

Успехи современной биологии, 2021, T. 141, № 4, стр. 399-416
К изучению линьки волосяного покрова мелких лесных млекопитающих (опыт аналитического обзора состояния проблемы)
1 Петрозаводский государственный университет
Петрозаводск, Россия
2 Карельский научный центр РАН
Петрозаводск, Россия
* E-mail: Ivanter@petrsu.ru
Поступила в редакцию 17.02.2021
После доработки 26.02.2021
Принята к публикации 26.02.2021
Аннотация
Для мелких насекомоядных и грызунов, существующих в условиях постоянных термических дефицитов и проводящих много времени в лесной подстилке и ходах нор, постоянно соприкасаясь с твердым субстратом, регулярная смена волосяного покрова имеет особое значение. Согласно проведенным исследованиям, сроки и интенсивность линьки изменяются в зависимости от экологических условий и физиологического состояния зверьков. В этой связи конкретные особенности хода и темпов смены волос у особей различных половых и возрастных групп могут служить своеобразным индикатором состояния всей популяции и сигнализировать о серьезных нарушениях важных эколого-физиологических и популяционных процессов. Полученные данные свидетельствуют об отсутствии существенных видовых различий в сроках, интенсивности и ходе сезонных линек, тогда как связь с полом, возрастом и состоянием популяции выступает достаточно отчетливо. На основе изучения массовых видов землероек и грызунов Карелии у них выявлены исключительная устойчивость и однообразие в последовательности смены волос, которые почти не зависят от образа жизни зверьков. У большинства видов она происходит по сублатеральному (дорсальному) типу, при котором новые волосы появляются сначала на нижних частях боков и головы, затем на брюшке и спине, и в последнюю очередь вылинивают верх головы и задняя часть спины.
ВВЕДЕНИЕ
Линька, то есть сезонная смена шерстного покрова и тесно связанные с ней изменения в коже млекопитающих, представляют собой очень тонкий биологический процесс¸ призванный обеспечить целостность покровов тела как основного защитного и теплоизоляционного образования. Для мелких насекомоядных и грызунов, проводящих много времени в лесной подстилке и ходах нор и постоянно соприкасающихся с твердым субстратом, регулярное перелинивание имеет особое значение, так как их волосяной покров быстро изнашивается и требует своевременной замены. Необходимость периодической смены меха диктуется также сезонными изменениями климата, являясь средством увеличения теплоотдачи летом и уменьшения – зимой. Как показали наши исследования, сроки и интенсивность линьки изменяются в зависимости от пола и возраста, а также от физиологического состояния зверьков, кормовых и погодных условий. Поэтому конкретные особенности хода и темпов линьки у животных различных возрастных и половых групп могут служить своеобразным индикатором состояния всей популяции и сигнализировать о серьезных нарушениях важных эколого-физиологических и популяционных процессов.
Как показывают исследования (Кузнецов, 1952; Соколов, 1973; Чернова, Целикова, 2004; Ивантер, 2018, 2021; Hyvärinen et al., 1969 и др.), развитие нового волоса начинается с закладки остевых волос, от сумок которых отпочковываются уже зачатки пуховых. Процесс замещения волос у различных групп млекопитающих протекает неодинаково. В отличие от хищных зверей, зачаток нового волоса которых закладывается из клеток дна луковицы старого, и по мере роста новый волос выталкивает отделившихся от луковицы, но довольно долго удерживающихся в волосяной сумке старой, у грызунов и насекомоядных закладка зачатков новых волос происходит совершенно независимо от волосяных сумок старых, выпадающих. Так что в противоположность хищникам группировка волос новой шерсти не соответствует у них таковым старой.
Своеобразно протекает у них и линька волосяного покрова. В зачатках новых волос концентрируются зерна пигмента. Просвечивая через подкожную клетчатку, они придают синеватый цвет мездре (на внутренней поверхности кожи). И поскольку в разных участках шкурки линька обычно протекает не одновременно, а в определенной последовательности, на мездре образуется характерный рисунок – узор линьки, состоящий из так называемых линных пятен. По их расположению и форме удается судить о наступлении той или иной стадии линьки. С ростом волос, выносящих пигмент из кожи, происходит посветление мездры, идущее обычно в той же последовательности, в какой шло ее потемнение. Полностью очистившаяся от пятен мездра знаменует окончание процесса линьки.
С линькой нередко бывает связано изменение структуры шерсти и ее окраски, выраженное порой очень резко. Подлежат изменению и другие структуры. Так, дерма в ходе линьки разрыхляется развивающимися зачатками новых волос и соответственно этому утолщается, а в периоды между линьками уплотняется. Жировая прослойка, неплохо развитая зимой, к лету истончается или полностью исчезает. В период линьки также увеличивается потребность в минеральном питании и витаминах, повышается белковый обмен, усиливается возбудимость. Таким образом, в связанном с линькой физиологическом процессе участвует весь организм животного, но особенно эффективно гормональное воздействие гипофиза и щитовидной железы.
Между тем изучение линьки и других приспособительных особенностей волосяного покрова мелких грызунов и насекомоядных представляет большой научный интерес, и прежде всего с позиций эволюционной морфологии, экологической физиологии и таксономии. Обитание этих животных в условиях постоянных термических дефицитов, а также свойственное им общее несовершенство химической терморегуляции выработали у них в процессе эволюции целый комплекс экологических и морфофизиологических приспособлений, направленных на сохранение оптимального энергетического баланса между организмом и средой при низких и крайне неустойчивых окружающих температурах. Отсюда возникла необходимость подробного исследования сезонно-возрастных изменений таких атрибутов физической терморегуляции массовых видов мелких млекопитающих, как строение и теплозащитные свойства шерсти, микроструктура волос, географическая и популяционная изменчивость этих показателей. Надо учитывать и общее слабое освещение этих вопросов в специальной литературе. В этих обстоятельствах представляется целесообразным представить заинтересованному читателю полученные нами данные по линьке волосяного покрова массовых видов мелких млекопитающих таежного Севера, тем более, что по объему, диапазону и адекватности используемых методик они значительно превосходят практически все предшествующие публикации, причем как в нашей стране, так и за рубежом.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом для специальных исследований послужили 293 шкурки мелких зверьков, добытых в 1966–2014 гг. в различных районах Карелии, из них 68 – крота, 43 – обыкновенной, 44 – средней, 38 – малой землероек-бурозубок, 32 – водяной куторы, 26 – лесной мышовки, 50 – рыжей полевки, 36 – темной полевки и 24 – полевки-экономки.
Изучение линьки волосяного покрова проводилось обычными, использовавшимися в аналогичных исследованиях методами (Кузнецов, 1932; Церевитинов и др., 1948; Депарма, 1951; Когтева, 1963; Соколов и др., 1986). При этом особенно широко применялся метод “картирования” – зарисовка на коллекционных карточках с трафаретами контура шкурки изменений окраски мездры и расположения линных пятен. Кроме того, в световом микроскопе определяли функциональное состояние наружных покровов, дифференцировали зрелые волосы от растущих, различали пигментированные и непигментированные участки закладки волоса у изученных образцов кожи и волос. Необходимость световой микроскопии была в данном случае вызвана тем, что только с ее помощью можно, например, безошибочно отличить растущий зимний волос от окончательно выросшего летнего той же длины. Для этого достаточно рассмотреть под бинокулярным микроскопом строение волосяной луковицы: у зрелых волос оно закрытого (колбовидного) типа, а у растущих – открытого (сосочкового). Кроме того, закладка волос не везде сопровождается потемнением мездры. Поскольку волосы землероек имеют зонарную окраску и их концевые сегменты на разных участках шкурки неодинаковы по цвету (на спине темно-коричневые, на боках – бурые, а на брюхе – белые), то на боках и спине линные пятна бывают видны сразу, а на брюхе – лишь после того, как пробьются через кожу былые верхушки волос. Сегменты же стержня, лежащие ниже вершины, окрашены в черный цвет. Поэтому при микроскопии кожи с брюшка без видимых признаков линьки легко обнаруживаются характерные изменения (в частности, функционирующие волосяные фолликулы), свидетельствующие о сезонной смене волос, даже при так называемой скрытой пигментации.
Кроме того, для установления сроков и стадий линьки крота на пунктах заготовки пушнины было просмотрено (в том числе с помощью бинокулярного микроскопа) более 150 его шкурок и привлечены данные о линьке различных видов мелких млекопитающих, содержащихся в 1800 коллекционных карточках лаборатории зоологии КарНЦ РАН.
Трудности в изучении линьки землероек связаны также с разновозрастным составом популяции. Продолжительность линьки одного зверька, как известно, невелика (Borowski, 1958). Но у всей популяции этот процесс растянут, и в один и тот же период в ее составе встречаются особи, находящиеся на разных стадиях линьки. Отсюда неизбежные сложности в определении последовательности и топографии сезонной смены шерсти.
