности грибов в отношении растений, то есть их присутствие может указывать на способность макромицета жить и размножаться на живой ткани растения-хозяина, постепенно разрушая ее. Вместе с тем полагают, что протеазы макромицетов, подобно протеолитическим токсинам микроорганизмов, способны при попадании в клетки животных организмов специфически обеспечивать деградацию регуляторных белков, контролирующих ответ на тепловой шок, SOS-путь репарации ДНК и программируемой клеточной гибели (Кудрявцева и др., 2008), а также путем активации плазминогена опосредовать лизис кровяного сгустка (Денисова, 2009).
Внимание исследователей привлекает способность протеаз макромицетов к гидролизу плазминогена и фибрина млекопитающих. Подобная активность востребована в медицинской практике в лечении тромботических осложнений, и в ходе клинического решения этой задачи уже продолжительное время используют стрептокиназу – токсин гемолитического стрептококка – и аналогичные ей белки. Способность к биосинтезу макромицетами протеинов фибрино- и тромболитического действия впервые была показана на примере протеаз Опенка зимнего (F. velutipes) (Фалина и др., 1978).
Несмотря на наличие основополагающих работ, до конца двадцатого столетия подобная активность среди протеолитических ферментов высших грибов оставалась практически не изученной. Группой отечественных исследователей путем оценки экстрактов около 400 видов выявлен список грибов, продуцирующих субстратспецифичные к фибрину и тромбину протеазы, снижающие свертываемость крови и ускоряющие лизис сосудистых тромбов (Денисова, 2009). Кроме этого, в решении актуальной проблемы ими были предложены лабораторные регламенты получения фибринолитических препаратов из глубинных культур и плодовых тел Навозников пушистого (Coprinus lagopides P. Karst) и домашнего (Coprinus domesticus (Bolton) Gray). Предложенный способ позволяет экстрагировать из плодовых тел C. lagopides фибринолизин с выходом 3.0 мг/кг и удельной активностью 50–100 Ед/мг. На модели индуцированного тромбоза бедренной артерии крыс показано, что тромбо- и фибринолитический эффекты протеазы Навозника пушистого возникают при ее введении в дозе 0.5 мг/кг, в то время как 2.5 мг/кг позволяет в течение суток восстанавливать нарушенный кровоток сосудов конечности на 93% от исходного уровня (Шамцян и др., 2011).
На моделях in vitro рассматривают (Денисова, 2009) терапевтический потенциал протеолитических ферментов Хлорофиллюма свинцового (Chlorophyllum molybdites (G. Mey.) Massee = Lepiota morganii Peck). Гриб известен своими токсичными свойствами, которые связывают с молибдофиллизином – Zn2+-зависимой протеазой массой 23.0 кДа. Интоксикации C. molybdites, как правило, ассоциированы с желудочно-кишечными расстройствами, нарушениями свертывающей системы крови и системной вазоплегией (Eilers, Nelson, 1974).
Молибдофиллизин при рН = 7.0 сохраняет активность при температурах до 60°С, теряя ее при 100°С в течение 20 мин. Внутривенная токсичность лиофиллизата модибдофиллизина для собак варьирует в диапазоне ЛД50 = 78–84 мг/кг (Eilers, Nelson, 1974). Как и в клинических наблюдениях, в условиях экспериментального исследования введенный внутривенно до и после норадреналина он оказывает выраженный гипотензивный эффект (Yamada et al., 2012). Это можно расценивать исключительно как проявление α1-обратного агонизма (Yamada et al., 2012) и отвергать возможность других механизмов влияния молибдофиллизина на системное сосудистое сопротивление. Но подобные выводы дискутируемы в связи с неокончательно охарактеризованной активностью фермента.
ЗАКЛЮЧЕНИЕ
Подводя итоги рассмотрения токсинов макромицетов со структурой высокомолекулярных белков, можно отметить, что данный обзор представляет практически первую попытку обобщить и системно проанализировать их ядовитые свойства. Анализ литературы свидетельствует о том, что высокомолекулярные белковые токсины грибов изучают, если возможна перспектива их использования в биотехнологии и медицинской практике либо их продуценты относятся к модельным организмам для изучения биологической роли синтезируемых ими ферментов. В этой связи необходимо заметить, что нередко полученные в ходе исследований данные могут казаться недостаточными для строгого отнесения ядовитого белка к тому или иному классу токсинов. Несмотря на некоторые проведенные параллели с аналогичными или более известными соединениями, продуцируемыми представителями других царств, краеугольным камнем в понимании токсинов макромицетов остается отсутствие их собственной систематизации.
