

Высокомолекулярные соединения (серия С), 2022, T. 64, № 2, стр. 277-292
МОЛЕКУЛЯРНЫЙ ПЕРЕКЛЮЧАТЕЛЬ НА ОСНОВЕ ЛИНЕЙНОЙ МАКРОМОЛЕКУЛЫ, ВНЕДРЕННОЙ В ЩЕТКУ ИЗ ПРИВИТЫХ ДЕНДРОНОВ ВТОРОЙ ГЕНЕРАЦИИ
А. А. Полоцкий a, *, Т. М. Бирштейн a
a Институт высокомолекулярных соединений Российской академии наук
199004 Санкт-Петербург, Большой пр., 31, Россия
* E-mail: alexey.polotsky@gmail.com
Поступила в редакцию 25.04.2022
После доработки 31.05.2022
Принята к публикации 14.06.2022
- EDN: VRAIDJ
- DOI: 10.31857/S230811472270011X
Аннотация
Методом самосогласованного поля Схойтенса–Флира теоретически исследовано поведение одиночной линейной цепи, внедренной в плоскую полимерную щетку из привитых дендронов второй генерации. Показано, что линейная макромолекула может претерпевать конформационный переход из состояния “клубка”, находящегося внутри щетки, в состояние “цветка”, состоящего из сильно растянутого стебля, погруженного в щетку, и головки, расположенной над щеткой. Этот переход имеет черты фазового перехода первого рода. Он осуществляется при увеличении длины линейной макромолекулы, увеличении плотности прививки, а также при улучшении качества растворителя, в котором находится щетка. Увеличение длины линейной цепи понижает высоту энергетического барьера, разделяющего состояния “клубка” и “цветка”, но сам переход происходит в растворителе худшего качества. В режиме плохого растворителя обнаружено новое состояние линейной цепи – “цветок в щетке”, когда и стебель, и головка цветка находятся внутри щетки.
ВВЕДЕНИЕ
Монослои полимерных цепей, густо привитых к непроницаемой твердой поверхности, или полимерные щетки, служат объектами пристального внимания со стороны теоретиков и экспериментаторов уже более 40 лет [1–5], начиная с пионерских работ S. Alexander [6] и P.-G. de Gennes [7]. Полимерные щетки находят применение в различных приложениях: для стабилизации коллоидных дисперсий [8–10], улучшения барьерных свойств поверхностей, их биосовместимости и предотвращения биозарастания [11–14], улучшения смешиваемости полимера-связующего и нанонаполнителя в нанокомпозитах [15–20], контролируемого высвобождения лекарств [21, 22], улучшения трибологических свойств поверхностей (в том числе для создания искусственных суставов и хрящей) [23–25]. Вместе с тем полимерные щетки являются частью более сложных биологических систем, таких как гликокаликс, нейрофиламенты, мицеллы каппа-казеина и других [26–28].
В последние годы интерес к полимерным щеткам вызван тем, что они выступают основой для создания “умных” (или стимул-чувствительных) поверхностей, свойства которых можно варьировать изменением внешних условий, вызывая конформационные перестройки макромолекул и их внутри- и межмолекулярную самоорганизацию. В роли таких воздействий может выступать изменение температуры, кислотности среды и качества растворителя, электрические и магнитные поля и т.д. Классическим примером “умных”, или стимул-чувствительных, щеток считаются полиэлектролитные щетки, состояние которых можно контролировать изменением ионной силы раствора или pH.
Более сложные сценарии наблюдаются в смешанных щетках, когда к поверхности прививаются макромолекулы различной химической природы. В этом случае образующие щетку полимеры могут быть частично или полностью несовместимыми, или отличаться по степени сродства к растворителю или же к поверхности прививки. Так, смешанные щетки из гидрофобных и гидрофильных цепей способны менять гидрофобность/гидрофильность поверхности в зависимости от того, в каком растворителе располагается щетка [29–32].
Другим свойством, которое можно использовать для управления структурой щетки, является сродство полимера к поверхности, т.е. способность полимерных цепей щетки адсорбироваться на поверхности прививки. В работах [33, 34] рассматривался случай щетки из цепей с адсорбирующейся группой на конце. Увеличение энергии притяжения концевых групп приводит к тому, что часть этих групп оказывается на поверхности, а цепи формируют петли, поэтому щетка превращается фактически в аналог бидисперсной щетки из коротких и длинных цепей, различающихся по длине в 2 раза. В недавней работе [35] исследовалась смешанная полимерная щетка, состоящая из двух типов привитых цепей одинаковой контурной длины, способных адсорбироваться на поверхности прививки и различающихся жесткостью. В отсутствие притяжения к поверхности или при очень слабом притяжении концы более жестких цепей располагаются над гибкими цепями. Однако благодаря тому, что критическая энергия адсорбции для жестких цепей меньше, чем для гибких, постепенное увеличение силы притяжения звеньев к поверхности приводит к преимущественной адсорбции жестких цепей, так что на периферии щетки остаются концы более гибких цепей. Происходит таким образом “переключение” гибких и жестких цепей в смешанной щетке.
Отдельный интерес вызывает предельный случай бинарной смешанной щетки, когда содержание одного из компонентов очень мало по сравнению с другим, и поэтому его можно назвать “минорным” компонентом. Впервые поведение “минорной” цепи, внедренной в щетку и способной адсорбироваться на поверхности прививки, было исследовано в работе А.М. Скворцова с соавторами [36]. Основной компонент щетки – цепи, составляющие щетку, – были инертными к поверхности прививки. Было показано, что если длина “минорной” цепи превышает длину основных цепей, то с увеличением энергии притяжения к поверхности такая цепь может претерпевать резкий конформационный переход из состояния “цветка” с сильно растянутым стеблем, находящимся внутри щетки, и “головой”, располагающейся над щеткой, в адсорбированное состояние (в литературе его также называют “блином” – pancake). Таким образом, адсорбционно-активная цепочка, внедренная в щетку из инертных цепей, может функционировать в качестве молекулярного переключателя. В последующих работах [37, 38] данный переход был исследован более подробно с привлечением взаимодополняющих подходов – методов аналитической теории, численного метода самосогласованного поля и компьютерного моделирования методом Монте–Карло. В работе [39] изучался переключатель на “матрице” из полидисперсной щетки. Наконец, в совсем недавней работе [40] была предложена идея, позволяющая понизить потенциальный барьер и оптимизировать работу переключателя, взяв вместо гомополимера диблок-сополимер с “внешним” нейтральным к поверхности блоком.