Определение теплоизоляционных свойств шкурок (по удельной теплопроводности) выполняли с помощью специального прибора ИТ-3, изготовленного по нашему заказу для измерения теплопроводности небольших по размеру неметаллических объектов в Киевском институте технической теплофизики АН УССР. Существенное преимущество этого прибора заключается в большой чувствительности и точности, а также в том, что все необходимые для расчета теплопроводности величины получают непосредственным измерением.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Согласно многим авторам (Огнев, 1950; Павлова, 1951; Фадеев, 1955, и др.), у млекопитающих первыми обычно развиваются направляющие и остевые волосы. Что же касается очередности появления волос разных категорий, то у обыкновенной бурозубки проследить ее затруднительно. На разных стадиях линьки в коже одновременно находятся широкие фолликулы будущих направляющих и остевых волос и более узкие – пуховых. Позднее, когда становятся заметны уже хорошо сформированные концевые сегменты волос всех категорий и они пробиваются наружу, мездра начинает постепенно светлеть и утончается.
Как видно из приведенных в табл. 1 и рис. 1 данных, первая линька прибылых обыкновенных бурозубок, покинувших гнезда, происходит только осенью – в сентябре–октябре. Первые особи с темными пятнами на мездре, занимающими 20–90% площади шкурки (в среднем 46%), встречались, начиная с десятых чисел сентября (11.09.1959 г.; 12.09.1970 г., 13 и 16.09.2005 г.), во второй декаде этого месяца линяет уже более 60% исследованных зверьков, а в третьей – 80%, причем интенсивность линьки (здесь и далее под термином “интенсивность” понимается относительная площадь шкурки, затронутая линькой, то есть пигментированием) составляет в среднем 63%. В октябре линька прибылых продолжается. В первой декаде этого месяца линяет 85% исследованных обыкновенных бурозубок с интенсивностью 8–100%, в среднем 73%, а во второй – 50%, со средней интенсивностью 68% (3–100%). В третьей декаде октября линька прибылых заканчивается. Последние линявшие прибылые со шкурками, пигментированными на 3–5%, были пойманы 3 и 6.11.1959 г. и 2.11.2005 г. Волосы на шкурках зверьков, отловленных в начале ноября, были разными, то есть на одной и той же шкурке можно было встретить и летние зрелые волосы с луковицами закрытого типа, и волосы на стадии роста (интенсивно пигментированная прикорневая часть стержня волоса, луковица открытого типа), и зрелые зимние волосы: шестисегментные остевые I порядка, семисегментные остевые II порядка и пуховые – все с закрытыми луковицами. Летние волосы постепенно выпадают, что подтверждается отсутствием их на шкурках зверьков из зимних и ранневесенних сборов, шерсть у таких землероек целиком состоит из зимних волос. Все это свидетельствует о полной смене волосяного покрова в период осенней линьки молодняка. Причем следует отметить необычайную стабильность сроков осенней линьки сеголеток всех видов рода Sorex, которые с удивительным постоянством повторяются из года в год.
Таблица 1.
Интенсивность линьки обыкновенной бурозубки по месяцам
| Месяц | Самцы | Самки | ||||
|---|---|---|---|---|---|---|
| число исследованных зверьков | из них линяющих, % | относительная площадь пигментированной шкурки, % от площади всей шкурки | число исследованных зверьков | из них линяющих, % | относительная площадь пигментированной шкурки, % от площади всей шкурки | |
| Прибылые | ||||||
| VI | 15 | 8.8* | <1 | 14 | 21.4* | <1 |
| VII | 310 | 2.2* | <1 | 249 | 5.2* | <1 |
| VIII | 328 | 0.3* | <1 | 331 | 0.9* | <1 |
| IX | 105 | 74.3 | 56 | 62 | 71.0 | 56 |
| X | 31 | 71.0 | 67 | 52 | 65.4 | 46 |
| XI | 18 | 11.1* | <1 | 22 | 4.5* | <1 |
| XII | 17 | 24.6* | <1 | 13 | 38.5* | <1 |
| I | 14 | – | – | 13 | – | – |
| II | 16 | 12.5* | <1 | 1 | – | – |
| III | 14 | – | – | 14 | – | – |
| Зимовавшие | ||||||
| IV | 15 | 6.7 | 20 | 3 | 33.3 | 30 |
| V | 7 | 47.0 | 50 | 7 | 28.5 | 90 |
| VI | 29 | 24.1 | 17 | 14 | 14.2 | 3 |
| VII | 48 | 14.6 | 6 | 61 | 16.5 | 9 |
| VIII | 19 | 31.6 | 2 | 45 | 9.7 | 22 |
| IX | 4 | 50.0 | 28 | 11 | – | – |
| X | 3 | 66.6 | 100 | 6 | 50.0 | <1 |
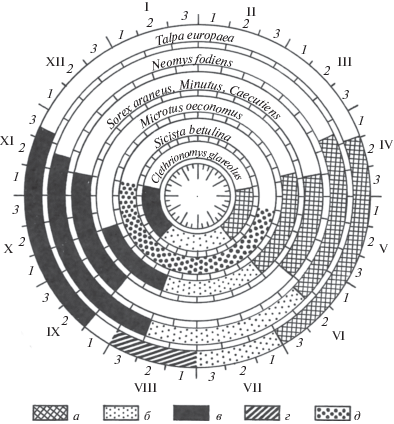
Рис. 1.
Сроки линьки мелких насекомоядных и грызунов Карелии. а – весенняя, б – ювенильная, в – осенняя, г – компенсаторная, д – летняя.
Отсутствуют и географические различия. В Англии, Нидерландах, Польше, Германии, Финляндии и различных областях России (в Московской обл., в Татарстане, в Сибири и т.д.) осенью линька прибылых происходит в одни и те же сроки – в сентябре–октябре (Огнев, 1928; Формозов, 1948; Строганов, 1957; Попов, 1960; Dehnel, 1949; Stein, 1954; Crowcroft, 1957; Skaren, 1964, 1973; Michielsen, 1966; Borowski, 1968).
Летней “промежуточной” линьки молодых зверьков, о которой пишет Штейн (Stein, 1954), мы ни у одного из исследованных видов землероек не обнаружили. Из 1873 сеголеток всех видов землероек-бурозубок, пойманных в июне–августе, лишь у 44 (2.3% от числа просмотренных) имелась темная точечная пигментация на мездре (интенсивность линьки не более 1%). Других признаков смены меха, в частности роста нового волоса, у них не было, так что считать их линяющими, очевидно, нет оснований. В нашем распоряжении материал позволяет установить приблизительный порядок смены меха на различных участках шкурки во время осенней линьки сеголеток (рис. 2). Судя про этим данным, начинается она обычно на спине у огузка и распространяется вперед к голове, переходя постепенно на бока и брюшко. Казалось бы, и волосы соответственно должны заканчивать рост сначала на спине, и в последнюю очередь на брюшке, однако при тщательном исследовании ноябрьских шкурок зрелые зимние волосы обнаруживались одновременно и на верхней, и на нижней стороне тела (на линии середины туловища). Возможно, это объясняется тем, что волосы всех категорий, растущие на спине, обычно превосходят по длине волосы на брюшке. Поэтому волосы на дорзальной стороне тела, хотя и пробиваются раньше, окончательной зрелости достигают одновременно с волосами нижней стороны тела (более длинные волосы дольше растут). В то же время описанный выше рисунок осенней линьки не всегда бывает достаточно четким еще и потому, что картина затушевывается неодновременным вступлением зверьков в линьку, а также многочисленными индивидуальными отклонениями.
Рис. 2.
Последовательность появления линных пятен у мелких насекомоядных млекопитающих. (а) – обыкновенная бурозубка, 1 – осенняя линька; 2 – весенняя I; 3 – весенняя II; (б) – водяная кутора; (в) – крот; 1 – ювенильная линька; 2 – весенняя; 3 – осенняя.

Большинство авторов, обсуждая ход весенней линьки у мелких насекомоядных (Огнев, 1928; Кузнецов, 1952; Докучаев, 1981; Jackson, 1928; Hamilton, 1940; Dehnel, 1949, 1950; Findley, Jones, 1956; Crowcroft, 1957; Meester, 1958; Williams, 1962; Michielsen, 1966 и др.), описывают волны длинного и короткого волоса, следующие друг за другом в особом порядке на разных участках тела зверька, но ничего не сообщают о потемнении мездры. Между тем, при рассмотрении осенней линьки они специально подчеркивают это явление. Все авторы единодушно отмечают, что осенняя линька начинается в крестцовой области и продолжается в направлении к голове, постепенно переходя на брюшную сторону. Весенняя же линька, наоборот, начинается с головы и распространяется к хвосту и брюху. Тем не менее, другие авторы (Попов, 1960; Scaren, 1973, 1979) утверждают, что весенняя линька обыкновенной бурозубки проходит в обратном порядке: начинается на брюшной стороне тела и заканчивается на спинной.