До настоящего времени ядовитые белки исследованы среди небольшой части видов огромного разнообразия таксонов и экологических групп высших грибов. Вместе с тем ввиду различной практической значимости они изучены в неодинаковой степени. Так, в отличие от достаточно полно охарактеризованного токсического процесса, провоцируемого эгеролизинами, в данный момент предприняты лишь единичные попытки по выявлению среди грибов продуцентов токсинов, влияющих на вторичные мессенджеры. За последнее время инструменты молекулярного и клеточного анализа предоставили новые возможности в их изучении.
Несомненно, что исследования в направлении детализации ядовитых свойств высших грибов актуальны и привлекают все больше внимания, а ответы на многие из этих вопросов будут получены уже в ближайшем будущем.
Список литературы
Белова Н.В. Рибонуклеазы высших грибов, перспективные в биомедицинских исследованиях // Микол. фитопатол. 2018. Т. 52. № 5. С. 301–305.
Вассер С.П. Наука о лекарственных шляпочных грибах: современные перспективы, достижения, доказательства и вызовы // Биосфера. 2015. Т. 7. № 2. С. 238–248.
Денисова Н.П. Тромболитические свойства ферментов базидиальных грибов // Пробл. мед. микол. 2009. № 4. С. 3–9.
Кудрявцева О.А., Дунаевский Я.Е., Камзолкина О.В., Белозерский М.А. Протеолитические ферменты грибов: особенности внеклеточных протеаз ксилотрофных базидиомицетов // Микробиология. 2008. Т. 77. № 6. С. 725–737.
Магазов Р.Ш., Степанов А.В., Чепур С.В., Савельев А.П. Токсины биологического происхождения: природа, структура, биологические функции и диагностика / Ред. Р.Ш. Магазов. Уфа: Башкирская энциклопедия, 2019. 352 с.
Современные методы изучения структуры и функций ионных каналов / Ред. О.С. Соколова. М.: КМК, 2020. 316 с.
Фалина Н.Н., Морозова Э.Н., Денисова Н.П. и др. Препарат фибринолитического действия из зимнего опенка (Flammulina velutipes) // Прикл. биохим. микробиол. 1978. Т. 14. № 5. С. 699–701.
Шамцян М.М., Петрищев Н.Н., Денисова Н.П. и др. Ферментный препарат тромболитического и фибринолитического действия из базидиального гриба рода Coprinus // Патент на изобретение RU 2435848 C1. 10.12.2011.
Al-Deen I.H.S., Twaij H.A.A., Al-Badr A.A., Istarabadi T.A. Toxicologic and histopathologic studies of Pleurotus ostreatus mushroom in mice // J. Ethnopharmacol. 1987. V. 21. № 3. P. 297–305.
Antonyuk V.O., Klyuchivska O.Y., Stoika R.S. Cytotoxic proteins of Amanita virosa Secr. mushroom: purification, characteristics and action towards mammalian cells // Toxicon. 2010. V. 55. № 7. P. 1297–1305.
Barbieri L., Valbonesi P., Bonora E. et al. Polynucleotide:adenosine glycosidase activity of ribosome-inactivating proteins: effect on DNA, RNA and poly(A) // Nucl. Acids Res. 1997. V. 25. № 3. P. 518–522.
Berry C., Crickmore N. Structural classification of insecticidal proteins – towards an in silico characterisation of novel toxins // J. Invertebr. Pathol. 2017. V. 142. P. 16–22.
Bonfini A., Buchon N. Pore-forming toxins trigger the purge // Cell Host Microbe. 2016. V. 20. № 6. P. 693–694.
Dal Peraro M., van der Goot F.G. Pore-forming toxins: ancient, but never really out of fashion // Nat. Rev. Microbiol. 2015. V. 14. № 2. P. 77–92.
Eilers F.I., Nelson L.R. Characterization and partial purification of the toxin of Lepiota morganii // Toxicon. 1974. V. 12. № 6. P. 557–563.
Ennamany R., Kretz O., Creppy E.E. Effect of bolesatine on phospholipid/calcium dependent protein kinase in Vero cells and in rat thymus // Arch. Toxicol. 1995a. V. 69. № 9. P. 624–630.
