

Высокомолекулярные соединения (серия А), 2019, T. 61, № 6, стр. 542-552
КОНФОРМАЦИИ МАКРОМОЛЕКУЛ И ВНУТРИМОЛЕКУЛЯРНЫЕ КОНФОРМАЦИОННЫЕ ПЕРЕХОДЫ
Т. М. Бирштейн *
Институт высокомолекулярных соединений Российской академии наук
199004 Санкт-Петербург, Большой пр., 31, Россия
* E-mail: birshtein@imc.macro.ru
Поступила в редакцию 14.06.2019
После доработки 06.07.2019
Принята к публикации 11.07.2019
Аннотация
Представлен обзор теоретических работ, посвященных конформациям макромолекул и выполненных автором (или при участии автора) в период от 50-х годов ХХ века до настоящего времени. Обзор демонстрирует многообразие конформаций макромолекул и показывает, как ход развития полимерной науки определял постановку последовательных задач в теоретическом исследовании конформаций и конформационных переходов в макромолекулах. Рассмотрено развитие молекулярной теории гибкости макромолекул и теории внутримолекулярных конформационных переходов спираль–клубок и глобула–клубок. Обсуждены конформации регулярно разветвленных (звездообразных и гребнеобразных) макромолекул при использовании концепции полимерных щеток и рассмотрен конформационный переход незаряженная глобула–ионизованная звезда в звездообразном pH-чувствительном гомополиэлектролите с амфифильными мономерными звеньями. Описан конформационный переход клубок–цветок в линейной полимерной цепи, внедренной в плоскую полимерную щетку. Обсуждены особенности структуры дендронных щеток и показано, что их использование в качестве матрицы позволяет создать модель термочувствительного молекулярного переключателя.
ВВЕДЕНИЕ
Эта статья, как и весь выпуск журнала “Высокомолекулярные соединения”, посвящена 80-летнему юбилею Александра Борисовича Зезина, члена-корреспондента РАН, крупнейшего специалиста в области высокомолекулярных соединений, особенно в области полиэлектролитов. Очень жаль, что Александр Борисович ушел из жизни, не дожив до этого юбилея.
Статья представляется мне не стандартной. Она является обзором теоретических работ, посвященных конформациям макромолекул и выполненных автором (или при участии автора) в период от 50-х годов ХХ века до настоящего времени. Цель этой ретроспективы – не только продемонстрировать многообразие конформаций макромолекул, но и показать, как ход развития полимерной науки, включающий и разработку теоретических представлений, и методов экспериментальных исследований, и возможности синтеза, определял постановку задач в теоретическом исследовании конформаций и конформационных переходов в макромолекулах.
КЛУБКООБРАЗНЫЕ МАКРОМОЛЕКУЛЫ: ОТ РАЗМЕРОВ К МОЛЕКУЛЯРНОЙ ТЕОРИИ ГИБКОСТИ
Начнем с 50-х годов ХХ века. Это очень ранние годы развития науки о полимерах. Лишь в 1920 г. Г. Штаудингер доказал само существование макромолекул. Одна из главных в науке о полимерах книг, монография P. Flory “Principles of Polymer Chemistry” [1], была опубликована только в 1953 г. (к сожалению, ее перевод на русский язык никогда не издавался). П. Флори в 1974 г. был удостоен Нобелевской премии по химии за “фундаментальные достижения, как теоретические, так и экспериментальные, в физической химии макромолекул”.
В теории конформаций макромолекул рассматривались гауссовы клубки и задача сводилась к установлению зависимости размеров клубкообразной макромолекулы от ее степени полимеризации и от простейших характеристик мономерных звеньев: длины связей, значений валентных углов, степени заторможенности внутреннего вращения. Мне представляется интересным, что в результате в 50–60 годы был пройден путь от простейших формул для размеров макромолекул ${{(--{\text{C}}{{{\text{H}}}_{2}}{\kern 1pt} --{\kern 1pt} {\text{CHR}}{\kern 1pt} --)}_{n}}$ к молекулярной теории гибкости макромолекул.
В 1951 г. М.В. Волькенштейн и О.Б. Птицын в журнале “Доклады АН СССР” опубликовали работу “Размеры полимерной цепи” [2] (по материалам дипломной работы О.Б. Птицына), в которой они, используя матрицы вращения систем координат, получили формулы, связывающие средний квадрат расстояния $h$ между концами цепи с числом $N$ и длиной $l$ звеньев, величиной валентного угла (π – α) и средним косинусом угла $\varphi $ внутреннего вращения около связи $\eta = \left\langle {\cos \varphi } \right\rangle $, характеризующим степень заторможенности внутреннего вращения. При $N \gg 1$ эта формула имела вид
(1)
$\overline {{{h}^{2}}} = N{{l}^{2}}\frac{{1 + \cos \alpha }}{{1 - \cos \alpha }}\frac{{1 + \eta }}{{1 - \eta }}$В литературе ее обычно называют формулой Тейлора [3], хотя, как выяснилось позднее, у нее было множество независимых авторов [4–7].
Формула показывает, что рост заторможенности вращения приводит к увеличению размера макромолекулы.
Следующим вопросом явился вопрос о влиянии асимметрии вращения в макромолекулах типа ${{(--{\text{C}}{{{\text{H}}}_{2}}{\kern 1pt} --{\kern 1pt} {\text{CHR}}{\kern 1pt} --)}_{n}}$, связанной с асимметрией мономерных единиц и характеризуемой средним синусом угла $\varphi $ внутреннего вращения $\varepsilon = \left\langle {\sin \varphi } \right\rangle $. Мы провели соответствующие расчеты и получили [8], что асимметрия мономерных звеньев приводит к уменьшению размеров цепи, т.е. к росту ее гибкости:
(2)
$\overline {{{h}^{2}}} = N{{l}^{2}}\frac{{1 + \cos \alpha }}{{1 - \cos \alpha }}\frac{{1 - {{\eta }^{2}} - {{\varepsilon }^{2}}}}{{{{{(1 - \eta )}}^{2}} + {{\varepsilon }^{2}}}}$Однако уже после публикации выяснилось, что в литературе [9], имеется противоположный вывод, следующий из полученной в этой работе формулы
(3)
$\overline {{{h}^{2}}} = N{{l}^{2}}\frac{{1 + \cos \alpha }}{{1 - \cos \alpha }}\frac{{{{{(1 + \eta )}}^{2}} + {{\varepsilon }^{2}}}}{{1 - {{\eta }^{2}} - {{\varepsilon }^{2}}}}$Данное расхождение послужило стимулом к последующему детальному анализу, который показал, что необходимо учитывать связь направлений преимущественного вращения около соседних связей с асимметрией мономерных единиц и со стереохимией цепи. Что касается формул (2) и (3), то оказалось, что формула (2) описывает изотактические макромолекулы ${{(--{\text{C}}{{{\text{H}}}_{2}}{\kern 1pt} --{\kern 1pt} {\text{CHR}}{\kern 1pt} --)}_{n}}$, для синдиотактических макромолекул остается справедливой формула (1), а формула (3) вообще не справедлива [10].
Но наиболее существенным в этой истории является не этот конкретный результат, а то, что она, подчеркнув связь характера внутреннего вращения со стереохимической структурой макромолекулы, оказалась дополнительным толчком к переходу от простейших формул для размеров клубкообразных макромолекул к разработке молекулярной теории гибкости полимерных цепей.
Основой молекулярной теории гибкости макромолекул послужило введенное М.В. Волькенштейном представление о поворотно-изомерном механизме внутреннего вращения в полимерах [11, 12]. Поворотные изомеры (локальные конформации мономерных единиц) зависят от химической и стереохимической структур. Они рассчитываются методом молекулярной механики с помощью полуэмпирических потенциалов взаимодействия.
Вращения около соседних связей взаимозависимы. Макромолекула – это линейная кооперативная система и теория гибкости строится именно как теория линейных кооперативных систем [13], которая позволяет перейти от локальных конформаций мономерных единиц к характеристикам цепи как целого. Впервые в молекулярной физике физические свойства сложных кооперативных неупорядоченных систем удалось количественно рассчитать исходя из химического строения.
Основы разработанной теории были представлены в монографии [14]. Серию монографий [12, 14] завершила монография P.J. Flory [15], расширившая класс рассмотренных макромолекул. Особенности внутреннего вращения в таких термостойких высокопрочных полимерах, как полиимиды, полиэфиримиды, полиоксифенилены и т.д., проанализированы в работе [16].
В завершение данного раздела отметим, что в настоящее время локальные конформации участков полимерной цепи и конформации цепи как целого удается непосредственно увидеть. Такую возможность дает полноатомное моделирование полимерных систем, реализация которого стала возможной благодаря расширению вычислительных возможностей при использовании суперкомпьютеров.
КЛУБКИ, СПИРАЛИ, ГЛОБУЛЫ И КОНФОРМАЦИОННЫЕ ПЕРЕХОДЫ В МАКРОМОЛЕКУЛАХ
Развитие новой науки – молекулярной биологии (датой рождения считается публикация в 1953 г. работы J. Watson и F. Crick [17] о структуре двойной спирали ДНК), а затем и молекулярной биофизики расширило представление о конформациях макромолекул. К конформациям клубков добавились конформации протяженных жестких спиралей и конформации компактных глобул. Это стимулировало постановку новых задач в статистической физике макромолекул. В первую очередь были рассмотрены задачи о стабильности структур и о внутримолекулярных конформационных переходах при изменении внешних условий.
Начнем со спиральных конформаций (двойная спираль ДНК, спирали синтетических полипептидов) и переходов спираль–клубок. В данном случае речь идет об изменении состояния линейной кооперативной системы, поскольку спираль стабилизована взаимодействиями близких по цепи групп. Статистическая теория линейных кооперативных систем, как уже подчеркивалось, была разработана в работе E. Ising [13]. Теории переходов спираль–клубок в синтетических полипептидах и ДНК, основой которых послужила эта классическая работа, были развиты Зиммом, Бреггом, Лифсоном и другими уже в конце 50-х–начале 60-х годов (см. обзор работ в гл. 9–11 монографии [14]). Стоял вопрос об экспериментальном определении степени стабильности спиральной структуры.
Используя основы теории, мы вместе с экспериментаторами искали рациональный метод обработки кривых перехода спираль–клубок, который дает возможность получить максимально возможную количественную информацию о термодинамических характеристиках спирали. В работе [18] было исследовано плавление ДНК при воздействии двух факторов (температуры и pH) и определена энергия стабилизации двойной спирали ΔH = 10–11 ккал/моль. В последующих работах [19, 20] удалось развить общий метод, позволяющий использовать экспериментальные кривые конформационных переходов между различными состояниями биологических и модельных макромолекул для определения стабильности этих конформационных состояний и кооперативности переходов между ними [21].
Переходы глобула–клубок с теоретической точки зрения существенно отличаются от переходов спираль–клубок. В данном случае речь идет об изменении состояния не одномерной, а трехмерной кооперативной системы. Глобула стабилизована притяжением не только близких по цепи, но и удаленных групп и к тому же обладает “линейной памятью” по терминологии И.М. Лифшица [22]. В работе [22], а также в работах [23–25], выполненных совместно с А.Ю. Гросбергом и А.Р. Хохловым, И.М. Лифшиц разработал строгую статистическую теорию гомополимерной глобулы и фазового перехода клубок–глобула. С решения этой проблемы началось становление Московской школы по физике полимеров.
Наряду со строгой статистической теорией ряд авторов [26–32], начиная с 60-х годов, использовали для построения теории перехода клубок–глобула простую схему метода Флори, развитого им для описания набухшего клубка [1]. К сожалению, как показал проведенный анализ, в указанных работах, по-видимому, по чисто историческим причинам неправильно оценивали конформационную энтропию глобулярного состояния. Следуя Флори, авторы использовали приближение, в котором функция распределения радиуса инерции цепи, определяющего объем цепи в пространстве и, следовательно, ее плотность, считалась подобной функции распределения расстояния между концами полимерной цепи. Это приближение, справедливое для области набухания цепи, не может быть использовано, однако, для области сжатия цепи. Расстояние между концами цепи не является в таких условиях характеристикой ее пространственного размера. Именно функция распределения радиуса инерции, а не расстояния между концами цепи, должна использоваться для расчета энтропии глобулярного состояния.
В работах [33, 34] на основе метода Флори мы построили простейшую теорию перехода клубок–глобула при корректной оценке энтропии глобулярного состояния. Согласно данной теории, переход клубок–глобула в гибкоцепных незаряженных молекулах гомополимеров является фазовым переходом второго рода, при котором не наблюдается скачка плотности.
Жесткость макромолекул обостряет переход, а при значительной жесткости, когда длина сегмента много больше его толщины, меняется и характер перехода, он становится фазовым переходом первого рода.
Упрощенная теория хорошо согласуется со строгой теорией [22–25], давая при этом простую и наглядную интерпретацию результатов [33–35].
ИДЕИ СКЕЙЛИНГА В ФИЗИКЕ ПОЛИМЕРОВ. ПОЛИМЕРНЫЕ ЩЕТКИ
В 1979 г. была опубликована (русский перевод 1982 г.) монография П. де Женна “Идеи скейлинга в физике полимеров” [36]. В 1991 г. П. де Женн был удостоен Нобелевской премии по физике “За открытие, что методы, разработанные для изучения явлений упорядочения в простых системах, могут быть обобщены на более сложные формы материи, в частности, на жидкие кристаллы и полимеры”.
Предлагаемые подходы дали обоснование для разработки скейлинговых теорий полимерных систем, с помощью которых можно получить степенные (асимптотические) зависимости характеристик системы от исходных параметров. Эти подходы ввели в рассмотрение радиус корреляции плотности в полимерных системах и понятие о блобе, не затрагиваемом внешними воздействиями, что позволило учитывать неоднородности в распределении плотности полимерных систем. В целом появилась возможность теоретического изучения полимерных систем сложной архитектуры. Одной из таких систем являлись полимерные щетки – монослои полимерных цепей, связанных одним концом с непроницаемой поверхностью.
Развитие теории полимерных щеток сыграло решающую роль в развитии теории регулярно разветвленных макромолекул.
Полимерные щетки были введены в науку о полимерах в теоретических работах S. Alexander [37] и P.-G. De Gennes [38], которые инициировали их дальнейшие исследования (см., в частности, работу [39]). Особенностью полимерных щеток, которая позволяет рассматривать их как специфические структуры, является перекрывание и, следовательно, взаимодействие привитых макромолекул, в большинстве случаев вызывающее растяжение макромолекул. Так, если щетка погружена в хороший растворитель, то звенья отталкиваются друг от друга (объемные взаимодействия), стремясь уменьшить концентрацию полимера в щетке. Это можно осуществить только растяжением привитых цепей в направлении от поверхности прививки. Толщина щетки определяется компромиссом между тенденцией к уменьшению ее плотности – концентрации полимера в щетке и невыгодностью растяжения привитых макромолекул.
Зависимость толщины щетки от исходных параметров (плотности прививки, размеров и гибкости привитых макромолекул, качества растворителя, характера взаимодействия макромолекул друг с другом и с растворителем и т.п.) строится на основе скейлинговой теории. Но так получается только характеристика щетки как целого. Использование теоретического метода самосогласованного поля дает возможность рассматривать структуру щетки: профиль плотности звеньев в щетке, распределение растяжения привитых макромолекул, т.е. распределение их свободных концов, и т.п. В данном методе изучаются равновесные конформации отдельной привитой макромолекулы, находящейся в самосогласованном поле щетки, т.е. поле, которое создают все макромолекулы, образующие щетку. Развитая аналитическая теория, самосогласованного поля [40–42] позволила разработать теорию структуры плоских полимерных щеток в условиях гауссового растяжения цепей. Один из важных выводов теории – неоднородность плотности щетки. В плоской щетке, погруженной в хороший растворитель, плотность убывает от плоскости прививки к периферии щетки по параболическому закону. Спад плотности обусловлен распределением привитых цепей по степени растяжения.
Метод численного решения уравнений самосогласованного поля (метод SF-SCF) дал дополнительные возможности детального изучения основных закономерностей самоорганизации в полимерных щетках любой геометрии при сложной структуре привитых макромолекул, многообразии их взаимодействий и т.п. Метод был разработан (см. работу [43]) J.M.H.M. Scheutjens (S) и G.J. Fleer (F) и в течение многих лет успешно развивается F.A.M. Leermakers [44]. Ряд представленных ниже результатов получен при использовании этого метода. Метод базируется на простейшей решеточной модели полимерной системы [1] и не требует использования суперкомпьютера.
Полимерные щетки оказались весьма распространенной системой. Модификация поверхностей раздела полимерными щетками путем химической прививки или адсорбции концов цепей на поверхности позволяет управлять барьерными свойствами поверхности, характером взаимодействий декорированных полимерной щеткой коллоидных и наночастиц и т.п. Полимерные щетки часто оказываются элементами структур, являющихся результатом самоорганизации в сополимерных системах, как, например, мицеллярных структур амфифильных диблок-сополимеров.
Разнообразие химических структур привитых макромолекул приводит к разнообразию взаимодействий между ними. На свойства щеток влияют также геометрия поверхности прививки, архитектура привитых макромолекул и т.п.
В обзоре 2000 г. [45] мы уже писали, что и после 20-летнего изучения “полимерные щетки продолжают оставаться неиссякаемым объектом исследования, обнаруживая все новые, часто весьма неожиданные свойства”. Интересно, что, хотя прошло еще почти 20 лет, этот вывод не устаревает. Так, недавно, при полноатомном моделировании была обнаружена необычная, ранее не обсуждавшаяся структура полимерной щетки, оказавшаяся связанной с наличием продольного дипольного момента у привитых цепей [46–48].
РЕГУЛЯРНО РАЗВЕТВЛЕННЫЕ МАКРОМОЛЕКУЛЫ. МИКРОФАЗОВО РАЗДЕЛЕННАЯ КОНФОРМАЦИЯ ПОЛИЭЛЕКТРОЛИТНОЙ ЗВЕЗДЫ
Вернемся к конформациям макромолекул и рассмотрим регулярно разветвленные звездообразные и гребнеобразные макромолекулы (рис. 1). Вопрос о связи их размеров с параметрами неоднократно обсуждался в литературе [49–51], но не получал решения. Именно концепция полимерных щеток сыграла и продолжает играть ключевую роль в разработке теории регулярно разветвленных макромолекул. Такие макромолекулы можно считать полимерными щетками с неплоской поверхностью прививки: звездообразная макромолекула является (при не слишком малом числе лучей) сферической щеткой, гребнеобразная макромолекула – цилиндрической щеткой. В теории полимерных щеток рассматривают деформацию привитых цепей, обусловленную их взаимодействием. В полимерной звезде это соответствующая деформация ветвей звезды. Тем самым, потеря энтропии всей звезды, связанная с внутримолекулярным взаимодействием, оценивается через энтропию деформации ее ветвей. В настоящее время общепринята скейлинговая теория структуры звездообразных полимеров, теория Даоуда–Коттона [52], базирующаяся на указанном подходе. Независимо, но с небольшим опозданием мы получили совпадающие результаты [53], рассмотрев также звезды не только с гибкими, но и с полужесткими ветвями. Была использована построенная нами диаграмма состояния раствора полужестких макромолекул [54]. Полученные степенные зависимости размеров звездообразных гомополимерных макромолекул от числа, длины и гибкости/жесткости ветвей хорошо согласуются с экспериментальными данными и результатами моделирования. В случае звезды в хорошем (или тета) растворителе скейлинговая теория позволила получить также профиль плотности звезды. Как оказалось, плотность убывает при росте расстояния $r$ от центра по степенному закону. Отметим, что, как было показано позднее [55, 56], дополнительный учет не рассматривавшихся ранее нелокальных “центробежных” эффектов (тенденции ветвей звезды сильнее растянуться в центре, чтобы достичь большего доступного объема) практически не влияет на результаты в случае незаряженных звезд.
Рис. 1.
Схематическое изображение звездообразной макромолекулы из 12 ветвей (а), блобная картина звездообразной макромолекулы (б), схематическое изображение гребнеобразной макромолекулы (в).
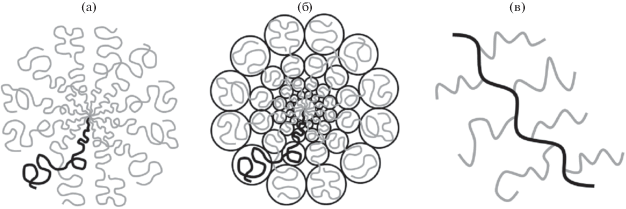
В отличие от звездообразных макромолекул, для которых вопрос об их конформациях был успешно решен около 40 лет назад, для гребнеобразных макромолекул проблема оказалась значительно более сложной. Она остается актуальной вплоть до настоящего времени, тем более, что развитие полимерной химии позволило синтезировать регулярно организованные гребнеобразные макромолекулы с большой линейной плотностью прививки гребней – молекулярные щетки (molecular bottle-brushes), которые стали популярным, практически значимым объектом исследований [57].
Теория конформаций гребнеобразных макромолекул, основанная на модели цилиндрической щетки, была построена в совместной работе представителей Ленинградской и Московской школ по теории полимеров [58]. Была получена зависимость радиуса цилиндрического слоя привитых цепей от параметров прививки. Размер данного слоя определяет толщину гребнеобразной макромолекулы. Эти результаты не потребовали дальнейшего пересмотра. Вместе с тем, важнейшей характеристикой цепной макромолекулы является ее гибкость/жесткость. В рассматриваемой работе был сделан вывод о том, что персистентная длина гребнеобразной цепи оказывается примерно равной ее толщине, т.е. радиусу слоя привитых цепей. Именно такой вывод положил начало длительной дискуссии. С одной стороны, согласно теоретической работе G.H. Fredrickson [59], персистентная длина гребнеобразных цепей должна зависеть от радиуса привитых цепей как этот радиус в степени, близкой к двум, из-за вытягивания части боковых привесков при изгибе и асимметрии сечения цепи в месте изгиба. Это приводит к высокой жесткости цепей. С другой стороны, многочисленные результаты компьютерного моделирования и экспериментальные данные не подтверждают высокую жесткость гребнеобразных макромолекул в растворе, согласуясь с первоначальным выводом [58]. С моей точки зрения вопрос состоит в том, в какой степени симметрия сечения и сохранение толщины цепи в месте изгиба могут быть достигнуты перераспределением боковых цепей. В случае молекулярных щеток в расплаве такое перераспределение обеспечивает их высокую гибкость, регулируемую химическим строением, что определяет возможность создания разнообразных покрытий [60]. В случае молекулярных щеток в растворе возможность симметризации сечения в месте изгиба оказывается зависящей от модели, и вопрос при такой постановке задачи требует дальнейших исследований.
Следующий вопрос в рассматриваемом ряду – конформация звездообразной макромолекулы, являющейся слабым (pH-чувствительным) полиэлектролитом. Макромолекулы полиэлектролитов часто амфифильны на уровне мономерных звеньев. Классическим и детально исследованным примером служит полиметакриловая кислота (ПМАК) ${{[ - {\text{C}}{{{\text{H}}}_{2}}{\kern 1pt} - {\kern 1pt} {\text{C(C}}{{{\text{H}}}_{3}}{\text{)(COOH)}}{\kern 1pt} - ]}_{n}}$, каждая мономерная единица которой содержит ионогенную карбоксильную группу $({\text{COOH}} \to {\text{CO}}{{{\text{O}}}^{ - }}$ + H+) и гидрофобную метильную группу. Гидрофильно-гидрофобный баланс зависит от кислотности водного растворителя и смещается в направлении от гидрофобности к гидрофильности при повышении рH, вызывающем рост степени ионизации карбоксильных групп. В связи с этим при низких pH слабо заряженная амфифильная макромолекула ПМАК локально структурирована за счет гидрофобных взаимодействий метильных групп, а с увеличением рH претерпевает внутримолекулярный конформационный переход в развернутую конформацию (см., например, работу [61]).
Для теоретического исследования конформаций полиэлектролитной звезды с амфифильными мономерными звеньями естественно, как уже говорилось, трактовать ее как сферическую полимерную щетку. Для детального анализа ее структуры при изменении внешних условий был использован метод SF-SCF. Звезда была помещена в большой объем водно-солевого растворителя, значение pH раствора обеспечивало сильную ионизацию кислотных (для определенности) групп, взятых по отдельности.
Оказалось, что при фиксированных значениях pH и параметра гидрофобного взаимодействия размер звезды как целого немонотонно зависит от концентрации низкомолекулярной соли в водно-солевом растворе, в который погружена звезда [62, 63]. Размер звезды минимален в бессолевом растворе, затем растет с увеличением солености и достигает максимума при некоторой умеренной концентрации соли (положение максимума зависит, в частности, от числа и длины ветвей в звезде). При дальнейшем росте солености раствора размер звезды уменьшается. Поведение в этой области характерно для всех полиэлектролитов. С повышением концентрации соли, т.е. ионной силы раствора, экранирующее действие низкомолекулярных противоионов на внутримолекулярное электростатическое отталкивание увеличивается. Степень набухания макромолекулы убывает.
Поведение амфифильной звезды в области низких концентраций соли оказывается необычным. Кривая зависимости ее размера от концентрации соли имеет S-образную форму, свидетельствуя о конформационном переходе. Как следует из детального анализа радиальных профилей плотности звезды и степени ионизации ее звеньев, этот конформационный переход является фазовым переходом первого рода от незаряженной коллапсированной звезды в бессолевом растворе к сильно заряженной и сильно набухшей звезде при концентрации соли вблизи максимума размеров звезды. Основная причина влияния ионной силы раствора на конформацию звезды состоит в электростатических взаимодействиях, благодаря которым заряженная звезда связывает низкомолекулярные противоположно заряженные противоионы. В бессолевом растворе роль противоионов могут играть лишь протоны (для определенности, речь идет о поликислоте). Рост их концентрации в звезде означает уменьшение локального pH, которое оказывается меньше, чем pH в массе растворителя. Низкое локальное pH не допускает ионизации звеньев звезды, и вследствие гидрофобных взаимодействий амфифильных звеньев, звезда приобретает глобулярную конформацию. При добавлении соли в раствор (для определенности, NaCl) часть связанных протонов замещается на Na, так что их концентрация убывает, локальное pH растет, стремясь к значению в растворителе. Возникают условия для ионизации звеньев. Это требует появления новой фазы, в которой ионизованные звенья погружены в раствор.
Интересной особенностью конформации звезды в области перехода глобула–клубок является сосуществование фаз, т.е. наблюдается внутримолекулярная микро- или точнее нанофазовая сегрегация. Звезда приобретает конформацию ядро–корона с незаряженным плотным ядром и заряженной короной. Такая конформация подобна конформации мицеллы из амфифильных диблок-сополимеров, однако в звезде микрофазы не отличаются по составу, отличаясь лишь по плотности и степени ионизации. “Затянутое” сосуществование микрофаз при фазовом переходе первого рода оказалось характерным свойством полимерных щеток. Оно было впервые обнаружено при исследовании численным методом SCF плоских щеток, способных к жидкокристаллическому упорядочению [45, 64]. Причиной эффекта является неоднородность плотности в щетке [65]. В звезде (сферической щетке) радиальный спад плотности сильно выражен, что и приводит гомополимерную звезду к образованию конформации “унимолекулярной мицеллы” (рис. 2).
Рис. 2.
Конформации полиэлектролитной звезды с амфифильными мономерными звеньями. Перестройка конформаций а → б → в → г при росте ионной силы раствора: глобула в бессолевом растворе (а), микрофазно-разделенная конформация (б), развернутая конформация с сильно вытянутыми ветвями (в), развернутая конформация с менее растянутыми ветвями (г).
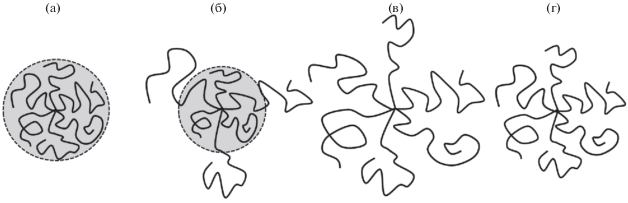
Распределение свободных концов ветвей звезды демонстрирует еще одну интересную особенность исследуемой структуры. Внутримолекулярное фазовое разделении реализуется путем сегрегации ветвей звезды. Они оказываются разделенными на две группы (популяции). Одна образует плотное незаряженное ядро, в котором она полностью размещается. Ветви второй популяции, пройдя через ядро, образуют рыхлую заряженную корону (рис. 2). Такая картина характерна и для плоских полимерных щеток при сосуществовании фаз [64, 65]. Отметим, что сегрегация ветвей была подтверждена результатами эксперимента для аналогичной системы. В работе [66] исследовали водные растворы молекулярных щеток ПИ–графт–ПМАК, где ПИ – водонерастворимый полиимид. В водном окружении главная цепь такой молекулярной щетки коллапсирует и щетка приобретает близкую к звездообразной структуру с амфифильными на уровне мономерных звеньев ветвями ПМАК. Методом поляризованной люминесценции при pH > 6 > pK (ПМАК) обнаружено два времени вращательной релаксации в ПМАК, что свидетельствует о сосуществовании двух различных конформаций ветвей ПМАК. Это соответствует их разделению на популяции в двухфазной структуре, где часть ветвей образовывает заряженную корону, а другая часть располагается в более плотной фазе около ПИ-ядра.
ЦЕПНАЯ МАКРОМОЛЕКУЛА В ЩЕТКЕ. ДЕНДРОННЫЕ ЩЕТКИ. КОНФОРМАЦИОННЫЙ ПЕРЕХОД КЛУБОК–ЦВЕТОК И ТЕРМОЧУВСТВИТЕЛЬНЫЙ МОЛЕКУЛЯРНЫЙ ПЕРЕКЛЮЧАТЕЛЬ
Рассмотрим еще одну конформацию макромолекулы – конформацию цветка и соответствующий внутримолекулярный конформационный переход – переход клубок–цветок. Такой переход может совершать линейная макромолекула, положение одного из концов которой зафиксировано на заданном расстоянии от границы внутри ограниченного объема. Пребывание макромолекулы в этом объеме термодинамически менее выгодно, чем пребывание во внешней среде. Для короткой макромолекулы граница недостижима, и ограниченность объема не влияет на ее конформацию. В частности, в растворе того же полимера эта макромолекула имеет конформацию клубка. Длинная макромолекула (способная достичь границы, которая предполагается проницаемой для нее) приобретает более сложную конформацию цветка. Часть цепи от фиксированного конца растягивается, образуя стебель, выносящий вторую часть со свободным концом во внешнюю среду (растворитель), где она образует клубкообразную головку цветка.
В литературе подробно исследован такой переход в полимерной цепи, внедренной (путем прививки за один из концов) в плоскую полимерную щетку из линейного полимера той же химической структуры [67]. Было показано, что конформация цепи определяется соотношением между степенями полимеризации этой цепи и цепей, образующих щетку. Переход от клубковой конформации, которую имеет короткая привитая макромолекула, к конформации цветка происходит при росте степени полимеризации макромолекулы до значений для цепей в щетке. В области перехода привитая цепь становится подобной цепям щетки и, как и цепи щетки, характеризуется распределением по растянутым конформациям. Переход клубок–цветок оказывается фазовым переходом второго рода. Он происходит при одной и той же степени полимеризации цепи вне зависимости от внешних условий, т.е. переходом нельзя управлять.
Разумеется, существенной является возможность управляемого перехода. В случае цепи в щетке это реализуется только при дополнительном взаимодействии, удерживающим свободный конец длинной привитой цепи внутри щетки [68–70].
Однако появились новые системы. Мы уже говорили о разнообразии полимерных щеток. Последнее время наряду с обычными полимерными щетками, образованными линейными макромолекулами, синтезируют и исследуют полимерные щетки, образованные прививкой макромолекул иной нелинейной архитектуры, в частности, щетки из древовидно-разветвленных макромолекул – дендронов. Начало было положено в 70–80-х годах ХХ века, когда были разработаны методы и начался синтез, изучение структуры, свойств и применение новых макромолекул – дендримеров. Дендримеры – это сферические полимерные щетки из дендронов. Несколько позднее начался синтез и изучение молекулярных (цилиндрических) и интересующих нас плоских дендронных щеток. Одним из стимулов к синтезу дендримеров было кажущееся очевидным представление об их необычной капсулоподобной структуре в растворе. Данное представление, распространяющееся и на плоские и цилиндрические щетки, а также и подтвердившая его теория [71] оказались ошибочными [72, 73]. Влияние архитектуры привитых цепей на структуру щеток не столь радикально. Тем не менее, использование дендронных щеток в качестве матрицы, сделало переход клубок–цветок во внедренной цепи управляемым стимуло-зависимым переходом.
Детальное исследование структуры плоских дендронных щеток в растворе методом SF-SCF было проведено в работах [74–76]. Было показано, что на первый взгляд их структура качественно подобна структуре обычных щеток из линейных цепей. Концентрация звеньев в дендронных, как и в обычных щетках спадает при удалении от плоскости прививки. Более того, при малых плотностях прививки в дендронной щетке, когда все привитые дендроны растянуты слабо, ей можно сопоставить полностью эквивалентную по структуре и свойствам обычную щетку из линейных цепей. Степень полимеризации цепей в эквивалентной щетке оказывается пропорциональной степени полимеризации дендронов, коэффициент пропорциональности меньше единицы и уменьшается с ростом степени разветвленности дендронов. Все закономерности перехода клубок–цветок в линейной цепи, внедренной в дендронную щетку со слабо растянутыми дендронами, полностью совпадают с закономерностями перехода в цепи, внедренной в эквивалентную обычную щетку [77].
Однако это состояние дендронной щетки реализуется, лишь когда ее толщина не превышает длину предельно растянутого корня дендрона.
Особенности дендронной щетки возникают при дальнейшем росте плотности прививки, приводящем к увеличению толщины щетки [74–76]. Они связаны с появлением популяции сильно вытянутых дендронов и качественным изменением структуры щетки, становящейся слоистой. Первый внутренний слой, который был образован при малых плотностях прививки слабо растянутыми дендронами, содержит теперь также предельно растянутые корни “проходных” дендронов. Ветви “проходных” дендронов формируют второй слой. При дальнейшем повышении плотности прививки толщина внутреннего слоя, фиксированная длиной растянутых корней, остается постоянной, толщина внешнего слоя увеличивается за счет роста доли “проходных” дендронов и степени растяжения их ветвей. Когда толщина второго слоя, ограниченная длиной предельно растянутых спейсеров первой генерации, доcтигнет предела, картина повторяется, возникает третья популяция дендронов, а щетка становится трехслойной. Предельное число слоев определяется степенью генерации дендронов g и равно (g + 1). Причиной разделения на популяции является ограниченность растяжения спейсеров. В привитом дендроне растяжение всегда крайне неоднородно. Из-за равновесия сил в точке ветвления корневой спейсер растянут сильнее, чем спейсеры первой генерации, растянутые сильнее, чем спейсеры второй генерации и т.д.
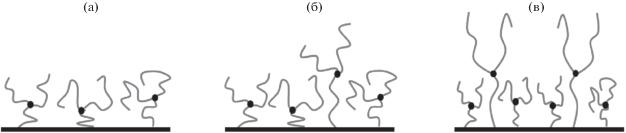
В простейшем случае дендронов первой генерации (звезд, привитых за конец одной из ветвей) дендронная щетка может быть либо однослойной при малой плотности прививки, когда ее толщина меньше контурной длины ветви (корня), либо двуслойной при большей плотности (рис. 3). В двуслойной щетке привитые звезды разделены на две популяции: слабо растянутые, организующие внутренний слой, и проходные с практически предельно растянутым корнем во внутреннем слое и ветвями, образующими второй слой. Отметим, что в целом картина разделения на популяции напоминает картину при фазовом разделении в полиэлектролитной звезде (раздел “Регулярно разветвленные макромолекулы. Микрофазово разделенная конформация полиэлектролитной звезды”). Однако разделение на слои не является фазовым разделением, на границе между слоями нет скачка плотности, различие между слоями не столь значительны. Тем не менее, именно слоистость дендронной щетки приводит к отличию ее свойств от свойств обычной щетки из линейных цепей, увеличивая восприимчивость к внешним условиям. При изменении внешних условий, меняющих толщину щетки, изменяется толщина только внешнего слоя (рис. 3), и соответственно изменяется соотношение слоев и вклад их характеристик в характеристики щетки в целом. Эта особенность структуры дендронной щетки приводит к новым свойствам перехода клубок–цветок в линейной цепи, внедренной в такую матрицу [77].
Рис. 3.
Схематическое изображение структуры щетки из дендронов первого поколения (звезд, привитых за один луч). Изменение структуры а → б → в при росте плотности прививки звезд: однослойная щетка при малой плотности прививки (а) и двуслойная с возрастающей толщиной внешнего слоя при большей (б, в).
Линейная цепь, внедренная в слоистую дендронную щетку, как и цепь, внедренная в “обычную” щетку, претерпевает переход клубок–цветок при росте контурной длины цепи. Однако этим сходство переходов исчерпывается, структура матрицы влияет и на характер и на условия перехода. В отличие от фазового перехода второго рода в случае цепи в “обычной” щетке, для цепи в слоистой дендронной щетке реализуется фазовый переход первого рода, переход по принципу “все или ничего”.
Далее контурная длина внедренной цепи, при которой осуществляется переход, зависит не только от характеристик привитых дендронов, но и является убывающей функцией плотности их прививки. Максимальная длина (наблюдаемая, когда дендронная щетка однослойна) совпадает с длиной цепей в эквивалентной “обычной” щетке. Минимальная длина определяется высотой полностью растянутого дендрона. Только первое из этих значений зависит от суммарной степени полимеризации дендрона, поэтому изменение длины внедренной цепи при переходе оказывается значительным. Так, например, в случае цепи, внедренной в щетку из 8-лучевых звезд (дендронов первой генерации) максимальная длина превышает минимальную (переходы клубок–цветок при малой и большой плотности прививки звезд, соответственно) более чем в 2 раза.
Влияние плотности прививки на положение перехода свидетельствует об обсужденных особенностях поведения слоистой щетки при изменении ее толщины. Вместе с тем зависимость длины внедренной цепи при переходе от плотности прививки показывает, что в слоистой дендронной щетке не только длина цепи оказывается параметром, контролирующим переход. Внедренная цепь фиксированной длины, лежащей в интервале между рассмотренными максимальной и минимальной длиной, всегда претерпевает конформационный переход клубок–цветок при увеличении плотности прививки дендронов. Чем короче внедренная цепь, тем выше плотность прививки дендронов, при которой осуществляется переход.
Этот результат приводит к еще одному очень важному выводу. Толщину дендронной щетки, и, следовательно, ее структуру, можно изменять, меняя качество растворителя, в который она погружена. Значит, переход клубок–цветок может быть индуцирован улучшением качества растворителя, вызывающим, как и рост плотности прививки, увеличение толщины щетки. Длина внедренной цепи должна быть согласована с характеристиками слоистой дендронной щетки (рис. 4).
Рис. 4.
Конформационный переход в линейной цепи, внедренной в слоистую щетку из трехлучевых звезд при переходе от тета- к атермическому растворителю. Показаны функции распределения свободного конца цепи из 240 мономеров, внедренной в щетку из трехлучевых звезд с лучами из 100 мономеров, полученные методом SF-SCF. Плотность прививки звезд (число звезд на единицу площади) равна 0.35.
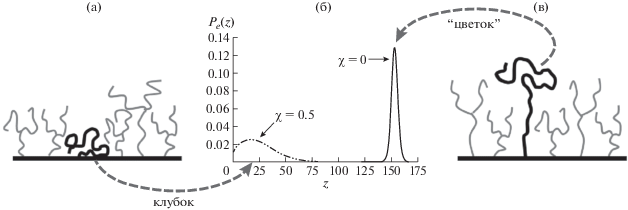
Таким образом, мы получили управляемый внешними условиями, термочувствительный (считаем, что качество растворителя меняется с температурой) конформационный переход клубок–цветок, который реализуется, как фазовый переход первого рода. Привитая макромолекула, претерпевающая переход, может играть роль молекулярного переключателя. Рассмотренная система, содержащая плоскую слоистую дендронную щетку с внедренной линейной макромолекулой, может служить моделью термочувствительного молекулярного переключателя.
Я благодарна всем соавторам цитированных здесь работ с моим участием за сотрудничество.
Список литературы
Flory P.J. Principles of Polymer Chemistry. New York: Cornell Univ.Press, 1953.
Волькенштейн М.В., Птицын О.Б. // Докл. АН СССР. 1951. Т. 78. С. 657.
Taylor W.J. // J. Chem. Phys. 1947. V. 15. P. 412.
Oka S. // Proc. Phys. Math. Soc. Japan. 1942. V. 24. P. 657.
Debye P. Report 637 of Rubber Reserve Company, 1945.
Kuhn Y.H. // J. Chem. Phys. 1947. V. 15. P. 843.
Benoit H. // J. Chim. Phys. 1947. V. 44. P. 18.
Бирштейн Т.М., Птицын О.Б. // Журн. физ. химии. 1952. Т. 26. С. 1215.
Au-Chin Tang, Chin I. // Chem. Soc. 1951. V. 18. P. 103.
Бирштейн Т.М., Птицын О.Б. // Журн. физ. химии. 1954. Т. 28. С. 213.
Волькенштейн М.В. // Докл. АН СССР. 1951. Т. 78. № 5. С. 879.
Волькенштейн М.В. Конфигурационная статистика полимерных цепей. М.; Л.: Изд-во АН СССР, 1959. (Volkenshtein M.V. Configurational statistics at polymeric chains. New York: Interscience. 1963).
Ising E. // Z. Phys. 1925. V. 31. P. 253.
Бирштейн Т.М., Птицын О.Б. Конформации макромолекул. М.: Наука, 1964. (Birshtein T.M., Ptitsyn O.B. Conformations of Macromolecules. New York: Wiley. 1966).
Флори П. Статистическая механика цепных молекул. М.: Мир, 1971. (Flory P.J. Statistical Мechanics of Сhain Мolecules. New York: Interscience. 1969).
Бирштейн Т.М. // Высокомолек. соед. А. 1977. Т. 19. № 1. P. 54.
Watson J., Crick F. // Nature. 1953. V. 171. P. 737.
Сухоруков Б.И., Мошковский Ю.Ш., Бирштейн Т.М., Лысцов В.Н. // Биофизика. 1963. Т. 8. С. 294.
Бирштейн Т.М., Птицын О.Б. // Молек. биология. 1969. Т. 3. № 1. С. 121.
Birshtein T.M., Ptitsyn O.B. // Biopolymers. 1969. V. 7. № 4. P. 435.
Privalov P.L., Ptitsyn O.B., Birshtein T.M. // Biopolymers. 1969. V. 8. № 5. P. 559.
Лифшиц И.М. // Журн. эксперим. и теорет. физики. 1968. Т. 55. № 6. С. 2408.
Лифшиц И.М., Гроссберг А.Ю. // Журн. эксперим. и теорет. физики. 1973. Т. 65. № 6. С. 12.
Lifshitz I.M., Grosberg A.Y., Khokhlov A.R. // Rev. Mod. Phys. 1978. V. 50. № 3. P. 68.
Лифшиц И.М., Гроссберг А.Ю., Хохлов А.Р. // Успехи физ. наук. 1979. Т. 127. № 3. С. 353.
Stockmayer W.H. // Makromol. Chem. 1960. B. 50. S. 54.
Птицын О.Б., Эйзнер Ю.Е. // Биофизика. 1965. Т. 10. № 3. С. 3.
De Gennes P.-G. // J. Phys. Lett. 1975. V. 36. P. 55.
Post C.B., Zimm B.H. // Biopolymers. 1979. V. 18. P. 1487.
Sanchez I.C. // Macromolecules. 1979. V. 12. P. 980.
DiMarzio E.A. // J. Chem. Phys. 1984. V. 81. P. 969.
Muthukumar M. J. // Chem. Phys. 1984. V. 81. P. 6272.
Бирштейн Т.М., Прямицын В.А. // Высокомолек. соед. А. 1987. Т. 29. № 9. С. 2039.
Birshtein T.M., Pryamitsyn V.A. // Macromolecules. 1991. V. 24. P. 1554.
Кузнецов Д.В., Бирштейн Т.М., Гросберг А.Ю. // Высокомолек. соед. Б. 1987. Т. 29. № 12. С. 951.
де Жен П. Идеи скейлинга в физике полимеров. М.: Мир, 1982. (De Gennes P.-G. Scaling Concepts in Polymer Physics. Ithaca, New York: Cornell Univ. Press. 1979).
Alexander S. // J. Phys. 1977. V. 38. P. 983.
De Gennes P.-G. // Macromolecules. 1980. V. 13. P. 1069.
Бирштейн Т.М., Жулина Е.Б. // Высокомолек. соед. A. 1983. Т. 25. № 9. С. 1862. (переиздано в томе избранных работ к 40 летнему юбилею журнала Высокомолекулярные соединения: Polymer Science A. 1999. V. 41. № 12. P. 1406.)
Semenov A.N. // Sov. Phys. JETP. 1985. V. 61. P. 733.
Milner S.T., Witten T.A., Cates M. // Macromolecules. 1988. V. 21. P. 2610.
Жулина Е.Б., Прямицын В.А., Борисов О.В. // Высокомолек. соед. A. 1989. V. 31. № 1. С. 185.
Fleer G.J., Cohen Stuart M.A., Scheutjens J.M.H.M., Cosgrove T., Vincent B. Polymers at Interfaces. London: Chapman and Hall, 1993.
van Male J., Leermakers F.A.M. “sfbox”, a computer program, 2010.
Birshtein T.M., Amoskov V.M. // Polymer Science C. 2000. V. 42. № 2. P. 172.
Glova A.D., Falkovich S.G., Larin S.V., Mezhenskaia D.A., Lukasheva N.V., Nazarychev V.M., Tolmachev D.A., Mercurieva A.A., Kenny J.M., Lyulin S.V. // Polym. Int. 2016. V. 65. № 8. P. 892.
Glova A.D., Larin S.V., Falkovich S.G., Nazarychev V.M., Tolmachev D.A., Lukasheva N.V., Lyulin S.V. // Soft Matter. 2017. V. 13. № 37. P. 6627.
Birshtein T.M., Polotsky A.A., Glova A.D., Amoskov V.M., Mercurieva A.A., Nazarychev V.M., Lyulin S.V. // Polymer. 2018. V. 147. P. 213.
Candau F., Rempp R., Benoit H. // Macromolecules. 1972. V. 5. P. 627.
Khokhlov A.R. // Polymer. 1978. V. 19. P. 1387.
Khokhlov A.R. // Polymer. 1981. V. 22. P. 447.
Daoud M., Cotton J.P. // J. phys. France. 1982. V. 43. № 3. P. 531.
Birshtein T.M., Zhulina E.B. // Polymer. 1984. V. 25. P. 1453.
Бирштейн Т.М. // Высокомолек. соед. А. 1982. Т. 24. № 10. P. 2110.
Zhulina E.B., Birshtein T.M., Borisov O.V. // Eur. Phys. J. E. 2006. V. 20. № 3. P. 243.
Birshtein T.M., Mercurieva A.A., Leermakers F.A.M., Rud’ O.V. // Polymer Science A. 2008. V. 50. № 9. P. 992.
Xie G., Martinez M.R., Olszewski M., Sheiko S.S., Matyjaszewski K. // Biomacromolecules. 2019. V. 20. № 1. P. 27.
Бирштейн Т.М., Борисов О.В., Жулина Е.Б., Хохлов А.Р., Юрасова Т.А. // Высокомолек. соед. A. 1987. Т. 29. №. 6. С. 1169.
Fredrickson G.H. // Macromolecules. 1993. V. 26. P. 2825.
Paturej J., Sheiko S.S., Panyukov S., Rubinstein M. // Science Advavances. 2016. V. 2. № 11. P. 1601478.
Anufrieva E.V., Birshtein T.M., Nekrasova T.N., Ptitsyn O.B., Sheveleva T.V. // J. Polym. Sci. C. 1968. V. 16. P. 3519.
Rud O.V., Mercurieva A.A., Leermakers F.A.M., Birshtein T.M. // Macromolecules. 2012. V. 45. № 4. P. 2145.
Rud O.V., Birshtein T.M. // Polymer Science A. 2013. V. 55. № 12. P. 757.
Amoskov V.M., Birshtein T.M., Pryamitsyn V.A. // Macromolecules. 1996. V. 29. P. 7240.
Klushin L.I., Birshtein T.M., Amoskov V.M. // Macromolecules. 2001. V. 34. № 25. P. 9156.
Pautov V.D., Nekrasova T.N., Anan’yeva T.D., Meleshko T.K., Ilgach D.M., Yakimansky A.V. // Polymer Science A. 2013. V. 55. № 9. P. 526.
Skvortsov A.M., Klushin L.I., Gorbunov A.A. // Macromolecules. 1997. V. 30. № 6. P. 1818.
Skvortsov A.M., Gorbunov A.A., Leermakers F.A.M., Fleer G.J. // Macromolecules. 1999. V. 32. № 6. P. 2004.
Klushin L.I., Skvortsov A.M., Polotsky A.A., Qi S., Schmid F. // Phys. Rev. Lett. 2014. V. 113. № 6. P. 068303.
Qi S., Klushin L.I., Skvortsov A.M., Polotsky A.A., Schmid F. // Macromolecules. 2015. V. 48. № 11. P. 3775.
De Gennes P.-G., Hervet H.J. // Phys. (Paris). 1983. V. 44. P. L351.
Picket G.T. // Macromolecules. 2001. V. 34. P. 8784.
Zook T.C., Picket G.T. // Phys. Rev. Lett. 2003. V. 90. P. 015502.
Polotsky A.A., Gillich T., Borisov O.V., Leermakers F.A.M., Textor M., Birshtein T.M. // Macromolecules. 2010. V. 43. № 22. P. 9555.
Polotsky A.A., Leermakers F.A.M., Zhulina E.B., Birshtein T.M. // Macromolecules. 2012. V. 45. № 17. P. 7260.
Zhulina E.B., Amoskov V.M., Polotsky A.A., Birshtein T.M. // Polymer 2014. V. 55. № 20. P. 5160.
Polotsky A.A., Kazakov A.D., Birshtein T.M. // Polymer. 2017. V. 130. P. 242.
Дополнительные материалы отсутствуют.
Инструменты
Высокомолекулярные соединения (серия А)