

Успехи физиологических наук, 2019, T. 50, № 3, стр. 14-33
Негативность рассогласования и пространственный слух
Л. Б. Шестопалова a, *, Е. А. Петропавловская a
a Федеральное государственное бюджетное учреждение науки Институт физиологии им. И.П. Павлова РАН
199034 Санкт-Петербург, Россия
* E-mail: shestopalovalb@infran.ru
Поступила в редакцию 21.05.2018
После доработки 08.10.2018
Принята к публикации 25.02.2019
Аннотация
Потенциал негативности рассогласования (НР) принадлежит к классу длиннолатентных слуховых вызванных потенциалов (ВП) и генерируется как реакция мозга на изменения соотношения сенсорных признаков стимула с предшествующим звуковым контекстом. В обзоре рассмотрены современные представления о негативности рассогласования и ее интерпретации. Подробно описаны накопившиеся к настоящему времени данные относительно формирования НР при локализации неподвижных и движущихся звуковых сигналов. Отдельное внимание уделено соотношению между НР и осознанным обнаружением стимулов, а также контекстным эффектам при исследовании восприятия слуховых различий. Детально рассматриваются вопросы межполушарной асимметрии НР при пространственном слуховом анализе.
Одно из актуальнейших направлений современной нейрофизиологии связано с обработкой сенсорной информации в мозге человека, происходящей без приложения сознательных усилий и без фокусировки внимания. При этом формируются так называемые автоматические (preattentive) реакции мозга. Механизмы возникновения этих реакций составляют фундаментальную проблему когнитивной нейрофизиологии и нейропсихологии. Ключевым звеном исследований в этой области служит потенциал негативности рассогласования (англоязычный термин – mismatch negativity, MMN), принадлежащий к классу длиннолатентных слуховых вызванных потенциалов (ВП). Впервые негативность рассогласования (далее НР) была описана профессором Хельсинкского Университета Ристо Наатаненом [111] и получила широчайшее распространение при оценке деятельности не только слуховой системы, но и других сенсорных систем человека (зрительной, обонятельной, соматосенсорной) [11, 112, 114, 115]. В последующих разделах данного обзора будут рассмотрены общие свойства НР и ее интерпретация, а затем будут подробно описаны накопившиеся к настоящему времени данные относительно НР, вызванной изменениями пространственного положения звуковых сигналов. Отдельное внимание будет уделено соотношению между НР и осознанным обнаружением стимулов, а также межполушарной асимметрии НР при пространственном слуховом анализе.
1. ПОЛУЧЕНИЕ НЕГАТИВНОСТИ РАССОГЛАСОВАНИЯ И ЕЕ ИНТЕРПРЕТАЦИЯ
В первые годы после открытия НР ее исследования проводились (и сейчас нередко проводятся) в условиях oddball-стимуляции, которая состоит в предъявлении испытуемому последовательности одинаковых звуковых стимулов (стандартов), в ряду которых с небольшой вероятностью появляются стимулы, отличающиеся по какому-либо параметру (девианты). При вычитании ВП на стандартный стимул из ВП на девиант во фронто-центральных отведениях наблюдается негативная волна НР, генерируемая главным образом в слуховой коре [23, 76], хотя также имеются данные о формировании НР во фронтальной коре [23, 121, 137].
Схематическое изображение компонентов ВП, регистрируемых в режиме oddball-стимуляции (рис. 1), иллюстрирует общий характер ответов на стандартные и девиантные стимулы, а также разностные потенциалы, генерируемые при пассивном прослушивании (ignore condition) и при фокусировке внимания на звуковой стимуляции (attend condition). Процессы обработки сенсорной информации в данной парадигме могут быть в целом описаны последовательной трехэтапной моделью [59, 60, 79, 80]. Первый этап связан с пассивной (неосознаваемой) обработкой сенсорного сигнала и его фильтрацией. В вызванных потенциалах этот процесс находит отражение в виде негативной волны N1, характеризующей обнаружение сенсорного события [116], и волны НР, реагирующей на различия в потоке стимулов [114]. N1 и НР принято считать модально-специфическими детекторами различий первого и второго порядка, соответственно. Латентность N1 варьирует в пределах от 80 до 120 мс от начала стимула, латентность НР – от 100 до 300 мс от начала девиации.
Рис. 1.
Схематическое изображение компонентов ВП и негативности рассогласования (MMN), регистрируемых в режиме oddball-стимуляции при пассивном прослушивании (ignore condition) и при фокусировке внимания на звуковой стимуляции (attend condition). Вверху слева: ВП на стандартные стимулы и негативности рассогласования, регистрируемые электродами Fz, Cz и Pz при пассивном прослушивании, т.е. без привлечения внимания к звуковой стимуляции. Эти компоненты отражают полностью автоматическую слуховую обработку. Вверху справа: ВП на стандартные и девиантные стимулы, регистрируемые теми же электродами. Параметры компонентов N1 и P2 могут различаться в зависимости от характера и вероятности предъявления стандартов и девиантов. Важно отметить более фронтальное формирование негативности рассогласования по сравнению с N1. Внизу слева: структура компонентов ВП при регистрации теми же электродами в активных условиях, т.е. при привлечении внимания слушателей к звуковой стимуляции. Важнейшее отличие от ВП, полученных в пассивных условиях, состоит в появлении комплекса N2b-P3a, перед которым часто наблюдается позитивная волна P165, а также медленных фронтальных негативных и теменных позитивных компонентов. Компонент N2b идентифицируется после удаления негативности рассогласования из компонента N2 при вычитании. В зависимости от инструкции, привлечение внимания может увеличивать амплитуду N1 и негативности рассогласования. Внизу справа: ВП на стандартные и девиантные стимулы, регистрируемые теми же электродами. Цит. по: Näätänen et al., 2011, figure 1.
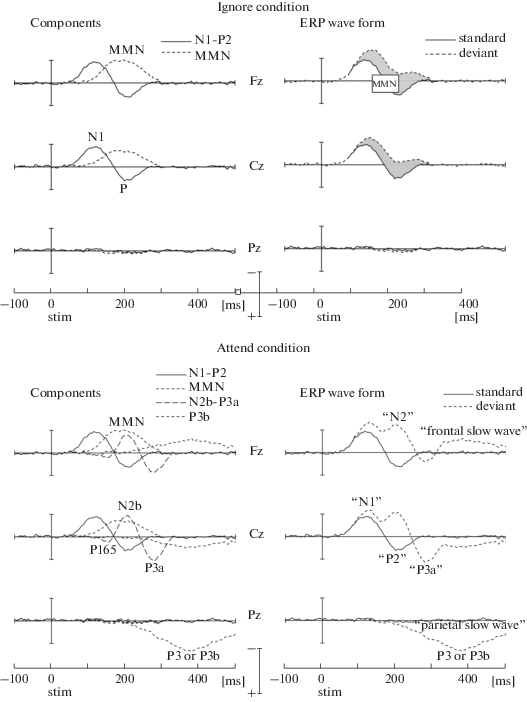
Процессы, лежащие в основе N1 и НР, могут привести к непроизвольному переключению внимания на девиантный стимул и переходу ко второму, активному, этапу сенсорной обработки. Индикатором непроизвольного переключения внимания считается позитивный компонент вызванного потенциала P3a, максимально выраженный во фронто-центральных отведениях. Он может отражать как сенсорные, так и когнитивные аспекты сенсорного события без выраженной модальной специфичности [33, 62]. В настоящее время связь между процессами, формирующими N1 и НР на первом этапе обработки информации и вызывающими P3a на втором этапе этой обработки, остается до конца не выясненной [79, 138].
Активные процессы, происходящие после переключения внимания, составляют третий этап сенсорной обработки. Нейрональная активность передается на височно-теменные области, где происходит генерация второго позитивного компонента вызванного потенциала – P3b, предположительно связанного с произвольным переключением внимания. P3a и P3b принято относить к субкомпонентам волны P300. Их латентность варьирует в широких пределах (от 250 до 800 мс) и зависит от сложности задачи по различению стимулов и от индивидуальных когнитивных особенностей испытуемых [48, 135]. Наконец, в тех случаях, когда девиантный стимул, вызвавший переключение внимания, не требует немедленных действий, установки избирательного внимания восстанавливаются к исходным. Этот процесс отражается в генерации модально неспецифического фронто-центрального негативного компонента, носящего название “reorienting negativity” (RON) с латентностью 400–600 мс [37, 79, 145, 148, 149].
Потенциал НР обладает следующими важными свойствами: 1) НР возникает как при пассивном прослушивании сигналов (при чтении книги или выполнении зрительного задания), так и при активном распознавании девиантных стимулов; 2) амплитуда НР увеличивается при уменьшении вероятности предъявления девиантного стимула в последовательности; 3) амплитуда НР растет, а латентность уменьшается при увеличении акустических различий между стандартом и девиантом [171].
Первоначальные интерпретации потенциала НР основывались на двух гипотезах. Адаптационная гипотеза, или гипотеза рефрактерности [81, итоговый обзор 102, 108, 109, 112] предполагает, что негативный фронто-центральный компонент ВП в реакции на девиантный стимул представляет собой составную часть измененного компонента N1. Гипотеза рассогласования со следом в памяти (memory mismatch) рассматривает фронто-центральную негативность в качестве специфического компонента ВП, генерируемого в ответ на девиантный стимул и отражающего различие между девиантом и следом в сенсорной памяти, сформировавшимся при регулярном предъявлении стандарта [108, 109, 118]. В результате интенсивной дискуссии и анализа большого количества экспериментальных данных, концепция рассогласования со следом в памяти была принята большинством исследователей [см. обзоры 110, 114, 171].
Однако, интерпретация разностного потенциала как специфического показателя автоматического различения требует важного уточнения. Во-первых, генераторы компонента N1 несомненно подвергаются адаптации при частом предъявлении стимула, и эффект адаптации для стандарта и девианта в одной последовательности будет разным, поскольку они появляются с разной частотой. Разное влияние адаптации может вносить вклад в разностный потенциал. Во-вторых, реакции на стандарт и девиант могут различаться даже при небольших акустических различиях между ними, и это тоже может отражаться на потенциале НР. Обе причины могут вести к искажениям при вычислении НР как разности между ответами на девиант и стандарт в пределах одной последовательности. Чтобы избежать искажений, для получения “истинной” (genuine) НР либо проводятся контрольные эксперименты, в которых оценивается величина возможных искажений, либо применяется так называемая “обратная oddball-парадигма” (reversed oddball), позволяющая вычислять НР как разность между ответами на идентичные стимулы, предъявленные в качестве девианта в одной последовательности и в качестве стандарта – в другой [10, 82, 83, 147]. Такой способ вычисления получил название “одностимульной” НР (same-stimulus MMN).
Определенные ограничения для понимания процессов автоматического распознавания различий были связаны с тем, что потенциал НР первоначально исследовался в условиях oddball-парадигмы, создающей слишком упрощенную модель акустической сцены. Важным шагом в развитии интерпретаций НР явился переход к сложным конфигурациям сигналов, более близким к реальной звуковой среде, но не позволяющим соотнести след в сенсорной памяти с одним или несколькими конкретными стандартными стимулами. Представление о стандарте как “повторяющемся стимуле” было расширено до “повторяющегося соотношения между стимулами”, а девиант стал рассматриваться как “нарушение закономерности” (regularity violation). Под закономерностью подразумевается неявное последовательное правило (sequential rule), извлекаемое слуховой системой из серии звуковых событий [см. обзор 173]. Примерами таких конфигураций могут служить нерегулярные изменения межстимульного интервала [61, 120], либо закономерности вида “чем выше частота – тем ниже интенсивность” [127] или “за длинным стимулом следует высокий, а за коротким – низкий” [124]. НР стала также применяться в качестве объективного показателя разделения конкурирующих слуховых потоков (stream segregation), формируемых при предъявлении чередующихся звуковых последовательностей [см. обзор 35]. Традиционная oddball-стимуляция рассматривается теперь как один из простых частных случаев парадигмы нарушения закономерности [49, 170].