Ennamany R., Lavergne J.P., Reboud J.P. et al. Mode of action of bolesatine, a cytotoxic glycoprotein from Boletus satanas Lenz. mechanistic approaches // Toxicology. 1995b. V. 100. № 1–3. P. 51–55.
Ennamany R., Bingen A., Creppy E.E. et al. Aspirin and heparin prevent hepatic blood stasis and thrombosis induced by the toxic glycoprotein bolesatine in mice // Hum. Experiment. Toxicol. 1998. V. 17. № 11. P. 620–624.
Faulstich H., Bühring H.J., Seitz J. Physical properties and function of phallolysin // Biochemistry. 1983. V. 22. № 19. P. 4574–4580.
Fernandez Espinar M.T., Labarère J. Cloning and sequencing of the Aa-Pri1 gene specifically expressed during fruiting initiation in the edible mushroom Agrocybe aegerita, and analysis of the predicted amino-acid sequence // Curr. Genet. 1997. V. 32. № 6. P. 420–424.
Ford W.W. The distribution of haemolysins, agglutinins, and poisons in fungi, especially the Amanitas, the Entolomas, the Lactarius and the Inocybes // J. Pharmacol. Exp. Ther. 1911. V. 2. P. 285–318.
Frangež R., Šuput D., Molgó J., Benoit E. Ostreolysin A / pleurotolysin B and equinatoxins: structure, function and pathophysiological effects of these pore-forming proteins // Toxins. 2017. V. 9. № 4. P. 128.
Gachet C., Ennamany R., Kretz O. et al. Bolesatine induces agglutination of rat platelets and human erythrocytes and platelets in vitro // Hum. Experiment. Toxicol. 1996. V. 15. № 1. P. 26–29.
Gouaux E. Channel-forming toxins: tales of transformation // Curr. Opin. Struct. Biol. 1997. V. 7. № 4. P. 566–573.
Guidebook to protein toxins and their use in cell biology / Eds R. Rappuoli, C. Montecucco. Oxford: Oxford Univ. Press, 1997. 275 p.
Hassan M.A., Rouf R., Tiralongo E. et al. Mushroom lectins: specificity, structure and bioactivity relevant to human disease // Int. J. Mol. Sci. 2015. V. 16. № 4. P. 7802–7838.
Henrici A.T. An endotoxin from Aspergillus fumigatus // J. Immunol. 1939. V. 36. P. 319–338.
Jin Y., Che T., Yin Y. et al. Lethal protein in mass consumption edible mushroom Agrocybe aegerita linked to strong hepatic toxicity // Toxicon. 2014. V. 90. P. 273–285.
Juntes P., Rebolj K., Sepcić K. et al. Ostreolysin induces sustained contraction of porcine coronary arteries and endothelial dysfunction in middle- and large-sized vessels // Toxicon. 2009. V. 54. № 6. P. 784–792.
Kretz O., Creppy E.E., Dirheimer G. Disposition of the toxic protein bolesatine in rats: its resistance to proteolytic enzymes // Xenobiotica. 1991a. V. 21. № 1. P. 65–73.
Kretz O., Creppy E.E., Dirheimer G. Characterization of bolesatine, a toxic protein from the mushroom Boletus satanas Lenz and it’s effects on kidney cells // Toxicology. 1991b. V. 66. № 2. P. 213–224.
Landi N., Pacifico S., Ragucci S. et al. Purification, characterization and cytotoxicity assessment of ageritin: the first ribotoxin from the basidiomycete mushroom Agrocybe aegerita // Biochim. Biophys. Act. Gen. Subj. 2017. V. 1861. № 5. Pt A. P. 1113–1121.
Lapadula W., Ayub M. Ribosome inactivating proteins from an evolutionary perspective // Toxicon. 2017. V. 136. P. 6–14.
Licastro F., Morini M. C., Kretz O. et al. Mitogenic activity and immunological properties of bolesatine, a lectin isolated from the mushroom Boletus satanas Lenz. // Int. J. Biochem. 1993. V. 25. № 5. P. 789–792.
Lin J., Lin Y., Chen C. et al. Cardiotoxic protein from edible mushrooms // Nature. 1974. V. 252. P. 235–237.
Lin S., Lo Y., Lin J., Liaw Y.-C. Crystal structures and electron micrographs of fungal volvatoxin A2 // J. Mol. Biol. 2004. V. 343. № 2. P. 477–491.
Lukoyanova N., Kondos S.C., Farabella I. et al. Conformational changes during pore formation by the perforin-related protein pleurotolysin // PLoS Biol. 2015. V. 13. № 2. P. e1002049.
Lv H., Kong Y., Yao Q. et al. Nebrodeolysin, a novel hemolytic protein from mushroom Pleurotus nebrodensis with apoptosis-inducing and anti-HIV-1 effects // Phytomedicine. 2009. V. 16. № 2–3. P. 198–205.
Mancheño J.M., Tateno H., Goldstein I.J. et al. Structural analysis of the Laetiporus sulphureus hemolytic pore-forming lectin in complex with sugars // J. Biol. Chem. 2005. V. 280. № 17. P. 17251–17259.
Martin F., Aerts A., Ahrén D. et al. The genome of Laccaria bicolor provides insights into mycorrhizal symbiosis // Nature. 2008. V. 452. P. 88–92.
Matsuura M., Yamada M., Saikawa Y. et al. Bolevenine, a toxic protein from the Japanese toadstool Boletus venenatus // Phytochemistry. 2007. V. 68. № 6. P. 893–898.
Matsuyama S., Hayakawa K., Sakuramoto K. et al. Vasodilating prostaglandin E1 does not reproduce interleukin-1β-induced oxygen metabolism abnormalities in rabbits // Acute Med. Surg. 2014. V. 2. № 1. P. 40–47.
Merlet A., Dauchy F.A., Dupon M. Hyperprocalcitonemia due to mushroom poisoning // Clin. Infect. Dis. 2012. V. 54. № 2. P. 307–308.
Mondego J.M., Carazzolle M.F., Costa G.G. et al. A genome survey of Moniliophthora perniciosa gives new insights into Witches’ Broom disease of cacao // BMC Genom. 2008. V. 9. P. 548.
Mori S., Nakagawa-Yoshida K., Tsuchihashi H. et al. Mushroom worker’s lung resulting from indoor cultivation of Pleurotus ostreatus // Occup. Med. 1998. V. 48. № 7. P. 465–468.
Nayak A.P., Green B.J., Beezhold D.H. Fungal hemolysins // Med. Mycol. 2013. V. 51. № 1. P. 1–16.
Ng T., Lam J., Wong J. et al. Differential abilities of the mushroom ribosome-inactivating proteins hypsin and velutin to perturb normal development of cultured mouse embryos // Toxicol. in vitro. 2010. V. 24. № 4. P. 1250–1257.
Ngai P., Ng T. A hemolysin from the mushroom Pleurotus eryngii // Appl. Microbiol. Biotechnol. 2006. V. 72. P. 1185–1191.
Novak M., Krpan T., Panevska A. et al. Binding specificity of ostreolysin A6 towards Sf9 insect cell lipids // Biochim. Biophys. Acta – Biomembr. 2020. V. 1862. № 9. P. e183307.
Odenthal K.P., Seeger R., Vogel G. Toxic effects of phallolysin from Amanita phalloides // Naunyn-Schmiedeberg’s Arch. Pharmacol. 1975. V. 290. № 2–3. P. 133–143.
Odenthal K.P., Seeger R., Braatz R. et al. Damage in vitro to various organs and tissues by rubescenslysin from the edible mushroom Amanita rubescens // Toxicon. 1982. V. 20. № 4. P. 765–781.
Olombrada M., Lázaro-Gorines R., López-Rodríguez J. et al. Fungal ribotoxins: a review of potential biotechnological applications // Toxins. 2017. V. 9. № 2. P. 1–21.
Ota K., Leonardi A., Mikelj M. et al. Membrane cholesterol and sphingomyelin, and ostreolysin A are obligatory for pore-formation by a MACPF/CDC-like pore-forming protein, pleurotolysin B // Biochim. 2013. V. 95. № 10. P. 1855–1864.
Rebolj K., Batista U., Sepcić K. et al. Ostreolysin affects rat aorta ring tension and endothelial cell viability in vitro // Toxicon. 2007. V. 49. № 8. P. 1211–1213.
Ruggiero A., Garcia-Ortega L., Moreira M. et al. Binding and enzymatic properties of ageritin, a fungal ribotoxin with novel zinc-dependent function // Int. J. Biol. Macromol. 2019. V. 136. P. 625–631.
Sabotič J., Ohm R.A., Künzler M. Entomotoxic and nematotoxic lectins and protease inhibitors from fungal fruiting bodies // Appl. Microbiol. Biotechnol. 2015. V. 100. № 1. P. 91–111.
Schmitt C.K., Meysick K.C., O’Brien A.D. Bacterial toxins: friends or foes? // Emerg. Infect. Dis. 1999. V. 5. № 2. P. 224–234.
Seeger R. Demonstration and isolation of phallolysin haemolytic toxin from Amanita phalloides // Naunyn-Schmiedeberg’s Arch. Pharmacol. 1975. V. 287. P. 277–287.
Seeger R., Bunsen E. Degranulation of rat mast cells in vitro by the fungal cytolysins phallolysin, rubescenslysin and fascicularelysin // Naunyn-Schmiedeberg’s Arch. Pharmacol. 1980. V. 315. № 2. P. 163–166.
Seeger R., Odenthal K.P., Mengs U. Toxic effects in mouse and rat of rubescenslysin from Amanita rubescens // Toxicon. 1981. V. 19. № 3. P. 409–417.
Skočaj M., Yu Y., Grundner M. et al. Characterisation of plasmalemmal shedding of vesicles induced by the cholesterol/sphingomyelin binding protein, ostreolysin A-mCherry // Biochim. Biophys. Acta. 2016. V. 1858. № 11. P. 2882–2893.
Stasyk T., Lutsik-Kordovsky M., Wernstedt C. et al. A new highly toxic protein isolated from the death cap Amanita phalloides is an L-amino acid oxidase // FEBS J. 2010. V. 277. № 5. P. 1260–1269.
Sze S., Ho J., Liu W. Volvariella volvacea lectin activates mouse T lymphocytes by a calcium dependent pathway // J. Cell. Biochem. 2004. V. 92. № 6. P. 1193–1202.
Vrecl M., Babnik M., Sepčić K. et al. Effect of the ostreolysin A/pleurotolysin B pore-forming complex on intracellular Ca2+ activity in the vascular smooth muscle cell line A10 // Toxicol. in vitro. 2015. V. 29. № 8. P. 2015–2021.
Wang H.X., Ng T.B., Liu W.K. et al. Isolation and characterization of two distinct lectins with antiproliferative activity from the cultured mycelium of the edible mushroom Tricholoma mongolicum // Int. J. Pept. Prot. Res. 1995. V. 46. № 6. P. 508–513.
Weng Y., Lin Y., Hsu C., Lin J.-Y. Functional domains of a pore-forming cardiotoxic protein, volvatoxin A2 // J. Boil. Chem. 2003. V. 279. № 8. P. 6805–6814.
White J., Weinstein S.A., De Haro L. et al. Mushroom poisoning: a proposed new clinical classification // Toxicon. 2019. V. 157. P. 53–65.
Wong J.H., Bao H., Ng T.B. et al. New ribosome-inactivating proteins and other proteins with protein synthesis-inhibiting activities // Appl. Microbiol. Biotechnol. 2020. V. 104. № 10. P. 4211–4226.
Xu X., Yan H., Chen J., Zhang X. Bioactive proteins from mushrooms // Biotechnol. Adv. 2011. V. 29. № 6. P. 667–674.
Yamada M., Tokumitsu N., Saikawa Y. et al. Molybdophyllysin, a toxic metalloendopeptidase from the tropical toadstool Chlorophyllum molybdites // Bioorg. Med. Chem. 2012. V. 20. № 22. P. 6583–6588.
Yao Q., Yu M., Ooi L. et al. Isolation and characterization of a type 1 ribosome-inactivating protein from fruiting bodies of the edible mushroom (Volvariella volvacea) // J. Agric. Food Chem. 1998. V. 46. № 2. P. 788–792.
Zheleva A., Tolekova A., Zhelev M. et al. Free radical reactions might contribute to severe alpha-amanitin hepatotoxicity – a hypothesis // Med. Hypoth. 2007. V. 69. № 2. P. 361–367.
Žužek M.C., Maček P., Sepčić K. et al. Toxic and lethal effects of ostreolysin, a cytolytic protein from edible oyster mushroom (Pleurotus ostreatus) in rodents // Toxicon. 2006. V. 48. № 3. P. 264–271.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии