

Успехи физиологических наук, 2019, T. 50, № 4, стр. 3-13
Перекрестная адаптация: от Ф.З. Меерсона до наших дней. Часть 1. Адаптация, перекрестная адаптация и перекрестная сенсибилизация
М. Ю. Зенько a, *, Е. А. Рыбникова a, **
a ФГБУН Институт физиологии им. И.П. Павлова РАН, Лаборатория регуляции функций нейронов мозга
199034 Санкт-Петербург, Россия
* E-mail: zenkomichail@mail.ru
** E-mail: rybnikova1@rambler.ru
Поступила в редакцию 20.03.2019
После доработки 30.04.2019
Принята к публикации 24.05.2019
Аннотация
Обзор посвящен обсуждению феномена перекрестной адаптации организма человека и животных к повреждающим факторам. В обзоре в лаконичной форме излагаются классические представления об адаптации, перекрестной адаптации и перекрестной сенсибилизации, а также тенденции их развития на современном этапе. Раскрывается значительный вклад российских физиологических школ в создание концепции о перекрестной адаптации. Обобщаются накопленные литературные и собственные данные о воздействиях, приводящих к формированию кросс-толерантности к различным неблагоприятным факторам. На примере протективных воздействий умеренной гипоксией демонстрируется зависимость эффектов от используемых форм гипоксии и ее режимов. Обзор завершается рассмотрением практических аспектов, касающихся возможностей и перспектив применения способов, направленных на активацию механизмов перекрестной адаптации, для профилактики и коррекции нарушений функций организма, возникающих в результате неблагоприятных воздействий различной модальности.
Термин “адаптация” происходит от латинского “adaptio”, что означает “приспособление”. Классическое определение гласит, что адаптация – это выработанное в процессе эволюционного развития приспособление биологической системы к условиям среды обитания [1]. Иными словами, под адаптацией подразумевалось приспособление к внешним факторам, однако это понятие интуитивно расширилось и включает в себя также адаптацию к ряду внутренних повреждающих факторов, в том числе гипоксии\ишемии, окислительному стрессу, токсическому действию нейромедиаторов и гормонов. На современном языке адаптация – это комплекс приспособительных реакций организма к изменяющимся условиям внешней и внутренней среды.
Согласно классику теории адаптации Ф.З. Меерсону, в адаптивной реакции организма можно выделить два последовательных этапа: это “срочная”, или “несовершенная” адаптация, и “совершенная” долговременная адаптации [25]. Первый этап адаптивной реакции, возникающий сразу после начала действия стрессора, может реализовываться только на базе имеющихся, заранее сформированных, физиологических механизмов. Примерами подобной “срочной” адаптации являются, например, реакция кардиореспираторой системы [2] и активация гипоксия-индуцибельных факторов семейства HIF в условиях гипоксии [22], увеличение теплопродукции организма в условиях низких температур [7], и теплоотдачи при повышенных. Однако, первый “срочный” этап, как правило, не обеспечивает проявление необходимого адаптационного эффекта в полной мере.
Реакция на любое новое и достаточно сильное воздействие среды – нарушение гомеостаза, реальное или вероятное, является системным процессом и обеспечивается, во-первых, системой, специфически реагирующей на данный раздражитель, и, во-вторых, неспецифическими стресс-реализующими симпатоадреналовой и гипоталамо-гипофизарно-адренокортикальной (ГГАС) системами. Неспецифический ответ представляет собой по сути общий адаптационный синдром Г. Селье [77], а специфический ответ оставляет на молекулярном уровне свой “структурный след”, как называет его Ф.З. Меерсон, который, при длительной или многократной мобилизации этой функциональной системы, приводит к увеличению ее функциональной мощности, и создает возможность превращения первоначальной, “срочной”, адаптации в устойчивую, “долговременную”.
Из-за постепенного количественного накопления изменений на молекулярном уровне организм становится адаптированным к этому неблагоприятному фактору, или толерантным, приобретая устойчивость к условиям, которые ранее были экстремальными или несовместимыми с жизнью – гипоксия, холод, повышенные температуры [43, 61, 62, 64]. Развиваются новые поведенческие реакции на стрессоры, реализуемые уже на высших когнитивных уровнях. Данный переход от “срочного” этапа к “долговременному”, по-видимому, представляет собой ключевой этап адаптационного процесса, так как именно он делает возможным длительное выживание организма, расширяет границы его комфортных условий и ареал.
Системный структурный “след” обеспечивает экономичность функционирования систем, ответственных за адаптацию; каждый неблагоприятный фактор запускает развитие своего специфичного “молекулярного следа”, при этом конечные эффекторы различных факторов могут частично быть общими, приводя к развитию или ослаблению толерантности к другим факторам, то есть возникновению перекрестной адаптации. Если при этом резистентность к другим факторам возрастает, то речь идет о перекрестной адаптации, а если толерантность снижается – о перекрестной сенсибилизации.
1. ПОНЯТИЕ О ПЕРЕКРЕСТНОЙ АДАПТАЦИИ
У каждого животного присутствует неотъемлемая способность справляться со стрессорами в форме приспособления, адаптации к нему. При этом наблюдается снижение чувствительности звеньев стрессорного ответа, таких как гипоталамо-гипофизарно-адренокортикальная система (ГГАС), во время повторных эпизодов воздействия того же стрессора на организм [42, 67]. Воздействие того же самого стрессора называется гомотипическим. Помимо такого типа стрессорной адаптации, существует еще одно хорошо документированное физиологическое и биохимическое явление, известное как перекрестная или “кросс-адаптация”, при котором наблюдается снижение стрессовой реакции на новый стрессор у ранее адаптированного к другому стрессору организма [58, 60]. Тем самым адаптивные изменения, вызванные одним стрессором, могут сделать организм более устойчивым к неблагоприятным последствиям другого типа стрессора [55, 60]. Организмы со сформировавшейся перекрестной адаптацией обладают лучшим ответом на стресс сразу на разных физиологических уровнях [64]. Клинические и доклинические исследования предполагают наличие явления перекрестной адаптации как с системными, так и эмоциональными стрессорами. Например, показана положительная перекрестная адаптация между высотной гипоксией и тепловым шоком [68], плаванием и электрошоковым раздражением хвоста [79], гипоксией, холодовым стрессом [64], психосоциальным стрессом и физической активностью [57], вынужденным плаванием и шумовым стрессом [45], холодовым и иммобилизационным стрессами [65] (табл. 1). В наших работах была показана перекрестная адаптация между гипобарической гипоксией и психоэмоциональными стрессорами в моделях тревожно-депрессивных расстройств. В частности, оказалось, что гипоксическая толерантность организма, индуцируемая гипобарическим гипоксическим прекондиционированием, обладает свойством универсальности, обеспечивая повышение резистентности не только к факторам гипоксической природы, но и психоэмоциональным стрессорам в экспериментальной модели посттравматического стрессового расстройства (ПТСР) и депрессии у крыс [17, 30].
Таблица 1.
Встречаемые по литературным данным виды перекрестной адаптации и сенсибилизации
| Гипоксия | ВП | ИММ | Шум | Холод | ТС | TS | ПТСР/ВБ | FSS | ПСС | |
|---|---|---|---|---|---|---|---|---|---|---|
| Гипоксия | – | + | + | + | + | |||||
| Вынужденное плавание | + | + | ||||||||
| Иммобилизация | + | – | +/– | + | ||||||
| Шум | + | |||||||||
| Физические нагрузки | + | +/– | ||||||||
| Холод | + | +/– | + | |||||||
| Тепловой стресс | + | + | ||||||||
| Хронический комбинированный стресс | +/– |
Как можно заметить, количество подобных сочетаний чрезвычайно разнообразно и отражает саму суть перекрестной адаптации – повышение резистентности к всевозможным гетеротипическим стрессорам. Явление “перекрестной адаптации” было описано в литературе достаточно давно, как у отечественных [25], так и зарубежных авторов [41, 55]. Однако на ранних этапах о глубоком понимании механизмов, лежащих в основе перекрестной адаптации, не было и речи. Наиболее близко к современному видению феномена подошла концепция “следа” Меерсона, где первоначальные изменения отпечатываются как “след” в протеоме и транскриптоме клеток. Более подробно эта концепция и возможные физиологические механизмы будут рассмотрены ниже.
1.1. Экспериментальные исследования перекрестной адаптации на животных
Исследования LeBlanc продемонстрировали наличие перекрестной адаптации между холодовым стрессом и острой гипоксией – что проявляется уменьшенным симпатическим ответом на острую гипоксию после сеанса холодового стресса [61]. Школа аэрокосмической медицины USAF провела исследования положительной кросс-адаптации у крыс – животные группы высотного стресса быстро теряли вес, в то время как крысы с температурной адаптацией показали меньшую потерю массы тела. Кроме того, подобный стрессор не вызывал значительных изменений в 24-часовых показателях экскреции креатинина, мочевой кислоты, магния и натрия в моче у животных с температурной адаптации, что свидетельствует о наличии перекрестной адаптации к данным факторам на физиологическом уровне. Аналогично, в других экспериментах также крысы, адаптированные к высоте и холоду, продемонстрировали кросс-адаптацию и показали меньшую потерю массы тела в ответ на лишение тепла и пищи по сравнению с интактными животными [68].
Тепловая акклиматизация индуцирует кросс-толерантность к факторам, связанным с изменением поступления кислорода (гипоксия, ишемия/реперфузия, гипероксия [43, 62]), ионизирующей радиацией и рядом других стрессирующих ситуаций (шум, травматическое повреждение [73, 78].
Weiss [79] изучал феномен адаптации и кросс-адаптации у крыс, подвергаемых электрошоковому раздражению и плаванию в холодной воде, оценивая попытки избегания перечисленных стрессоров. Подобный холодовой стресс приводил к большему числу попыток избегания электрокожного раздражения, что свидетельствует о наличии перекрестной адаптации уже на поведенческом уровне. Кроме того, поведенческие реакции, свидетельствующие о кросс-толерантности, были также обнаружены в тесте “вынужденного плавания” (тест Порсолта) в холодной воде у крыс, ранее подвергнутых хроническому электрокожному раздражению хвоста [79]. При этом у крыс, подвергшихся острому электрокожному раздражению хвоста, уровни норадреналина в гипоталамусе и коре головного мозга были ниже чем у интактных. Однако у крыс, подвергшихся подобному хроническому воздействию или вынужденному плаванию в холодной воде, не было показано снижения уровней норадреналина при воздействии острого электрокожного раздражения, что демонстрирует адаптацию и перекрестную адаптацию на нейрохимическом уровне. Другие исследования также подтверждают аналогичную нейрохимическую адаптацию в ответ на повторное воздействие стрессора [49, 66, 81]. Подобные изменения уровня норадреналина у особей, подвергавшихся повторным стрессорным воздействиям, объяснялось увеличением активности тирозингидроксилазы (скорость-лимитирующего фермента в цепочке синтеза норадреналина) и снижением поглощения норадреналина в коре больших полушарий. Исходя из этого, было высказано предположение, что за адаптацию, а также кросс-адаптацию в ответ на различные стрессоры в основном отвечает восстановление уровней норадреналина в мозге [79].
Cox с соавторами [53] продемонстрировали положительный профилактический эффект плавания на гипертензию, индуцированную электрокожным раздражением хвоста у погранично-гипертензивных крыс. Эти крысы подвергались неконтролируемым ударам хвоста вместе с ежедневным плавательным стрессом, результаты выявили значительно меньший рост систолического и диастолического кровяного давления у крыс с обоими стрессорами. На нейрохимическом уровне острое снижение уровня норадреналина, индуцированное электрокожным раздражением хвоста у крыс (TS, tail shock), было менее выражено при повторном вынужденном плавании [53]. Armario c коллегами [46] изучали влияние хронического электрокожного раздражения хвоста на физиологические реакции у самцов крыс, ранее подвергшихся воздействию хронического иммобилизационного стресса. В частности, нормализация ответа АКТГ наблюдалась у животных, которые неоднократно подвергались иммобилизации или TS, что указывало на развитие классической адаптации к стрессу. Однако при проведении части исследования с чередованием этих стрессоров, которое должно было выявить наличие кросс-адаптации на уровне АКТГ, было показано, что при воздействии нового острого стрессора (TS) у хронически иммобилизованных крыс, напротив, наблюдалось увеличение ответа АКТГ, свидетельствующее об отсутствии кросс-адаптации на уровне ГГАС. В то же время, несмотря на эндокринные изменения, авторы зафиксировали кросс-адаптацию на уровне других физиологических реакций. Было показано, что хронический TS уменьшает массу тела у нестрессированных до этого животных, тогда как у многократно иммобилизованных крыс, подвергнутых хроническому TS той же интенсивности, наблюдалось увеличение массы тела, что может свидетельствовать о наличии перекрестной адаптации между этими стрессорами. Авторы полагают, что изменения в метаболизме могут быть ответственны за перекрестную адаптацию при подобном стрессе, поскольку ранее иммобилизованные животные оказались метаболически адаптированы для снижения потребления пищи. Их повышенная метаболическая эффективность усвоения пищи, формирующаяся в результате предшествующего иммобилизационного стресса, возможно, может предотвратить потерю веса при хроническом TS. Отсутствие кросс-адаптации на уровне гипофизарного АКТГ и наличие перекрестной адаптации на физиологическом уровне, говорит о том, что перекрестная адаптация может фактически зависеть от типа измеряемой переменной [46].
Pol c коллегами [74] также исследовали эффекты повторяющихся эпизодов иммобилизационного стресса и TS. Отклонения на поведенческом уровне в тесте ОП с отверстиями в полу были довольно значительно выражены у крыс с хронической иммобилизацией. Однако никаких изменений в поведении не наблюдалось в ответ на острое воздействии TS у нестрессированных и предварительно иммобилизированных крыс, что указывало на отсутствие кросс-адаптации на поведенческом уровне в тесте с отверстиями в полу (разновидность теста “открытое поле”). При этом в тесте Порсолта наличие перекрестной адаптации на поведенческом уровне было достоверно выявлено. Это предполагает, что развитие перекрестной адаптации на поведенческом уровне может также зависеть от типа применяемого поведенческого теста. Острые воздействия, как иммобилизация, так и TS, уменьшали уровни норадреналина в различных участках мозга у наивных к стрессу крыс, тогда как подобное снижение в ответ на острый стресс (иммобилизация или шок) не наблюдалось при многократном стрессировании крыс. Стрессовое сохранение уровней норадреналина свидетельствует о существовании нейрохимической перекрестной адаптации [74]. Японские исследователи [69] продемонстрировали феномен перекрестной адаптации между физической активностью и иммобилизационным стрессом у спонтанно гипертензивных крыс (SHR). После 4 недель хронического нагрузочного стресса реакции повышения норадреналина и систолического давления в ответ на острый иммобилизационный стресс значительно снизились. Понижение систолического артериального давления и уровня норадреналина в ответ на новый стрессирующий фактор, очевидно, свидетельствуют о наличии перекрестной адаптации между хроническими упражнениями и иммобилизационным стрессом [69].
Показано, что хроническое воздействие иммобилизационного стресса на 8-й день оказывает протекцию от острого электрошокового раздражения (FSS, foot shock stress) и анксиогенного действия β-карболина (обратного агониста бензодиазепинового сайта связывания ГАМК-рецепторов), что свидетельствует о наличии перекрестной адаптации [50, 52]. Исследование Cancela и коллег также продемонстрировало, что повторные воздействия иммобилизационного стресса улучшали поведение крыс в тесте на принудительное плавание [51]. Однако существуют также менее однозначные примеры. В частности, предварительное прохождение хронического комбинированного стресса не изменяло активную компоненту поведения (“борьбу”) при последующем остром иммобилизационном воздействии, что указывало на развитие только частичной кросс-адаптации [72].
1.2. Перекрестная адаптация у человека
Lunt и коллеги продемонстрировали существование кросс-толерантности между холодовым и гипоксическим воздействиями у людей [64]. Авторы работы продемонстрировали нормализацию гипоксических симптомов, симпатической активности и параметров изменчивости сердечного ритма во время второго гипоксического воздействия после холодового привыкания (погружение участников в воду при 12°C) по сравнению с первым воздействием. Было предположено, что погружение в холодную воду уменьшало симпатическое раздражение (снижение концентрации циркулирующих катехоламинов) и увеличивало парасимпатическую активность при последующих гипоксических воздействиях, вызывая общую вегетативную перекрестную адаптацию. Klaperski c коллегами продемонстрировали частичную перекрестную адаптацию у молодых женщин (18–28 лет), исследуя физиологический и психологический стресс. У активных женщин (6 ч упражнений в неделю) наблюдалось небольшое увеличение частоты сердечных сокращений и уровня кортизола слюны в ответ на острый стресс (социальный стресс-тест Трира) по сравнению с группой редких тренировок (менее 2 ч в неделю) и умеренной физической нагрузки (2–6 ч в неделю), что свидетельствует о наличии перекрестной адаптации на физиологическом уровне. Однако подобная перекрестная адаптация не проявлялась на психологическом уровне, так как активная группа женщин также проявляла беспокойство по поводу воздействия психосоциального стресса [57].
1.3. Отсутствие перекрестной адаптации или перекрестная сенсибилизация
Следует отметить, что последовательное или комбинированное действие неблагоприятных факторов среды на функциональное состояние физиологических систем организма далеко не всегда приводит к формированию перекрестной адаптации. Более того, в ряде случаев действие одних факторов приводит к уязвимости по отношению к другим.
Kant c коллегами исследовали влияние гомотипического или гетеротипического стрессора у крыс, ранее привыкших к повторному гомотипическому стрессу [56]. Крыс неоднократно подвергали воздействию либо иммобилизационного, либо FSS или TS один раз в день в течение 10 дней. На 11-й день крысы подвергались воздействию гомотипического или гетеротипического стресса. Никакой значительной перекрестной адаптации не наблюдались судя по изменениям уровня кортикостерона. Несмотря на то, что повторные воздействия сопровождались изменениями уровней пролактина плазмы крови и гипофизарного циклического АМФ, тем не менее, кросс-адаптации при этом не наблюдалось. Armario с соавторами также продемонстрировали, что адаптация может быть специфичной для стресса. Результаты показали наличие адаптации на гормональном уровне (ЛГ, ГР и кортикостерона) в ответ на неоднократное воздействие шумового стресса. Однако острое предъявление нового стрессора (принудительное плавание) привело к аналогичному увеличению уровней кортикостерона у акклиматизированных крыс, что свидетельствует об отсутствии перекрестной адаптации между этими стрессорами [44].
Кросс-сенсибилизация – это явление, характеризующееся увеличенным стрессорным ответом на новый стрессор после хронического стресса. Существует несколько исследований, подтверждающих потенцирование активности симпатоадреналовой системы у хронически стрессированных крыс в ответ на новое воздействие стрессора [47, 59]. Крысы, акклиматизированные к холоду и гипоксии, демонстрируют значительно более высокий рост плазменных катехоламинов в ответ на иммобилизационный стресс [47]. У животных, подвергнутых повторному стрессовому воздействию, показан повышенный уровень катехоламинов в плазме в ответ на новый стресс по сравнению со стресс-наивными животными [59]. Исследования также продемонстрировали повышение уровней катехоламина в плазме при повторном прохождении холодового/иммобилизационного стресса у крыс в ответ на различные новые стрессоры [54]. Сообщалось о значительном увеличении активности надпочечниковой тирозингидроксилазы у акклиматизированных к холоду крыс после острого воздействия нового стрессора [58, 71]. Rusnak продемонстрировал, что повторное воздействие гомотипического стрессора вызывает адаптацию норадреналиновых нейронов, в то время как острое воздействие гетеротипического нового стрессора сверх-активирует симпатоадреналовую систему, что выражено увеличением экспрессии гена тирозингидроксилазы в locus coeruleus [75]. В статье Marti и коллег сообщается, что острое воздействие гетеротипического стрессора (холодового стресса) приводит к большему увеличению уровней кортикостерона в плазме и АКТГ у крыс с хронической иммобилизацией по сравнению с крысами без предшествующего воздействия хронического стресса. Повышенные уровни норадреналина, адреналина и кортикостерона в плазме после воздействия нового стрессора свидетельствуют о индукции явления кросс-сенсибилизации [65].
1.4. Акклиматизация, тренировка, прекондиционирование и посткондиционирование
Случаи положительной перекрестной адаптации довольно разнообразны и, вероятно, могут лечь в основу адаптационной профилактики многих болезней. Уже сейчас феномен “перекрестной адаптации” применяется в медицине, в том числе военной и спортивной, где используют гипоксические воздействия для повышения неспецифической резистентности организма и физиологических ресурсов организма как с целью профилактики, так и с целью коррекции дезадаптивных состояний различной этиологии. Важным аспектом при реализации адаптотогенного действия факторов, вызывающих перекрестную адаптацию, является режим воздействия. Различают четыре основных – акклиматизация, тренировка, прекондиционирование и посткондиционирование.
Акклиматизация – это адаптация к хронической горной гипоксии при долгом пребывании в горах на высоте. Следует заметить, что понятие “акклиматизация” имеет более широкий смысл, подразумевая адаптацию к любым хроническим климатическим условиям, включая температуру, влажность, инсаляцию и др. Однако поскольку в настоящее время не вызывает сомнений, что ведущая роль в адаптогенном воздействии горной акклиматизации отводится именно гипоксическому фактору, в рамках данного обзора мы будем использовать термин акклиматизация по отношению только к хронической горной гипоксии. Этот феномен хорошо известен издавна, он был использован еще в XVI веке испанскими завоевателями Перу, столица которого в те времена находилась на высоте 3200 м над уровнем моря. Неадаптированные к горам испанские солдаты страдали в условиях высокогорья горной болезнью, поэтому были не способны управлять инками, живущими высоко в горах. Для того, чтобы войско всегда было готово к действиям в любых атмосферных условиях, испанцы акклиматизировали войска путем их проживания в среднегорье по 2 месяца. Научные исследования по проблеме акклиматизации, изначально тесно связанные с изучением физиологических механизмов горной болезни, берут свое начало с работ Лортета [63], второй половины XIX века, который впервые охарактеризовал кривые дыхания на больших и малых высотах. Над этой проблемой плодотворно работал И.М. Сеченов, определив, что пребывание в горах или барокамере приводит к кислородному голоданию тканей организма [34], хотя в основном исследователи того времени придавали ведущее значение другим горно-климатичеким факторам (гипобарии, акапнии, ацидозу, холоду и др.). П. Бэр [48], производя повторные воздействия высоты на организм, впервые описал адаптацию к горной гипоксии, и именно эти исторические исследования заложили основу всех видов современной гипокситерапии. Далее физиологические основы горной акклиматизации изучались достаточно интенсивно, значительный вклад в раскрытие физиологических и биохимических механизмов акклиматизации внесли отечественные ученые А.Г. Гинецинский, Г.Е. Владимиров, З.И. Барбашова, Н.Н. Сиротинин, К.М. Быков, А.Д. Слоним, Н.А. Агаджанян и др. К.М. Быков [8] впервые охарактеризовал физиологическую сущность акклиматизации, заключающуюся в направленных модификациях системных механизмов (эритропоэз, сдвиг кривой диссоциации оксигемоглобина, легочной гипервентиляции, основного и тканевого обмена), приводящих к повышению эффективности механизмов утилизации кислорода в условиях его дефицита. Иными словами, основные приспособительные реакции организма, возникающие при действии хронической горной гипоксии, направлены на ликвидацию или уменьшение кислородной недостаточности. Именно в связи со спецификой этих перестроек, составляющих “структурный след”, акклиматизация приводит к “перекрестной адаптации” к факторам, так или иначе имеющим отношение к гипоксии, если не системной, то тканевой. Это прежде всего повышение устойчивости к физическим нагрузкам, широко применяемое при тренировке спортсменов циклических видов спорта [26]. Однако Ф.З. Меерсоном и коллегами при акклиматизации также были описаны изменения эндокринной и нервной систем, которые могут приводить к снижению при акклиматизации общей реактивности гормональных стресс-систем организма и повышению функциональных возможностей мозга, в частности увеличению степени сохранения условных рефлексов, ускоренному переходу кратковременной памяти в долговременную и увеличению устойчивости мозга к чрезмерным раздражителям, конфликтным ситуациям, эпилептогенам и т.д. [27]. Тем не менее, анализ методических аспектов проведенных коллегами Ф.З. Меерсона экспериментальных исследований по данному вопросу не позволяют однозначно утверждать, что не гипоксическая тренировка, а именно акклиматизация, как адаптация к хронической непрерывной гипоксии, обеспечивает повышение устойчивости не только к гипоксии/ишемии и физическим нагрузкам, но и к острому стрессу. Высказывалось также мнение, что акклиматизация может быть рекомендована для лечения ряда психических расстройств, в том числе депрессии, что связывали с устойчивым усилением кровоснабжения мозга, способствующим поддержанию нормального его функционирования [10], однако убедительных доказательств этой точки зрения пока нет.
Гипоксическая тренировка отличается от режима акклиматизации прерывающейся экспозицией гипоксическому фактору, то есть чередованием периодов гипоксии и реоксигенации. Вероятно, термин “тренировка” использован в данном случае в свете концепции Л.Х. Гаркави и коллег [13] об адаптационных реакциях организма в зависимости от качественно-количественного принципа. Согласно этой концепции, на действие повторяющихся слабых раздражителей (малые дозы) в организме развивается адаптационная реакция, названная “реакцией тренировки” [13].
Наиболее распространенным вариантом гипоксической тренировки является интервальная гипоксическая тренировка (ИГТ), при которой эпизоды умеренной кратковременной гипоксии чередуются с эпизодами нормоксии или, в более редких случаях, гипероксии [20, 32]. Обычно для гипоксических тренировок используют вдыхание газовых смесей с помощью различных гипоксикаторов (например, гипоксикатор А.Я. Чижова, Р.Б. Стрелкова, Т.В. Серебровской, а также их более современные модификации), однако, возможно также использование для этих целей и гипобарической гипоксии.
Теоретически концепция о гипоксических тренировках базировалась на укоренившихся представлениях о целебном воздействии на организм адаптации к горной гипоксии. Возможность полноценной замены горноклиматического и барокамерного гипоксического воздействия на значительно более простой и дешевый способ дыхания газовыми смесями с пониженным содержанием кислорода (базовыми гипоксическими смесями) была доказана в 1970–80 гг. проф. Р.Б. Стрелковым и его коллегами. Было показано, что хорошо переносимая человеком нормобарическая гипоксия развивается в организме человека при дыхании газовыми смесями, содержащими не менее 10% кислорода и 90% азота. ИГТ успешно зарекомендовала себя при подготовке спортсменов, альпинистов и космонавтов, а также гипоксипрофилактики и гипокситерапии различных хронических заболеваний, например, болезней органов дыхания [11, 36]. Обычно применяют 10–24 сеансов суммарной длительностью гипоксического воздействия 300–420 мин, при этом достигается эффект, аналогичный результату месячного пребывания в среднегорье [19, 20], то есть эффективность гипоксической тренировки выше чем акклиматизации. Более того, спектр факторов, к которым формируется перекрестная адаптация при гипоксической тренировке значительно шире и включает гипербарию, радиацию, инфекции, токсические вещества, переохлаждение и перегревание, галлюциногены, алкоголь, аллергены [5, 15, 18, 25, 35]. Кроме того, разными исследователями показан антистрессорный и антидепрессивный эффект гипоксических тренировок как ингаляционных, так и в барокамере [14, 27]. Как уже упоминалось, принципиальным отличием тренировки от акклиматизации является чередование с периодами реоксигенации. Известно, что молекулярно-клеточные процессы, происходящие в период реоксигенации, играют важную роль в формировании ряда патологических и/или адаптивных реакций, более универсальных нежели реакции адаптации к гипоксии. Вероятно, именно в связи с этим наличие периодов реоксигенации повышает эффективность гипоксической тренировки и неспецифичность формирующейся при этом резистентности организма.
Наличием периодов реоксигенации характеризуются и два других режима – прекондиционирование и посткондиционирование, различающихся между собой лишь временем предъявления – до или после повреждающего воздействия, соответственно. Суть данных воздействий заключается в использовании нескольких эпизодов сублетальной гипоксии, интенсивность которой значительно выше, чем при гипоксических тренировках. Соответственно, от режима тренировки пре- и посткондиционирование отличаются более высокой интенсивностью гипоксического воздействия и значительно меньшим числом сеансов. В отличие от акклиматизации и тренировки, в качестве пре- и посткондиционирующих воздействий могут использоваться ишемические воздействия (так называемое ишемические пре- и посткондиционирование, в том числе дистантные). Феномены пре- и посткондиционирования описаны сравнительно недавно, в 1986 и 2003 г., соответственно [70, 80], однако можно сказать, что предпосылки были заложены еще в концепции Л.Х. Гаркави и коллег [13], согласно которой на действие раздражителей средней силы развивается “реакция активации”. Это достаточно точно характеризует сущность феноменов пре- и посткондиционирования, индуцирующих срочную мобилизацию адаптационных резервов организма на разных уровнях организации, от молекулярно-клеточного до системного. По меткому выражению Л.Х. Гаркави, это “более высокий уровень (зона) адаптационных реакций” нежели тренировка.
На современном этапе эффекты и механизмы гипоксического/ишемического кондиционирования, как пре-, так и пост-, довольно хорошо исследованы и продолжают изучаться в экспериментальных моделях на животных. Доказана их высокая эффективность для кардиопротекции, нефропротекции, нейропротекции, повышения стрессоустойчивости, а также реабилитации после повреждающих воздействий, улучшении когнитивных функций, коррекции возрастных деменций и геропротекции [6, 21, 23, 24, 31, 33].
Следует подчеркнуть, что среди всех рассмотренных выше режимов, гипоксическое/ишемическое пре- и посткондиционирование вызывает наиболее выраженную кросс-толерантность к повреждающим факторам психоэмоциональной природы (стрессорам) [3, 4, 30, 76]. Как, показали наши исследования, это связано с одновременной стимуляцией экспрессии факторов нейропластичности в нейронах мозга [9, 16] и сложными перестройками функции ГГАС на всех уровнях ее регуляции [3, 17, 28, 29].
Накопленные к настоящему моменту литературные и собственные данные об особенностях перекрестной резистентности при применении различных видов и режимов гипоксических воздействий представлены в обобщенном виде на рис. 1.
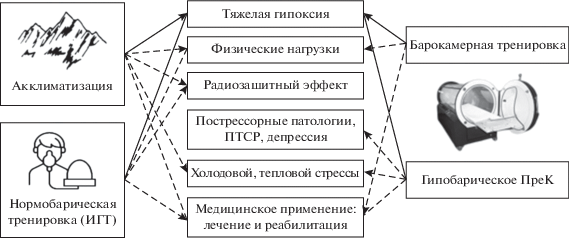
ЗАКЛЮЧЕНИЕ
Таким образом, адаптация – это процесс таких перестроек функциональных систем, а также отдельных органов и тканей, которые обеспечивают возможность длительной активной жизнедеятельности организма животного и человека в измененных условиях существования с сохранением способности к воспроизведению здорового потомства. В этом отношении перекрестная адаптация – это адаптация наиболее высокого уровня, поскольку вызываемые ею перестройки затрагивают базисные адаптивные механизмы и носят неспецифический характер, поэтому обусловленная ими адаптация к конкретному подействовавшему фактору сопровождается одновременным формированием толерантности к другим, потенциальным. Иными словами, понятие перекрестной адаптации непосредственно связано с понятием “устойчивость”, но не равнозначно ему. Адаптация становится перекрестной в том случае, когда наряду с приспособлением к действующему фактору повышается устойчивость к другим факторам.
Несмотря на то, что за прошедшие почти пол века накоплено много сведений о феноменологии и механизмах перекрестной адаптации, остается много нерешенных вопросов. Один из них касается взаимоотношений перекрестной адаптации и перекрестной сенсибилизации, которые, по-видимому, могут существовать одновременно, контролируя баланс между устойчивостью и восприимчивостью к стрессорам высокой биологической важности. Этот вопрос приобретает большую актуальность в связи с развитием нового направления исследований пре- и посткондиционирования – стрессорного. В 1980-е гг. специалисты считали нецелесообразным использовать стрессорные воздействия для повышения резистентности организма, аргументируя свою точку зрения тем, что в этих условиях повышается главным образом пассивная резистентность, причем это вызывает большие и неоправданные энергетические затраты, а также риски повреждения [18]. Однако Л.Х. Гаркави с коллегами [12] показали, что можно повысить и активную резистентность, если повторные стрессорные факторы вызывают развитие “реакции активации” ввиду снижения чувствительности. В результате противоречивости сведений и взглядов того времени внимание к этой проблеме угасло, однако, на современном витке наблюдается новый всплеск интереса к феномену стрессорного кондиционирования. Связанно это с современным прочтением классической концепции Г. Селье о стрессе, подчеркивающим исходно про-адаптивную природу стрессорной реакции [37, 38]. С этих позиций умеренный стресс способен повысить устойчивость к более тяжелому стрессорному воздействию за счет вызываемой им активации классических гормональных механизмов адаптации [39, 40]. В этом отношении, учитывая факт о том, что чрезмерная или длительная активация ГГАС оказывает не про-адаптивное а повреждающее действие, необходимо при стрессорном кондиционировании точно установить те границы, в которых действие умеренного стресса способствует повышению устойчивости, а не сенсибилизации к последующему тяжелому стрессу. Отдельным аспектом данной проблематики является вопрос о том, возможно ли использование в качестве пре- и посткондиционирующего фактора не физического стресса (гипоксия, ишемия, изменение температуры и пр.) по-отдельности или в сочетании с психоэмоциональным, а только умеренного стресса психоэмоциональной природы. На наш взгляд, этот вопрос не может быть решен вне рамок классической теории о феномене перекрестной адаптации, поэтому открывает новые перспективы для ее развития и практического применения.
Работа поддержана грантом РФФИ № 19-015-00336.
Список литературы
Агаджанян Н.А. и др. Экология человека: словарь-справочник. М.: Крук. 1997. 208 с.
Агаджанян Н.А., Двоеносов В.Г. Особенности адаптивных реакций кардиореспираторной системы у лиц с различным уровнем легочной вентиляции при сочетанном воздействии гипоксии и гиперкапнии // Вестник Уральской медицинской академической науки. 2010. № 4. С. 17–21.
Баранова К.А., Пивина С.Г., Рыбникова Е.А. Анксиолитические эффекты умеренной гипоксии и дистантной ишемии в модели посттравматического стрессового расстройства сопровождаются модификацией функций гипофизарно-адренокортикальной системы // Нейрохимия. 2018. Т. 35. № 2. С. 126–131.
Баранова К.А., Зенько М.Ю. Анксиолитический эффект дистантного ишемического пре- и посткондиционирования в модели посттравматического стрессового расстройства // Журн. ВНД им. И.П. Павлова. 2018. Т. 68. № 5. С. 663–672.
Барбашова З.И. Акклиматизация к гипоксии и ее физиологические механизмы // Л.: АН СССР. 1960. С. 12.
Беляков А.В. и др. Патент РФ на изобретение № 2593345. 2016.
Бочаров М.И. Терморегуляция организма при холодовых воздействиях (обзор). Сообщение I // Журн. медико-биологических исследований. 2015. № 2. С. 5–15.
Быков К.М., Мартинсон Э.Э. Материалы по физиологии горного климата //Архивы биологических наук. 1933. Т. 33. № 1–2. С. 72.
Ветровой О.В., Рыбникова Е.А., Самойлов М.О. Церебральные механизмы гипоксического/ишемического посткондиционирования // Биохимия. 2017. Т. 82. № 3. С. 542–551.
Войткевич В.И. Хроническая гипоксия. Приспособительные реакции организма Л. 1973. 189 с.
Волков Н.И. Гипоксическая тренировка для реабилитации и профилактики заболеваний / В сб.: Реабилитация и терапия в условиях курорта. М. 1993. С. 12–25
Гаркави Л.Х., Квакина Е.Б. Диапазоны адаптационных реакций организма // Математическое моделирование биол. процессов. 1979. С. 27–33.
Гаркави Л.Х., Квакина Е.Б., Уколова М.А. Адаптационная реакция организма // Ростов-на-Дону: Изд. РГУ. 1977. 178 с.
Гуревич М.О., Сумская А.М., Хачатуриан А.А. Опыт лечения депрессии аноксемией // Журн. невропатологии и психиатрии им. С.С. Корсакова. 1941. № 9. С. 3.
Жиронкин В.Г. О повышении устойчивости животных к токсическому действию избытка кислорода путем гипоксической тренировки // Кислородная недостаточность. Киев: Изд. АН УССР. 1963. С. 353–357.
Зенько М.Ю., Рыбникова Е.А., Глущенко Т.С. Экспрессия нейротрофина BDNF в гиппокампе и неокортексе у крыс при формировании постстрессового тревожного состояния и его коррекции гипоксическим посткондиционированием // Морфология. 2014. Т. 146. № 5. С. 14–18.
Зенько М.Ю. и др. Патогенетическая роль стрессорного выброса глюкокортикоидных гормонов в генезе посттравматического стрессового расстройства (экспериментальное исследование) // Доклады Академии наук. Наука, 2018. Т. 479. № 3. С. 348–350.
Караш Ю.М., Стрелков Р.Б., Чижов А.Я. Нормобарическая гипоксия в лечении, профилактике и реабилитации // М.: Медицина. 1988. С. 351–352.
Колчинская А.З. Интервальная гипоксическая тренировка. Эффективность, механизмы действия. Киев: Елта. 1992. 159 с.
Колчинская А.З., Цыганова Т.Н., Остапенко Л.А. Нормобарическая интервальная гипоксическая тренировка в медицине и спорте. М.: Медицина. 2003. 408 с.
Лишманов Ю.Б. и др. Стресс- и инфаркт-лимитирующий эффекты раннего гипоксического прекондиционирования // Российский физиологический журн. им. И.М. Сеченова. 2015. Т. 101. № 9. С. 1013–1021.
Лукьянова Л.Д., Кирова Ю.И., Сукоян Г.В. Сигнальные механизмы адаптации к гипоксии и их роль в системной регуляции // Биологические мембраны: Журн. мембранной и клеточной биологии. 2012. Т. 29. № 4. С. 238–238.
Маслов Л.Н. и др. Гипоксическое прекондиционирование как новый подход к профилактике ишемических и реперфузионных повреждений головного мозга и сердца // Ангиология и сосудистая хирургия. 2011. Т. 17. № 3. С. 27–36.
Маслов Л.Н., Лишманов Ю.Б. Ишемическое посткондиционирование сердца. Часть I // Сибирский медицинский журн. (Томск). 2012. Т. 27. № 3.
Меерсон Ф.З. Адаптация, стресс и профилактика. М.: Наука. 1981. 278 с.
Меерсон Ф.З. Общий механизм адаптации и профилактики. М.: Медицина, 1973. 360 с.
Меерсон Ф.З., Кругликов Р.И. Высшие адаптационные реакции организма // Физиология адаптационных процессов (Руководство по физиологии). М.: Наука. 1986. С. 492–518.
Миронова В.И., Рыбникова Е.А. Нейроэндокринные механизмы протективного эффекта гипоксического прекондиционирования в экспериментальных моделях депрессии и тревоги // Обзоры по клинической фармакологии и лекарственной терапии. 2010. Т. 8. № 1.
Рыбникова Е.А. и др. Гормональные механизмы гипоксического прекондиционирования у крыс // Доклады Академии наук. Наука. 2008. Т. 421. № 5. С. 713–715.
Рыбникова Е.А., Воробьев М.Г., Самойлов М.О. Гипоксическое посткондиционирование корректирует нарушения поведения крыс в модели посттравматического стрессового расстройства // Журн. высш. нерв. деят. им. И.П. Павлова. 2012. Т. 62. № 3. С. 364–364.
Рыбникова Е.А., Самойлов М.О. Современные представления о церебральных механизмах гипоксического пре и посткондиционирования // Усп. физиол. наук. 2016. Т. 47. № 4. С. 3–17.
Сазонтова Т.Г. и др. Периодическая гипоксическая и гипероксическая тренировка: тканеспецифичность действия / В сб: Биохимия: от исследования молекулярных механизмов – до внедрения в клиническую практику под ред. Твердохлиба В.П., 2003. С. 64–67.
Самойлов М.О., Рыбникова Е.А. Молекулярно-клеточные и гормональные механизмы индуцированной толерантности мозга к экстремальным факторам среды // Рос. физиол. журн. им. И.М. Сеченова. 2012. Т. 98. № 1. С. 108–126.
Сеченов И.М. К вопросу о дыхании разряженным воздухом // Врач. 1880. № 22. С. 358–359.
Сиротинин Н.Н. Возрастные изменения обмена и реактивность организма. Киев. 1951. С. 62.
Стрелков Р.Б., Чижов А.Я. Прерывистая нормобарическая гипоксия в профилактике, лечении и реабилитации // Екатеринбург: Уральский рабочий. 2001. Т. 258.
Филаретова Л.П. Активация гипоталамо-гипофизарно-адренокортикальной системы как важный гастропротективный компонент стрессорной реакции // Рос. физиол. жур. им. И.М. Сеченова. 2006. Т. 92. № 2. С. 249–262.
Филаретова Л.П. Стресс в физиологических исследованиях // Российский физиологический журн. им. ИМ Сеченова. 2010. Т. 96. № 9. С. 924–935.
Филаретова Л.П. Стресс и язвообразование в желудке: гастропротективная роль глюкокортикоидных гормонов // Рос. физиол. журн. им. И.М. Сеченова. 2009. Т. 95. № 10. С. 1160–1170.
Филаретова Л.П., Багаева Т.Р., Морозова О.Ю. Гастропротективное действие кортикотропин-рилизинг фактора (КРФ): вовлечение глюкокортикоидных гормонов и КРФ рецепторов 2-го типа // Рос. физиол. журн. им. И.М. Сеченова. 2012. V. 98. № 12. P. 1555–1566.
Adolph E.F. Perspectives of adaptation: some general properties // Handbook of physiology. Section. 1964. V. 4. P. 27–35.
Agrawal A., Jaggi A.S., Singh N. Pharmacological investigations on adaptation in rats subjected to cold water immersion stress // Physiology & behavior. 2011. V. 103. № 3–4. P. 321–329.
Arieli Y. et al. Heat acclimation prolongs the time to central nervous system oxygen toxicity in the rat: possible involvement of HSP72 // Brain research. 2003. V. 962. № 1–2. P. 15–20.
Armario A. et al. Response of anterior pituitary hormones to chronic stress. The specificity of adaptation // Neuroscience & Biobehavioral Reviews. 1986. V. 10. № 3. P. 245–250.
Armario A., Castellanos J.M., Balasch J. Effect of chronic noise on corticotropin function and on emotional reactivity in adult rats // Neuroendocrinology letters. 1984. V. 6. № 2. P. 121–127.
Armario A., Hidalgo J., Giralt M. Evidence that the pituitary-adrenal axis does not cross-adapt to stressors: comparison to other physiological variables // Neuroendocrinology. 1988. V. 47. № 3. P. 263–267.
Balaz V. et al. The effect of one-year acclimatization of rats to mountain conditions on plasma catecholamines and dopamine-β-hydroxylase activity // Catecholamines and Stress: Recent Advances, edited by Usdin E., Kvetnansky R., Kopin I.J. New York: Elsevier North Holland. 1980. P. 259–264.
Bert P. La pression barométrique: recherches de physiologie expérimentale. G. Masson, 1878.
Bliss E.L., Zwanziger J. Brain amines and emotional stress // J. psychiatric research. 1966. V. 4. № 3. P. 189–198.
Cancela L.M., Bregonzio C., Molina V.A. Anxiolytic-like effect induced by chronic stress is reversed by naloxone pretreatment // Brain research bulletin. 1995. V. 36. № 3. P. 209–213.
Cancela L.M., Rossi S., Molina V.A. Effect of different restraint schedules on the immobility in the forced swim test: modulation by an opiate mechanism // Brain research bulletin. 1991. V. 26. № 5. P. 671–675.
Chauhan E. et al. Pharmacological investigations on cross adaptation in mice subjected to stress immobilization // Life sciences. 2015. V. 127. P. 98–105.
Cox R.H. et al. Cardiovascular and sympathoadrenal responses to stress in swim-trained rats // J. Applied physiology. 1985. V. 58. № 4. P. 1207–1214.
Dronjak S. et al. Effects of novel stressors on plasma catecholamine levels in rats exposed to long-term cold // Stress neural, endocrine and molecular studies. 2002. C. 83–89.
Hale H.B. Cross-adaptation // Environmental Research. 1969. V. 2. № 5–6. P. 423–434.
Kant G.J. et al. Habituation to repeated stress is stressor specific // Pharm. Biochem. and Behav. 1985. V. 22. № 4. P. 631–634.
Klaperski S. et al. Does the level of physical exercise affect physiological and psychological responses to psychosocial stress in women? // Psychology of Sport and Exercise. 2013. V. 14. № 2. P. 266–274.
Kvetnansky R. et al. 21. Novel Stressors Exaggerate Tyrosine Hydroxylase Gene Expression in the Adrenal Medulla of Rats Exposed to Long-Term Cold Stress // Stress: neural, endocrine and molecular studies. 2002. P. 121.
Kvetnansky R. et al. Plasma catecholamines in rats during adaptation to intermittent exposure to different stressors // Stress: The role of catecholamines and other neurotransmitters. 1984. V. 1. P. 537–562.
Launay J.C. et al. Acclimation to intermittent hypobaric hypoxia modifies responses to cold at sea level // Aviation, space, and environmental medicine. 2006. V. 77. № 12. P. 1230–1235.
LeBlanc J. Stress and interstress adaptation // Federation Proc. 1969. V. 28. P. 996–1000.
Levi E. et al. Heat acclimation improves cardiac mechanics and metabolic performance during ischemia and reperfusion // J. Appl. Physiol. 1993. V. 75. № 2. P. 833–839.
Lortet L. Deux ascensions au Mont Blanc et le mal des montagnes. Recherches sur la circulation et la respiration à une grande altitude. Tracés sphygmographiques pris à 4.810 mètres d’altitude. Paris. 1869.
Lunt H.C. et al. “Cross-adaptation”: habituation to short repeated cold-water immersions affects the response to acute hypoxia in humans // The J. Physiology. 2010. V. 588. № 18. P. 3605–3613.
Marti O. et al. Direct evidence for chronic stress-induced facilitation of the adrenocorticotropin response to a novel acute stressor // Neuroendocrinology. 1994. V. 60. № 1. P. 1–7.
Maynert E.W., Levi O. Stress-induced release of brain norepinephrine and its inhibition by drugs // J. Pharmacology and Experimental Therapeutics. 1964. V. 143. № 1. P. 90–95.
McEwen B.S., Stellar E. Stress and the individual: mechanisms leading to disease //Archives of internal medicine. 1993. V. 153. № 18. P. 2093–2101.
Mefferd JR R.B., Hale H.B. Effects of abrupt temperature changes on excretion characteristics of rats acclimated to cold, neutral or hot environments // American J. Physiology-Legacy Content. 1958. V. 195. № 3. P. 726–734.
Miyoshi M., Watanabe T. Running training attenuates blood pressure and norepinephrine responses to immobilization stress in spontaneously hypertensive rats // The J. Physical Fitness and Sports Medicine. 2013. V. 2. № 3. P. 365–372.
Murry C.E., Jennings R.B., Reimer K.A. Preconditioning with ischemia: a delay of lethal cell injury in ischemic myocardium // Circulation. 1986. V. 74. № 5. P. 1124–1136.
Nisenbaum L. K., Abercrombie E. D. Enhanced tyrosine hydroxylation in hippocampus of chronically stressed rats upon exposure to a novel stressor // J. Neurochemistry. 1992. V. 58. № 1. P. 276–281.
Pastor-Ciurana J. et al. Prior exposure to repeated immobilization or chronic unpredictable stress protects from some negative sequels of an acute immobilization // Behav. brain research. 2014. V. 265. P. 155–162.
Paz Z. et al. Prior heat acclimation confers protection against noise-induced hearing loss // Audiology and Neurotology. 2004. V. 9. № 6. P. 363–369.
Pol O. et al. Behavioral and neurochemical changes in response to acute stressors: influence of previous chronic exposure to immobilization // Pharmacology Biochemistry and Behavior. 1992. V. 42. № 3. P. 407–412.
Rusnák M. et al. Effect of novel stressors on gene expression of tyrosine hydroxylase and monoamine transporters in brainstem noradrenergic neurons of long-term repeatedly immobilized rats // Brain research. 2001. V. 899. № 1–2. P. 20–35.
Rybnikova E. et al. Involvement of the hypothalamic-pituitary-adrenal axis in the antidepressant-like effects of mild hypoxic preconditioning in rats // Psychoneuroendocrinology. 2007. V. 32. № 7. P. 813–823.
Selye H. Stress and the general adaptation syndrome // British medical J. 1950. V. 1. № 4667. P. 1383.
Shohami E., Novikov M., Horowitz M. Long term exposure to heat reduces edema formation after closed head injury in the rat // Brain Edema IX. Springer, Vienna, 1994. P. 443–445.
Weiss J.M. et al. Effects of chronic exposure to stressors on avoidance-escape behavior and on brain norepinephrine // Psychosomatic medicine. 1975. V. 37. № 6. P. 522–534.
Zhao Z.Q. et al. Inhibition of myocardial injury by ischemic postconditioning during reperfusion: comparison with ischemic preconditioning // American J. Physiology-Heart and Circulatory Physiology. 2003. V. 285. № 2. P. H579–H588.
Zigmond M.J., Harvey J.A. Resistance to central norepinephrine depletion and decreased mortality in rats chronically exposed to electric foot shock // J. Neuro-visceral relations. 1970. V. 31. № 4. P. 373-381.
Дополнительные материалы отсутствуют.
Инструменты
Успехи физиологических наук