Во всех рассмотренных выше примерах компоненты смешанных щеток отличались друг от друга по химической структуре, а с точки зрения архитектуры они все представляли собой линейные макромолекулы. Оказалось, что другой способ получения управляемой смешанной щетки состоит в смешении полимеров разной топологии (архитектуры), но при этом аналогичных химически и имеющих одинаковое сродство к поверхности. В работе [41] были рассмотрены смешанные щетки из привитых за луч звезд и линейных цепей. Было установлено, что конформации звезд и линейных цепей в щетке определяются соотношением их полных молекулярных масс и длины “максимального пути”, соединяющего точку прививки со свободным концом. В частности, когда линейные цепи имеют бóльший “максимальный путь”, но меньшую ММ, чем звезды, конформации звезд и линейных цепей существенно зависят от состава щетки. Если количество линейных цепей невелико, они являются проходными и располагают свои концы на периферии щетки, а при большом содержании линейных цепей – располагаются вблизи поверхности прививки. Переход между двумя режимами характеризуется бимодальным распределением линейных цепей и большими флуктуациями положения их свободных концов. Также переключение линейных цепей и звезд в смешанной щетке может быть вызвано изменением качества растворителя, в котором находится щетка. Впоследствии эти выводы были подтверждены в работе [42], посвященной моделированию аналогичной системы методом молекулярной динамики.
Более подробно был исследован предельный случай смешанной щетки из звезд и линейных цепей – случай “минорной” линейной цепи, внедренной в щетку из привитых звезд [43]. Было показано, что цепочка в щетке может претерпевать переход из состояния привитого “клубка”, расположенного внутри щетки, в состояние “цветка”. Переход “клубок–цветок” может быть вызван изменением внутренних параметров системы (длина цепи или плотность прививки щетки) или внешних условий (качество растворителя). При редкой прививке переход “клубок–цветок” происходит непрерывно в виде фазового перехода второго рода, тогда как при средней и высокой плотности прививки переход имеет черты фазового перехода первого рода. Следовательно, цепочка в щетке может быть использована как молекулярный переключатель, чувствительный к изменению температуры, влияющей на качество растворителя.
Щетки из привитых, регулярно разветвленных дендронов сами по себе выступают интересными объектами, имеющими сложную иерархически организованную внутреннюю слоистую структуру: дендроны в щетке разделяются на несколько групп (популяций), различающихся по степени растяжения [44–50]. Одновременно в щетке присутствуют дендроны, все спейсеры которых растянуты слабо, дендроны, у которых сильно (предельно) растянут корневой спейсер, или спейсер первого слоя, дендроны, у которых сильно растянуты спейсеры первого и второго слоев, дендроны, у которых сильно растянуты спейсеры первого, второго и третьего слоев и т.д. Максимальное число популяций дендронов npop,max и количество субслоев связано с числом генераций в дендроне g и превышает его на единицу: npop,max = g + 1.
Поначалу казалось, что слоистая структура – это всего лишь необычная внутренняя особенность, слабо отражающаяся на макроскопических свойствах щеток, которые во многом были похожи на привычные щетки из линейных привитых макромолекул. Например, слоистая структура никак не отражается на форме профиля плотности полимера в щетке. Изучение поведения внедренной “минорной” линейной цепи в щетке из звезд – дендронов первой генерации – показало, что именно в слоистой щетке внедренная линейная цепь может претерпевать резкий конформационный переход “клубок–цветок” при увеличении плотности прививки или увеличении длины цепи.
Несмотря на важность полученных результатов [43], несколько интересных вопросов оставались неисследованными. В частности, известно, что щетка из привитых дендронов имеет сложную слоистую (многопопуляционную) структуру: повлияет ли увеличение числа слоев на характер перехода “клубок–цветок” или картина останется такой же, как и для дендронов первой генерации? Другой, несомненно, интересный вывод [43] касался возможности индуцировать конформационный переход изменением качества растворителя. Однако в работе [43] была лишь продемонстрирована такая возможность, хотя важные для практических приложений особенности такого перехода – острота перехода, амплитуда перехода, зависимость положения точки перехода от длины цепи и плотности прививки – исследованы не были. Поэтому в настоящей работе изучено поведение линейной цепи, внедренной в щетку из дендронов следующей, второй генерации (g = 2). Также подробно рассмотрен случай атермического растворителя и на основании полученных результатов исследовано поведение внедренной “минорной” линейной цепи при ухудшении качества растворителя.
МОДЕЛЬ И МЕТОДЫ ИССЛЕДОВАНИЯ
В работе исследовали полимерную щетку, образованную дендритно-разветвленными макромолекулами, привитыми к поверхности посредством своей фокальной группы с плотностью (число дендронов на единицу поверхности) σ, и погруженную в растворитель (рис. 1). Дендроны состоят из химически идентичных спейсеров одинаковой длины n = 100, характеризуются числом поколений g и функциональностью точки ветвления q (отношение числа спейсеров, исходящих из точки ветвления, к числу входящих). В данной работе рассматривали щетки из дендронов второй (g = 2) генерации c функциональностью точки ветвления q ≥ 2. Полное число мономерных звеньев в дендроне составляет N = $n({{q}^{{g + 1}}}$ – – 1)/(q – 1). Другая важная характеристика дендронных щеток – длина максимального пути, соединяющего точку прививки дендрона к поверхности с любым из его свободных концов. Для дендрона g-й генерации со спейсерами одинаковой длины число мономерных звеньев в максимальном пути равняется $\mathcal{N} = \left( {g + 1} \right)n$.
Рис. 1.
Изображение “минорной” линейной полимерной цепи, внедренной в плоскую щетку из дендронов второй генерации: а – короткая цепь в конформации “клубка”, б – длинная цепь в конформации “цветка”.
В щетку из дендронов внедряли одиночную линейную макромолекулу, состоящую из M мономерных звеньев. Химическая структура мономерного звена линейной и дендритных макромолекул идентична. Систему погружали в растворитель, качество которого характеризует параметр Флори χ. Качество растворителя изменялось от хорошего (атермического) χ = 0 до умеренно плохого χ = 1.5.
Большая часть результатов представлена для атермического растворителя. В первую очередь нас будет интересовать, какую конформацию примет внедренная в дендронную щетку линейная цепь.
Метод самосогласованного поля
Систему изучали с помощью метода самосогласованного поля Схойтенса–Флира [51]. Этот подход очень активно используется для моделирования неоднородных полимерных слоев уже более 30 лет. Так, в методе самосогласованного поля различные взаимодействия между частицами в системе заменяются средними эффективными взаимодействиями, что позволяет в случае монодисперсной полимерной щетки свести многоцепочечную задачу к одноцепочечной, решать которую гораздо проще. Преимущество данного метода заключается в его вычислительной эффективности с высокой скоростью сходимости в сочетании с воспроизводимостью результатов.
Метод самосогласованного поля основан на приближении среднего поля и дискретном представлении пространства с помощью решетки. Предполагается, что пространство состоит из ячеек решетки размера a и объема a3. В дальнейшем размер ячейки решетки принимается за единицу длины: a = 1. Ячейки решетки организованы в слои, которые расположены параллельно поверхности прививки и имеют номера z = 0, …, zmax. Слой z = 0 соответствует поверхности прививки, а в слое z = 1 закреплены первые сегменты всех привитых цепей. Прививка цепей должна быть достаточно плотной, чтобы цепи сильно перекрывались и образовывали относительно однородную щетку. В этих условиях объемная доля полимера φ и обменный химический потенциал u остаются постоянными в пределах одного и того же слоя и изменяются только в направлении z (поэтому систему или решетку можно назвать одноградиентной). Привитые цепи моделируются как случайные блуждания на простой кубической решетке в эффективном внешнем поле U(z), индуцированном меж- и внутримолекулярными взаимодействиями. Внешнее поле U(z) связано с распределением плотности полимера φ(z) в системе:
(1)
$\frac{{U(z)}}{{{{k}_{{\text{B}}}}T}} = \log \left[ {1 - {{\varphi }}(z)} \right] - 2{{\chi }}\langle {{\varphi }}(z)\rangle $Угловые скобки 〈…〉 обозначают локальное усреднение по ближайшим соседям: $\langle {{\varphi }}(z)\rangle = {{\lambda \varphi }}(z - 1)$ + 4λφ(z) + λφ(z + 1), где λ = 1/6 – вероятность шага на простой кубической решетке в одном из шести возможных направлений. Два соседних звена цепи занимают соседние узлы решетки; корреляции более дальнего порядка игнорируются. Следовательно, перекрывание двух сегментов на узле решетки, в принципе, разрешено, но эффективно предотвращается с помощью условия несжимаемости:
которое должно соблюдаться в каждом слое решетки, φs(z) – объемная доля растворителя в слое z. В рамках метода самосогласованного поля макроскопические свойства полимерной щетки, такие как толщина щетки или профиль объемной доли полимера, могут быть рассчитаны непосредственно на основе минимизации свободной энергии.Для изучения поведения одиночной цепи наиболее рациональным представляется проведение вычислений в два этапа. На первом этапе решается задача для дендронной щетки без внедренной цепи – рассчитываются профили плотности и соответствующий самосогласованный потенциал. Эти вычисления проводили с использованием программы SFBox, разработанной в Университете г. Вагенинген (Нидерланды). На втором этапе решается задача о поведении одиночной цепи заданной длины в полученном самосогласованном потенциале U(z), поскольку предполагается, что внедренная “минорная” линейная цепь не оказывает влияния на щетку в целом, т.е. не возмущает самосогласованный потенциал U(z).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Структура дендронных щеток
Рассмотрим структуру дендронных щеток в условиях атермического растворителя (χ = 0). На рис. 2 представлены профили плотности полимера в щетке из дендронов второй генерации (рис. 2а) и соответствующий ему самосогласованный потенциал $U(z){\text{/}}{{k}_{{\text{B}}}}T = \log \left[ {1 - {{\varphi }}(z)} \right]$ (рис. 2б). Плотность прививки на этих зависимостях варьируется в широком диапазоне, вплоть до близкой к максимальной, которая равна отношению числа мономерных звеньев в длинном пути и в дендронной макромолекуле в целом ${{{{\sigma }}}_{{max}}} = \mathcal{N}{\text{/}}N$ [52]. Видно, что профили плотности не содержат в себе никаких фундаментальных особенностей: плотность полимера непрерывно спадает с увеличением расстояния от плотности прививки, т.е. чем гуще привиты дендроны, тем плотнее и толще становится щетка.
Рис. 2.
Профили объемной доли полимера φ(z) (a), самосогласованный потенциал U(z) = –ln[1 – φ(z)] (б) и преобразованный самосогласованный потенциал (в) в щетке, сформированной дендронами второй генерации со степенью ветвления q = 2, привитыми с плотностью σ = 0.01, 0.02, 0.04, … 0.40 (значения указаны на рисунке).

Если перестроить потенциалы в виде Umax – U(z), где Umax – максимальное значение потенциала, то окажется, что все кривые ложатся на единую универсальную зависимость (рис. 2в). При z = H зависимость потенциала отклоняется от универсальной зависимости. Если строить потенциал как функцию от квадрата расстояния от плотности прививки, то в диапазоне 0 < z < n потенциал U ~ z2. Этот результат находится в полном согласии с аналитической теорией самосогласованного поля дендронных щеток, простроенной в приближении сильного растяжения цепей. При низкой плотности прививки самосогласованный потенциал дендронной щетки, является квадратичным, как и в случае обычной щетки из привитых линейных цепей:
Разница между щетками из дендронов и линейных цепей заключена в зависимости топологического коэффициента ϰ от параметров макромолекул, образующих щетку. В случае щетки из линейных цепей ϰlin представляет собой функцию единственного параметра – степени полимеризации Nlin:
Для щетки из дендронов ϰden – функция всех параметров, описывающих структуру дендрона: ϰden = ϰden (n, g, q). В рассматриваемом в настоящей работе случае дендронов второй генерации g = 2 [53]:
(5)
$\begin{gathered} {{{{\kappa }}}_{{den}}} = {{n}^{{ - 1}}}{\text{arctan}}{{\left[ {q\left( {q + 2} \right)} \right]}^{{ - 1/2}}} = \\ = N_{{den}}^{{ - 1}}(1 + q + \,{{q}^{2}})\arctan {{\left[ {q\left( {q + 2} \right)} \right]}^{{ - 1/2}}}, \\ \end{gathered} $(6)
${{\varphi }}\left( z \right) = 1 - \exp \left[ { - U\left( z \right)} \right]\sim U\left( z \right)$При ϰden = ϰlin профили плотности дендронной и обычной щетки из линейных цепей полностью совпадают, и такие щетки могут рассматриваться как эквивалентные. Как видно из уравнений (4)–(7), щетка из дендронов второй генерации эквивалентна обычной щетке из линейных макромолекул с параметрами

Здесь ${{\eta }} = {{{{\kappa }}}_{{den}}}{\text{/}}{{{{\kappa }}}_{{lin}}} \geqslant 1$ – так называемое топологическое отношение, или степень разветвленности дендрона при сохранении полной молекулярной массы ${{N}_{{den}}} = {{N}_{{lin}}}$:
(9)
${{\eta }} = \frac{2}{{{\pi }}}(1 + q + {{q}^{2}})\arctan {{\left[ {q\left( {q + 2} \right)} \right]}^{{ - 1/2}}}$В табл. 1 представлены теоретические значения Nlin,eq и η для щеток из дендронов второй генерации с q = 2, 3 и 4, рассмотренных в настоящей работе, а также показаны простые оценки значений Nlin,eq и η, полученные на основе скейлингового анализа упрощенной модели дендронной щетки [45]:
(10)
${{N}_{{lin.eq}}}{\text{/}}{{N}_{{den}}} = {{\left( {{{\mathcal{N}}_{{den}}}{\text{/}}{{N}_{{den}}}} \right)}^{{1/2}}},$Таблица 1.
Значения длины линейной цепи в эквивалентной щетке (Nlin.eq/n), топологического отношения η и их скейлинговых оценок (${{\left( {{{\mathcal{N}}_{{den}}}{{N}_{{den}}}} \right)}^{{1/2}}}{\text{/}}n$ и ${{\left( {{{N}_{{den}}}{\text{/}}{{\mathcal{N}}_{{den}}}} \right)}^{{1/2}}}$ соответственно) для щеток из дендронов второй генерации с функциональностью ветвления q и полным числом спейсеров (Nden/n)
| q | Nden/n | Nlin.eq/n | ${{\left( {{{\mathcal{N}}_{{den}}}{{N}_{{den}}}} \right)}^{{1/2}}}{\text{/}}n$ | η | ${{\left( {{{N}_{{den}}}{\text{/}}{{\mathcal{N}}_{{den}}}} \right)}^{{1/2}}}$ |
|---|---|---|---|---|---|
| 2 | 7 | 4.62 | 4.58 | 1.51 | 1.53 |
| 3 | 13 | 6.22 | 6.24 | 2.09 | 2.08 |
| 4 | 21 | 7.81 | 7.93 | 2.69 | 2.65 |
Отметим, что дендронная щетка оказывается эквивалентной обычной щетке из линейных цепей при условии, что самосогласованный потенциал имеет форму (3) по всей толщине дендронной щетки. Из данных, представленных на рис. 2в, следует, что для сохранения параболической формы потенциала должно выполняться условие H < n. Как будет показано ниже, в этих условиях дендронная щетка является однослойной. Когда же это условие нарушается, щетка приобретает двух- или трехслойную структуру.
Обращают на себя внимание небольшие “сингулярности” в окрестности z = 100 = n и z = 200 = = 2n, наблюдаемые на профилях плотности и потенциала при высокой плотности прививки. Это результат сильного растяжения спейсеров дендронов (рис. 3а, 3б) и соответствующего расположения точек первого и второго ветвлений, каждая из которых соединяет три спейсера на этом расстоянии z от плоскости прививки: чем выше плотность прививки, тем более выраженными оказываются эти “сингулярности”.
Рис. 3.
Распределение точек первого (a), второго (б) ветвления и свободных концов (в) дендронов в щетке, образованной дендронами второй генерации со степенью ветвления q = 2, привитыми с плотностью σ = 0.01, 0.02, 0.04, 0.06, … 0.4 (значения указаны на рисунке).

Распределения точек ветвления и концов на рис. 3 показывают, что с увеличением плотности прививки дендроны разделяются на популяции: сначала на две, затем на три. Дендроны первой популяции растянуты слабо, в дендронах второй популяции сильно растянут “корневой” спейсер, за который дендроны привиты к поверхности, или спейсер первого слоя, а в дендронах третьей популяции сильно растянуты спейсеры первого и второго слоя.
Долю дендронов в каждой из популяций можно оценить по площади под распределениями первой и второй точек ветвления. Площадь под пиком распределения первой точки ветвления в окрестности z = n = 100 дает суммарную долю дендронов во второй и третьей популяции (α2 + α3), а площадь под пиком распределения вторых точек ветвления в окрестности z = 2n = 200 дает долю дендронов в третьей популяции (α3). Благодаря этому можно найти α2 и α3, а также вычислить долю слабо растянутых дендронов (α1 = 1 – α2 – α3).
Зависимости доли дендронов в первой, второй и третьей популяциях представлены на рис. 411, они отвечают распределениям точек ветвления на рис. 3а и 3б. Видно, что при редкой прививке все дендроны растянуты слабо, и доля таких дендронов α1 равна единице. Формирование второй популяции происходит скачкоообразно – ее доля α2 в точке образования отлична от нуля. Доля слабо растянутых дендронов α1 скачкообразно уменьшается в этой точке, а с дальнейшим ростом плотности прививки уменьшается непрерывно. Формирование третьей популяции α3 также происходит скачком, причем для этого используются исключительно дендроны второй популяции, а непрерывность уменьшения доли слабо растянутых дендронов α1 в этой точке не нарушается. Формирование третьей популяции изменяет характер поведения доли дендронов во второй популяции: она начинает уменьшаться с ростом плотности прививки. Рост третьей, наиболее растянутой, популяции оказывается настолько сильным, что в эту популяцию переходят как частично растянутые, так и полностью нерастянутые дендроны.
Рис. 4.
а – Зависимости доли дендронов в первой (1), второй (2) и третьей (3) популяциях в щетке, образованной дендронами второй генерации со степенью ветвления q = 2, от плотности прививки σ. б – Схематические изображения щеток с одной (I), двумя (II) и тремя (III) популяциями дендронов.

Можно также сравнить формирование популяционной структуры в щетках из дендронов с разной функциональностью ветвления q (рис. 5). Поскольку увеличение степени ветвления увеличивает полную молекулярную массу дендрона, а “длинный путь” при этом остается неизменным, чтобы сравнение было более корректным, следует отложить зависимости α1, α2, α3 от плотности прививки, приведенной к максимально возможному ее значению. Поскольку ${{{{\sigma }}}_{{max}}} = \mathcal{N}{\text{/}}N$, отношение ${{\sigma /}}{{{{\sigma }}}_{{max}}} = {{\sigma }}N{\text{/}}\mathcal{N}\sim {{\sigma }}N$ представляет собой величину, пропорциональную полной массе полимера, привитого к единице поверхности прививки (полимерной “нагрузке” на поверхность прививки).
Рис. 5.
Зависимости доли дендронов в популяциях: а – нерастянутые; б – с растянутым корневым спейсером; в – с растянутыми спейсерами первого и второго слоя в щетке, образованной дендронами второй генерации со степенью ветвления q = 2, 3 и 4, от плотности прививки σ / σmax.

В целом, доли дендронов в популяциях слабо зависят от функциональности точек ветвления при одинаковой нагрузке. Особенно это характерно для дендронов второй, частично растянутой, популяции. В то время как доля слабо растянутых дендронов α1 монотонно убывает, а доля сильно растянутых дендронов (у которых растянуты спейсеры первого и второго слоя) α3 монотонно растет, доля дендронов во второй “промежуточной” популяции α2 ведет себя немонотонно: рост до начала формирования третьей популяции и убывание “в пользу” третьей популяции.
Таким образом видно, что дендронная щетка обладает нетривиальной “слоистой” внутренней структурой, при этом ее профиль плотности фактически является непрерывным, а не слоистым. Причиной формирования слоистой структуры, как уже неоднократно указывалось в предыдущих работах [45–47, 54], являются плотная прививка дендронов и необходимость упаковать их в щетке, и конечная растяжимость их спейсеров.
Переход “клубок–цветок” в линейной цепи, внедренной в дендронную щетку
Теперь перейдем к ситуации, когда в дендронную щетку внедрена одиночная “минорная” линейная макромолекула. Как было показано для щетки из дендронов первой генерации [43], линейная цепочка в такой щетке может совершать переход между конформациями “клубка” и “цветка” при увеличении плотности прививки или длины цепи.
Прививка дендронов более высокой второй генерации не изменяет эту картину. На рис. 6 приведены зависимости средней высоты конца линейной цепи над плоскостью прививки от длины цепи при фиксированной плотности прививки и от плотности прививки при фиксированной длине цепи, которые показывают, что увеличение длины цепи и плотности прививки вызывает переход “клубок–цветок”.
Рис. 6.
Среднее положение свободного конца линейной цепи, внедренной в щетку, образованную дендронами второй генерации со степенью ветвления q = 2 (а, в) и 3 (б, г), как функции длины линейной цепи фиксированной плотности прививки дендронов (а, б) и как функции плотности прививки при фиксированной длине цепи (в, г). Плотность прививки σ = 0.01, 0.02, 0.04, 0.06 … 0.40 (а) и 0.01, 0.02, 0.04, 0.06 … 0.22 (б); длина цепи M = 330, 340, 350 … 500 (в) и 340, 360, 380 … 600 (г).

С увеличением фиксированной плотности прививки σ переход по M (рис. 6а, 6б) происходит более резко, увеличивается амплитуда перехода, а критическая длина цепи, при которой она начинает выходить из щетки и формировать конформацию цветка, уменьшается. При σ → σmax = $\mathcal{N}{\text{/}}N$ эта критическая длина стремится к длине $\mathcal{N}$ длинного пути в дендроне. Аналогичным образом при фиксированной длине цепи M резкость и амплитуда перехода, вызванного увеличением σ, увеличиваются с уменьшением M (рис. 6в, 6г). Переход “клубок–цветок” осуществляется через сосуществование двух состояний в области перехода.
Диаграмма состояний “минорной” линейной цепи в щетке и различные способы ее представления
Рассмотрим более подробно точку конформационного перехода “клубок–цветок”, которую можно определять по положению максимума флуктуаций конца цепи. Линии точек перехода образуют диаграмму состояний, разделяющую области “клубка” и “цветка”. Эту диаграмму можно представлять в различных координатах. Так, на рис. 7а диаграмма изображена в исходном представлении – в виде зависимости длины цепи, при которой цепь начинает выходить из щетки, формируя конформацию “цветка”, от плотности прививки. При σ → σmax длина выходящей цепи ${{M}_{{tr}}} \to \mathcal{N}$. При σ → 0 длина выходящей цепи стремится к конечному значению: оно тем больше, чем выше функциональность точки ветвления.
Рис. 7.
Диаграммы состояний — линии точек перехода “клубок–цветок” для линейной цепи длины M, внедренной в щетку, образованную дендронами второй генерации со степенью ветвления q = 2, 3 и 4, в четырех представлениях: a – (M, σ), б – (M, σ/σmax), в – (M, φ) и г – (M, H).

На рис. 7б плотность прививки приведена к максимально возможной плотности прививки σmax. Как уже отмечалось выше, отношение σ/σmax пропорционально массе полимера, привитого на единицу площади поверхности прививки, или нагрузке, и мы таким образом можем сравнивать точки перехода для щеток, построенных из дендронов с разной функциональностью точек ветвления q, при одинаковой нагрузке. Видно, что более короткие цепи выходят из щеток из дендронов с более низким q. Значит, при фиксированной нагрузке на поверхность цепочка держится лучше внутри щетки из более разветвленных дендронов.
Можно также сравнить выход цепочек из щеток с одинаковой средней плотностью (рис. 7в). Здесь можно обнаружить, что щетка из более разветвленных дедронов способна удерживать внутри себя более длинные цепи. Средняя плотность полимера в дендронной щетке рассчитывалась из профиля плотности полимера φ(z) как
(11)
${{\varphi }} = \frac{{\sum\limits_z^{} {{{{{\varphi }}}^{2}}\left( z \right)} }}{{\sum\limits_z^{} {{{\varphi }}\left( z \right)} }}$Наконец на рис. 7г длина выходящей цепи представлена в виде зависимости от толщины щетки, которая определяется через среднюю плотность полимера. Действительно, среднюю плотность полимера можно выразить через толщину щетки, число звеньев в дендронах и плотность их прививки как ${{\varphi }} = N{{\sigma /}}H$, следовательно, $H = N{{\sigma /\varphi }}$. Как и на рис. 7б и 7в, при сравнении щеток одинаковой толщины минимальная длина цепи, при которой цепочка может выходить из щетки и формировать конформацию клубка, увеличивается с ростом степени разветвленности дендронов. Это еще раз убеждает нас в том, что более разветвленные дендроны сильнее удерживают цепь в щетке.
Переход “клубок–цветок” при ухудшении качества растворителя
В статье [43] отмечалось, что качество растворителя, в который погружена щетка, влияет на переход “клубок–цветок” во внедренной линейной цепи, и поэтому изменение качества растворителя может быть использовано как стимул для переключения конформаций линейной цепи. В настоящей работе этот эффект изучается более подробно.
На рис. 8 представлены зависимости среднего положения конца внедренной цепи от значения параметра Флори, характеризующего качество растворителя. Поскольку исследуемая система включает одиночную цепь, которая, в свою очередь, способна формировать конформацию “цветка” с “головой”, расположенной на поверхности щетки, и при этом используется одноградиентная версия метода самосогласованного поля, то исследуемый диапазон значений параметра Флори будет в основном ограничен областью не очень высоких значений: 0 ≤ χ ≤ 1.5. В плохом растворителе отдельная цепь коллапсирует в глобулу сферической формы, и такая структура не отвечает плоской геометрии одноградиентной модели. Поэтому мы будем избегать больших значений параметра Флори, соответствующих сильному осадителю. Выбранная область значений параметра Флори вместе с тем полностью соответствует области рабочих значений температуры для ряда водорастворимых полимеров, используемых для создания полимерных щеток. Таковыми являются, в частности, поли-N-изопропилакриламид (ПНИПАМ) и полиэтиленгликоль (ПЭГ) [54]; дендроны на основе ПЭГ применяют для создания дендронных щеток [45, 55, 56].
Рис. 8.
Зависимости среднего положения свободного конца линейной цепи, внедренной в щетку, образованную дендронами второй генерации со степенью ветвления q = 2, привитыми с плотностью σ = 0.1 (а) и 0.2 (б) от параметра Флори. Длина пробной цепи М варьируется от 440 до 480 (a) и от 406 до 450 (б) с интервалом ΔM = 2 на каждом из графиков. Штриховая линия показывает зависимость средней высоты щетки от качества растворителя.
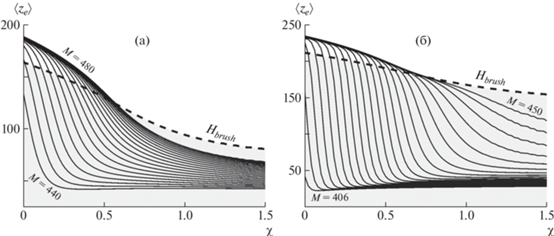
С ухудшением качества растворителя (увеличением χ) щетка как целое коллапсирует, а линейная цепочка может совершать переход из состояния “цветка” в состояние “клубка”. Для этого желательно, чтобы при χ = 0 параметры цепочки и щетки находились на диаграмме состояний (см. рис. 7) вблизи линии перехода. Таким образом, коллапсируя с ухудшением качества растворителя, щетка “втягивает” в себя линейную цепь, которой оказывается предпочтительнее целиком находиться в среде полимерной щетки, близкой по концентрации к полуразбавленному полимерному раствору, чем выставлять часть своих звеньев наружу. Для коротких цепей переход происходит более резко и выраженно, с бóльшей амплитудой. С увеличением длины линейной цепи переход в состояние “клубка” становится более плавным и осуществляется при более высоких значениях χ.
Распределения свободного конца и всех звеньев цепи, т.е. зависимости числа звеньев пробной цепи в плоском слое от номера слоя nc(z), представленные на рис. 9, указывают на то, что переход осуществляется по-разному в зависимости от длины линейной цепи. На этих рисунках распределения представлены совместно с профилем плотности полимера в щетке, поэтому можно хорошо различать состояния “цветка”, в котором его “головка” и концевое звено цепи располагаются над щеткой, и “клубка”, целиком находящегося внутри щетки. Видно, что если цепь короткая (но тем не менее длинная настолько, чтобы находиться в состоянии “цветка” при χ = 0; рис. 9а), то переход осуществляется через сосуществование двух состояний – распределение конца в области перехода, имеющей конечную ширину, бимодально, а максимумы, отвечающие этим двум состояниям, хорошо разделены. Стебель цепи в конформации “цветка” растянут практически равномерно.
Рис. 9.
Распределения свободного конца (верхний ряд) и звеньев (нижний ряд) линейной цепи из М = 420 (а) и 450 (б) мономерных звеньев, внедренной в щетку, образованную дендронами второй генерации со степенью ветвления q = 2, привитыми с плотностью σ = 0.2. Значения параметра Флори указаны на графиках. Также показаны профили плотности полимера в щетке (штриховые линии с закрашенной площадью).

Картина перехода для более длинной цепи оказывается иной. Характерный пример для M = = 450 показан на рис. 9б. Видно, что в условиях хорошего растворителя цепь находится в состоянии “цветка”, максимум распределения ее свободного конца располагается снаружи щетки и лишь незначительно перекрывается с профилем плотности. С ухудшением качества растворителя щетка коллапсирует и втягивает в себя цепь. При этом щетка по-прежнему находится в состоянии “цветка”: на распределении звеньев хорошо различимы “стебель” и “головка”, а максимум распределения конца по-прежнему находится далеко от поверхности прививки, хотя на внешней границе щетки степень перекрывания с профилем плотности с увеличением χ растет. При χ = 1 можно заметить, что оба распределения фактически целиком находятся внутри щетки, а сама граница щетка–растворитель становится резкой. Таким образом, мы сталкиваемся здесь с новым вариантом конформации “цветка”, полностью погруженного в щетку, в отличие от традиционного варианта, когда в щетке расположен только “стебель”, а “головка” находится снаружи щетки в растворителе.
Появление такой конформации связано с тем, что ухудшение качества растворителя оказывает двойной эффект на внедренную цепь. С одной стороны, нахождение в чистом растворителе становится невыгодным для звеньев цепи, поэтому ей предпочтительнее “спрятаться” в щетке (это отвечает второму слагаемому в потенциале, см. выражение (1)). С другой стороны, ухудшение качества растворителя вызывает коллапс и уплотнение щетки в целом, поэтому звеньям цепи становится труднее находить себе место в щетке, полимер щетки действует как выталкивающее поле (это отвечает первому слагаемому в потенциале, уравнение (1)). Поскольку плотность полимера на периферии щетки все же несколько меньше, чем у поверхности прививки, возникает сила, выталкивающая цепь к внешней границе щетки. Она и приводит к формированию конформации “цветка в щетке”, ранее в литературе не обсуждавшейся.
При дальнейшем увеличении параметра Флори происходит еще бóльшее уплотнение щетки и уплощение профиля плотности полимера в щетке. Это делает возможным формирование конформации “клубка в щетке”, располагающегося вблизи поверхности прививки. Так, при χ = 1.5 прослеживается сосуществование конформаций “клубка” и “цветка в щетке”. Очевидно, что при дальнейшем ухудшении качества растворителя вероятность нахождения внедренной цепи в состоянии “цветка в щетке” будет уменьшаться, а в состоянии “клубка в щетке” – увеличиваться.
Формирование конформации “цветка в щетке” является, несомненно, интересным, однако с практической точки зрения, более интересным все же представляется традиционный “цветок с головой”, расположенной над щеткой.
Логарифмируя функцию распределения конца цепи, мы получаем свободную энергию системы как функцию расстояния конца от поверхности прививки, выполняющего роль параметра порядка (свободная энергия представляет собой функцию Ландау): $F\left( z \right) = - \log {{P}_{e}}\left( z \right)$. Максимумы на распределениях ${{P}_{e}}\left( z \right)$ соответствуют минимумам свободной энергии F.
Положение точки перехода можно определять двумя способами – по положению максимума (пика) флуктуаций положения конца или как значение параметра Флори, при котором два минимума свободной энергии имеют одинаковую глубину. На рис. 10 представлена зависимость свободной энергии F(z) при для щеток с q = 2, σ = = 0.2 и цепями разной длины при значениях χ, отвечающих точке перехода. Два минимума свободной энергии разделены барьером, который необходимо преодолеть для осуществления перехода “цветок–клубок” (для удобства сравнения свободная энергия Ландау была построена таким образом, при которой минимум отвечает нулевому уровню свободной энергии). С увеличением длины цепи уменьшается высота барьера, величина минимума, отвечающего состоянию “клубка” увеличивается, а “цветка” – уменьшается.
Рис. 10.
Свободная энергия как функция положения свободного конца линейной цепи, внедренной в щетку, образованную дендронами второй генерации со степенью ветвления q = 2, привитыми с плотностью σ = 0.2 в точке перехода “цветок–клубок”. Длина пробной цепи М варьируется от 410 до 440 с интервалом 10, значение параметра Флори χ отвечает точке перехода M = 410, χ = 0.085 (1); M = 420, χ = 0.32 (2); M = 430, χ = 0.564 (3) и M = 440, χ = 1.08 (4).

На рис. 11 представлена зависимость положения точки перехода от длины внедренной цепи, определенной двумя способами. Для коротких цепей и более высокой плотности прививки, когда переход идет резко и ярко выражено, результаты, полученные двумя способами, фактически совпадают, тогда как для более длинных цепей и редкой прививке наблюдаются различия. Пик флуктуации положения конца примерно соответствует равенству площадей под пиками ${{P}_{e}}\left( z \right)$, отвечающим двум состояниям цепи, что не всегда эквивалентно равенству свободной энергии. Максимум, отвечающий состоянию “клубка”, всегда чуть шире, чем максимум, относящийся к “цветку”, поэтому пик флуктуаций отвечает чуть меньшему значению χ, чем при равенстве глубины минимумов свободной энергии.
Рис. 11.
Зависимость положения точки перехода “цветок–клубок” от длины линейной цепи, внедренной в щетку, образованную дендронами второй генерации со степенью ветвления q = 2, привитыми с плотностью σ = 0.1 (1), 0.2 (2) и 0.3 (3). Положение точки перехода определено по максимуму флуктуаций положения конца (штриховые линии) и равенству значений свободной энергии Ландау в точке минимума (точки).

Зависимости высоты барьера в точке перехода от длины линейной цепи и качества растворителя показаны на рис. 12. Видно, что высота барьера уменьшается с ростом длины цепи и увеличивается с ростом плотности прививки, при этом зависимости высоты барьера от M при разных σ ложатся на универсальную кривую. Уменьшение высоты барьера с длиной цепи связано в том числе с тем, что переход более длинных цепей происходит при более высоких значениях χ, что, несомненно, способствует втягиванию цепи в щетку.
Рис. 12.
Зависимости высоты барьера в точке перехода от длины линейной цепи (a), внедренной в щетку, образованную дендронами второй генерации со степенью ветвления q = 2, привитыми с плотностью σ = 0.2, и от параметра Флори в точке перехода, по равенству значений свободной энергии Ландау в точке минимума.

Из зависимости высоты барьера от качества растворителя в точке перехода следует, что при заданном фиксированном значении χ более низкие значения высоты барьера получаются в редко привитых щетках, при этом длина выходящей цепочки оказывается больше, чем в густо привитых щетках.
Таким образом, пользуясь зависимостями, представленными на рис. 11 и 12, можно построить молекулярный переключатель на основе одиночной линейной цепи, внедренной в дендронную щетку, “настроив” его на переход при определенном значении параметра Флори χ. Высоту барьера и резкость перехода можно регулировать с помощью плотности прививки дендронов к поверхности.
Нетривиальная обратная зависимость высоты барьера от длины цепи наблюдалась нами и в прошлой работе, выполненной для щетки из звезд (дендронов первой генерации) в атермическом растворителе (см. рис. 7). В работе [40] идея удлинения цепи была предложена для уменьшения высоты барьера в переключателе на основе адсорбционно-активной “минорной” цепи в щетке из линейных макромолекул. При этом “минорная” (пробная) цепь удлинялась путем присоединения к ее свободному концу блока из нейтральных, не адсорбирующихся на поверхности, звеньев.
ЗАКЛЮЧЕНИЕ
Настоящая работа посвящена теоретическому изучению конформационного поведения одиночной линейной макромолекулы, внедренной в полимерную щетку из аналогичных по химической структуре привитых дендронов второй генерации. Показано, что линейная макромолекула может претерпевать конформационный переход из состояния “клубка”, находящегося внутри щетки, в состояние “цветка”, состоящего из сильно растянутого стебля, погруженного в щетку, и головки, расположенной над щеткой. Этот переход полностью аналогичен переходу, наблюдаемому в щетке из дендронов первой генерации (привитых звезд) и исследованному ранее [43]. Переход “клубок–цветок” имеет черты фазового перехода первого рода: в области перехода состояния “клубка” и “цветка” сосуществуют одновременно с некоторой вероятностью. Переход из состояния “клубка” в состояние “цветка” может быть индуцирован увеличением длины линейной макромолекулы или увеличением плотности прививки. Увеличение степени разветвленности дендронов способствует удержанию цепи внутри щетки.
При исследовании перехода “цветок–клубок”, вызываемого ухудшением качества растворителя, в который погружена щетка, обнаружено новое состояние линейной цепи – “цветок в щетке”, когда не только стебель, но и головка находятся внутри щетки. Увеличение длины линейной цепи понижает высоту энергетического барьера, разделяющего состояния “клубка” и “цветка”, тем самым облегчая переход, но в то же время повышает значение параметра Флори, при котором осуществляется переход, делая щетку более плотной, что, напротив, способно замедлить переход. Взаимосвязь этих двух противоположно направленных факторов следует учитывать при создании молекулярного переключателя на основе линейной цепи, внедренной в дендронную щетку.
С практической точки зрения переход “клубок–цветок”, индуцируемый изменением температуры, безусловно, должен представлять интерес. Экспериментально такой переход можно было бы исследовать, пометив свободный конец линейной цепи флуоресцентной меткой, которая будет иметь разную интенсивность флуоресценции внутри щетки (когда цепь находится в состоянии “клубка”) и на ее поверхности (когда цепь находится в состоянии “цветка”). Такой подход уже был успешно применен для визуализации конформационных переходов в полимерных щетках [57, 58].
Авторы выражают благодарность Ф.А.М. Леермакерсу (Университет Вагенингена, Нидерланды) за возможность использования программы SFBox для проведения численных расчетов методом самосогласованного поля.
Список литературы
Milner S.T. // Science. 1991. V. 251. P. 905.
Halperin A., Tirrell M., Lodge T.P. // Adv. Polym. Sci. 1992. V. 100. № 1. P. 31.
Birshtein T.M., Amoskov V.M. // Polymer Science C. 2000. V. 42. № 1. P. 172.
Mittal V. // Polymer Brushes: Substrates Technologies, and Properties. Boca Raton: CRC Press, 2012.
Chen W.-L., Cordero R., Tran H., Ober C.K. // Macromolecules. 2017. V. 50. № 11. P. 4089.
Alexander S. // J. Phys. (Paris). 1977. V. 38. P. 983.
de Gennes P.-G. // Macromolecules. 1980. V. 13. P. 1069.
Witten T.A., Pincus P.A. // Macromolecules. 1986. V. 19. № 10. P. 2509.
Zhulina E.B., Pryamitsyn V.A., Borisov O.V. // J. Coll. Int. Sci. 1990. V. 137. P. 495.
Pincus P. // Macromolecules. 1991. V. 24. № 10. P. 2912.
Ye Q., Zhou F. // Antifouling Surfaces and Materials / Ed. by F. Zhou. Berlin; Heidelberg: Springer, 2015.
Higaki Y., Kobayashi M., Murakami D., Takahara A. // Polym. J. 2016. V. 48. P. 325.
Fu Y., Yang Y., Xiao S., Zhang L., Huang L., Chen F., Fan P., Zhong M., Tan J., Yang J. // Progr. Organic Coatings. 2019. V. 130. P. 75.
Liu Y., Zhang D., Ren B., Gong X., Xu L., Feng Z.-Q., Chang Y., He Y., Zheng J. // J. Mater. Chem. B. 2020. V. 8. P. 3814.
Balazs A.C., Singh C., Zhulina E., Lyatskaya Y. // Acc. Chem. Res. 1999. V. 32. P. 651.
Ginzburg V.V., Singh C., Balazs A.C. // Macromolecules. 2000. V. 33. P. 1089.
Kumar S.K., Jouault N., Benicewicz B., Neely T. // Macromolecules. 2013. V. 46. № 9. P. 3199.
Glova A.D., Falkovich S.G., Larin S.V., Mezhenskaia D.A., Lukasheva N.V., Nazarychev V.M., Tolmachev D.A., Mercurieva A.A., Kenny J.M., Lyulin S.V. // Polym. Int. 2016. V. 65. № 8. P. 892.
Nie G., Li G., Wang L., Zhang X. // Polym. Chem. 2016. V. 7. P. 753.
Giovino M., Pribyl J., Benicewicz B., Bucinell R., Schadler L. // Nanocomposites. 2018. V. 4. № 4. P. 244.
Li D., Xu L., Wang J., Gautrot J.E. // Adv. Healthcare Mater. 2020. V. 10. № 5. P. 2000953.
Abdo G.G., Zagho M.M., Khalil A. // Emergent Mater. 2020. V. 3. P. 407.
Kyomoto M., Moro T., Takatori Y., Kawaguchi H., Ishihara K. // Clin Orthop. Relat. Res. 2011. V. 469. № 8. P. 2327.
Kreer T. // Soft Matter. 2016. V. 12. № 15. P. 3479.
Yang W., Zhou F. // Biosurf. Biotribol. 2017. V. 3. № 3. P. 97.
Reitsma S., Slaaf D.W., Vink H., van Zandvoort M.A.M.J., oude Egbrink M.G.A. // Eur. J. Physiol. 2007. V. 454. № 3. P. 345.
Zhulina E.B., Leermakers F.A.M. // Soft Matter. 2009. V. 5. № 15. P. 2836.
de Kruif G.G., Zhulina, E.B. // Coll. Surf. A. 1996. V. 117. № 1−2. P. 151.
Merlitz H., He G.-L., Sommer J.-U., Wu C.-X. // Macromolecules. 2009. V. 42. № 1. P. 445.
He G.-L., Merlitz H., Sommer J.-U., Wu C.-X. // Macromolecules. 2009. V. 42. № 18. P. 7194.
Romeis D., Sommer J.-U. // ACS Appl. Mater. Int. 2015. V. 7. № 23. P. 12496.
Bos I., Merlitz H., Rosenthal A., Uhlmann P., Sommer J.-U. // Soft Matter. 2018. V. 14. № 35. P. 7237.
Skvortsov A.M., Pavlushkov I.V., Gorbunov A.A., Zhulina E.B. // J. Chem. Phys. 1996. V. 105. № 5. P. 2119.
Koch M., Romeis D., Sommer J.-U. // Macromolecules. 2020. V. 53. № 17. P. 7356.
Yang X., Huang J.-H., Mikhailov I.V., Polotsky A.A., Luo M.-B. // Macromolecules. 2020. V. 53. № 17. P. 7369.
Skvortsov A.M., Gorbunov A.A., Leermakers F.A.M., Fleer G.J. // Macromolecules. 1999. V. 32. № 6. P. 2004.
Klushin L.I., Skvortsov A.M., Polotsky A.A., Qi S., Schmid F. // Phys. Rev. Lett. 2014. V. 113. № 6. P. 068303.
Qi S., Klushin L.I., Skvortsov A.M., Polotsky A.A., Schmid F. // Macromolecules. 2015. V. 48. № 11. P. 3775.
Qi S., Klushin L.I., Skvortsov A.M., Liu M., Zhou J., Schmid F. // Adv. Funct. Mater. 2018. V. 28. № 49. P. 1800745.
Qi S., Klushin L.I., Skvortsov A.M., Schmid F. // Macromolecules. 2020. V. 53. № 13. P. 5326.
Polotsky A.A., Leermakers F.A.M., Birshtein T.M. // Macromolecules. 2015. V. 48. № 7. P. 2263.
Su C.-F., Merlitz H., Wu C.-X., Sommer J.-U. // J. Chem. Phys. 2016. V. 145. № 23. P. 234905.
Polotsky A.A., Kazakov A.D., Birshtein T.M. // Polymer. 2017. V. 130. P. 242.
Pickett G.T. // Macromolecules. 2001. V. 34. № 25. P. 8784.
Polotsky A.A., Gillich T., Borisov O.V., Leermakers F.A.M., Textor M., Birshtein T.M. // Macromolecules. 2010. V. 43. № 22. P. 9555.
Polotsky A.A., Leermakers F.A.M., Zhulina E.B., Birshtein T.M. // Macromolecules. 2012. V. 45. № 17. P. 7260.
Merlitz H., Wu C.-X., Sommer J.-U. // Macromolecules. 2011. V. 44. № 17. P. 7043.
Borisov O.V., Polotsky A.A., Rud O.V., Zhulina E.B., Leermakers F.A.M., Birshtein T.M. // Soft Matter. 2014. V. 10. № 13. P. 2093.
Cui W., Su C.-F., Merlitz H., Wu C.-X., Sommer J.-U. // Macromolecules. 2014. P. 47. № 11. P. 3645.
Li C.-W., Merlitz H., Wu C.-X., Sommer J.-U. // Polymer. 2016. V. 98. P. 437.
Fleer G.J., Cohen Stuart M.A., Scheutjens J.M.H.M., Cosgrove T., Vincent B. // Polymer Interfaces. London: Chapman and Hall, 1993.
Zhulina E.B., Amoskov V.M., Polotsky A.A., Birshtein T.M. // Polymer. 2014. V. 55. № 20. P. 5160.
Lebedeva I.O., Zhulina E.B., Leermakers F.A.M., Borisov O.V. // Langmuir. 2017. V. 33. № 5. P. 1315.
Léonforte F., Müller M. // ACS Appl. Mater. Int. 2015. V. 7. № 23. P. 12450.
Gillich T., Benetti E.M., Rakhmatullina E., Konradi R., Li W., Zhang A., Schlüter A.D., Textor M. // J. Am. Chem. Soc. 2011. V. 133. № 28. P. 10940.
Gillich T., Acikgöz C., Isa L., Schlüter A.D., Spencer N.D., Textor M. // ACS Nano. 2013. V. 7. № 1. P. 316.
Akkilic N., Molenaar R., Claessens M.M.A.E., Blum C., de Vos W.M. // Langmuir. 2016. V. 32. № 35. P. 8803.
Tas S., Kopec M., van der Pol R., Cirelli M., de Vries I., Bölükbas D.A., Tempelman K., Benes N.E., Hempenius M.A., Vancso G.J., de Beer S. // Macromol. Chem. Phys. 2019. V. 220. № 5. P. 1800537.
Дополнительные материалы отсутствуют.
Инструменты
Высокомолекулярные соединения (серия С)