Согласно современным представлениям, основная функция генерации НР – поддержание и обновление нейрональных моделей, лежащих в основе обнаружения и разделения звуковых объектов. Мозг кодирует слуховую и абстрактную (внесенсорную) информацию, характеризующую недавнюю историю стимуляции, и на основании обнаруженных закономерностей формирует предсказательные модели относительно стимулов, которые появятся в ближайшем будущем. Результатом этого процесса является оценка не только сенсорных признаков стимула, но и соотношения стимула со звуковым контекстом и его соответствия выявленным закономерностям. Когда поступающий девиантный стимул нарушает обнаруженную закономерность, генерируется потенциал НР, величина которого отражает степень девиации. Возникновение НР служит сигналом к обновлению предсказательной модели, осуществляя обратную связь между моделью и обработкой текущей информации.
Будучи индикатором реакции мозга на ошибку предсказания, формирование НР может предшествовать реакциям, обнаружимым на поведенческом уровне, и служить “сигналом тревоги”, инициирующим переключение внимания на девиантное событие. Таким образом, даже на ранних кортикальных уровнях слуховой системы, предшествующих подключению внимания (preattentive level), могут выполняться достаточно сложные когнитивные операции, связанные с обобщением информации [см. обзоры 110, 122, 171].
2. НЕГАТИВНОСТЬ РАССОГЛАСОВАНИЯ ПРИ ИЗМЕНЕНИИ ПРОСТРАНСТВЕННЫХ ПРИЗНАКОВ ЗВУКОВОГО СТИМУЛА
2.1. Негативность рассогласования при изменении положения неподвижного стимула.
Правильная акустическая ориентация человека во многом определяется способностью различать изменения пространственных характеристик источников звука. В изучении ранних этапов слухового анализа важная роль принадлежит работам по регистрации НР как в условиях дихотического предъявления сигналов, так и при стимуляции в свободном звуковом поле.
Метод дихотической стимуляции получил широкое распространение в связи с возможностью изолированного или комбинированного предъявления бинауральных локализационных признаков звука – межушных различий по времени (ΔT) и интенсивности (ΔI). Регистрация НР в дихотических условиях была применена для изучения механизмов обработки ΔT и ΔI, и в частности, для решения вопроса о том, является ли анализ этих признаков раздельным, или это совместный процесс. В исследовании Шрёгера с соавторами применялись стимулы с ΔT, с ΔI и с комбинацией этих признаков [144, 146]. Амплитуда каждой из НР, возникающих при введении в девиант только одного из признаков, была меньше амплитуды НР, генерируемой при сочетании признаков ΔT и ΔI в девианте. Авторы предполагают, что частичная аддитивность НР указывает на параллельное и независимое (по крайней мере, отчасти) нейрональное кодирование этих двух признаков на уровне коры. В известной степени такой вывод согласуется с данными по положению дипольных источников компонента N1 ВП, свидетельствующими о наличии относительно независимой обработки ΔT и ΔI в слуховой коре человека [169]. Кроме того, он вполне укладывается в рамки дуплексной теории пространственного слуха, постулирующей наличие двух различных механизмов локализации источника звука по ΔT и ΔI [2–4]. Предположение о раздельном анализе локализационных признаков звука подкрепляется результатами исследования магнитных аналогов НР с использованием виртуальных источников звука, сформированных по передаточной функции головы [64]. Топографический анализ показывает, что различение бинауральных признаков, определяющих азимут источника звука, и различение спектральных признаков, связанных с вертикальным положением источника звука, обеспечиваются разными участками височной коры мозга. При этом бинауральные локализационные признаки (ΔT и ΔI) обрабатываются раньше, чем спектральные признаки (100–150 мс и 200–250 мс, соответственно).
Значительная часть исследований была посвящена изучению характеристик НР как показателя пространственного различения сигналов. Впервые связь НР с угловым расстоянием между девиантным и стандартным стимулами была установлена в работе Паавилайнена с соавторами [125]. В условиях латерализации девиантного стимула за счет межушных различий по фазе (тон 600 Гц) или по интенсивности (тон 3000 Гц) амплитуда НР возрастала как функция угла смещения девианта относительно стандарта. Та же закономерность была выявлена в работе [124] с использованием тональных сигналов с межушными различиями по времени. Сходные данные были получены в других работах при использовании различных шумовых сигналов и различном пространственном разнесении стандартного и девиантного стимулов по азимуту [56, 119, 154].
В отличие от исследований в условиях дихотической стимуляции, результаты работ, выполненных в свободном звуковом поле, оказались не столь однозначными [47, 52, 54, 125, 175]. Увеличение амплитуды НР в зависимости от пространственного разнесения стандартного и девиантного стимулов обнаружено только в работе Дэуэлла и др. [54]. В остальных исследованиях наблюдались изменения НР по закону “все или ничего”.
НР может отражать не только воспринимаемое изменение в пространственном положении стимулов, но также субъективные различия их пространственной протяженности. В работе Винклера и др. [175] два источника девиантного сигнала располагались под углом 45° справа и слева от слушателя, и включались либо по очереди, либо одновременно. В последнем случае девиантный стимул воспринимался в том же месте, что и стандартный сигнал (по центру головы), но отличался от него большей пространственной протяженностью. Во всех трех случаях возникали НР приблизительно равной амплитуды. Появление НР в ответ на изменение субъективной протяженности стимулов согласуется с представлением о том, что в основе генерации НР могут лежать не только физические, но также и воспринимаемые (субъективные) различия в сигналах.
2.2. Негативность рассогласования и локализация движущегося стимула
Одной из важных составляющих пространственного слуха является локализация движущихся источников звука. Анализ акустических признаков движения звука слуховой системой уже давно составляет предмет как психофизиологических, так и нейрофизиологических исследований. Полученные к настоящему времени экспериментальные факты указывают на наличие областей коры, избирательно активируемых движущимися звуковыми стимулами. Об этом свидетельствуют как данные нейровизуализации мозга человека [34, 42, 72–75, 96, 101, 169], так и результаты регистрации активности одиночных нейронов на животных [17, 26, 156, 167]. Была выдвинута гипотеза о наличии в слуховой системе специализированных нервных элементов (“детекторов движения”), обеспечивающих непрерывный анализ параметров источника звука по мере его движения [26]. Электро- и магнитоэнцефалографические исследования показали, что при движении источника звука в коре генерируются выраженные слуховые вызванные потенциалы и их магнитные аналоги [28, 39, 57, 87, 177]. Избирательность реакции к направлению движения, по мнению некоторых авторов, могла служить подтверждением наличия дирекциональной чувствительности нейронов слуховой коры [103].
Эффективным новым подходом к исследованию кортикальных процессов обработки движения стимула становится использование потенциала НР как объективного показателя при решении вопроса о том, сводится ли первичная обработка движения стимула к локализации начальных и конечных точек его траектории. Установлено, что формирование НР происходит за счет анализа более длительного участка стимула, чем при формировании компонента N1. Генераторы N1 кодируют акустическую информацию в пределах 40–50 мс после включения сигнала, поэтому компонент N1 представляет собой в большей степени реакцию на включение сигнала, чем на его динамическую структуру. В то же время, НР отражает структуру стимула на протяжении 300 мс и более [113, 114]. Следовательно, различия, введенные в функцию межушной задержки сигнала (ΔT и/или ΔI) в пределах 300 мс, могут быть адекватным образом отражены в параметрах потенциала НР. Движение стимула с разными скоростями может быть успешно моделировано введением межушной задержки ΔT, изменяющейся по разным временным законам, а движение по траекториям разной длины – путем введения ΔT разной величины. Сравнительный анализ характеристик НР, вызванной динамическими изменениями ΔT, позволяет определить, считывается ли на ранних корковых этапах слухового анализа информация о временной структуре сигнала, содержащаяся в средней части траектории его движения.
В первом исследовании формирования НР, вызванной линейными изменениями межушной задержки ΔT, использовались центральное и два латеральных (у левого и правого уха) положения стандартного стимула, и три типа девиантов: неподвижные либо движущиеся по направлению к стандартным стимулам или от них [30]. При движении девиантов по направлению к стандарту потенциал НР определялся главным образом величиной ΔT в начальной точке траектории, а последующие линейные изменения ΔT практически не отражались на параметрах НР. В противоположной ситуации, при движении девианта от стандарта, потенциал НР генерировался с большей латентностью и приобретал чувствительность к изменениям ΔT. Величина НР была больше, когда сравнение стимулов происходило преимущественно во фронтальном секторе.
Количественная связь НР и параметров движения стимула во фронтальном акустическом секторе была установлена в работе Альтмана с соавторами [29]. Используя два способа дихотической стимуляции (варьирование межушной задержки ΔT при постоянной длительности стимула либо изменение длительности при фиксированной ΔT), авторы разделяют влияние факторов расстояния и времени движения на величину НР. Согласно полученным результатам, амплитуда НР не зависела от длительности сигнала. Девиантные стимулы, смещавшиеся на одни и те же угловые расстояния с разными скоростями за счет различия в длительности, вызывали НР одинаковой амплитуды, но разной латентности. Это свидетельствует о том, что угловое расстояние, пройденное звуковым стимулом, является более важным признаком для формирования НР, чем собственно скорость.
Зависимость параметров НР от временного паттерна динамических изменений межушной задержки ΔT исследована в серии работ с применением плавного и скачкообразного движения стимулов [6, 9, 153]. Авторы исходили из предположения, что если бы величина углового смещения действительно являлась единственным определяющим параметром при формировании НР, то это означало бы, что механизм автоматического пространственного различения основывается на оценке положения только начальных и конечных точек траектории. Такая гипотеза была выдвинута ранее в классических психофизических исследованиях слуха [70, 106] и получила название “теории мгновенных отпечатков” (“snapshot theory”). Согласно этой теории, восприятие движения звука основывается не на специализированных нейрональных “детекторах движения”, а на последовательной локализации дискретных точек траектории звукового источника. В случае коротких (менее 250 мс) стимулов слуховая система обрабатывает только информацию о положении концов траектории движения [140]. Таким образом, согласно теории “мгновенных отпечатков”, слуховая система не способна различить стимулы, смещающиеся на одно и то же угловое расстояние за одинаковое время по разным траекториям. Для проверки этой гипотезы в упомянутой серии работ [6, 9, 153] были использованы два вида изменений ΔT (плавное и мгновенное нарастание) в движущихся звуковых стимулах (рис. 2). Плавное изменение ΔT и скачок приводили к разным зависимостям НР от величины углового смещения. При ΔT ≥ 40 мкс скачок вызывал бóльшую НР, чем плавное движение, при одной и той же величине углового смещения. Следовательно, величина НР может определяться не только пройденным угловым расстоянием, но и характером динамических изменений ΔT. Кроме того, в этой же серии работ была получена НР при предъявлении скачка в качестве девианта в контексте плавно движущихся стандартов, при совпадении положения их начальных и конечных точек. Это означает, что динамические свойства стимула могут быть вовлечены в обработку движения звукового стимула наряду с информацией о концах его траектории.
Рис. 2.
Негативность рассогласования при различении плавного и мгновенного перемещения в контексте неподвижных звуковых стимулов. (а): слева – мгновенное и плавное (линейное) изменение ΔT в стимулах, синтезированных на основе белого шума, фильтрованного в полосе 100–1300 Гц; справа – соответствующие траектории перемещения звуковых стимулов в горизонтальной плоскости. Межушная задержка возрастала от 0 до ±20, ±40, ±120, ±200 мкс, что соответствовало угловым смещениям в ±2°, ±5°, ±14° и ±23°. На схеме показаны только траектории движения от центра вправо; в эксперименте использовалось также движение от центра влево. (б): амплитуды негативности рассогласования (НР), вызванной мгновенным и плавным перемещением стимулов (белые и черные кружки, соответственно) в контексте неподвижных стандартных стимулов, локализуемых по средней линии головы. Отведение Fz. Вертикальными черточками показана стандартная ошибка среднего (N = 9). Цит. с изменениями по: Shestopalova et al., 2012, figure 4.

Формирование НР, вызванной скачком и плавным движением, можно рассмотреть с точки зрения понятия “временного окна интеграции”, производящего усреднение бинауральной информации с определенной постоянной времени [40, 71]. Временное интегрирование акустической информации в бинауральной слуховой системе рассматривается в виде экспоненциально убывающей функции, присваивающей более низкие веса более удаленным событиям в прошлом [50, 93]. Обработку стимула слуховой системой можно представить, как свертку функции межушной задержки сигнала с функцией временного окна. Результат временного интегрирования окажется больше для скачкообразного изменения ΔT, чем для линейной функции, соответствующей плавному движению. Большие амплитуды НР в реакции на скачок (по сравнению с плавным движением) подтверждают правомерность рассмотрения обработки плавного и скачкообразного движения слуховой системой как процесса временного интегрирования. Наличие в бинауральной слуховой системе интеграционных процессов позволяет относить стимулы с мгновенным изменением ΔT к категории движущихся плавно с очень большой скоростью и использовать их в качестве модели “самого быстрого стимула” в задачах сравнения скоростей.
3. НЕГАТИВНОСТЬ РАССОГЛАСОВАНИЯ И СУБЪЕКТИВНЫЕ ПОКАЗАТЕЛИ СЛУХОВОГО РАЗЛИЧЕНИЯ
3.1. Негативность рассогласования и психофизическое обнаружение стимулов
Соотношение объективных и субъективных показателей различения стимулов является одним из основных вопросов когнитивной психофизиологии. Как было сказано выше, первичная обработка слуховой информации происходит на уровне, предшествующем вниманию (preattentive). Генерация НР происходит в результате нарушения обнаруженных закономерностей звуковой стимуляции, включающих сенсорную информацию как о стимуле, так и о контексте, в котором он был предъявлен. Наименьшая величина акустических различий между стандартом и девиантом, приводящая к возникновению НР, приблизительно соответствует минимальному обнаружимому изменению данного параметра (например, длительности, частоты, пространственного положения и т.п.). На групповом уровне возникновение и амплитуда НР изменяются вместе с различительной способностью к исследуемому признаку вследствие созревания, обучения или повреждения мозга [16, 92, 94, 98, 117]. Амплитуда НР может определяться величиной перцептивных различий в большей степени, чем акустических [163, 174, 175].
Нарушение закономерности в звуковой последовательности зачастую может быть обнаружено на сознательном уровне благодаря переключению внимания, происходящему после предъявления девианта [108, 113, 171]. Многочисленные работы показывают, что формирование следа стимула в сенсорной памяти, лежащее в основе генерации НР, может быть исследовано также психофизическими методами [см. обзор 97]. Однако, в некоторых случаях акустические различия, вызывающие НР, оказываются ниже порога активного распознавания [25, 41, 51, 86, 107]. Описаны также экспериментальные ситуации, когда перцептивно различные стимулы не вызывали НР, либо формировались одинаковые НР при наличии перцептивных отличий в стимулах [38, 133, 142, 161]. Так, например, отсутствие НР наблюдалось при предъявлении отчетливо различимых коротких стимулов [21, 126]. В целом, корреляция между параметрами НР и субъективной различимостью стимулов оказывается неустойчивой, особенно на индивидуальном уровне.
Отсутствие стойкой корреляции между объективными и субъективными показателями различения может объясняться различиями механизмов формирования психофизических реакций и НР, связанными с разными уровнями обработки сенсорной информации в мозге. С другой стороны, отсутствие корреляции может быть вызвано чисто методическими причинами. Известно, что разные психофизические методы измерения дифференциальных порогов (например, метод “да–нет”, метод вынужденного выбора) основываются на разных критериях субъективной оценки и могут давать разные результаты [84]. Кроме того, измерения психофизических порогов и регистрация НР в разных исследованиях проводились в неодинаковых условиях звуковой стимуляции. При измерении психофизических порогов обычно использовались процедуры интервального наблюдения, тогда как при регистрации НР предъявляется последовательный ряд стимулов. Очевидно, что для корректного сравнения психофизических и электрофизиологических данных условия звуковой стимуляции должны совпадать. Работы, в которых выполнено это требование, будут рассмотрены ниже (раздел 3.3).
3.2. Негативность рассогласования при околопороговых различиях в пространственных звуковых стимулах
Применительно к пространственному слуху, тема взаимосвязи субъективных и объективных характеристик слухового различения сигналов имеет два аспекта: 1) обнаружение движущегося стимула в контексте неподвижных и 2) различение сигналов, движущихся с разными скоростями.
Для оценки субъективного различения движущихся стимулов от неподвижных в работах с применением плавного и скачкообразного движения [6, 153] применяли метод двухальтернативного вынужденного выбора, и определяли порог различения при помощи процедуры лестниц. Минимальное угловое смещение, при котором движущиеся стимулы были отличимы от неподвижных, составило 27 ± 3 мкс для плавного движения и 33 ± 3 мкс для скачка. В то же время, минимальная величина ΔT, при которой получена значимая НР, составила 20 мкс, что может указывать на более высокую разрешающую способность процессов автоматической обработки пространственных стимулов.
Непосредственное сравнение объективных и субъективных показателей различения движущихся стимулов при варьировании длины траектории выполнено в работе Шестопаловой и др. [12]. Для каждой из траекторий применялись стимулы, различающиеся динамикой изменения ΔT (плавное и скачкообразное перемещение). Уровень субъективного различения определяли методом двухальтернативного вынужденного выбора. Стимулы, перемещающиеся по коротким и длинным траекториям, соответствующие подпороговому и надпороговому субъективному различению, использовали в качестве стандартов и девиантов в экспериментах с регистрацией НР. Подпороговые (субъективно неразличимые) стимулы с плавным и скачкообразным движением вызывали формирование значимой НР, что подтверждает выводы работ по различению движущихся и неподвижных стимулов [6, 153] о способности мозга распознавать пространственные сигналы на уровнях, предшествующих формированию осознанного восприятия. Переход к надпороговым различиям в перемещении стимулов приводил к улучшению как субъективного, так и электрофизиологического показателей различения в среднем по группе, но не на индивидуальном уровне. При увеличении длины траектории движения, амплитуда НР и процент правильных ответов в психофизическом тестировании не достигали значимой корреляции.
Переходя к вопросу о различении сигналов, движущихся с разными скоростями, следует отметить, что существующие данные по дифференциальной чувствительности слуховой системы к скорости движения стимулов не позволяют напрямую соотнести характеристики субъективного восприятия скорости и его электрофизиологические показатели. Взаимозависимость параметров движения (времени, пути и скорости) составляет ключевую проблему субъективного различения скорости. Скорость движения определяется пройденным расстоянием и временем движения, которые сами по себе могут служить признаками для различения стимулов, движущихся с разными скоростями. При варьировании скорости стимула за счет времени движения либо за счет длины траектории проблема множественности признаков остается нерешенной. Тем не менее, психофизические исследования показали, что слуховая система способна напрямую оценивать скорость движения, однако присутствие в движущихся стимулах пространственных различий может существенно влиять на их различение по скорости [46, 70]. Установлено, что чувствительность слуховой системы человека ниже при различении быстро движущихся источников звука, чем при различении медленных. С увеличением скорости движения источника звука слуховая чувствительность к его пространственному смещению постепенно снижается. Количественно это выражается в линейном возрастании минимально различимого угла движения стимула с ростом его скорости [4 (обзор)]. Одновременно с ухудшением пространственной разрешающей способности слуховая система становится менее чувствительной к изменению скорости движения источника звука. Абсолютные значения дифференциального порога по скорости монотонно возрастают по мере увеличения скорости стимула [27, 31, 46, 89, 174].
Нейрофизиологические механизмы восприятия человеком скорости движения источника звука до конца не изучены. Исследования электрической активности мозга показывают, что характеристики слуховых вызванных потенциалов (ВП), в частности, комплекса N1P2, могут изменяться в зависимости от скорости движения. Увеличение скорости вызывает возрастание амплитуды и уменьшение латентности компонентов N1 и P2. Эти свойства реакции достаточно хорошо выражены на высоких скоростях движения (более 90 град/с) и полностью исчезают при переходе к более низким скоростям [5]. Нечувствительность ВП к медленным движениям звукового источника подтверждают исследования, в которых движение стимула производилось с задержкой относительно момента включения сигнала [8, 65, 66, 95]. В этих условиях ВП, соответствующий движущемуся фрагменту сигнала, отсутствует на малых скоростях движения, появляется при скоростях 90–100 град/с и монотонно возрастает по величине при последующем увеличении скорости. При исследовании ВП показателем реакции на скорость стимула служит степень изменения величины ответа при изменении скорости. Аналогичным показателем может служить амплитуда НР.
Сравнение разрешающей способности слуховой системы к скорости движения стимула, измеренной путем регистрации НР и путем определения дифференциальных порогов по скорости движения, выполнено в работе Шестопаловой с соавторами [13]. Авторами был специально разработан набор движущихся стимулов, в которых переменным параметром являлась скорость, а длительность и угловое смещение оставались постоянными (рис. 3). Такие сигналы позволяли избежать проблемы множественности признаков в задаче различения стимулов по скорости их движения, но делали задачу различения достаточно сложной для слушателей, поскольку они были лишены возможности опираться на дополнительные признаки времени и пройденного пути. Дифференциальные пороги по скорости определялись в режиме двухальтернативного вынужденного выбора при помощи метода лестниц. Полученные значения дифференциальных порогов оказались ожидаемо выше величин, известных из литературы, и составили 242 ± 13 град/с при перемещении стимулов влево и 234 ± 19 град/с при перемещении вправо, или 54 и 52% в относительных единицах, соответственно. Электрофизиологическая часть исследования показала, что значимый потенциал НР формируется в комбинации плавно движущихся стимулов со скоростями 450 град/с (стандарт) и 620 град/с (девиант), т.е. при разнице скоростей стандарта и девианта 170 град/с (или 38% в относительных единицах). Таким образом, при сравнении с психофизическими измерениями, амплитуда НР оказалась более чувствительным показателем различения как при обнаружении движущихся стимулов в контексте неподвижных, так и при различении сигналов, движущихся с разными скоростями.
Рис. 3.
Негативность рассогласования при различении скоростей плавного движения стимулов. (a): набор движущихся стимулов, в которых переменным параметром являлась скорость, а длительность и угловое смещение оставались постоянными. Полный набор скоростей плавного движения разработан для психофизического эксперимента по определению дифференциального порога по скорости (Шестопалова и др., 2015). Для получения негативности рассогласования использовались стимулы, движущиеся со скоростями 450 град/с и 620 град/с (темные жирные сплошные линии), а также скачок (толстый пунктир). (б): Негативность рассогласования, вызванная плавным движением (620 град/с) и скачком в контексте плавно движущихся стандартов (450 град/с). Отведение Fz. Усреднение по данным 11 испытуемых.

3.3. Контекстные эффекты в исследованиях перцептивных различий звуковых стимулов
Как субъективные, так и объективные показатели слухового различения могут существенно изменяться в зависимости от акустического контекста стимуляции [напр., 156]. Простейшим примером влияния контекста служат данные по перестановке местами пары сигналов, используемых в качестве стандарта и девианта. В этом случае физические различия между сигналами сохраняются неизменными, тогда как функциональное значение сигналов меняется на противоположное. При этом могут существенно изменяться и перцептивные различия между сигналами. Если уровень различения (объективный или субъективный) не изменяется при инверсии конфигурации стимулов, то данный показатель отражает только физические различия между сигналами. Если же уровень различения изменяется при перестановке стандарта и девианта, то используемый показатель характеризует, помимо физических различий, контекстные особенности стимульного ряда.
В современных исследованиях слухового восприятия контекста широко применяются также последовательности, в которых вероятности предъявления стандарта и девианта изменяются плавно, что приводит к постепенной смене ролей. Согласно принятым представлениям, амплитуда НР обычно больше в последовательностях с медленной сменой ролей стандарта и девианта, чем с быстрой, поскольку быстрая смена ролей ведет к снижению точности предсказательной модели, сформированной в мозге относительно поступающего потока стимулов [63, 171]. Однако, величина НР зависит также от того, какой из двух стимулов появился в роли девианта первым [order-driven bias; 165, 166]. Остается неясным, какие области мозга участвуют в наблюдаемых контекстных эффектах, связанных с извлечением информации о вероятности появления стимулов. Накопленные данные свидетельствуют, что контекстные эффекты могут оказывать весьма существенное влияние на самых ранних этапах слухового анализа.
Режим звуковой стимуляции, при котором каждый вид стимулов выступает в роли стандарта в одних сериях и в роли девианта – в других, при одной и той же структуре всех серий, называется “обратной oddball-парадигмой”. Такая стимуляция использовалась в ряде работ, чтобы устранить возможное влияние чисто акустических различий между стандартом и девиантом на разностный потенциал [напр., 22, 83, 131]. Авторы применяли метод вычисления “одностимульной” НР, при котором НР получают вычитанием ответов на один и тот же стимул, служащий девиантом в одной и стандартом в другой серии. Этот метод успешно применялся в случаях различий по частоте и длительности [напр., 82, 83, 131, 139]. Аналогичная методика получения НР, вызванной пространственными различиями, применена в работе Шрёгера и Вольфа [147]. Авторы использовали неподвижные звуковые стимулы различной локализации. Модифицированная НР вычислялась путем вычитания ВП на контрольные равновероятные стимулы из ВП на девианты, т.е. вычитание производилось между ответами на идентичные стимулы, предъявленные с одинаковой вероятностью. Амплитуда модифицированной НР оказалась меньше, чем традиционной. Авторы приходят к заключению, что пространственные различия могут вызывать истинную НР, в основе которой лежат различительные процессы высокого порядка.
Подробный анализ контекстных эффектов при различении динамических пространственных стимулов выполнен в серии работ Шестопаловой с соавторами [14, 15, 151, 152]. Важной особенностью этих работ является параллельное проведение регистрации ЭЭГ и психофизического тестирования в режиме oddball-стимуляции. В экспериментах применялись дихотически предъявляемые стимулы с быстрым или медленным изменением акустических параметров. Эксперименты с азимутальным движением звуковых стимулов, создаваемым за счет изменений межушной задержки ΔT [15, 152], показали более высокую вероятность активного обнаружения девианта и бóльшую амплитуду НР в тех последовательностях сигналов, в которых девиант перемещался быстрее стандарта. Хотя физические различия между стандартом и девиантом при смене контекста на противоположный оставались неизменными, перцептивные различия оказались больше при быстром перемещении девианта в контексте более медленных стандартов. Амплитуда НР демонстрировала большую чувствительность к скорости движения, чем уровень активного обнаружения девианта и уровень ложных тревог. Этим определялись различия контекстных эффектов в активных и пассивных условиях. Результаты психофизического тестирования указывали на категориальное восприятие движущихся стимулов: скачок и медленное движение могли быть отнесены к одной перцептивной категории (движущиеся сигналы), а неподвижный стимул – к другой.
Более детальное изучение активного и пассивного различения при смене контекста проведено при использовании сигналов с нарастанием или спадом интенсивности [14, 151]. Характерной особенностью восприятия таких сигналов является неоднократно описанное в психофизической литературе перцептивное преимущество стимулов с нарастающей интенсивностью, отражающее высокую биологическую значимость распознавания приближающихся объектов. Как показало исследование, автоматическое (preattentive) различение амплитудно-модулированных сигналов определялось главным образом соотношением интенсивности девианта и стандарта в последовательности, в то время как субъективные показатели различения зависели от направления модуляции: активное различение было более успешным для сигналов с ростом интенсивности. Иными словами, потенциал НР демонстрировал реакцию на структуру последовательности в целом, тогда как психофизические показатели отражали параметры отдельных сигналов. Такое расхождение между нейрональными механизмами генерации НР и процессами, лежащими в основе осознанного различения, служит иллюстрацией диссоциации между функциями “мозга” и “сознания”. На ранних этапах сенсорной обработки нейрональные структуры мозга могут различать слуховые события, неразличимые на более поздних этапах восприятия. Субъективные показатели различения могут не всегда проявлять чувствительность к эффектам, происходящим на низших уровнях обработки и предшествующим переключению внимания, подготовке и выполнению моторного ответа [78, 122]. Формирование НР меньшей латентности для нарастающих различий между стандартом и девиантом отражает временное преимущество их обработки на этапе автоматического различения. С другой стороны, высокий уровень субъективного различения для нарастающих стимулов свидетельствует в пользу гипотезы о том, что рост интенсивности может служить для слушателя внутренним, безусловным предупреждающим сигналом [32]. Вопрос о взаимосвязи этого предупреждающего сигнала с автоматическим различением требует дальнейших исследований. На данном этапе можно заключить, что диссоциация между ранними и поздними этапами слуховой обработки может базироваться на большей биологической значимости роста интенсивности. Информация о структуре последовательности играет меньшую роль при перераспределении ресурсов внимания в сторону потенциально опасных объектов, и поэтому перцептивное преимущество роста интенсивности не нашло отражения в параметрах НР.
4. МЕЖПОЛУШАРНАЯ АСИММЕТРИЯ РЕАКЦИЙ МОЗГА ПРИ ЛОКАЛИЗАЦИИ ИСТОЧНИКА ЗВУКА
4.1 Модели функциональной асимметрии активности мозга
Относительно обработки бинауральных признаков звукового стимула в левом и правом полушариях мозга существуют две основные гипотезы: преимущественная обработка в полушарии, контралатеральном стороне звуковой стимуляции (“контралатеральное доминирование”), и преобладающая роль правой слуховой коры в анализе пространственных признаков звука (“правостороннее доминирование”). Однако в последнее время получено множество экспериментальных свидетельств того, что эти гипотезы не являются полностью взаимоисключающими.
Основой для гипотезы о контралатеральном доминировании полушарий мозга при обработке звуковых стимулов послужили результаты исследований на животных, убедительно показавшие большую величину контралатеральной реакции нейронов при монауральной и бинауральной звуковой стимуляции [134]. Контралатеральное доминирование реакций мозга человека при анализе пространственных признаков звука было также выявлено во многих работах [43 – регистрация ЭЭГ; 91, 96 – метод фМРТ; 104, 129, 130, 141 – регистрация магнитных реакций мозга]. Контралатеральное преобладание ВП описано также в исследованиях, изучавших по отдельности реакцию на включение звука и реакцию на пространственные изменения стимуляции, начало которых отсрочено относительно момента включения. Реакция на начало движения формировалась раньше и была сильнее в полушарии, контралатеральном направлению движения, причем контралатеральное преимущество более выражено при движении звуковых стимулов влево, чем вправо [95]. В работе Гецмана [66] была установлена связь между степенью асимметрии реакции и скоростью движения стимула. Контралатеральное преобладание реакций было сильнее при высоких скоростях движения, а при медленном движении амплитуда ответа становилась больше в правом полушарии.
В то же время, в некоторых работах контралатеральное доминирование при обработке слуховой информации не было обнаружено [85, 176, 180, 181]. Исследования, выполненные при повреждении участков мозга, а также методами фМРТ и МЭГ, выявили правостороннее доминирование реакций на пространственные звуковые стимулы [34, 44, 74, 77, 155, 159, 177, 179].
Результаты ряда исследований не укладывались полностью ни в одну из описанных теорий. Так, в работе Тиитинена [164] при сравнении активности двух полушарий большая амплитуда компонентов N1m и P2m отмечалась в правосторонних отведениях; однако, при сравнении реакций на каждый вид звуковых стимулов большая амплитуда реакции регистрировалась от полушария, контралатерального звуковому стимулу, чем от ипсилатерального. Авторы делают вывод о специализации правого полушария при пассивной обработке пространственных признаков стимула. Согласно данным Паломяки с соавторами, в области левой и правой височной доли наблюдается выраженная контралатеральная активация; однако, в целом амплитуда компонента N1m больше при отведении от правого полушария, чем от левого [129]. Установлено также, что реакции левого полушария активируются преимущественно правосторонним движением стимула, тогда как правое полушарие реагирует на движение звуковых стимулов с обеих сторон (иными словами, движение вправо активирует оба полушария, а движение влево – в основном правую кору) [18, 43, 96]. Контралатеральность реакций левого полушария (при симметричности реакций в правом полушарии) рассматривается в литературе как проявление феномена “левостороннего игнорирования” (в англоязычной литературе – neglect model), под которым подразумевают преимущественную обработку контралатеральной пространственной информации в левом полушарии и обработку информации по всему внешнему пространству в правом полушарии мозга [105, 162]. Более выраженная контралатеральность реакций левого полушария в процессе обработки пространственной слуховой информации наблюдалась при исследовании электрофизиологических и гемодинамических реакций мозга человека, и предположительно связана с тем, что активность левой теменной коры модулируется фронто-теменными нейрональными структурами правого полушария [52, 91, 143].
Возможной причиной столь сильных расхождений в представлениях о том, как организована обработка пространственных признаков звука в центральных отделах слуховой системы человека, могут быть качественные различия в способах звуковой стимуляции, применяемых разными авторами. Установлено, что асимметрия реакций мозга существенно зависит от того, каким именно признаком определяется пространственное положение стимула: межушной задержкой ΔT или ΔI, либо их сочетанием, либо передаточной функцией головы [напр., 74, 88, 129, 168, 177]. Влияние способа стимуляции на асимметрию реакции на начало движения непосредственно исследовалось в работе Гетцмана и Левальда [68]. Контралатеральное преобладание ВП выявлено как при дихотической стимуляции с созданием эффекта движения за счет изменений ΔT и ΔI, так и при стимуляции в свободном поле, а также при использовании трехмерного виртуального движения, тогда как правополушарное преимущество реакций наблюдалось только для виртуального движения и движения в свободном поле.
Другая причина противоречивых результатов исследований латерализации реакций мозга может заключаться в разной направленности внимания испытуемых. В некоторых работах применялись условия пассивного прослушивания, в других внимание испытуемых было направлено на вторичное зрительное задание либо на определенные звуковые стимулы. Межполушарная асимметрия зависит от фокусировки внимания при локализации звука [см. обзор 24]. Базовые аспекты асимметрии могут быть связаны с автоматической обработкой слуховой информации, поэтому феномен НР в последнее время активно используется при изучении функциональной роли левого и правого полушарий в пространственном слуховом анализе. Имеются определенные свидетельства, что процесс автоматического распознавания различий вызывает более выраженную полушарную специализацию соответствующих нейронных структур, чем независимое предъявление стимулов вне процедуры регистрации НР [90].
4.2. Функциональная асимметрия негативности рассогласования для неподвижных стимулов
По данным ряда первых исследований, амплитуда НР была больше в правом полушарии, чем в левом, независимо от стороны звуковой стимуляции [19, 20, 53, 69, 99, 123]. В то же время, детальное исследование топографии НР при дихотической стимуляции и в свободном поле выявило сложную картину асимметрии разных ее компонентов [52]. Височный компонент НР демонстрировал контралатеральное преобладание; фронтальный компонент был значительно смещен вправо при предъявлении левосторонних девиантов, но оставался совершенно симметричным в случае правосторонних. По мнению авторов, эти результаты соответствуют гипотезе о связи фронтального компонента НР с процессами непроизвольного переключения внимания [58]. Аналогичный результат был получен при регистрации магнитного эквивалента НР: правосторонние девианты приводили к формированию НР с одинаковой латентностью в обоих полушариях, тогда как при предъявлении левосторонних девиантов НР в правом полушарии опережала НР левого полушария [91]. Последующие работы также подтвердили, что при отведении от полушария, контралатерального неподвижным латерализованным девиантным стимулам, регистрируется большая амплитуда НР [90, 119, 158, 160]. Преимущественно контралатеральная автоматическая обработка пространственных различий звуковой стимуляции получена также в работе Рихтер с соавторами [136]. Авторы подчеркивают, что степень межполушарной асимметрии НР зависит от положения девиантных стимулов относительно стандартных: контралатеральное преобладание НР значительно более выражено при смещении стимулов от неподвижных стандартов к периферии, чем к средней линии головы.
4.3. Функциональная асимметрия негативности рассогласования для движущихся стимулов
Несмотря на значительное количество и разнообразие упомянутых выше исследований, имеющихся экспериментальных свидетельств недостаточно для окончательного решения вопроса о том, какая модель асимметрии в большей степени соответствует потенциалу НР: правостороннее доминирование, контралатеральное доминирование или модель левостороннего игнорирования. В уже упоминавшейся серии работ по исследованию ВП и НР, вызванных движением стимулов в пределах фронтального акустического сектора [6, 9, 29, 153], также был проведен анализ межполушарной асимметрии НР. Обнаружена тенденция к преобладанию реакции от контралатеральных отведений по сравнению с ипсилатеральными относительно латерализации стимула. В целом асимметрия НР была выражена достаточно слабо [6], но все же результаты указывали на увеличение степени контралатеральности НР с ростом ΔT от 0 до 200 мкс. Авторами было выдвинуто предположение, что асимметрия НР может проявиться более отчетливо при перемещении стимулов от центра к уху до положения максимальной латерализации. В последующей работе [150] исследовалось влияние стороны отведения (левой или правой) и направления перемещения стимула (ипси- или контралатерального стороне отведения) на компоненты N1 и P2 вызванного потенциала и на НР при перемещении звуковых стимулов от средней линии головы к левому или правому уху с разными скоростями по длинным траекториям (от 0 до 800 мс, рис. 3). Кроме того, была исследована временная динамика развития межполушарной асимметрии N1, P2 и НР, а также зависимость асимметрии от скорости движения.
Полученные результаты указывают на явное правостороннее доминирование компонентов N1 и P2: в отведениях от правого полушария регистрировались потенциалы большей амплитуды и латентности, чем в левом полушарии. Правосторонняя активность преобладала во временном интервале 120–160 мс (на нисходящей ветви компонента N1) независимо от скорости стимула. Эти результаты служат подтверждением общепринятого взгляда на функциональное разделение полушарий мозга, при котором правое полушарие играет решающую роль в анализе пространственной и пространственно-временной информации [напр., 44, 141], а левое полушарие обеспечивает начальную базовую обработку звука и дальнейший анализ речевых стимулов [55, 132]. Правая задняя верхневисочная извилина, наряду с правой латеральной фронтотеменной областью, может образовывать супрамодальную нейрональную сеть, предназначенную для контроля внешнего пространства [100, 162, 178].
Потенциалы НР, напротив, не демонстрировали правополушарной асимметрии. При регистрации от левого полушария НР характеризовались преобладанием контралатеральных реакций (на движение вправо), тогда как НР правого полушария не зависели от направления движения стимулов. Скорость движения не оказывала систематического влияния на межполушарную асимметрию НР. Тем самым было продемонстрировано различие в асимметрии НР, регистрируемой от обоих полушарий мозга при перемещении стимулов влево и вправо, и получено подтверждение модели левостороннего игнорирования для автоматического различения движущихся звуковых стимулов.
Совокупность полученных данных свидетельствует о разном характере асимметрии ВП и НР в реакциях на движение звука в oddball-парадигме. Аналогичные эффекты были описаны в электрофизиологических исследованиях реакций мозга на неподвижные и движущиеся стимулы, и было выдвинуто предположение о наличии двух последовательных этапов слуховой обработки движения звука [57, 67]. На первом этапе предположительно происходит интеграция акустических признаков стимула, а на втором – определение направления пространственного сдвига. По мнению Дюкоммуна с соавторами [57], эти два этапа обеспечиваются активностью различных нейронных сетей. Гетцманн [67] пришел к выводу, что ранняя и поздняя фаза ответа на начало движения, демонстрирующие разный характер функциональной асимметрии, могут считаться электрофизиологическими коррелятами двух этапов обработки движения.
Результаты описанного выше исследования [150] вполне соответствуют точке зрения, что ранние и поздние компоненты реакции (N1, P2 и НР) могут отражать последовательные фазы слуховой обработки. Области пика волны N1 могут быть показателями эффекта включения звука, а независящее от скорости правостороннее преимущество, возникающее на нисходящей ветви волны N1, может быть связано с общим контекстом автоматического пространственного восприятия. Компоненты P2 и НР могут отражать непосредственную оценку скорости и перцептивного расстояния между движущимися стимулами.
5. ЗАКЛЮЧЕНИЕ
Анализ имеющейся литературы подтверждает, что НР может рассматриваться в качестве достаточно надежного инструмента для изучения автоматического различения пространственных изменений звуковых стимулов, но только на групповом уровне. Величина НР может отражать не только положение начальной и конечной точек траектории, но и динамические изменения стимула, происходящие в процессе движения.
На индивидуальном уровне НР может слабо коррелировать (или совсем не коррелировать) с субъективными показателями, измеренными психофизическими методами. Слабая корреляция наблюдается для самых различных видов стимуляции и экспериментальных парадигм [см. обзоры 97, 122]. Проблемы взаимосвязи между механизмами формирования НР и активным различением, а также роли избирательного внимания в обработке внезапных изменений акустической среды, требуют детального исследования.
До сих пор не получил окончательного решения вопрос о том, в какой мере параметры НР отражают физические различия между стандартным и девиантным стимулом, а в какой – структуру звуковой последовательности в целом. В этом направлении ведется множество работ, изучающих НР высокого порядка, связанную с распознаванием абстрактных закономерностей звукового потока, в том числе с использованием последовательностей с непостоянной временной структурой [122, 165, 166, 173].
С точки зрения изучения пространственного слуха, по-прежнему представляет интерес получение НР в условиях стимуляции в свободном звуковом поле. Сочетание нескольких локализационных признаков звука (межушной задержки по времени и по интенсивности, спектральных признаков и изменений интенсивности) позволит моделировать пространственные различия не только в пределах горизонтальной плоскости, но и в трехмерном пространстве. Исследования НР, вызванной движением звука в вертикальной плоскости, до сих пор не проводились. Анализ параметров НР в таких условиях может дать новые сведения об активации мозговых структур, обеспечивающих ориентацию человека и его адаптацию к пространственным акустическим изменениям.
Список литературы
Александров А.А., Бабанин М.Е. Негативность рассогласования в вызванных потенциалах при использовании акустических стимулов небольшой продолжительности // Физиология человека. 2000. Т. 26. № 6. С. 111–115.
Альтман Я.А. Локализация звука. Ленинград: Наука, 1972. 214 с.
Альтман Я.А. Локализация движущегося источника звука. Ленинград: Наука, 1983. 176 с.
Альтман Я.А. Пространственный слух. Санкт-Петербург: Институт физиологии им. И.П. Павлова РАН, 2011. 311 с.
Альтман Я.А., Вайтулевич С.Ф. Слуховые вызванные потенциалы человека и локализация источника звука. Санкт-Петербург: Наука, 1992. 136 с.
Альтман Я.А., Вайтулевич С.Ф., Петропавловская Е.А., Шестопалова Л.Б. Различение человеком динамических изменений пространственного положения звуковых образов (электрофизиологическое и психофизическое исследование) // Физиология человека. 2010. Т. 36. № 1. С. 1–9.
Вайтулевич С.Ф., Петропавловская Е.А., Шестопалова Л.Б., Никитин Н.И. Межполушарная асимметрия суммарной активности мозга человека при локализации источника звука // Сенсорные системы. 2015. Т. 29. № 1. С. 63–77.
Варфоломеев А.Л., Старостина Л.В. Слуховые вызванные потенциалы человека при иллюзорном движении звукового образа // Росс. физиол. жур. им. И.М. Сеченова. 2006. Т. 92. № 9. С. 1046.
Василенко Ю.А., Шестопалова Л.Б. Различение движущихся звуковых образов: негативность рассогласования и психофизическое тестирование // Физиология человека. 2010. Т. 36. № 2. С. 23–31.
Евстигнеева М.Д., Александров А.А. Негативность рассогласования: вклад различий в рефрактерности стимул-специфических нейрональных популяций // Росс. физиол. жур. им. И.М. Сеченова. 2008. Т. 94. № 10. С. 1147–1157.
Наатанен Р. Внимание и функции мозга. Под ред. Соколова Е.Н. Изд-во МГУ, 1998. 559 с.
Шестопалова Л.Б., Петропавловская Е.А., Вайтулевич С.Ф., Никитин Н.И. Объективные и субъективные показатели слухового различения движущихся стимулов // Физиология человека. 2014. Т. 40. № 3. С. 36–44.
Шестопалова Л.Б., Петропавловская Е.А., Вайтулевич С.Ф., Никитин Н.И. Оценка чувствительности слуховой системы к скорости движения звуковых стимулов: дифференциальные пороги и негативность рассогласования // Физиология человека. 2015. Т. 41. № 2. С. 14–22.
Шестопалова Л.Б., Петропавловская Е.А., Семенова В.В., Никитин Н.И. Негативность рассогласования при предъявлении амплитудно-модулированных звуковых сигналов // Журн. высшей нервной деятельности. 2018. Т. 2.
Шестопалова Л.Б., Петропавловская Е.А., Семенова В.В., Никитин Н.И., Вайтулевич С.Ф. Влияние акустического контекста на перцептивные различия пространственных звуковых сигналов // Физиология человека. 2017. Т. 43. № 6. С. 1–11.
Aaltonen O., Tuomainen J., Laine M., Niemi P. Cortical differences in tonal versus vowel processing as revealed by an ERP component called mismatch negativity (MMN) // Brain and Language. 1993. V. 44. P. 139–152.
Ahissar M., Ahissar E., Bergman H., Vaadia E. Encoding of sound-source location and movement: Activity of single neurons and interactions between adjacent neurons in the monkey auditory cortex // J. Neurophysiol. 1992. V. 67. P. 203–215.
Ahveninen J., Kopco N., Jääskeläinen I.P. Psychophysics and neuronal bases of sound localization in humans // Hear. Res. 2014. V. 307. P. 86–97. https://doi.org/10.1016/j.heares.2013.07.008
Alain C., Woods D.L., Knight R. A distributed cortical network for auditory sensory memory in humans // Brain Res. 1998. V. 812. P. 23–37.
Alain C., Woods D.L., Ogawa K.H. Brain indices of automatic pattern processing // Neuroreport. 1994. V. 6. № 1. P. 140–144.
Aleksandrov A.A., Babanin M.E., Stankevich L.N. Mechanisms of generation of mismatch negativity and their role in discrimination of short acoustical stimuli // Ross. Fiziol. Zh. Im. I.M. Sechenova. 2002. V. 88. № 7. P. 809–816.
Alexandrov A.A., Boricheva D.O., Pulvermüller F., Shtyrov Yu. 2011. Strength of word-specific neural memory traces assessed electrophysiologically // PLoS One. 2011. V. 6. № 8. e22999. https://doi.org/10.1371/journal.pone.0022999
Alho K. Cerebral generators of mismatch negativity (MMN) and its magnetic counterpart (MMNm) elicited by sound changes // Ear and Hearing. 1995. V. 16. P. 38–51.
Alho K., Rinne T., Herron T.J., Woods D.L. Stimulus-dependent activations and attention-related modulations in the auditory cortex: A meta-analysis of fMRI studies. // Hear. Res. 2014. V. 307. 29–41. https://doi.org/10.1016/j.heares.2013.08.001
Allen J., Kraus N., Bradlow A. Neural representation of consciously imperceptible speech sound differences // Percept Psychophys. 2000. V. 62. P. 1383-1393.
Altman J.A. Are there neurons detecting direction of sound source motion? // Exp. Neurol. 1968. V. 22. P. 13–25.
Altman J.A., Romanov V.P. Psychophysical characteristics of the auditory image movement perception during dichotic stimulation // Int. J. Neurosci. 1988. V. 38. P. 369–379.
Altman J.A., Vaitulevich S.Ph. Auditory image movement in evoked potential // Electroenceph. Clin. Neurophysiol. 1990. V. 75. P. 323–333.
Altman J.A., Vaitulevich S.Ph., Shestopalova L.B., Petropavlovskaia E.A. How does mismatch negativity reflect auditory motion? // Hear. Res. 2010. V. 268. P. 194–201.
Altman J.A., Vaitulevich S.Ph, Shestopalova L.B., Varfolomeev A.L. Mismatch negativity evoked by stationary and moving auditory images of different azimuthal positions // Neurosci. Lett. 2005. V. 384. P. 330–335.
Altman J.A., Viskov O.V. Discrimination of perceived movement velocity for fused auditory image in dichotic stimulation // J. Acoust. Soc. Am. 1977. V. 61. № 3. P. 816–819.
Bach D.R., Schächinger H., Neuhoff J.G., Esposito F., Di Salle F., Lehmann C., Herdener M., Scheffler K., Seifritz E. (2008). Rising sound intensity: An intrinsic warning cue activating the amygdala // Cereb. Cortex. 2008. V. 18. P. 145–150. https://doi.org/10.1093/cercor/bhm040
Barcelo F., Escera C., Corral M.J., Periáñez J.A. Task switching and novelty processing activate a common neural network for cognitive control // J. Cognitive Neurosci. 2006. V. 18. P. 1734–1748.
Baumgart F., Gaschler-Markefski B., Woldorff M.G., Heinze H.J., Schleich H. A movement-sensitive area in auditory cortex // Nature. 1999. V. 400. P. 724–726.
Bendixen A. (2014). Predictability effects in auditory scene analysis: A review // Front Neurosci. 2014. V. 8. P. 60. https://doi.org/10.3389/fnins.2014.00060
Bernstein L.R., Trahiotis C., Akeroyd M.A., Hartung K. Sensitivity to brief changes of interaural time and interaural intensity // J. Acoust. Soc. Am. 2001. V. 109. P. 1604–1615.
Berti S., Schröger E. A comparison of auditory and visual distraction effects: behavioural and event-related indices // Cogn. Brain Res. 2001. V. 10. P. 265–273.
Bertoli S., Heimberg S., Smurzynski J., Probst R. Mismatch negativity and psychoacoustic measures of gap detection in normally hearing subjects // Psychophysiology. 2001. V. 38. P. 334–342.
Bidet-Caulet A., Bertrand O. Dynamics of a temporo-fronto-parietal network during sustained spatial or spectral auditory processing // J. Cogn. Neurosci. 2005. V. 17. № 11. P. 1691–1703.
Blauert J. On the lag of lateralization caused by interaural time and intensity differences // Audiology. 1972. V. 11. P. 265–270.
Bradlow A.R., Kraus N., Nicol T.G., McGee T.J., Cunningham J., Zecker S.G. Effects of lengthened formant transition duration on discrimination and neural representation of synthetic CV syllables by normal and learning-disabled children // J. Acoust. Soc. Am. 1999. V. 106. P. 2086–2096.
Bremmer F., Schlack A., Shah N.J., Zafiris O., Kubischik M., Hoffmann K., Zilles K., Fink G.R. Polymodal motion processing in posterior parietal and premotor cortex: a human fMRI study strongly implies equivalencies between humans and monkeys // Neuron. 2001. V. 29. № 1. P. 287–296.
Briley P.M., Kitterick P.T., Summerfield A.Q. Evidence for Opponent Process Analysis of Sound Source Location in Humans // JARO. 2013. V. 14. P. 83–101. https://doi.org/10.1007/s10162-012-0356-x
Brunetti M., Belardinelli P., Caulo M., Del Gratta C., Della Penna S., Ferretti A., Lucci G., Moretti A., Pizzella V., Tartaro A., Torquati K., Olivetti Belardinelli M., Romani G.L. Human brain activation during passive listening to sounds from different locations: an fMRI and MEG study // Hum. Brain Mapp. 2005. V. 26. P. 251–261.
Deouell L.Y., Bentin S., Giard M.H. Mismatch negativity in dichotic listening: Evidence for interhemispheric differences and multiple generators // Psychophysiol. 1998. V. 35. № 4. P. 355–365.
Carlile S., Best V. Discrimination of sound source velocity in human listeners // J. Acoust. Soc. Am. 2002. V. 111. № 2. P. 1026–1035. https://doi.org/10.1121/1.1436067
Colin C., Radeau M., Soquet A, Dachy B., Deltenre P. Electrophysiology of spatial scene analysis: the mismatch negativity (MMN) is sensitive to ventriloquism illusion // Clin. Neurophysiol. 2002. V. 113. P. 507–518.
Coull J.T. Neural correlates of attention and arousal: insights from electrophysiology, functional neuroimaging and psychopharmacology // Prog. Neurobiol. 1998. V. 55. P. 343–361.
Cowan N., Winkler I., Teder W., Näätänen R. Memory prerequisites of the mismatch negativity in the auditory event-related potential (ERP) // J. Exp. Psychol. Learn. Mem. Cogn. 1993. V. 19. P. 909–921.
Culling J.F., Summerfield Q. Measurements of the binaural temporal window using a detection task // J. Acoust. Soc. Am. 1998. V. 103. P. 3540–3553.
Dalebout S., Stack J. Mismatch negativity to acoustic differences not differentiated behaviorally // J. Am. Acad. Audiol. 1999. V. 10. P. 388–399.
Deouell L.Y., Bentin S., Giard M. Mismatch negativity in dichotic listening: evidence for interhemispheric differences and multiple generators // Psychophysiology. 1998. V. 35. P. 355–365.
Deouell L.Y., Bentin S., Soroker N. Electrophysiological evidence for an early (pre-attentive) information processing deficit in patients with right hemisphere damage and unilateral neglect // Brain. 2000. V. 123. № 2. P. 353–365.
Deouell L.Y., Parnes A., Pickard N., Knight R. Spatial locations accurately tracked by human auditory sensory memory: evidence from the mismatch negativity // Eur. J. Neurosci. 2006. V. 24. P. 1488–1494.
Devlin J.T., Raley J., Tunbridge E., Lanary K., Floyer-Lea A., Narain C., Cohen I., Behrens T., Jezzard P., Matthews P.M., Moore D.R. Functional asymmetry for auditory processing in human primary auditory cortex // J. Neurosci. 2003. V. 23 № 37. P. 11516–11522.
Doeller C.F., Opitz B., Mecklinger A., Krick C., Reith W., Schröger E. Prefrontal cortex involvement in preattentive auditory deviance detection: neuroimaging and electrophysiological evidence // NeuroImage. 2003. V. 20. P. 1270–1282.
Ducommun C.Y., Murray M.M., Thut G., Bellmann A., Viaud-Delmon I., Clarke S., Michel C.M. Segregated processing of auditory motion and auditory location: an ERP mapping study // NeuroImage. 2002. V. 16. № 1. P. 76–88.
Escera C., Alho K., Winkler I., Näätänen R. Neural mechanisms of involuntary attention to acoustic novelty and change // J. Cogn. Neurosci. 1998. V. 10. № 5. P. 590–604.
Escera C., Corral M.-J. The distraction potential (DP), an electrophysiological tracer of involuntary attention control and its dysfunction. In: Reinvang I., Greenlee M. W., Herrmann M. (Eds) The Cognitive Neuroscience of Individual Differences – New Perspectives (p. 63–76). Bibliotheks-und Informationssystem der Universität. Oldenburg. Germany. 2003.
Escera C., Corral M.J. Role of mismatch negativity and novelty-P3 in involuntary auditory attention // J. Psychophysiol. 2007. V. 21. P. 251–264.
Ford J.M., Hillyard S.A. Event-related potentials, ERPs, to interruptions of steady rhythm // Psychophysiology. 1981. V. 18. P. 322–330.
Friedman D., Cycowicz Y.M., Gaeta H. The novelty P3: an event-related brain potential (ERP) sign of the brain’s evaluation of novelty // Neurosci. Biobehav. 2001. R. V. 25. P. 355–373.
Friston K. A theory of cortical responses // Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005. V. 360. P. 815–836. https://doi.org/10.1098/rstb.2005.1622
Fujiki N., Riederer K.A.J., Jousmäki V., Mäkelä J.P., Hari R. Human cortical representation of virtual auditory space: differences between sound azimuth and elevation // Eur. J. Neurosci. 2002. V. 16. P. 2207–2213.
Getzmann S. Effects of velocity and motion-onset delay on detection and discrimination of sound motion // Hear. Res. 2008. V. 246. P. 44-51.
Getzmann S. Effect of auditory motion velocity on reaction time and cortical processes // Neuropsychol. 2009. V. 47. P. 2625-2633.
Getzmann S. Auditory motion perception: onset position and motion direction are encoded in discrete processing stages // Eur. J. Neurosci. 2011. V. 33. P. 1339–1350. https://doi.org/10.1111/j.1460-9568.2011.07617.x
Getzmann S., Lewald J. Effects of natural versus artificial spatial cues on electrophysiological correlates of auditory motion // Hear. Res. 2010. V. 259. P. 44–54. https://doi.org/10.1016/j.heares.2009.09.021
Giard M.H., Perrin F., Pernier J., Bouchet P. Brain generators implicated in the processing of auditory stimulus deviance: a topographic event-related study // Psychophysiol. 1990. V. 27. P. 627–640.
Grantham D.W. Detection and discrimination of simulated motion of auditory targets in the horizontal plane // J. Acoust. Soc. Am. 1986. V. 79. № 6. P. 1939-1949.
Grantham D.W., Wightman F.L. Detectability of varying interaural temporal differences // J. Acoust. Soc. Am. 1978. V. 63. P. 511–523.
Griffiths T.D., Bench C.J., Frackowiak R.S. Human cortical areas selectively activated by apparent sound motion // Curr. Biol. 1994. V. 4. P. 892–895.
Griffiths T.D., Green G.G., Rees A., Rees G. Human brain areas involved in the analysis of auditory movement // Hum. Brain Mapp. 2000. V. 9. P. 72–80.
Griffiths T.D., Rees G., Rees A., Green G.G., Witton C., Rowe D., Büchel C., Turner R., Frackowiak R.S. Right parietal cortex is involved in the perception of sound movement in humans // Nat. Neurosci. 1998. V. 1. P. 74–79.
Hall D.A., Hart H.C., Johnsrude I.S. Relationships between human auditory cortical structure and function // Audiol. Neurootol. 2003. V. 8. № 1. P. 1–18.
Hari R., Hämäläinen M., Ilmoniemi R., Kaukoranta E., Reinikainen K., Salminen et al. Responses of the primary auditory cortex to pitch changes in a sequence of tone pips: Neuromagnetic recordings in man // Neurosci. Lett. 1984. V. 50. P. 127–132.
Hart H.C., Palmer A.R., Hall D.A. Different areas of human non-primary auditory cortex are activated by sounds with spatial and nonspatial properties // Hum. Brain Mapp. 2004. V. 21. P. 178–190.
Hopfinger J.B., Parks E.L. Involuntary attention. In Mangun G.R. (Eds), The neuroscience of attention: Attentional control and selection (p. 30–53). New York: Oxford University Press. 2012.
Horváth J., Winkler I. Distraction in a continuous-stimulation detection task // Biol. Psychol. 2010. V. 83. P. 229–238.
Horváth J., Winkler I., Bendixen A. Do N1/MMN, P3a, and RON form a strongly coupled chain reflecting the three stages of auditory distraction? // Biol. Psychol. 2008. V. 79. P. 139–147.
Jääskeläinen I.P., Ahveninen J., Bonmassar G., Dale A.M., Ilmoniemi R.J., Levänen S. et al. Human posterior auditory cortex gates novel sounds to consciousness // PNAS. 2004. V. 17. P. 6809–6814.
Jacobsen T., Schröger E. Is there preattentive memory-based comparison of pitch? // Psychophysiol. 2001. V. 38. P. 723–727.
Jacobsen T., Schröger, E. Measuring duration mismatch negativity // Clin. Neurophysiol. 2003. V. 114. P. 1133–1143.
Jäkel F., Wichmann F. A. Spatial four-alternative forced-choice method is the preferred psychophysiological method for naïve observers // J. Vis. 2006. V. 6. P. 1307–1322. https://doi.org/10.1167/6.11.13
Jäncke L., Wustenberg T., Schulze K., Heinze H. J. Asymmetric hemodynamic responses of the human auditory cortex to monaural and binaural stimulation // Hear. Res. 2002. V. 170. № 1–2. P. 166–178.
Jaramillo M., Ilvonen T., Kujala T., Alku P., Tervaniemi M., Alho K. Are different kinds of acoustic features processed differently for speech and non-speech sounds? // Cogn. Brain Res. 2001. V. 12. P. 459-466.
Jerger J., Estes R. Asymmetry in event-related potentials to simulated auditory motion in children, young adults, and seniors // J. Am. Acad. Audiol. 2002. V. 13. № 1. P. 1–13.
Johnson B.W., Hautus M.J. Processing of binaural spatial information in human auditory cortex: neuromagnetic responses to interaural timing and level differences // Neuropsychol. 2010. V. 48. P. 2610–2619.
Kaczmarek T., Niewiarowicz M. Auditory motion perception in normal hearing and in hearing impaired people // Acta. Acust. United Ac. 2013. V. 99. P. 283–291.
Kaiser J., Lutzenberger W. Location changes enhance hemispheric asymmetry of magnetic fields evoked by lateralized sounds in humans // Neurosci. Lett. 2001. V. 314. P. 17–20.
Kaiser J., Lutzenberger W., Preissl H., Ackermann H., Birbaumer N. Right-hemisphere dominance for the processing of sound-source lateralization // J. Neurosci. 2000. V. 20. P. 6631–6639.
Koelsch S., Schröger E., Tervaniemi M. (). Superior attentive and preattentive auditory processing in musicians // Neuroreport. 1999. V. 10. P. 1309–1313.
Kollmeier B., Gilkey R.H. Binaural forward and backward masking: Evidence for sluggishness in binaural detection // J. Acoust. Soc. Am. 1990. V. 87. P. 1709–1719.
Kraus N., McGee T.J., Carrell T.D., Zecker S.G., Nicol T.G., Koch D.B. Auditory neurophysiologic responses and discrimination deficits in children with learning problems // Science. 1996. V. 273. P. 971–973.
Krumbholz K., Hewson-Stoate N., Schönwiesner M. Cortical response to auditory motion suggests an asymmetry in the reliance on inter-hemispheric connections between the left and right auditory cortices // J. Neurophysiol. 2007. V. 97. P. 1649-1655.
Krumbholz K., Shönwiesner M., von Cramon D.Y., Rübsamen R., Shah N.J., Zilles K., Fink G.R. Representation of interaural temporal information from left and right auditory space in the human planum temporale and inferior parietal lobe // Cereb. Cortex. 2005. V. 15. P. 317–324.
Kujala T., Tervaniemi M., Schröger E. The mismatch negativity in cognitive and clinical neuroscience: Theoretical and methodological considerations // Biol. Psychol. 2007. V. 74. P. 1-19.
Lang H.A., Nyrke T., Ek M., Aaltonen O., Raimo I., Näätänen R. Pitch discrimination performance and auditory event related potentials. In Brunia C.H.M., Gaillard A.W.K., Kok A., Mulder G., & Verbaten M.N. (Eds), Psychophysiological brain research (Vol. 1, p. 294–298). Tilburg, The Netherlands: Tilburg University Press. 1990.
Levänen S., Ahonen A., Hari R., McEvoy L., Sams M. Deviant auditory stimuli activate human left and right auditory cortex differently // Cereb. Cortex. 1996. V. 6. P. 288–296.
Lewald J., Riederer K.A., Lentz T., Meister I.G. Processing of sound location in human cortex // Eur. J. Neurosci. 2008. V. 27. № 5. P. 1261–1270. https://doi.org/10.1111/j.1460-9568.2008.06094.x
Lewis J.W., Beauchamp M.S., DeYoe E.A. A comparison of visual and auditory motion processing in human cerebral cortex // Cereb. Cortex. 2000. V. 10. № 9. P. 873–888.
May P.J.C., Tiitinen H. Mismatch negativity (MMN), the deviance-elicited auditory deflection, explained // Psychophysiol. 2010. V. 47. P. 66–122.
Mäkelä J.P., McEvoy L. Auditory evoked fields to illusory sound source movements // Exp. Brain Res. 1996. V. 110. № 3. P. 446–454.
McEvoy L., Hari R., Imada T., Sams M. Human auditory cortical mechanisms of sound lateralization. II. Interaural time differences at sound onset // Hear. Res. 1993. V. 67. P. 98–109.
Mesulam M.M. Spatial attention and neglect: parietal, frontal and cingulate contributions to the mental representation and attentional targeting of salient extrapersonal events // Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1999. V. 354. P. 1325–1346.
Middlebrooks J.C., Green D.M. Sound localization by human listeners // Annu. Rev. Psychol. 1991. V. 42. P. 135–159.
Molholm S., Gomes H., Lobosco J., Deacon D., Ritter W. Feature versus gestalt representation of stimuli in the mismatch negativity system of 7- to 9-year-old children // Psychophysiol. 2004. V. 41. P. 385–393.
Näätänen R. The role of attention in auditory information processing as revealed by event-related potentials and other brain measures of cognitive function // The Behavioral and Brain Sciences. 1990. V. 13. P. 201–233.
Näätänen R. Attention and brain function. Hillsdale, NJ: Erlbaum. 1992.
Näätänen R., Astikainen P., Ruusivirta T., Huotilainen M. Automatic auditory intelligence: An expression of the sensory–cognitive core of cognitive processes // Brain Res. Rev. 2010. V. 64. P. 123–136.
Näätänen R., Gaillard A.W.K., Mäntysalo S. Early selective attention effect on evoked potential reinterpreted // Acta Psychologica. 1978. V. 42. P. 313–329.
Näätänen R., Jacobsen T., Winkler I. Memory-based or afferent processes in mismatch negativity (MMN): A review of the evidence // Psychophysiol. 2005. V. 42. P. 25–32.
Näätänen R., Kujala T., Winkler I. Auditory processing that leads to conscious perception: A unique window to central auditory processing opened by the mismatch negativity and related responses // Psychophysiol. 2011. V. 48. P. 4–22.
Näätänen R., Paavilainen P., Rinne T., Alho K. The mismatch negativity (MMN) in basic research of central auditory processing: A review // Clin. Neurophysiol. 2007. V. 118. P. 2544–2590.
Näätänen R., Pakarinen S., Rinne T., Takegata R. The mismatch negativity (MMN): towards the optimal paradigm // Clin. Neurophysiol. 2004. V. 115. P. 140–144.
Näätänen R., Picton T.W. The N1 wave of the human electric and magnetic response to sound: A review and an analysis of the component structure // Psychophysiol. 1987. V. 24. P. 375–425.
Näätänen R., Schröger E., Karakas S., Tervaniemi M., Paavilainen P. Development of a memory trace for a complex sound in the human brain // NeuroReport. 1993. V. 4. P. 503–506.
Näätänen R., Winkler I. The concept of auditory stimulus representation in cognitive neuroscience // Psychol Bull. 1999. V. 125. P. 826–859.
Nager W., Kohlmetz C., Joppich., Möbes J., Münte T. Tracking of multiple sound sources defined by interaural time differences: brain potential evidence in human // Neurosci. Lett. 2003. V. 344. P. 181–184.
Nordby H., Roth W.T., Pfefferbaum A. Event-related potentials to time deviant and pitch deviant tones // Psychophysiol. 1988. V. 25 P. 249–261.
Opitz B., Rinne T., Mecklinger A., von Cramon D.Y., Schröger E. Differential contribution of frontal and temporal cortices to auditory change detection: fMRI and ERP results // Neuroimage. 2002. V. 15. P. 167–174.
Paavilainen P. The mismatch-negativity (MMN) component of the auditory event-related potential to violations of abstract regularities: A review // Int. J. Psychophysiol. 2013. V. 88. P. 109–123.
Paavilainen P., Alho K., Reinikainen K., Sams M., Näätänen R. Right hemisphere dominance of different mismatch negativities // Electroenceph. Clin. Neurophysiol. 1991. V. 78. P. 466–479.
Paavilainen P., Arajärvi P., Takegata R. Preattentive detection of nonsalient contingencies between auditory features // NeuroReport. 2007. V. 18. P. 159–163.
Paavilainen P., Karlsson M.L., Reinikainen K., Näätänen R. Mismatch negativity to change in spatial location of an auditory stimulus // Electroenceph. Clin. Neurophysiol. 1989. V. 73. P. 129–141.
Paavilainen P., Jiang D., Lavikainen J., Naatanen R. Stimulus duration and the sensory memory trace: Aп event-related potential study // Biol Psychology. 1993. V. 35. P. 139.
Paavilainen P., Simola J., Jaramillo M., Näätänen R., Winkler I. Preattentive extraction of abstract feature conjunctions from auditory stimulation as reflected by the mismatch negativity (MMN) // Psychophysiol. 2001. V. 38. P. 359–365.
Pakarainen S., Takegata R., Rinne T., Huotilainen M., Näätänen R. Measurement of extensive auditory discrimination profiles using the mismatch negativity (MMN) of the auditory event-related potential (ERP) // Clin. Neurophysiol. 2007. V. 118. P. 177–185.
Palomäki K.J., Tiitinen H., Mäkinen V., May P.J., Alku P. Spatial processing in human auditory cortex: the effects of 3D, ITD, and ILD stimulation techniques // Brain Res. Cogn. Brain Res. 2005. V. 24. P. 364–379.
Pavani F., Macaluso E., Warren J.D., Driver J., Griffiths T.D. A common cortical substrate activated by horizontal and vertical sound movement in the human brain // Curr. Biol. 2002. V. 12. P. 1584–1590.
Peter V., McArthur G., Thompson W.F. Effect of deviance direction and calculation method on duration and frequency mismatch negativity (MMN) // Neurosci. Lett. 2010. V. 482. P. 71–75. https://doi.org/10.1016/j.neulet.2010.07.010
Petit L., Simon G., Joliot M., Andersson F., Bertin T., Zago L., Mellet E., Tzourio-Mazoyer N. Right hemisphere dominance for auditory attention and its modulation by eye position: an event-related fMRI study // Restor. Neurol. Neurosci. 2007. V. 25 № 3–4. P. 211–225.
Pettigrew C.M., Murdoch B.E., Ponton C.W., Finnigan S., Alku P., Kei J. Automatic auditory processing of English words as indexed by the mismatch negativity, using a multiple deviant paradigm // Ear Hear. 2004. V. 25. P. 284–301.
Phillips D.P., Brugge J.F. Progress in neurophysiology of sound localization // Annu. Rev. Psychol. 1985. V. 36. P. 245–274.
Polich J. Updating P300: an integrative theory of P3a and P3b // Clin. Neurophysiol. 2007. V. 118. P. 2128–2148.
Richter N., Schröger E., Rübsamen R. Hemispheric specialization during discrimination of sound sources reflected by MMN // Neuropsychol. 2009. V. 47. P. 2652–2659.
Rinne T., Alho K., Ilmoniemi R.J., Virtanen J., Näätänen R. Separate time behaviors of the temporal and frontal mismatch negativity sources // Neuroimage. 2000. V. 12. P. 14–19.
Rinne T., Sarkka A., Degerman A., Schröger E., Alho K. Two separate mechanisms underlie auditory change detection and involuntary control of attention // Brain Res. 2006. V. 1077. P. 135–143.
Ruhnau Ph., Herrmann B., Schröger E. Finding the right control: The mismatch negativity under investigation // Clin. Neurophysiol. 2012. V. 123. P. 507–512. https://doi.org/10.1016/j.clinph.2011.07.035
Saberi K., Hafter E.R. Experiments on Auditory Motion Discrimination. In Gilkey R.H., & Anderson T.R. (Eds), Binaural and spatial hearing in real and virtual environments (p. 315–327). NJ: Erlbaum Publ. 1997.
Salminen N.H., Tiitinen H., Miettinen I., Alku P., May P.J. Asymmetrical representation of auditory space in human cortex // Brain Res. 2010. V. 1306. P. 93–99.
Savela J., Kujala T., Tuomainen J., Ek M., Aaltonen O., Näätänen R. The mismatch negativity and reaction time as indices of the perceptual distance between the corresponding vowels of two related languages // Cogn. Brain Res. 2003. V. 16. P. 250-256.
Schonwiesner M., Krumbholz K., Rubsamen R., Fink G.R., von Cramon D.Y. Hemispheric asymmetry for auditory processing in the human auditory brain stem, thalamus, and cortex // Cereb. Cortex. 2007. V. 17. P. 492–499.
Schröger E. Interaural time and level differences: integrated or separated processing? // Hear. Res. 1996. V. 96. P. 191–198.
Schröger E., Giard M.H., Wolff C. Auditory distraction: event-related potential and behavioural indices // Clin. Neurophysiol. 2000. V. 111. P. 1450–1460.
Schröger E., Tervaniemi M., Winkler I., Wolff C., Näätänen R. Processing of interaural cues used for auditory lateralization by the mismatch negativity. Contribution to Psychological Acoustics (p. 49–56). Bibliotheks-und Informationssystem der Carl von Ossietzky Universität Oldenburg. 1997.
Schröger E., Wolff C. Mismatch response to changes in sound location // NeuroReport. 1996. V. 7. P. 3005–3008.
Schröger E., Wolff C. Attentional orienting and reorienting is indicated by human event-related brain potentials // Neuroreport. 1998a. V. 9. P. 3355–3358.
Schröger E., Wolff C. Behavioural and electrophysiological effects of task-irrelevant sound change: a new distraction paradigm // Cogn. Brain. Res. 1998б. V. 7. P. 71–87.
Shestopalova L.B., Petropavlovskaia E.A., Vaitulevich S.Ph., Nikitin N.I. Hemispheric asymmetry of ERPs and MMNs evoked by slow, fast and abrupt auditory motion // Neuropsychologia. 2016. V. 91. P. 465–479. https://doi.org/10.1016/j.neuropsychologia.2016.09.011
Shestopalova L.B., Petropavlovskaia E.A., Semenova V.V., Nikitin N.I. Mismatch negativity and psychophysical detection of rising and falling intensity sounds // Biol. Psychol. 2018.
Shestopalova L.B., Petropavlovskaia E.A., Vaitulevich S.Ph., Nikitin N.I. Contextual effects on preattentive processing of sound motion as revealed by spatial MMN // Int. J. Psychophysiol. 2015. V. 96 P. 49–56. https://doi.org/10.1016/j.ijpsycho.2015.02.021
Shestopalova L.B., Petropavlovskaia E.A., Vaitulevich S.Ph., Vasilenko Yu.A., Nikitin N.I., Altman J.A. Discrimination of auditory motion patterns: mismatch negativity study // Neuropsychol. 2012. V. 50. P. 2720–2729.
Sonnadara R., Alain C., Trainor L.J. Occasional changes in sound location enhance middle latency evoked responses // Brain Res. 2006. V. 1076. P. 187–192.
Spierer L., Bellmann-Thiran A., Maeder pH, Murray M.M., Clarke S. Hemispheric competence for auditory spatial representation // Brain. 2009. V. 132. P. 1953–1966. https://doi.org/10.1093/brain/awp127
Spitzer M.W., Semple M.N. Interaural phase coding in auditory midbrain: influence of dynamic stimulus features // Science. 1991. V. 254 № 5032. P. 721–724.
Sussman E. S., Chen S., Sussman-Fort J., Dinces E. The Five Myths of MMN: Redefining how to use MMN in basic and clinical research // Brain Topogr. 2014. V. 27. № 4. P. 553–564. .https://doi.org/10.1007/s10548-013-0326-6
Takegata R., Huotilainen M., Rinne T., Näätänen R., Winkler I. Changes in acoustic features and their conjunctions are processed by separate neuronal populations // Neuroreport. 2001. V. 12. P. 525–529.
Tanaka H., Hachisuka K., Ogata H. Sound lateralization in patients with left or right cerebral hemispheric lesions :relation with unilateral visuospatial neglect // J. Neurol. Neurosurg. Psychiatry. 1999. V. 67. P. 481–486.
Tata M.S., Ward L.M. Early phase of spatial mismatch negativity is localized to a posterior “where” auditory pathway // Exp. Brain Res. 2005. V. 167. P. 481–486.
Tervaniemi M., Just V., Koelsch S., Widmann A., Schröger E. Pitch-discrimination accuracy in musicians vs. non-musicians: an event-related potential and behavioral study // Exp. Brain Res. 2005. V. 161. P. 1–10.
Teshiba T.M., Ling J., Ruhl D.A., Bedrick B.S., Peña A., Mayer A.R. Evoked and Intrinsic Asymmetries during Auditory Attention: Implications for the Contralateral and Neglect Models of Functioning // Cereb. Cortex. 2013. V. 23. № 3. P. 560–569. https://doi.org/10.1093/cercor/bhs039
Tiitinen H., May P., Reinikainen K., Näätänen R. Attentive novelty detection in humans is governed by preattentive sensory memory // Nature. 1994. V. 372. P. 90–92.
Tiitinen H., Salminen N.H., Palomaki K.J., Makinen V.T., Alku P., May P.J. Neuromagnetic recordings reveal the temporal dynamics of auditory spatial processing in the human cortex // Neurosci. Lett. 2006. V. 396. P. 17–22.
Todd J., Heathcote A., Whitson L.R., Mullens D., Provost A., Winkler I. Mismatch negativity (MMN) to pitch change is susceptible to order-dependent bias // Front. Neurosci. 2014. V. 8. P. 180. https://doi.org/10.3389/fnins.2014.00180
Todd J., Provost A., Whitson L.R., Cooper G., Heathcote A. Not so primitive: Context-sensitive meta-learning about unattended sound sequences // J. Neurophysiol. 2013. V. 109. № 1. P. 99–105.
Toronchuk J.M., Stumpf E., Cynader M.S. Auditory cortex neurons sensitive to correlates of auditory motion: underlying mechanisms // Exp. Brain Res. 1992. V. 88. P. 169–180.
Ungan R., Yagcioglu S., Goksoy C. Differences between the N1 wave of the responses to interaural time and intensity disparities: scalp topography and dipole sources // Clin. Neurophysiol. 2001. V. 112. P. 485–498.
Warren J.D., Zielinski B.A., Green G.G., Rauschecker J.P., Griffiths T.D. Perception of sound-source motion by the human brain // Neuron. 2002. V. 34. № 1. P. 139–148.
Waugh W., Strybel T.Z., Perrott D.R. Perception of moving sounds: velocity discrimination // J. Aud. Res. 1979. V. 19. P. 103–110.
Winkler I. Interpreting the Mismatch Negativity // J. Psychophysiol. 2007. V. 21. №3–4. P. 147–163.
Winkler I., Korzykov O., Gumenyuk V., Cowan N., Linkenkaer-Hansen K., Alho K. et al. Temporary and longer retention of acoustic information // Psychophysiol. 2002. V. 39. P. 530–534.
Winkler I., Schröger E. Auditory perceptual objects as generative models: Setting the stage for communication by sound // Brain Lang. 2015. V. 148. P. 1–22. https://doi.org/10.1016/j.bandl.2015.05.003
Winkler I., Tervaniemi M., Näätänen R. Two separate codes for missing fundamental pitch in the auditory cortex // J. Acoust. Soc. Am. 1997. V. 102. P. 1072–1082.
Winkler I., Tervaniemi M., Schröger E., Wolff C., Näätänen R. Preattentive processing of auditory spatial information in human // Neurosci. Lett. 1998. V. 242. P. 49–52.
Woldorff M.G., Tempelmann C., Fell J., Tegeler C., Gaschler-Markefski B. Hinrichs H., Heinz H.J., Scheich H. Lateralized auditory spatial perception and the contralaterality of cortical processing as studied with functional magnetic resonance imaging and magnetoencephalography // Hum. Brain Mapp. 1999. V. 7. № 1. P. 49–66.
Xiang J., Chuang S., Wilson D., Otsubo H., Pang E., Holowka S., Sharma R., Ochi A., Chitoku S. Sound motion evoked magnetic fields // Clin. Neurophysiol. 2002. V. 113. P. 1–9.
Zatorre R.J., Mondor T.A., Evans A.C. Auditory attention to space and frequency activates similar cerebral systems // NeuroImage. 1999. V. 10. P. 544–554.
Zatorre R.J., Penhune V.B. Spatial localization after excision of human auditory cortex // J. Neurosci. 2001. V. 21. P. 6321–6328.
Zimmer U., Lewald J., Erb M., Karnath H.O. Processing of auditory spatial cues in human cortex: An fMRI study // Neuropsychol. 2006. V. 44. P. 454–461.
Zimmer U., Macaluso E. High binaural coherence determines successful sound localization and increased activity in posterior auditory areas // Neuron. 2005. V. 47. P. 893–905.
Дополнительные материалы отсутствуют.
Инструменты
Успехи физиологических наук