То, что весной не было замечено характерных изменений в коже (пигментации мездры), привело к рождению гипотезы, согласно которой у землероек нет нормальной весенней линьки (роста нового волоса), а происходит так называемая редукция – обламывание конечных сегментов зимних волос по перетяжкам и переход части остевых волос в пуховые (Borowski, 1952). Эту гипотезу критиковали более поздние исследователи (Шварц, 1955; Попов, 1960; Stein, 1954), имевшие в своих коллекционных сборах экземпляры в стадии нормальной весенней линьки с темными пятнами на мездре и ростом нового волоса. Случаи же, когда зверек имел одновременно короткий и длинный волос на разных частях шкурки (например, длинный на брюшке и короткий на спине) с резкой границей между ними, но без пигментации на мездре, они рассматривали как перерыв в линьке. Позже, отказавшись от гипотезы “редукции”, к этому пришел и Боровский (1968). Согласно его новым представлениям, волны короткого и длинного волоса дважды проходят по телу животного: один раз с брюшной стороны на спинную и вскоре после этого в обратном направлении – со спины на брюшко. В свете этих данных нетрудно примирить упомянутые выше высказывания относительно направления весенней линьки. Попов и Скарен наблюдали первую фазу весенней линьки, а Деннель, Кроукрофт и другие авторы – вторую фазу.
В более поздних обстоятельных работах Боровского (1968, 1973), получивших затем подтверждение в исследованиях ряда других зоологов (Ивантер, 1972, 2021; Докучаев, 1981; Stein, 1960), было показано, что у землероек-бурозубок весной бывает две следующие друг за другом линьки, различные по характеру, срокам и направлению, в котором они протекают. Весенняя линька I (ВЛ-I) заключается в смене шестисегментного зимнего волоса на пятисегментный весенний и проходит с брюшной стороны на спинную (рис. 2). В процессе весенней линьки II (ВЛ-II) этот пятисегментный весенний волос заменяется четырехсегментным летним. Начинается она на спине и заканчивается на брюшке. Эта линька может захватывать большую часть или всю шкурку животного (“полная линька”, по терминологии Боровского) или проходить в пределах узкой (1–5 мм шириной), постепенно перемещающейся полосы на шкурке (“волновая линька”). Кроме того, часто наблюдаются интервалы (перерывы) в линьке, и тогда землеройка может иметь одновременно длинный волос на одной части тела и короткий – на другой, без пигментации мездры. Такая “прерванная” линька наблюдается во время ВЛ-I у 40% особей, а ВЛ-II – у 22%.
Таким образом, рассмотренная выше картина двухэтапной весенней линьки перезимовавших зверьков по срокам и направлению соответствует ВЛ-I и ВЛ-II, описанным Боровским. Различна лишь трактовка этого явления. По Боровскому, у каждого зимовавшего зверька бывают две следующие друг за другом, но разграниченные во времени, весенние линьки I и II, при которых смена волос идет в противоположном направлении: сначала от живота к спине, а затем от спины к животу. Мы же считаем, что каждому зверьку свойственна лишь одна обычная (непрерывная) весенняя линька, а несовпадение ее сроков и хода у отдельных особей объясняется принадлежностью их к различным возрастным генерациям. Первыми начинают линьку пережившие зиму зверьки немногочисленных в наших условиях прошлогодних весенних пометов, то есть более старшие по возрасту. Они и претерпевают мнимую ВЛ-I. Что же касается второго этапа весенней линьки (по Боровскому – это ВЛ-II), то он соответствует массовой линьке особей поздних (летних) генераций. Иначе говоря, зимовавшие бурозубки вступают в весеннюю линьку в зависимости от своего календарного возраста как бы поэтапно. Это и создает ложное впечатление о существовании у одного и того же зверька двух последовательных весенних линек. К этой точке зрения приходят и некоторые другие исследователи, например Хивяринен с соавт. (Hyvärinen et al., 1971).
И все же требует пояснения сравнительно небольшая доля линяющих особей в наших весенних сборах (за апрель–май – всего 21.8%). Причин здесь несколько. Во-первых, это частично обусловлено большой индивидуальной скоростью линьки. Специальные наблюдения показали, что землеройка может сменить весь свой шерстный покров за 2–5 дней (Wilke, 1938; Borowski, 1963). Таким образом, вероятность отлова линяющих особей сравнительно мала. Во-вторых, весенняя линька зверьков, как показали наблюдения польских зоологов за зверьками, содержавшимися в неволе, и в самом деле может перемежаться паузами (по Боровскому, это так называемая прерванная линька). Мездра в этих случаях непигментированная, хотя смена волосяного покрова еще не закончилась, о чем говорит наличие зимних и летних волос на одной и той же шкурке. Такие зверьки тоже не попадают в категорию линяющих. И, наконец, относительно небольшая доля линяющих особей в весенних уловах может быть связана с их пониженной активностью. Передвижения землероек с утолщенной в 4–5 раз в результате линьки кожей, по-видимому, более ограничены, поэтому вероятность их отлова уменьшается (Borowski, 1968).
В результате весенней линьки, как уже указывалось, происходит смена зимних волос (шестисегментных остевых I порядка, семисегментных остевых II порядка и семисегментных пуховых) летними: трехсегментными остевыми I порядка, четырехсегментными остевыми II порядка и четырехсегментными пуховыми. Соответственно (с удлиненных на более короткие) изменяется и длина волос. К концу мая весенняя линька у землероек-бурозубок заканчивается, и зверьки добываются уже без пигментации на мездре, с коротким летним мехом (табл. 1). В последний раз зверьки с признаками весенней линьки пойманы 4.06.1967 г. и 8.06.1998 г. Со второй декады июня по середину сентября шерстный покров землероек-бурозубок, по-видимому, находится в состоянии покоя, о чем свидетельствует тонкая, светлая мездра, отсутствие линных пятен и прекращение роста волос всех категорий. На зрелость волоса указывают луковицы – они закрытого типа.
По поводу осенней линьки землероек мнения различных исследователей в общем довольно близки (Огнев, 1928; Формозов, 1948; Ивантер, 1975; Докучаев, 1981). Все они сходятся на том, что она проходит в более узкие, чем весной, сроки и начинается на спине, близ основания хвоста, распространяется вперед к голове, а затем переходит на брюшко (рис. 2). Менее единодушны они в вопросе о так называемой промежуточной линьке. Например, согласно Штейну (Stein, 1954), небольшая часть популяции бурозубок, помимо нормальных весенней и осенней линек, проходит еще три: одну – в их первое лето, другую – во второе и последнюю (третью промежуточную) – незадолго до смерти, осенью (“старческая линька”). В отношении зимовавших особей существование старческой линьки, продолжающейся с мая по ноябрь, подтвердили исследования Боровского (Borowski, 1964). В то же время Кроукрофт (1957) полагает, что промежуточная летняя линька представляет собой задержавшуюся весеннюю или рано начавшуюся осеннюю. С этим согласен и Скарен (Skaren, 1964), такова и наша позиция.
Нормальной осенней линьки у зимовавших бурозубок не наблюдается, однако у них бывает так называемая старческая (“промежуточная” или “остаточная”) летне-осенняя линька, заключающаяся в росте нового короткого летнего волоса на некоторых частях тела, большей частью по его дорсальной стороне (Borowski, 1964). Длится эта линька с июня по октябрь, причем у некоторых особей она может проявиться сразу после завершения нормальной весенней линьки, а у других – после большого перерыва. В соответствии с этим намечаются два пика интенсивности старческой линьки: в июне и в сентябре–октябре, когда доля линяющих животных достигает 30–60%; в остальные месяцы она обычно не превышает 25% (табл. 1).
На основании полученных нами данных вполне можно сделать вывод об отсутствии существенных видовых различий в сроках, интенсивности и топографии сезонных линек (рис. 3). Между тем связь с полом, возрастом и состоянием популяции выступает достаточно отчетливо. Установлено, в частности, что весенняя линька у размножающихся самок начинается несколько раньше, чем у самцов и не участвующих в репродукции самок. Осенняя линька прибылых зверьков у всех Soricidae происходит в близкие сроки (сентябрь–октябрь) и заключается в смене коротких летних волос на более длинные и густые. Появлению новых волос предшествуют морфообразовательные процессы в коже (разрыхление, утолщение, пигментация). Начинаются они обычно на спине у огузка, далее распространяются вперед к голове, затем переходят на бока и заканчиваются на брюшке.
Рис. 3.
Закладка и рост волос при сезонной линьке: (а) – появление пигмента; (б) – образование луковиц новых волос; (в) – растущие волосы в толще кожи (микрофото. ув. × 600).

Весной, в апреле–мае линяют взрослые (перезимовавшие) особи. Смена волос начинается у них на брюшной стороне тела с постепенным охватом боков, а завершается – на спине или голове. Двухэтапный характер весенней линьки с противоположным направлением смены волос (у одних зверьков она идет от живота к спине, а у других – от спины к животу) мы, в отличие от Боровского (Borowski, 1968, 1973), объясняем не существованием двух весенних линек, а неодновременным вступлением в линьку разных возрастных генераций. Первыми начинают линять особи прошлогодних весенних пометов, то есть более старшие по возрасту. Они и образуют мнимую ВЛ-I с характерным вентродорсальным направлением процесса. Что же касается второго этапа весенней линьки (по Боровскому, это ВЛ-II), то он соответствует массовой линьке зверьков поздних (летних) генераций и имеет дорсовентральтный порядок смены волос. Настоящая осенняя линька у этих животных, видимо, вообще отсутствует. Вместо этого у них наблюдается старческая линька, затрагивающая, как правило, лишь отдельные участки и не имеющая четкого рисунка.
Напрашивается вывод о том, что у землероек-бурозубок любая сезонная линька – будь то весенняя или осенняя, если она первая в жизни зверька, начинается на спинной стороне тела, а если вторая – на брюшной. К отрицанию двух весенних линек приходят и финские исследователи (Hyvärinen et al., 1971). Таким образом, землеройки-бурозубки претерпевают в условиях Севера две нормальные сезонные линьки (весеннюю и осеннюю), а также старческую. У водяной куторы, помимо этого, имеется ювенильная линька, а у крота – компенсаторная.
Линьке мышевидных грызунов, особенно представителям родов Clethrionomys и Microtus (рыжие и серые полевки), в отличие от Soricidae, посвящена сравнительно более широкая литература (Крыльцов, 1958, 1959, 1962; Марвин, 1974; Громов, Поляков, 1977; Stein, 1938; Lehmann, 1962; Mazak, 1962; Zejda, Mazak, 1965; Khateeb, Johnson, 1971a, 1971b, 1971c; Rowsemitt et al., 1975; Viro, 1979 и др.).
На основе изучения массовых видов грызунов Казахстана А. И. Крыльцов приходит к выводу об исключительной устойчивости и однообразии в последовательности смены волос у всех полевок Старого Света, которая почти не зависит от образа жизни зверьков. У обитателей суходольных и заболоченных лесов и лугов – рыжих, красных и темных (пашенных) полевок и полевок-экономок, у типичных полупустынных форм – общественных полевок, у полуводных – водяных крыс и ондатр, даже у таких специализированных подземных грызунов, как слепушонки, наблюдается один и тот же свойственный большинству изученных видов ход смены волос. Происходит она по сублатеральному (дорсальному) типу, при котором новые волосы появляются сначала на нижних частях боков и головы, затем процесс распространяется на брющко и спину, и в последнюю очередь вылинивают верх головы и задняя часть спины. В общих чертах сублатеральный тип подроста волос сохраняется при всех видах возрастных и сезонных линек, варьирует только последовательность и скорость вылинивания головы, средней и задней части спины. Лишь у некоторых эндемичных для Сибири и Казахстана представителей рода Clethrionomys все или часть особей вида в период одной из сезонных линек сменяют мех по цефалосакральному типу. Порядок смены волос в этом случае – обратный описанному: начинается двумя овальными пятнами на задней части спины, затем переходит на голову и завершается на боках и брюшке. Старые зверьки у всех видов имеют диффузный тип линьки, при котором не наблюдается какой-либо закономерной последовательности в ее топографии (Крыльцов, 1962; Марвин, 1974; Громов, Поляков, 1977; Lehmann, 1962; Zejda, Mazak, 1965; Quay, 1968; Ling, 1970; Viro, 1979).
Проведенные нами исследования в общем подтверждают выводы названных выше авторов. Линька изученных нами грызунов близка по срокам и проходит по аналогичному плану (рис. 2, 4, табл. 2). Для всех полевок установлено существование трех линек: ювенильной, которая в зависимости от времени рождения зверька может проходить весной, летом и осенью и завершается сменой детского волосяного покрова взрослым, и двух сезонных – весенней и осенней, сопровождающихся полной сменой волосяного покрова – соответственно на летний и зимний. Лесная мышовка, как, вероятно, и прочие зимоспящие млекопитающие, линяет почти весь бесснежный период с мая по октябрь, при этом линька, по-видимому, протекает диффузно, во всяком случае, закономерный характер в смене волос установить у нее не удается.
Рис. 4.
Последовательность появления линных пятен у мелких грызунов. (а) – рыжая полевка; (б) – полевка-экономка; 1 – ювенильная линька, 2 – весенняя линька, 3 – осенняя линька.

Таблица 2.
Встречаемость линяющих особей и интенсивность линьки рыжей полевки по месяцам
| Месяц | Самцы | Самки | ||||
|---|---|---|---|---|---|---|
| число исследованных зверьков | из них линяющих, % | относительная площадь пигментированной шкурки, % от площади всей шкурки | число исследованных зверьков | из них линяющих, % | относительная площадь пигментированной шкурки, % от площади всей шкурки | |
| Прибылые ранних выводков | ||||||
| VI | 28 | 82.1 | 31.6 | 17 | 76.4 | 54.1 |
| VII | 141 | 91.4 | 33.1 | 77 | 69.0 | 56.4 |
| VIII | 55 | 87.3 | 15.7 | 35 | 65.7 | 41.0 |
| IX | 4 | 100.0 | 23.0 | 3 | 100.0 | 16.3 |
| Прибылые поздних выводков | ||||||
| VII | 56 | 62.5 | 45.0 | 47 | 29.8 | 29.0 |
| VIII | 77 | 66.2 | 47.8 | 60 | 68.3 | 56.6 |
| IX | 23 | 73.9 | 53.5 | 14 | 79.6 | 44.0 |
| X | 51 | 76.5 | 23.8 | 44 | 86.3 | 27.4 |
| XI– XII | 5 | – | – | 5 | – | – |
| Зимовавшие | ||||||
| III | 3 | – | – | 2 | – | – |
| IV | 3 | 64.7 | 16.0 | 3 | 66.7 | 10.0 |
| V | 27 | 66.7 | 4.9 | 11 | 54.5 | 22.5 |
| VI | 20 | 85.0 | 12.1 | 11 | 45.4 | 35.8 |
| VII | 46 | 65.2 | 8.0 | 27 | 22.2 | 6.4 |
| VIII | 15 | 93.3 | 9.3 | 5 | 80.0 | 10.0 |
Осенняя линька у всех грызунов проходит обычно более интенсивно, чем весенняя, сроки которой из-за неоднородности популяции в возрастном отношении крайне растянуты. Сроки и скорость линек зависят также от пола и физиологического состояния зверьков. Так, линька лактирующих самок запаздывает по сравнению с самками без признаков размножения, но начинается на 2–3 нед. раньше, чем у самцов. Ювенильная линька молодых поздних выводков обычно проходит быстрее, чем ранних, и тем не менее может переходить в осеннюю. Существенные коррективы в общий ход, темпы и порядок сезонной линьки вносят климатические условия года и состояние популяции (уровень численности и фаза популяционного цикла).
Наиболее четкая картина сезонных смен шерстного покрова вырисовывается на нашем материале для рыжей полевки, по которой удалось собрать наиболее богатый фактический материал. При этом сама шерсть этого грызуна значительно отличается от описанного выше волосяного покрова землероек и лесной мышовки не только густотой и линейными размерами волос, но и формой волосяного стержня, его микроскопическим строением, а также групповым расположением волос на шкурке. Отсутствует характерная для насекомоядных сегментация волос. Стержни волос расположены наклонно к поверхности кожи и образуют отчетливые “потоки”, направленные к хвосту. Волосы растут пучками, состоящими из 2–4 пуховых или одного остевого и 2–3 пуховых волос, реже встречаются пучки из одних остевых. Вокруг каждого направляющего волоса группируются по 2–3 таких пучка, образуя сложные комплексы с различным сочетанием волос всех категорий. Мех – относительно длинный и густой. Окраска его яркая с заметным преобладанием темных, ржаво-коричневых и рыже-красных тонов на спине, рыже-серых на боках и светло-серых на брюшке, причем переходы в окраске различных частей тела постепенные.
По данным ряда авторов (Zejda, Mazak, 1965; Viro, Koskela, 1978; Viro, 1979), у рыжей полевки в течение жизни шерстный покров сменяется несколько раз. Первая ювенильная линька начинается в возрасте 5 недель. Вскоре после нее проходит постювенильная линька, в процессе которой редкий и короткий серовато-коричневый волос сменяется на летний у прибылых, родившихся весной и в начале лета, или на зимний – у родившихся в конце лета и осенью. В дальнейшем регулярная смена меха происходит весной и осенью. Она тесно связана с экологическими и внутренними факторами, половой активностью, беременностью и лактацией.
Наши исследования в целом подтверждают эти сведения (табл. 2, рис. 3). В условиях Карелии первую возрастную (ювенильную) линьку рыжие полевки претерпевают в полуторамесячном возрасте. Она связана, по-видимому с выходом из гнезд и переходом к более активному наземному образу жизни, в процессе которого зверьки подвергаются воздействию новых, более изменчивых экологических условий. Смена волос происходит почти одновременно по всей шкурке, но все-таки на вентральной стороне она начинается раньше. Согласно принятой номенклатуре (Крыльцов, 1962; Марвин, 1974; Lehmann, 1962; Ling, 1970; Viro, 1979), это так называемая дорсальная, или сублатеральная линька. Общая продолжительность ювенильной линьки не более 10 дней, и в результате у прибылых зверьков ранних выводков появляется постювенильный летний волосяной покров, а у поздних – взрослый летний или даже зимний.
Многие авторы связывают начало ювенильной линьки у полевок с наступлением половой зрелости (Ченцова, 1949; Огнев, 1950; Тупикова, 1971 и др.). Однако мы склонны разделить мнение А. И. Крыльцова (1958), который, опровергает эту точку зрения и приводит примеры возрастной (ювенильной) и даже одной из двух сезонных линек, наблюдаемых у зверьков задолго до их полового созревания.
После ювенильной линьки следует постювенильная (в процессе нее полевки весеннего и ранне-летнего поколения приобретают взрослый летний, а осенью – зимний волос), а затем сезонные – осенняя и весенняя (будущего года). При этом у прибылых самцов и самок ранних выводков ювенильная, постювенильная и осенняя линьки сменяют одна другую практически без перерыва, так что разграничить их сроки не удается. То же можно сказать и о прибылых поздних выводках. В конце октября молодые полевки всех возрастов заканчивают линьку и в ноябре носят уже зимний наряд.
У зимовавших самцов весенняя линька, судя по изменению числа линяющих и интенсивности смены меха, очень растянута и проходит с середины апреля по конец июня, а разгар ее (линяет 65–100% зверьков со средней интенсивностью 5–25%) – с третьей декады мая по вторую декаду июня. У самок линька начинается и проходит на 2–3 недели раньше – с начала апреля по май (67–100% линяющих с интенсивностью 10–52%), но захватывает и первую половину июня. В дальнейшем (в конце июня–июле) число линяющих среди зимовавших полевок сокращается, а затем, в августе, снова увеличивается, достигая 80–100%. Возможно, в конце лета у них начинается осенняя линька, но проверить это трудно, так как большинство зимовавших зверьков к сентябрю погибает.
В мае чаще всего попадались зверьки с темной мездрой на брюшке и боках и светлой – на спине. При этом подрост новых волос на брюшке составлял у них 2 мм, на боках – 1–1.8, а на спине новых волос еще не было и даже пигментация мездры обнаруживалась только при микроскопировании. Это свидетельствует о том, что весенняя линька начинается на вентральной стороне, а заканчивается – на дорсальной (рис. 3). У зимовавших самцов первыми весной линяют бока и конечности, затем сменяется шерсть на спине, брюшке и огузке. В июле линяют главным образом бока, а в августе – спина. У самок весенняя линька тоже начинается с боков, затем распространяется на брюшко и в последнюю очередь – на спину и голову. В июле у них линяют бока и плечи, а в августе – отдельные участки на брюшке и боках. Летняя смена волос у прибылых самцов ранних выводков начинается с боков, после чего распространяется на голову, плечи, спину и живот. У самок этой возрастной группы во время летней (ювенильной и постювенильной) линьки первым сменяется волос на брюшке и боках, а затем на остальной части тела, кроме спины, вылинивающей позднее.
Осенняя линька прибылых и зимовавших зверьков сопровождается полной сменой волосяного покрова и растянута с конца августа по октябрь включительно (табл. 2). Выпадение старых волос в этот период почти незаметно, так как идет медленно и начинается лишь после того, как длина новых волос достигнет уровня старых. Подрост зимнего меха при осенней линьке у половозрелых и неполовозрелых прибылых полевок начинается на боках с почти одновременным охватом спины. Затем линное пятно, распространяясь к голове, переходит на брюшную сторону и заканчивается в паховой области. Таким образом, последовательность осенней линьки прибылых особей в общих чертах соответствует порядку появления волос у новорожденных и относится к типичной для рыжих полевок дорсальной схеме. В то же время у прибылых самцов поздних выводков ювенильная смена шерстного покрова начинается нередко с боков и брюшка и заканчивается на спине, а осенняя – с огузка и отсюда постепенно переходит на остальную часть спины, голову, плечи, брюшко и бока (по цефальному и каудальному типу). У старых зверьков, родившихся в конце лета или осенью предыдущего года, подрост волос происходит ограниченными участками на различных частях тела и более всего подходит к диффузному или лоскутному типам линьки. Отсутствие четкой картины в последовательности осенней линьки можно объяснить варьированием ее сроков и растянутостью у отдельных зверьков (исходя из продолжительности линьки одной особи равной 15–18 дней), в результате чего разные топографические стадии смены волос накладываются друг на друга и затушевывают закономерный порядок линьки.
Общеизвестно, что одна из основных целей сезонных линек волосяного покрова млекопитающих заключается в адаптивных изменениях его свойств по сезонам, приводящим к соответствующей смене качеств меха и прежде всего его теплопроводности по сезонам. Это подтверждают и выполненные нами ранее исследования на мелких млекопитающих (Ивантер и др., 1985). Согласно этим данным, плотность волосяного покрова обыкновенной и других видов изученных бурозубок при смене на летний изменяется в основном за счет пуховых при сравнительно одинаковом количестве остевых и направляющих. Наблюдающееся же на протяжении всей жизни зверька закономерное изменение густоты волос при сохранении постоянного числа волосяных фолликулов Боровский (1958) связывает с эффектом Денеля – изменением массы и общей поверхности тела животных по сезонам (Dehnel, 1949).
В результате осенней линьки волосы становятся гуще, тоньше и почти в 2 раза длиннее. В то же время относительная толщина сердцевинного канала от лета к зиме заметно увеличивается: от 50–63% в бесснежный период до 62–84% зимой. При этом волосяной покров зверьков, отловленных летом, значительно ниже, чем у зимних. Пуховые волосы, входящие в состав летнего меха, – четырехсегментные, зимнего – семисегментные; остевые соответственно трех- и шестисегментные. В общем зимний мех выше летнего в 1.8–2 раза, причем длина концевого сегмента изменяется незначительно. Это относится и к другим категориям волос. При этом сезонные изменения высоты волосяного покрова связаны не только с изменением числа сегментов, но и их длины.
Мех водяной куторы достаточно густой, чтобы защитить ее от переохлаждения как в воде, так и на суше, но обращает внимание относительная его изреженность на брюшке, что, по-видимому, компенсируется большей толщиной растущих здесь волос. Впрочем, Б.Ф. Церевитинов (1958), упоминая, что у некоторых из представителей земноводных млекопитающих волосы на спине все-таки гуще, чем на других частях туловища, и объясняет это разной степенью приспособления к водным условиям. Вероятно, к таким “некоторым” следует отнести и кутору. По сезонно густота волос у этого вида изменяется за счет пуховых. Наконец, характерным для куторы следует считать то, что в отличие от бурозубок, у которых остевые почти равны по толщине направляющим, у куторы они почти вдвое тоньше направляющих.
Сопоставление результатов наших измерений с данными В.Е. Соколова (1973) для кутор из разных регионов выявляет достаточно четкие географические различия рассматриваемого показателя. Согласно этим материалам (Ивантер и др., 1985), мех кутор Карелии состоит из наиболее длинных и соответственно наиболее тонких волос, что особенно важно для животным, обитающих у северных границ видового ареала в условиях постоянных термических дефицитов. Данные же по длине волос куторы показывают, что по крайней мере в летний период волосяной покров у нее значительно выше (“пышнее”), чем у землероек-бурозубок. В общем же у куторы длина волос всех категорий постепенно уменьшается с крестца и боков до спины и брюшка, что по номенклатуре Б.Ф. Церевитинова, соответствует эквалатеральному типу топографии волосяного покрова, характерному для большинства полуводных (“земноводных” – по терминологии автора) млекопитающих. У мелких грызунов к отмеченным выше особенностям сезонных линек следует добавить их растянутость и зависящую от участия в размножении четкую дифференцированность по возрастным генерациям.
Теплозащитные свойства покровов, их изменения в процессе сезонной линьки и механизмы терморегуляции
В процессе эволюции и освоения среды обитания мелкие млекопитающие таежного Севера приобрели целый ряд адаптаций, направленных в первую очередь на наиболее целесообразное, с наименьшими энергетическими затратами, поддержание теплового баланса организма со средой. Поскольку сохранение тепла за счет физической терморегуляции не требует дополнительных пищевых ресурсов, оно является основным способом экономии энергетических затрат (Jansky, 1967). В связи с этим эволюция терморегуляции идет в направлении замены химических механизмов физическими (Слоним, 1952, 1964, 1971; Шилов, 1962), о чем говорит и более отчетливая выраженность физической терморегуляции у типично субарктических и высокогорных видов (Шварц и др., 1968; Большаков, 1972).
Как известно, в системе приспособления млекопитающих к пониженным температурам важное место занимают реакции волосяного покрова. Многочисленными исследованиями показано, что теплоизоляция пропорциональна густоте и толщине мехового покрова, и у мелких млекопитающих она ниже, чем у и более крупных (Калабухов, 1951, 1969; Слоним, 1952, 1971; Марвин, 1966; Большаков, Некрасов, 1968; Кривошеев и др., 1975; Давыдов, 1982; Hart, Heroux, 1955; Irving, 1964; Morrison, 1964 и др.). Чтобы иметь возможность успешно передвигаться, мелкие животные должны обладать тонким и легким волосяным покровом, а то, что шерсть у них по необходимости короче, снижает теплоизоляционные свойства покровов (Шмидт-Ниельсен, 1982).
Согласно современным представлениям, теплозащитные качества волосяного покрова зависят главным образом от образующегося в его толще “неподвижного” и “инертного” воздуха, исключающего конвекционные токи и создающего эффективную тепловую изоляцию (Соколов, 1973). Чем пышнее и выше мех и тоньше волосы, тем больше воздушная прослойка между волосками и, следовательно, лучше сохраняется тепло. Имеет значение и строение волос – развитие сердцевины, содержащей теплоизолирующий воздух, наличие продольных бороздок на стержне, его конфигурация, число сегментов, способ укладки и т.п.
Связь теплоизоляционных свойств меха с морфологией и общим состоянием наружных покровов млекопитающих подтверждается экспериментальным определением коэффициента теплопроводности сухих коллекционных шкурок зверьков, добытых в Карелии в разные сезоны года (табл. 3). Согласно этим данным, интегральная теплопроводность шерсти имеет достаточно четкие таксономические и сезонные различия, причем последние приобретаются в процессе сезонных линек.
Таблица 3.
Коэффициент теплопроводности (10–3 Вт/м К) сухих шкурок мелких млекопитающих Карелии
| Вид | Сезон | n | Пределы | M ± m |
|---|---|---|---|---|
| Обыкновенная бурозубка | Лето | 16 | 38.2–50.0 | 47.6 ± 0.8 |
| Зима | 15 | 35.0–46.4 | 37.4 ± 0.6 | |
| Осень | 12 | 33.1–41.2 | 34.7 ± 0.5 | |
| Средняя бурозубка | Лето | 16 | 50.0–60.0 | 54.0 ± 1.2 |
| Зима | 15 | 38.3–42.6 | 40.1 ± 0.5 | |
| Осень | 13 | 37.1–44.8 | 39.4 ± 0.3 | |
| Малая бурозубка | Лето | 15 | 49.0–57.2 | 53.5 ± 0.9 |
| Зима | 14 | 38.0–49.0 | 42.2 ± 0.5 | |
| Осень | 9 | 36.0–45.0 | 39.0 ± 0.4 | |
| Обыкновенная кутора | Лето | 18 | 30.0–42.1 | 32.5 ± 0.9 |
| Осень | 14 | 28.0–39.0 | 30.3 ± 0.8 | |
| Обыкновенный крот | Лето | 9 | 24.2–30.0 | 26.7 ± 0.3 |
| Зима | 8 | 19.0–22.4 | 20.5 ± 0.3 | |
| Осень | 8 | 18.9–21.0 | 19.1 ± 0.3 | |
| Лесная мышовка | Лето | 22 | 48.0–50.3 | 48.6 ± 0.3 |
| Осень | 4 | 37.9–38.2 | 38.1 ± 0.4 | |
| Рыжая полевка | Лето | 20 | 45.2–53.1 | 49.8 ± 0.9 |
| Зима | 16 | 37.0–50.7 | 41.0 ± 0.8 | |
| Осень | 14 | 36.1–42.1 | 38.7 ± 0.8 | |
| Полевка-экономка | Лето | 11 | 46.7–53.2 | 47.8 ± 0.4 |
| Зима | 5 | 34.1–38.4 | 36.5 ± 0.4 | |
| Осень | 8 | 34.1–40.9 | 36.1 ± 0.6 |
Волосяной покров насекомоядных млекопитающих значительно уступает шерсти грызунов по высоте, но превосходит его по плотности. Кроме того, характерное для насекомоядных сегментированное строение волос, предотвращая сминание и обеспечивая укладку волос в любом направлении, способствует удержанию в нем неподвижного воздуха, что положительно сказывается на теплоизолирующих свойствах шкурки в целом и в значительной мере компенсирует общее несовершенство структуры их волосяного покрова.
Хорошо известно, что структура волосяного покрова млекопитающих зависит от сезона года: зимний мех гуще и длиннее и, как правило, лучше сохраняет тепло, чем более легкий (редкий и короткий) летний мех. Это сказывается и на уровне энергетического обмена. Тем не менее более совершенная терморегуляция организма “зимних” зверьков определяет специфику их адаптации к холоду: последняя обычно сопровождается не адекватным повышением теплопродукции, а значительным ограничением теплоотдачи (Слоним, 1964; Смирнов, 1968). Таким образом, улучшение теплоизоляционных свойств покровов в результате осенней линьки и обусловленное наступающими холодами снижение химической терморегуляции представляют собой общую закономерность (Слоним, 1961; Марвин, 1966; Смирнов, 1968; Калабухов, 1969; Башенина, 1966, 1977; Scholander et al., 1950; Hart, Heroux, 1953; Irving et al., 1955; Jung, 1964). И хотя это больше свойственно крупным животным, улучшение теплозащитных свойств зимнего меха и соответствующие сезонные изменения уровня теплоотдачи отмечены и у мелких млекопитающих (Лещинская, 1952; Волчанецкая, 1954; Смирнов, 1968; Калабухов, 1969; Башенина, 1977; Hammel, 1955; Irving, Krogh, 1955; Hart, 1956; Gębczyński, Olszewsky, 1963; Viro, 1979 и др.).
Согласно нашим измерениям (табл. 3), коэффициент теплопроводности летнего волосяного покрова по отношению к зимнему увеличивается (а теплоизоляция снижается) у обыкновенной бурозубки на 28.2%, средней – на 34.6, малой – на 26.8, у крота – на 30.2, у рыжей полевки – на 21.5, у полевки-экономки – на 30.9%. Это соответствует отмеченным выше сезонным изменениям длины и густоты меха.
Показатель ранговой корреляции Спирмена (rs) между длиной волос и удельной теплопроводностью шкурки составил (по всему материалу) –0.41 (t = 2.3; p > 0.95), что говорит о средней по силе, но статистически вполне достоверной связи. Еще теснее зависимость коэффициента теплопроводности от густоты меха: rs = –0.66 (при t = 3.6; p > 0.99). Коэффициент общей (совокупной) корреляции, оценивающий связь теплозащитных свойств шкурки с длиной и густотой волос вместе, составил 0.90 (р > 0.999).
Таким образом, проведенные исследования убедительно показывают, что шкурки мелких зверьков в разгар линьки не только не уступают зимним по термозащитным свойствам, но даже превосходят их, о чем свидетельствует максимально низкий коэффициент теплопроводности (табл. 3). Как уже указывалось, это объясняется характерным для линьки значительным утолщением кожи и увеличением густоты меха за счет совмещения не выпавших, еще старых волос, с подрастающими новыми. Таким образом, вопреки распространенному мнению во время линьки, по крайней мере у мелких млекопитающих, механизмы физической терморегуляции не только не утрачивают своего значения, а, напротив, служат важным звеном в цепи гомеостатических реакций, направленных на снижение непроизводительных трат энергии и потери тепла в сложные для жизни зверков переходные периоды. Более глубокая, чем в другие сезоны, реакция газообмена на воздействие холода указывает на то, что осенью грызуны и землеройки еще сохраняют определенные черты физиологической адаптации высоким летним температурам, и воздействие низких температур, усиленных повышенной влажностью воздуха, вызывает у них обостренную реакцию химической терморегуляции (Смирнов, 1968). Как считает сам П.К. Смирнов, такая реакция требует повышенных затрат энергии на избыточную теплопродукцию, необходимую для покрытия увеличенной потери тепла и, следовательно, для поддержания теплового баланса на уровне, обеспечивающем нормальную жизнедеятельность. Улучшение же теплоизоляционных свойств наружных покровов в период линьки в значительной мере компенсирует несовершенство химической терморегуляции и служит важным приспособлением для более рационального теплообмена организма при его физиологической перестройке в условиях неустойчивого температурного режима среды.
ЗАКЛЮЧЕНИЕ
Выполненное исследование посвящено мелким лесным млекопитающим – одной из наиболее типичных экологических групп, представляющих норно-наземных млекопитающих, или, как их еще называют, полунорников, мелких зверьков, обитающих в лесной подстилке и ходах неглубоких нор, но нередко появляющихся и на поверхности почвы, то есть живущих здесь в условиях низких и весьма непостоянных температур. По образу жизни, условиям среды, а значит и по морфофизиологическим свойствам они совмещают в себе многие черты, характерные как для настоящих наземных, так и для специализированных подземных млекопитающих, и по существу представляют собой переходный адаптивный тип. В числе его важнейших адаптивных свойств – достаточно совершенная химическая терморегуляция, как известно, начинающаяся у гомойотермных животных лишь после того как исчерпываются возможности физической терморегуляции. Отсюда и особое значение, которое приобретает изучение особенностей механизмов физической терморегуляции у мелких млекопитающих – зверьков, характеризующихся высокой энергетикой при низкой индивидуальной стойкости организма к условиям внешней среды.
Обитая в умеренной и особенно холодной климатических зонах, в условиях низких и весьма изменчивых температур, эти животные приобрели легкий, не мешающий движениям, но достаточно теплый с высокими термоизоляционными свойствами волосяной покров, характеризующийся умеренной густотой и длиной волос, неравномерностью опушения отдельных частей тела, а у землероек – еще и булавовидной удлиненной апикальной гранной и более сильным, чем у других групп животных, развитием сердцевинного канала. Тем не менее у мелких млекопитающих, как у животных с высоким потреблением энергии и недостаточно развитыми механизмами физической терморегуляции, сохранение температурного гомеостазиса достигается в основном интенсификацией обменных процессов. Особенно велико значение химической терморегуляции для широко распространенных видов мелких млекопитающих, ареалы которых охватывают несколько ландшафтных зон (обыкновенная бурозубка, рыжая полевка). Между тем у животных-автохтонов Субарктики приспособления направлены на уменьшение энергетических затрат и связаны с хорошим развитием теплоизолирующих свойств наружных покровов (темная полевка, лесной и тундровый лемминги). Наконец у зимоспящих грызунов, то есть не нуждающихся зимой в эффективных механизмах физической теплорегуляции, теплозащитная роль покровов в этот период сводится к нулю.
В этой связи в условиях Субарктики совершенствование энергетически выгодных и биологически целесообразных механизмов физической терморегуляции оказалось одним из основных направлений в эволюции адаптаций гомойотермных животных. Однако достигалось это разными путями. Выявленную многими исследователями высокую устойчивость северных млекопитающих к холодовым нагрузкам (отсюда крайне нижнее положение “критической точки” обмена), их способность длительное время поддерживать нормальную температуру тела при низких температурах без дополнительных энергетических затрат нельзя, конечно, объяснить одним лишь повышением теплоизоляционных свойств покровов. Не меньшую роль, очевидно, играют и биохимические адаптации, заключающиеся в способности поверхностных тканей организма к нормальному обмену при температурах, значительно более низких, чем сердцевины тела. Эффективность физической терморегуляции в качестве одного из важнейших компонентов адаптивного теплообмена продемонстрирована в процессе предпринятого нами исследования структуры и теплозащитных свойств покровов различных видов мелких насекомоядных и грызунов Карелии. Показано, в частности, что вопреки распространенному мнению во время линьки механизмы физической терморегуляции не только не утрачивают своего значения, а, напротив, служат важным звеном в цепи гомеостатических реакций, направленных на снижение непроизводительных трат энергии и потери тепла в сложные для жизни зверьков переходные периоды. В свою очередь это приводит к характерным изменениям эколого-физиологических реакций – понижению уровня обмена, установлению и поддержанию постоянной температуры тела, сдвигам в деятельности эндокринной системы и т. д.
Список литературы
Башенина Н.В. О сезонных изменениях химической терморегуляции у полевок // Бюл. МОИП. Отд. биол. 1966. Т. 71. Вып. 3. С. 27–40.
Башенина Н.В. Адаптивные особенности теплообмена мышевидных грызунов. М.: Наука, 1977. 294 с.
Большаков В.Н. Пути приспособления мелких млекопитающих к горным условиям. М.: Наука, 1972. 100 с.
Большаков В.Н., Некрасов Е.С. О теплоизоляционных свойствах наружных покровов горных грызунов // Науч. докл. высш. школы. Сер. биол. наук. 1968. № 10. С. 23–27.
Волчанецкая Г.И. Сезонные изменения реакции некоторых видов полевок на влияние температуры среды // Уч. зап. Харьковск. ун-та. 1964. Вып. 52. С. 225–288.
Громов И.М., Поляков И.Я. Полевки (Мicrotinae). Л.: Наука, 1977. 504 с.
Давыдов А.Ф. Гомойотермные организмы бореальной климатической зоны // Экологическая физиология животных. Л.: Наука, 1982. Ч. 3. С. 110–130.
Депарма Н.К. Линька кротов, ее последовательность и сроки // Тр. Всесоюзн. научно-исслед. ин-та охотничьего хоз-ва. М., 1951. Вып. 10. С. 93–127.
Докучаев Н.Е. Материалы по линьке средней (Sorex caecutiens Laxm.) и крупнозубой (S. daphaenodon Thomas) бурозубок на северо-востоке Сибири // Экология млекопитающих Северо-Восточной Сибири. М.: Наука, 1981. С. 22–32.
Ивантер Т.В. Особенности линьки землероек Карелии // Науч. конф. биологов Карелии. Петрозаводск, 1972. С. 180–182.
Ивантер Э.В. Очерки популяционной экологии мелких млекопитающих на северной периферии ареала. М.: Тов-во науч. изд. КМК, 2018. 770 с. Ивантер Э.В. Строение, теплозащитные свойства и линька волосяного покрова обыкновенной бурозубки (Sorex araneus) // Зоол. журн. 2021. Т. 100. № 4. С. 419–433.
Ивантер Э.В., Ивантер Т.В., Туманов И.Л. Адаптивные особенности мелких млекопитающих. Л.: Наука, 1985. 318 с.
Калабухов Н.И. Методика экспериментальных исследований по экологии наземных позвоночных. М.: Наука, 1951. 177 с.
Калабухов Н.И. Периодические (сезонные и годичные) изменения в организме грызунов, их причины и последствия. Л.: Наука, 1969. 248 с.
Когтева Е.З. Сезонная изменчивость и возрастные особенности строения кожи и волосяного покрова крота, зайца-беляка и енотовидной собаки // Промысловая фауна и охотничье хозяйство Северо-Запада РСФСР. Л.: Госиздат, 1963. С. 213–271.
Кривошеев В.Г., Варич А.А., Уманцева Н.Д., Кривошеева В.П. Механизмы физической терморегуляции сибирского лемминга и субарктических популяций красной полевки и полевки-экономки // Мат. по экологии мелких млекопитающих Субарктики. Новосибирск: Наука, 1975. С. 110–144.
Крыльцов А.И. Материалы по линьке мышевидных грызунов. Сообщение 1. Линька массовых видов полевок северного Казахстана // Зоол. журн. 1958. Т. 37. Вып. 2. С. 271–286.
Крыльцов А.И. Материалы по линьке мышевидных грызунов. Сообщение 2. Линька различных представителей подсемейства полевок // Зоол. журн. 1959. Т. 38. Вып. 5. С. 745–755.
Крыльцов А.И. Топография линьки грызунов и возможность использования ее в качестве таксономического признака // Тр. Ин-та защиты раст. КазССР. Алма-Ата, 1962. Т. 7. С. 418–451.
Кузнецов Б.А. Товароведение пушно-мехового сырья СССР. М: Заготиздат, 1932. 464 с.
Кузнецов Б.А. Основы товароведения пушно-мехового сырья М.: Заготиздат, 1952. 508 с.
Лещинская Е.М. Сезонные изменения кожного покрова млекопитающих // Зоол. журн. 1952. Т. 31. Вып. 3. С. 745–751.
Марвин М.Я. Влияние внешней среды на строение меха млекопитающих // Уч. зап. Уральск. гос. ун-та. Сер. биол. Свердловск, 1966. Вып. 3. С. 52–57.
Марвин М.Я. Строение и линька волосяного покрова грызунов Урала // Науч. тр. Свердловск. гос. пед. ин-та, 1974. Сб. 235. С. 11–22.
Огнев С.И. Звери Восточной Европы и Северной Азии. Т. I. Насекомоядные и летучие мыши. М.-Л.: Главнаука, Госиздат, 1928. 632 с.
Огнев С.И. Звери СССР и прилежащих стран. М.-Л.: Академия Наук СССР, 1950. Т. 7. 706 с.
Павлова Е.А. Сезонные изменения волосяного покрова зайца-беляка и прогноз сроков выходности меха // Тр. Всесоюзн. научно-исслед. ин-та охотничьего хозяйства. 1951. Вып. 10. С. 59–77.
Попов В.А. Млекопитающие Волжско-Камского края. Казань: Госиздат, 1960. 466 с.
Слоним А.Д. Животная теплота и ее регуляция в организме млекопитающих. М.; Л.: Госиздат, 1952. 327 с.
Слоним А.Д. Основы общей экологической физиологии млекопитающих. М.; Л.: Биомедгиз, 1961. 432 с.
Слоним А.Д. О физиологических механизмах природных адаптаций животных и человека. М.; Л.: Госиздат, 1964. 64 с.
Слоним А.Д. Экологическая физиология животных. Учебное пособие. М.: Высшая школа, 1971. 448 с.
Смирнов П.К. Эколого-физиологические исследования некоторых видов грызунов. Л.: Изд. ЛГУ, 1968. 135 с.
Соколов В.Е. Кожный покров млекопитающих. М.: Наука, 1973. 487 с.
Соколов В.Е., Скурат Л.Н., Степанова Л.В. и др. Руководство по изучению кожного покрова млекопитающих. М.: Наука, 1986. 279 с.
Строганов С.У. Звери Сибири. Насекомоядные. М.: Изд. АН СССР, 1957. 267 с.
Тупикова Н.В. Методы определения возраста грызунов в целях изучения возрастного состава их популяции // Тез. докл. II экол. конф. Киев, 1950. Ч. 2. С. 113–115.
Фадеев Е.В. Строение меха нутрии // Тр. Всесоюзн. научно-исслед. ин-та охотн. хоз-ва. М.: Заготиздат, 1955. Вып. 15. С. 93–104.
Формозов А.Н. Мелкие грызуны и насекомоядные Шарьинского района Костромской области в период 1930–1940 гг. // Материалы по грызунам. М.: МОИП, 1948. Вып. 3. С. 3–110.
Церевитинов Б.Ф., Кирсанов И.К., Бигман В.В. Товароведение животного сырья. М.: Госиздат, 1948. 248 с.
Церевитинов Б.Ф. Топографические особенности волосяного покрова пушных зверей // Тр. ВНИИЖП. М., 1958. Вып. 17. С. 256–307.
Ченцова Н.Ю. Определение возраста полевок (Microtus gregalis Pall.) по пластическим признакам // Тр. ВИЗР. Л., 1949. Вып. 2. С. 56–68.
Чернова О.Ф., Целикова Т.Н. Атлас волос млекопитающих. М.: Тов-во науч. изданий КМК, 2004. 429 с.
Шварц С.С. Биология землероек лесостепного Зауралья // Зоол. журн. 1955. Т. 34. Вып. 3. С. 915–927.
Шварц С.С., Смирнов В.С., Добринский Л.Н. Метод морфофизиологических индикаторов в экологии наземных позвоночных. Свердловск: Госиздат, 1968. 387 с.
Шилов И.А. О некоторых понятиях, связанных с изучением проблемы терморегуляции // Зоол. журн. 1962. Т. 41. Вып. 4. С. 491–497.
Шмидт-Ниельсен К. Физиология животных. Приспособление и среда: В 2 кн. М.: Мир, 1982. 785 с.
Borowski S. Sezonowe zmiany u wlosienia i Soricidae // Ann. Univ. M. Curie-Sklod. Sect. C. 1952. V. 7. № 2. S. 65–117.
Borowski S. Variation in density of coat during the life cycle of Sorex araneus araneus Linnaeus, 1758 // Acta Theriol. 1958. V. 2. № 14. P. 286–289.
Borowski S. Old-age moult in the common shrew, Sorex araneus Linnaeus, 1758 // Acta Theriol. 1963. V. 7. № 19. P. 374–375.
Borowski S. Moult of shrews (Sorex L.) under laboratory conditions // Acta Theriol. 1964. V. 8. № 8. P. 125–135.
Borowski S. On the moult in the common shrew // Acta Theriol. 1968. V. 13. № 30. P. 483–498.
Borowski S. Variations in coat and colour in representatives of the genera Sorex L. and Neomys Kaup. // Acta Theriol. 1973. V. 18. № 14. P. 247–279.
Crowcroft P. The life of the shrew. L.: M. Reinhardt, 1957. 166 p.
Dehnel A. Studies in the genus Sorex L. // Ann. Univ. M. Curie-Sklod. Sect. C. 1949. V. 4. № 2. S. 17–102.
Dehnel A. Studies on the genus Neomys Kaup // Ann. Univ. M. Curie-Sklod. Sect. C. 1950. V. 5. № 1. S. 1–63.
Findley J.S., Jones J.K. Moult of the short-tailded shrew, Blarina brevicauda // Amer. Midl. Nat. 1956. V. 56. № 1. P. 246–249.
Gębczyński M., Olszewsi J. Katathermometric measurements of insulating properties of the fur in small mammals // Acta Theriol. 1963. V. 7. № 19. P. 369–371.
Hammel H.T. Termal properties of fur // Amer. J. Phisiol. 1955. V. 182. P. 369–376.
Hamilton W.J. The moult of Blarina brevicauda // J. Mammal. 1940. V. 21. № 4. P. 457–458.
Hart J.S. Seasonal changes in insulation of the fur // Canad. J. Zool. 1956. V. 34. № 1. P. 53–57.
Hart J.S., Heroux O. Exercise and temperature regulation in lemming and rabbits // Canad. J. Biochem. Phisiol. 1955. V. 33. № 3. P. 528–532.
Hyvärinen H. On the seasonal changes in the skeleton shrew (Sorex araneus L.) and their physiological background // Aquil. Ser. Zool. 1969. V. 7. P. 1–32.
Hyvärinen H., Pelttari A., Saure L. Seasonal changes in the histology and alkaline phosphatase distribution of the skin of the common shrew and of the bank vole as a function of hair cycle // Aquil. Ser. Zool. 1971. V. 12. P. 43–52.
Irving L. Terrestrial animals in cold: birds and mammals // Handbook of physiology. Sect. 4. Adaptation to environment. Washington, 1964. P. 361–378.
Irving L., Krog J. Temperature of skin in the arctic as a regulator of heat // J. Appl. Phisiol. 1955. V. 7. P. 355–364.
Irving L., Krogh H., Monson M. The metabolism of some alascan animals in winter and summer // Phisiology. 1955. V. 28. № 3. P. 173–185.
Jackson H.H.T. A taxonomic review of the American longtailed shrew // North Amer. Fauna. 1928. V. 51. P. 1–30.
Jansky L. Evolutionary adaptations of temperature regulation in mammals. Cond. Z, Saugetier–Kunde, 1967. B. 32. № 3. P. 167–172.
Jung K.D. Untersuchungen über Wärmestrahlungsmechanismen an Säugerintegumenten und ihre Biologische Bedeutung // Zool. Jahrb. Abt. I. 1964. B. 91. № 4. S. 56–74.
Khateeb A.A., Johnson E. Seasonal changes of pelage in vole (Microtus agrestis). I. Correlation with changes in the endocrine clands // Gen. Comp. Endocrinol. 1971a. V. 16. P. 217–228.
Khateeb A.A., Johnson E. Seasonal changes of pelage in the vole (Microtus agrestis). II. The effect of daylength // Gen. Comp. Endocrinol., 1971b. V. 16. P. 239–235.
Khateeb A.A., Johnson E. Seasonal changes of pelage in the vole (Microtus agrestis). III. The role of the endocrine system // Gen. Comp. Endocrinol., 1971c. V. 16. P. 236–240.
Lehmann von E. Uber die Seitendrusen der Mitteleuropaischen Rotelmaus (Clethrionomys glareolus Schreber) // Z. Morphol. Okol. Tiere. 1962. B. 51. S. 335–344.
Ling J.K. Pelage and moulting in wild mammals with special reference to aquatic forms // Quart. Rev. Biol. 1970. V. 45. P. 16–54.
Mazak V. Zur Kenntnis der Postnatalen Entwicklung der Rotelmaus, Clethrionomys glareolus Schreber, 1780 (Mammalia, Microtidae) // Acta Soc. Zool. Bahemoslov. 1962. B. 26. S. 77–104.
Meester J. The fur and moult in the shrew, Myosorex cafer // J. Mammal., 1958. V. 39. № 4. P. 494–498.
Michielsen N.C. Intraspecific and interspecific competition in the shrews Sorex araneus L., and S. minutus L. // Arch. Neerland Zool. V. 17. № 1. P. 73–174.
Morrison P. Adaptation of small mammals to the arctic // Fed. Proc. 1964. V. 23. № 6. P. 1292–1206.
Quay W.B. The specialized posterolateral sebaceous glandular regions in microtine rodents // J. Mammal. 1968. V. 49. P. 427–445.
Rowsemitt C., Kunz T.N., Tamarin R.H. The timing and patterns of moult in Microtus breweri // Oecas. Pap. Mus. Nat. Hist. Univ. Kansas, 1975. V. 34. P. 1–11.
Scholander P.F., Hock R., Walters W., Irving L. Heat regulation in same arctic and tropical mammals and birds // Biol. Bull. 1950. V. P. 237–258.
Skaren U. Variation in two shrews, Sorex unguiculatus Dobson and S. a. araneus L. // Ann. Zool. Fenn. 1964. V. 1. № 2. P. 94–124.
Skaren U. Spring moult and onset of the breeding season of the common shrew (Sorex araneus L.) in the Central Finland // Acta Theriol. 1973. V. 18. № 23. P. 443–458.
Stein G.H.W. Biologische Studien an Deutchen Kleinsaugern // Arch. Naturgesch. 1938. B. 7. № 4. S. 477–513.
Stein G.H.W. Materialen zum Haawechsel Deutscher Insectivoren // Mitt. Zool. Mus. Berlin. 1954. B. 30. S. 12–34.
Stein G.H.W. Zum Haarwechsel der Feldmaus (Microtus arvalis) und Weiterer Murroidea // Acta Theriol. 1960. V. 4. № 6. P. 12–34.
Viro P. Moulting and structure of the fur in the bank vole, Clathrionomys glareolus (Schreber 1780), in the vicinity of Oulu, Finland // Aquil. Ser. Zool. 1979. V. 19. P. 49–58.
Viro P., Koskela P. Moult topography, moulting and the structure of the fur in the harvest mouse // Acta Theriol. 1978. V. 23. № 34. P. 503–517.
Wilke G. Freilands- und Gefangenscaftsbeobachtungen an Sorex araneus L. // Z. Saugetierk. 1938. B. 12. S. 332–335.
Williams L.W. Moult in Blarina brevicauda // J. Mammal. 1962. V. 43. № 3. P. 423–424.
Zejda J., Mazak V. Cycle de changement du pelage chez le compagnol roussatre, Clethrionomys glareolus Schreber 1780 (Microtidae, Mammalia) // J. Mammalia. 1965. V. 29. P. 577–597.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии