

Водные ресурсы, 2020, T. 47, № 4, стр. 426-437
Сезонная динамика гиперсоленого озера Ойбург (Крым) как модель для изучения последствий изменения климата
И. И. Руднева a, В. Г. Шайда a, *
a ФИЦ “Институт биологии южных морей им. А.О. Ковалевского” РАН
299011 Севастополь, Россия
* E-mail: svg-41@mail.ru
Поступила в редакцию 18.01.2019
После доработки 14.05.2019
Принята к публикации 24.12.2019
Аннотация
Проведенные в 2016 г. исследования сезонных изменений некоторых гидрохимических показателей воды (солености, содержания кислорода, температуры, рН, Eh) и состояния популяции артемии в типичном гиперсоленом оз. Ойбург (с. Штормовое, Крым) четко показали их зависимость от метеорологических условий. Установлены колебания численности особей в популяции артемии, их жизненных стадий в озере, а также степень вылупления науплиев из цист рачка – от 0.5% в марте до 11.0% в феврале. Благодаря древнему происхождению гиперсоленых озер Крыма (их насчитывается 45), образовавшихся 6500–7000 лет назад, происходящие в них процессы могут быть использованы для реконструкции климатических и эволюционных событий на Земле в отдаленные геологические эпохи. Обсуждается возможность использования годовой динамики климатических, гидрохимических и экологических показателей соленого озера в модели для исследования последствий потепления климата, обусловленных этим тенденций изменения абиотических условий в водоеме и влияния их на биоту.
ВВЕДЕНИЕ
Глобальные климатические изменения и локальное антропогенное воздействие существенно влияют на состояние водных экосистем, нарушают эволюционно сложившиеся гидродинамические и гидрохимические характеристики, что приводит к истощению их ресурсов, используемых человеком. Увеличение среднегодовой температуры воздуха на планете на 1.8–4.0°C изменяет климатические условия, гидродинамику и стратификацию вод, их минерализацию, что негативно влияет на условия жизни гидробионтов [5]. Предполагается, что к концу ХХI в. температура воды в океане повысится на 2°С [18], что может привести к пагубным последствиям как для водных обитателей, так и для прибрежных территорий и акваторий, к снижению биоразнообразия, сокращению численности популяций и нарушению их структуры, воспроизводства, распределения, к модификации взаимоотношений между видами, в том числе их пищевых связей [21]. Это наносит непоправимый ущерб водным ресурсам. Происходит увеличение приходящей солнечной радиации, особенно ультрафиолетовой. Это влияет как непосредственно на живые организмы, вызывая повреждения на клеточном и молекулярном уровне, так и косвенно через фотосенсибилизирующее действие ультрафиолетовой радиации на растворенные в воде химические вещества антропогенного происхождения, что приводит к возникновению новых соединений с повышенными токсическими свойствами [24]. Все перечисленные эффекты проявляются в разной степени в зависимости от географического положения местности, устойчивости организмов к изменяющимся условиям среды обитания, их фенотипической пластичности, адаптивных возможностей и репродуктивного цикла, а также определяются интенсивностью антропогенной деятельности, сопряженной с загрязнением окружающей среды, эксплуатацией ресурсов, включая минеральные и биологические.
Гиперсоленые озера распространены в различных районах мира, часто расположены в аридных зонах с высокими температурой воздуха и испарением [28]. Они характеризуются разными соленостью, рН, минеральным составом, биоразнообразием. Эти водные объекты интенсивно используются в хозяйственной деятельности человека, так как минералы, содержащиеся в рапе, служат сырьем для предприятий химической промышленности, грунты применяются в лечебных целях в бальнеологии, косметической и фармацевтической индустрии, а основной доминирующий вид жаброногий рачок артемия – ценнейший стартовый корм для объектов аквакультуры. При этом артемия – галофильный вид, приспособленный к жизни при широком диапазоне солености. Рачок относится к фильтраторам, питается микроводорослями и взвешенными органическими частицами, что способствует просветлению воды в водоеме и обеспечению его нормального функционирования [25]. Особое внимание уделяется галофильным бактериям – источникам биологически активных веществ с уникальными свойствами, в том числе применяемых для очистки загрязненных органическими веществами соленых вод [4, 6, 7]. Кроме того, имея древнее происхождение (образовались 6500–7000 лет назад), эти водные объекты могут быть моделью, реконструирующей климатические и эволюционные процессы, происходившие на Земле в отдаленные геологические эпохи. В настоящее время интенсивное хозяйственное воздействие на гиперсоленые озера, эксплуатация их ресурсов имеют нерегламентированный характер, что наносит существенный ущерб этим уникальным экосистемам, особенно в сочетании с изменениями климата на планете.
В Крыму насчитывается более 300 озер, среди которых 45 относятся к гиперсоленым [1]. Площадь их составляет ~52 000 га, а глубина колеблется в пределах 1–4 м. Соленость также варьирует в широком диапазоне, меняется в разные периоды годового цикла. Гиперсоленые озера Крыма по происхождению делятся на две группы – приморские и внутренние, которые, как считают некоторые исследователи, в отдаленные геологические эпохи также были морскими заливами и эстуариями. По географическому расположению они разделены на 5 групп – Перекопская, Евпаторийская, Тарханкутская, Керченская и Херсонесская (рис. 1). Эти водоемы в основном используются в рекреационных и бальнеологических целях, в некоторых случаях в них добывают сырье для химической промышленности. Биологические ресурсы соленых озер Крыма до сих пор остаются недостаточно изученными, хотя в некоторых из них лечебные илы и цисты артемии изымаются браконьерским и бесконтрольным способом. Кроме того, артемия служит чувствительным индикатором экологического состояния соленых озер, в частности – уровня их распреснения, при котором популяция ракообразного погибает [3].
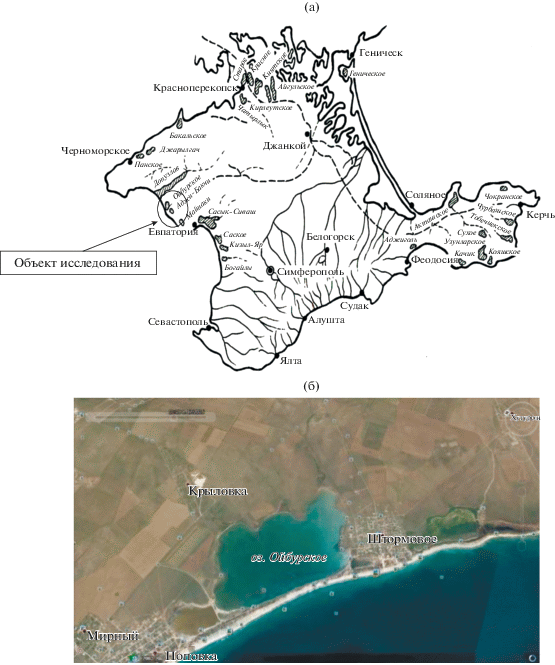
Цель настоящей работы – исследование сезонных изменений некоторых гидрохимических показателей воды и характеристик популяции артемии, обитающей в типичном гиперсоленом оз. Ойбург, относящемся к Евпаторийской группе, и их зависимости от метеорологических условий в данном районе.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЙ
Ойбурское озеро – водоем на северо-западе Сакского района между селами Поповка и Штормовое (45°16′55.8″ с.ш. и 33°04′18.7″ в.д.) (рис. 1). Озеро соленое, лиманное, бессточное. Имеет округлую форму, вытянутую с СЗ на ЮВ. Его площадь составляет 6.3 км2, длина – 4 км, средняя ширина – 1.3 км, максимальная – 1.5 км, средняя глубина – 2 м, максимальная – 3.9 м. Берега пологие, за исключением небольшого участка на ЮВ с обрывистым пляжем. На В и СЗ (у с. Поповка) к озеру прилегают солончаки. Реки не впадают. Акватория озера разделена на две основные части насыпными дамбами. Высота над уровнем моря 0.4 м. Отделено от Черного моря перешейком, по которому проходит дорога без твердого покрытия. Местами есть надводные камни. В районе озера идет интенсивная добыча песка. Согласно исследованиям сотрудников Сакской гидрогеологической режимно-эксплуатационной станции (ГГРЭС), в разрезе грязевая залежь (сверху вниз) представлена темно-серыми и черными иловыми отложениями мощностью 20 см. Поверхность грязевой залежи покрыта тонким (до 5 мм) буровато-коричневым слоем пластичной глины с примесью песчаной фракции. Минерализация рапы составляла в мае 2008 г. 97.7 г/дм3, а в августе 2015 г. – 122.5 г/дм3. Среднегодовое количество осадков ~400 мм в год. Питание осуществляется за счет поверхностных и подземных вод.
Илы озера используются для самолечения местными жителями. По берегам озера они загрязнены пластиковыми бутылками, целлофановыми пакетами и другим бытовым и строительным мусором, имеются несанкционированные мусорные свалки твердых бытовых отходов и строительных материалов. Источники загрязнения – склады ГСМ, заправочная станция, курятники и загоны для домашних животных [3].
Анализ метеорологической ситуации в районе озера
Координаты месторасположения озера и мест отбора проб определяли с помощью смартфона GSM A51OF/DS (“Samsung Electronics, CoLtd”, Южная Корея). Анализ метеорологической ситуации в районе оз. Ойбург (с. Штормовое) проводили ежемесячно, используя данные [17], на основе которых вычисляли среднюю ежемесячную температуру воздуха, давление, наличие или отсутствие облачности и осадков, направление, силу и скорость ветра.
Отбор и анализ образцов воды в озере
Температуру воды измеряли непосредственно в воде озера с помощью электронного термометра “HANNA Instruments Check Temp-1” (Россия). рН, окислительно-восстановительный потенциал (Еh), содержание кислорода определяли в лабораторных условиях с помощью анализатора Expert-001 (“Econix–Expert Moexa CoLtd”, Россия) с использованием соответствующих селективных электродов (фирма “Вольта”, Россия). Перевод Еh в редокс-потенциал Rh проводили по формуле Нернста [15, 16]:
Соленость воды (‰) измеряли с помощью рефрактометра “PAL-06S LTA GO” (Япония). Образцы воды из озера перед определением параметров хранили в холодильнике при +4°C в течение 4–6 ч после отбора проб.Отбор и анализ цист артемии
В образцах воды определяли наличие разных онтогенетических стадий артемии обычными гидробиологическими методами с помощью камеры Богорова. Для этого цисты собирали в трех–четырех строго определенных местах согласно методу, описанному в [27], ежемесячно в течение 2016 г. Отбор образцов проводили в воде с помощью специального сконструированного авторами статьи устройства, представляющего собой пластиковую емкость объемом 5 л, на дне которой находилась сетка с величиной ячеи 50 микрон. Через данное устройство пропускали 5 л воды и вели подсчет цист, науплиев и взрослых особей артемии, осевших на сетку, из расчета количество экземпляров в 1 л воды.
Для определения качества цист артемии их собирали на побережье озера ежемесячно в течение 2016 г., отмывали согласно общепринятым рекомендациям [8, 27], после чего образцы помещали в стандартные условия (среда инкубации соленостью 35‰, температура +25°С) при периодическом перемешивании для определения доли вылупления науплиев из цист, которую рассчитывали как отношение числа выклюнувшихся через 48 ч личинок к числу помещенных в инкубационную среду цист.
Статистический анализ результатов
Все определения проводили в трех повторностях. Статистические различия между исследуемыми показателями находили с помощью теста Стьюдента при уровня значимости р < 0.05 [2]. Корреляционный анализ проводили с помощью компьютерной программы CURFVIT (версия 2.10–L).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Сезонная динамика метеорологических показателей в районе расположения оз. Ойбург
Метеорологические характеристики в районе оз. Ойбург приведены на рис. 2–4 и в табл. 1, где представлен анализ погоды в момент отбора проб воды и артемии в озере.
Рис. 2.
Сезонные колебания среднемесячной температуры воздуха, скорости ветра, атмосферного давления и количества дней с выпадением атмосферных осадков в районе оз. Ойбург.
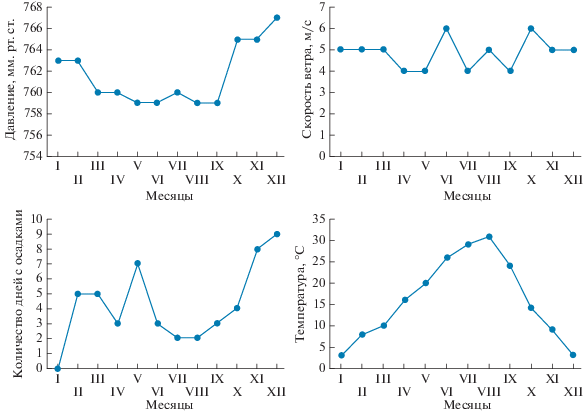
Рис. 3.
Сезонное соотношение ясных, малооблачных, облачных и пасмурных дней в районе оз. Ойбург в 2016 г.
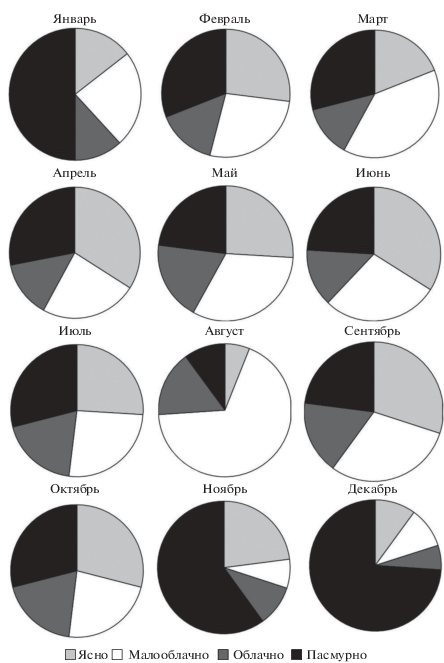
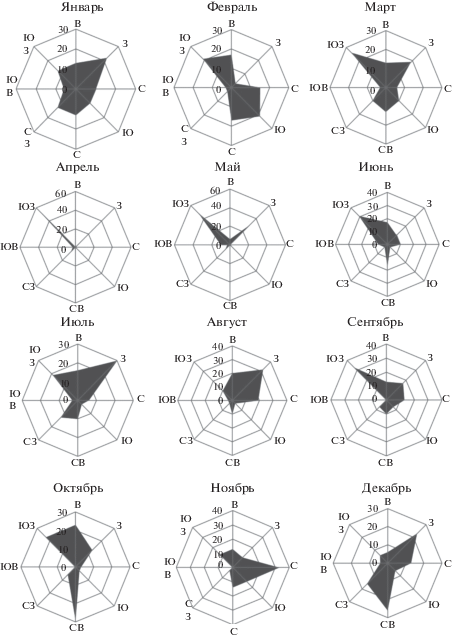
Таблица 1.
Характеристика метеорологической обстановки в период отбора проб в районе оз. Ойбург в 2016 г.
| Дата и состояние атмосферы | Температура воздуха, °С |
Давление, мм рт. ст. |
Направление ветра | Скорость ветра, м/с |
|---|---|---|---|---|
| 6 января, пасмурно | +2 | 760 | СВ | 6 |
| 10 февраля, ясно | +11 | 761 | Ю | 3 |
| 6 марта, малооблачно | +17 | 761 | Ю | 2 |
| 2 апреля, облачно | +12 | 764 | СЗ | 3 |
| 2 мая, ясно | +19 | 764 | СВ | 11 |
| 18 июня, малооблачно | +28 | 764 | ЮЗ | 4 |
| 6 июля, ясно | +28 | 760 | ЮВ | 3 |
| 1 августа, малооблачно | +35 | 759 | З | 5 |
| 1 сентября, малооблачно | +25 | 764 | В | 6 |
| 1 октября, ясно | +24 | 764 | З | 4 |
| 1 ноября, пасмурно | +8 | 764 | С | 5 |
| 1 декабря, пасмурно, снег | +2 | 761 | ЮЗ | 5 |
Как видно на рис. 2–4, атмосферное давление незначительно снижалось до 759 мм рт. ст. в весенне-летний период, но затем увеличивалось в холодное время года до 767 мм рт. ст. (рис. 2). Среднемесячная температура воздуха последовательно возрастала в теплый сезон, достигает максимума в июле–августе (+31°С), после чего падала до минимума (+3°С) в декабре–январе. Было несколько дней с минусовой температурой воздуха в зимнее время, вследствие чего среднемесячная температура воздуха была положительной в течение всего 2016 г. Средняя скорость ветра варьировала в пределах 4–6 м/с, наибольшая отмечена в июне и октябре. Максимальное количество дней с осадками пришлось на декабрь (9 дней) и май (7 дней), тогда как в остальные месяцы 2016 г. количество дней с осадками было 0–5, что свидетельствует о тенденции формирования засушливого климата в данном районе. Это подтверждается сокращением количества пасмурных и облачных дней в весенне-летний период, особенно в июле–сентябре, и постепенного их увеличения в ноябре–декабре (рис. 3).
Формирование погодных условий во многом зависит от направления ветра, которое также менялось в зависимости от времени года, что можно видеть на рис. 4: если в декабре–январе преобладали западные ветры, в феврале доминировали юго-западные и южные, а в марте–июне – юго-западные, в июле–августе – западные, в сентябре – юго-западные, в октябре – северо-восточные, а в ноябре – северные. Преобладание западных морских ветров в этом районе способствовало понижению температуры воздуха и воды, образованию облачности, снижению интенсивности испарения, особенно в летний период. Направление и сила ветра способствовали перемешиванию вод озера, переливу морской воды через дамбу во время зимних штормов и попаданию ее в акваторию, изменению стратификации и гидрохимических характеристик вод, а также распределению планктонных организмов, в частности артемии. Анализ погодных условий в дни отбора проб в оз. Ойбург представлен в табл. 1. Погодные условия в дни исследований соответствовали среднестатистическим для тестируемого месяца (рис. 2–4).
Сезонная динамика некоторых гидрохимических характеристик воды в оз. Ойбург в 2016 г.
Соленость воды в озере в период 2016 г. колебалась от 91‰ в апреле до 155‰ в августе (рис. 5). Температура воды менялась от –5.4°С в январе до +28.8°С в июне и августе. Содержание кислорода в воде также колебалось от 8.3 мг/л в сентябре до 13.8 мг/л в декабре. Величины рН, Eh и Rh варьировали в меньшей степени, однако наименьшее значение рН (6.60) отмечено в январе, наибольшее (7.38) – в июне и июле, Rh –19.9 условных единиц, самое низкое значение Eh ( –51.5 мВ) отмечено в июне, самое высокое (–8 мВ) – в январе.
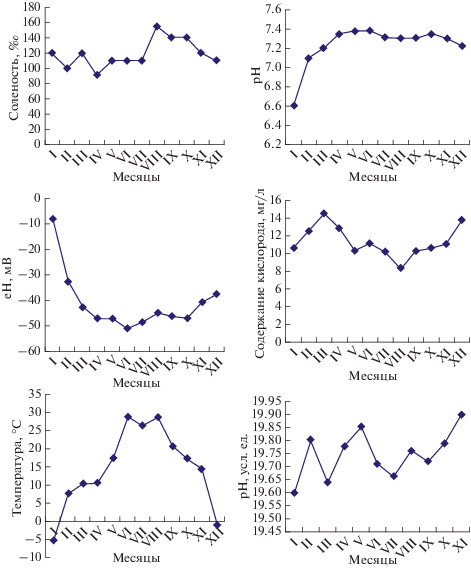
Обнаружена высокая корреляция между температурой воздуха и температурой воды в озере (r = 0.88, р < 0.05), между температурой воды, соленостью и содержанием кислорода не выявлено прямой зависимости, высокая корреляция установлена между значениями рН, Eh и Rh (r = 0.96, р < 0.05). Совокупность физико-химических условий в воде озера и их изменение в течение года существенно повлияли на состояние популяции артемии (табл. 2).
Таблица 2.
Сезонная динамика распределения и численности, экз/л, разных онтогенетических стадий артемии в оз. Ойбург (в скобках – среднее из трех–пяти определений)
| Месяц | Цисты | Науплии | Метанауплии | Взрослые |
|---|---|---|---|---|
| Январь | На поверхности льда | 0 | 0 | 0 |
| Февраль | На берегу, в воде 0–8 (5 ± 1) | 0 | 0 | 0 |
| Март | На берегу, в воде 1–8 (5 ± 2) | 0 | 0 | 0 |
| Апрель | В воде 1–4 (2 ± 1) | 0–1 (1 ± 0) | 0 | 0 |
| Май | В воде 1–56 (12 ± 7) | 1–4 (2 ± 1) | 1–8 (5 ± 2) | 8–16 (12 ± 3) ювенильные |
| Июнь | В воде 6–27 (17 ± 5) | 1–4 (2 ± 1) | 1–10 (7 ± 1) | 13–42 (16 ± 3) ювенильные, 1 половозрелая самка |
| Июль | В воде 40–326 (263 ± 86) | 5–220 (153 ± 28) | 5–120 (86 ± 18) | 2–25 (14 ± 3), половозрелые и неполовозрелые самки, есть мертвые |
| Август | В воде 105–432 (278 ± 96) | 12–162 (87 ± 12) в том числе мертвые |
6–165 (75 ± 23) в том числе мертвые |
1 половозрелая самка, мертвые раки и их фрагменты |
| Сентябрь | В воде 105–348 (186 ± 57) | 0 | 8–123 (66 ± 18) в том числе мертвые |
Мертвые половозрелые самки |
| Октябрь | В воде 105–260 (125 ± 75) | 0 | 0 | Мертвые половозрелые самки и их фрагменты |
| Ноябрь | На берегу, в воде 1–14 (5 ± 2) | 1–6 (3 ± 1) |
1–4 (2 ± 1) |
То же |
| Декабрь | На берегу, в воде 1–6 (4 ± 1) | 0 | 0 | 0 |
Сезонные изменения состояния популяции артемии
В холодные месяцы в январе–марте цисты были найдены на льду озера и в отдельных местах на побережье в небольшом количестве (табл. 2). В воде также обнаружены яйца (1–8 экземпляров в 1 л). В апреле начался массовый выклев науплиев из цист в разных частях акватории озера. В весенне-летний сезон в воде присутствовали все онтогенетические стадии артемии в значительных количествах. При этом в июле и августе обнаружены мертвые раки, в сентябре их число существенно увеличилось. В октябре в воде озера не были найдены ранние онтогенетические стадии артемии, тогда как в прибойной зоне обнаружено большое количество отмерших организмов. В ноябре встречались единичные науплии, также присутствовали мертвые самки, а на берегу – цисты. В декабре на берегу и в воде находили цисты ракообразного.
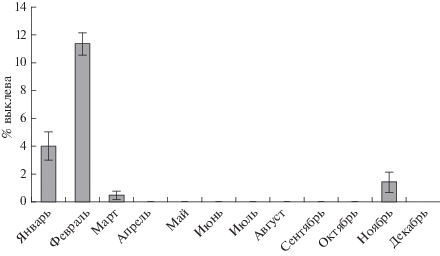
В образцах цист, собранных на берегу, определена доля выклева науплиев (рис. 6). Она оказалась невысокой, в зимние месяцы выше (4% в январе, 11.3% в феврале), чем в марте (0.5%) и в ноябре (1.4%), что свидетельствует о разном качестве яиц артемии в разные периоды годового цикла, и это следует учитывать при их сборе для аквакультурных и рыбоводческих целей.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Для понимания механизмов функционирования водных экосистем в районах с экстремальными условиями, в том числе в зонах с засушливым климатом, необходимо проводить комплексное изучение факторов, определяющих динамику параметров водных объектов в течение определенного периода времени. Для этого важно знать годовой цикл основных показателей как непосредственного состояния водного объекта и его ресурсов, так и внешних условий, влияющих на это состояние. С этой целью в настоящем исследовании изучена взаимосвязь годовой динамики метеорологических характеристик в районе оз. Ойбург и сезонных колебаний некоторых его гидрохимических и гидробиологических параметров. Учитывая тот факт, что это гиперсоленое озеро расположено в зоне активной хозяйственной деятельности, полученные результаты могут иметь и практическое приложение и быть учтены при разработке способов оптимального использования его ресурсов – как минеральных, так и биологических. Более того, в летние месяцы при достаточно высокой температуре воздуха ( >+30°С) и минимальном количестве осадков или их полном отсутствии оно может быть моделью для описания и прогнозирования изменений, обусловленных потеплением климата и его последствиями для водных экосистем. При этом в зонах с засушливым климатом достаточно много эстуариев и внутренних водоемов, соленость которых существенно выше, чем в океане и морях. Часто они располагаются в непосредственной близости к морю. В теплое время года эти водные объекты могут высыхать, затем снова наполняться морской водой или осадками вплоть до их значительного распреснения. Таким образом, гиперсоленость для таких водоемов – сезонное явление, она обусловлена интенсивным испарением в теплый период года, тогда как для других водных объектов высокая минерализация постоянна [28].
Настоящие исследования показали, что в апреле–мае в районе оз. Ойбург начинался резкий подъем температуры воздуха, которая достигала максимальных значений в июле–августе (>+30°С, максимум +35°С), после чего она постепенно снижалась до минимальных значений в декабре–январе (+2°С). Атмосферное давление изменялось незначительно и противоположно температуре – было выше в холодное время года, снижалось в теплый сезон. Однако не обнаружено прямой корреляции между изменением температуры воздуха и колебаниями давления в районе оз. Ойбург. Количество дней с осадками зависело от сезона: в январе 2016 г. осадков не было, в феврале–июне – 3–7 дней, затем в июле–августе – 2, после чего в сентябре и в последующие месяцы число последовательно возрастало, достигало максимума в декабре. Эта ситуация коррелировала с количеством пасмурных и ясных дней в исследуемом районе, в осенне-зимний период преобладали облачные дни, а в весенне-летний – ясные или малооблачные.
Ветер определяет интенсивность перемешивания вод и их вертикальную стратификацию, а также перелив морской воды через песчаный барьер в озеро [14, 19]. Для района оз. Ойбург характерна скорость ветра 4–6 м/с. В то же время температура воздуха, давление и количество осадков во многом зависят от направления ветра. Если в первую половину 2016 г. в основном преобладали южные и юго-западные ветры, то с июля–августа – западные, в сентябре – вновь юго-западные, в октябре и ноябре – северные и северо-восточные, в декабре – западные и северо-восточные. Изменение направления ветра и его скорости также влияло на выпадение осадков, гидрологические характеристики озера, обусловленные перемешиванием воды в водоеме, на распределение биоты.
Соленость и рН в гиперсоленых водоемах существенно колеблются в течение дня и в течение сезонов, что определяется как концентрацией отдельных ионов, так и доминирующими буферными системами, соотношение которых в процессе испарения может меняться, что приводит к сдвигу рН [11, 20, 26]. В воде оз. Ойбург отчетливо прослеживались сезонные колебания солености: минерализация рапы была ниже с февраля по июль, после этого резко возрастала в августе (максимум 155‰), затем постепенно снижалась. Наиболее высокие показатели солености совпадали с засушливым периодом на фоне резкого повышения температуры воздуха. В этом случае наблюдали частичное пересыхание отдельных участков озера, что характерно и для других водоемов в жаркое время года. При возрастании минерализации вследствие интенсивного испарения возможно повышение концентрации различных токсикантов, содержащихся в воде, в том числе попадающих туда в результате нерегламентированной хозяйственной деятельности человека [10, 12], что может представлять опасность для биоты и для человека. Следует отметить, что при изменении климата и дальнейшем повышении температуры процессы испарения в озере могут проходить более активно, минерализация нарастать, как и содержание в рапе опасных веществ, что отмечено в ряде публикаций [24].
Максимальные значения температуры воды (28.8°С) в озере отмечены в июне–августе, после чего она уменьшалась и достигала минимальных значений в январе и декабре. Температура воды в водоеме была, как правило, на 3–4°С ниже, чем температура воздуха, что согласуется с данными [19]. Обнаружена высокая корреляция между температурой воды и воздуха (r = 0.96), тогда как между температурой воды и соленостью не установлено прямой зависимости. Это может быть обусловлено тем, что озеро питается грунтовыми водами, которые поддерживают соленость на определенном уровне и подпитывают водоем даже при критически высокой температуре, что не допускает его полного пересыхания. Далее, во время штормов происходит перелив морской воды через песчаный барьер, который отделяет озеро от моря. Следствие этого – не только снижение солености рапы, но и насыщение ее кислородом. Сходные явления наблюдали и в других гиперсоленых озерах в прибрежных зонах морей и океанов [9, 22]. Содержание кислорода было существенно ниже в теплое время года по сравнению с холодным, что обусловлено процессами эвтрофирования в данном водоеме в наиболее жаркие летние месяцы (июнь–август), после чего уровень его заметно повышался.
На экологическое состояние водоема и условия существования гидробионтов в нем существенное влияние оказывают активная реакция рН среды, Eh и Rh, от которых зависят фотосинтез, дыхание и окислительно-восстановительные процессы в водоеме [13]. Известно, что в естественных водоемах Eh колеблется от –400 до +700 мВ, что определяется совокупностью всех происходящих в нем окислительных и восстановительных процессов. В воде оз. Ойбург в течение года этот показатель последовательно снижался от –8 мВ в январе до –51.5 мВ в июне, после чего опять возрастал. Rh – показатель химической и биологической активности воды, который существенно влияет на жизнедеятельность гидробионтов. pH и Rh воды взаимосвязаны: чем выше Rh, тем ниже pH. Значения этих показателей, полученные в настоящих исследованиях, свидетельствуют о том, что в течение года Rh менялся незначительно – в пределах 19.6–19.9, что соответствует среде со слабыми восстановительными свойствами (15–20 условных единиц) [17]. В то же время возрастание Rh означает увеличение окислительных свойств воды. Низкий показатель Rh благоприятен для анаэробных организмов, т.е. некоторых микробов, грибов, вирусов.
В водной среде постоянно происходят как окислительные, так и восстановительные реакции. При загрязнении водоема и попадании в него большого количества органических веществ, при разложении отмерших частей растений и животных, а также при эвтрофировании Rh может резко возрасти. Окислительные процессы уменьшают рН, восстановительные – способствуют увеличению рН. В свою очередь, величины рН влияют на Rh. Rh имеет более низкие значения в придонных слоях воды, а у поверхности грунта он больше, чем в самом грунте. Поскольку величина рН колеблется в течение суток, меняется и величина Rh, зависящая также от температуры [15, 16]. В настоящих исследованиях установлено возрастание Rh в мае и ноябре–декабре. В первом случае это обусловлено бурным развитием фитопланктона в воде озера, во втором – увеличением количества органических веществ за счет отмирания артемии и прекращения вегетации микроводорослей, что согласуется с данными о динамике популяции рачка (табл. 2).
Организмы-фильтраторы, к которым относится артемия, – ключевые компоненты многих водных экосистем. Они оказывают существенное влияние на их динамику, в том числе сезонную. В связи с этим исследования сезонных флуктуаций количества и состава популяции артемии в соленых водоемах имеет важное значение для понимания функционирования экосистемы, в том числе в условиях изменения окружающей среды и потепления климата. Поскольку артемия – доминирующий вид в гиперсоленых озерах, изменения в ее популяции отражают динамику процессов в экосистеме, происходящих под влиянием внешних факторов, включая климатические [23]. Жизненный цикл ракообразного в оз. Ойбург зависит от сезонных изменений в водоеме. Об этом свидетельствуют как наличие и флуктуации численности различных онтогенетических стадий рачка, так и доля выклева науплиев из цист. Увеличение выклева науплиев из цист до 11% в зимний период по сравнению с величиной этого показателя в другие сезоны обусловлено низкой температурой и отсутствием осадков. Именно снижение солености воды весной в результате таяния снега и атмосферных осадков приводит к необратимой гидратации цист, стимулирует эмбриогенез, но дальнейшее развитие личинок невозможно в условиях пониженной температуры. Этим объясняется резкое снижение доли выклева науплиев из цист, собранных в марте и последующие месяцы. Полученные данные могут быть полезны для организации добычи цист артемии в оз. Ойбург.
На основании проведенных исследований показано, что экологическое состояние гиперсоленого оз. Ойбург имело четко выраженную сезонную изменчивость, связанную с климатическими особенностями территории. Увеличение температуры воздуха в летний период вплоть до экстремальных значений, отсутствие осадков и сильное испарение – важнейшие факторы изменения гидрохимических характеристик озера, приведшие к повышению температуры воды, солености, снижению содержания в ней кислорода, Eh и Rh. Сезонные изменения, происходящие в исследуемом водоеме, могут служить в качестве модели для изучения последствий изменения климата. Показательна динамика численности различных жизненных стадий артемии, которая широко распространена повсюду на планете и адаптирована к экстремальным условиям существования, включая повышенную температуру, низкое содержание кислорода и высокую соленость, а также колебания этих параметров в течение суток и года. Комплексное исследование сезонных флуктуаций метеорологических, гидрохимических и гидробиологических характеристик гиперсоленых водоемов может дать существенную информацию для понимания последствий изменения климата как в глобальных масштабах, так и в конкретных географических районах, в которых наблюдается тенденция к формированию засушливого климата, а также будет способствовать разработке мероприятий по оптимальному использованию и охране их ресурсов.
ВЫВОДЫ
Установлена высокая корреляция между температурой воздуха в районе расположения оз. Ойбург и температурой воды в водоеме (r = 0.96).
В результате увеличения температуры воздуха и воды в летний период соленость в озере увеличилась в 1.7 раза по сравнению с осенними, зимними и весенними месяцами, а содержание кислорода в воде снизилось.
Сезонные изменения рН, Eh, Rh были выражены в меньшей степени.
В холодное время года в воде и на берегу озера обнаружены цисты артемии, первые науплии появляются в апреле, все онтогенетические стадии рачка выявлены в мае–июне. В августе при максимальной температуре воды начинается массовая гибель рачков и в воде отмечено большое количество мертвых особей.
Наблюдаемые сезонные изменения гидрохимических показателей воды и состояния популяции артемии в оз. Ойбург обусловлены климатическими флуктуациями. Динамика этих изменений может быть использована для анализа последствий изменения климата.
Список литературы
Гулов О.А. Экоцид крымских соляных озер // Теория и практика восстановления внутренних водоемов. СПб.: Лема, 2007. С. 60–78.
Халафян А.А. Статистика 6. Гл. 8. М.: Биом, 2008. С. 133–152.
Чабан В.В. Применение биоиндикатора Artemia salina при экологическом изучении соленых озер Крымского полуострова // Стр-во и техногенная безопасность. 2012. № 43. С. 102–105.
Aljohny B.O. Halophilic bacterium-A review of new studies (Review) // Biosci. Biotechnol. Res. Asia. 2015. V. 12. Is. 3. P. 2061–2069.
Bradley M., van Putten I., Sheaves M. The pace and progress of adaptation: marine climate change preparedness in Australia’s coastal communities // Marine Policy. 2015. V. 53. P. 13–20.
Di Meglio, Santos L., Gomariz F., Almansa M., López C.C., Antón J., Nercessian D. Seasonal dynamics of extremely halophilic microbial communities in three Argentinian salterns (Article) // FEMS Microbiol. Ecol. 2016. V. 92. Is. 12. Article number fiw184.
Edbeib M.F., Wahab R.A., Huyop F. Halophiles: biology, adaptation, and their role in decontamination of hypersaline environments (Review) // World J. Microbiol. Biotechnol. 2016. V. 32. Is 8. Article number 135.
El-Magsodi M.O., El-Ghebli H.M., Hamza M., Van Stappen G., Sorgeloos P. Characterization of Libyan Artemia from Abu Kammash Sabkha // Libyan J. Marine Sci. 2005. V. 10. P. 19–29.
Geldenhuys Ch., Cotiyane Ph., Rajkaran A. Understanding the creek dynamics and environmental characteristics that determine the distribution of mangrove and saltmarsh communities at Nahoon Estuary // South African J. Botany. 2016. V. 107. P. 137–147.
Glavaš N., Šmuc N.R., Dolenec M., Kovač N. The seasonal heavy metal signature and variations in the microbial mat (petola) of the Sečovlje Salina (northern Adriatic) // J. Soils and Sediments. 2015. V. 15 Is. 12. P. 2359–2368.
Golan R., Itta Gavrieli I., Ganor J., Lazarc B. Controls on the pH of hyper-saline lakes – A lesson from the Dead Sea // Earth and Planetary Sci. Letters. 2016. V. 434. P. 289–297.
Guesdon G., Santiago-Martin A., Raymond S., Messaoud H., Michaux A., Roy S., Galvez R. Impacts of salinity on Saint-Augustin Lake, Canada: Water Remediation measures of watershed scale // Water (Switzerland). 2016. V. 8. Is. 7. Article № 285.
Hargrave B.T., Holmer M., Newcombe C.P. Towards a classification of organic enrichment in marine sediments based on biochemical indicators // Marine Pollut. Bull. 2008. V. 56. P. 810 –824.
Hetzel Y., Pattiaratchi C., Lowe R., Hofmeister R. Wind and tidal mixing controls on stratification and dense water outflows in a large hypersaline bay // J. Geophys. Res.: Oceans, 2015. V. 120. Is. 9. P. 6034–6056.
http://liveinsuccess.ru/voda-ph-eh-rh/
http://ribosite.ucoz.ru/publ/1-1-0-17
https://www.gismeteo.ru/
IPCC. Report Fifth Assessment Report on Climate Change. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change / Eds Stocker T.F., Qin D., Plattner G-K., Tignor M., Allen S.K., Boschung J., Nauels A., Xia Y., V Bex V., Midgley P.M. Cambridge: Cambridge Univ. Press, 2013.
Kompaniets L.A., Iakubailik T.V. Observations of hydrophysical characteristics of stratified salt lake Shira (Siberia) as an important part of environmental monitoring. // 15th Int. Multidisciplinary Sci. GeoConf. SGEM 2015. V. 2(3). P. 635–641.
Li L., Song W., Deng C., Zhang D., Al-Misned F.A., Mortuza M.G., Gadd G.M., Pan X. Effects of pH and salinity on adsorption of hypersaline photosynthetic microbial mat exopolymers to goethite: A study using a quartz crystal microbalance and fluorescence spectroscopy // Geomicrobiol. J. 2016. V. 33. Is. 3–4. P. 332–337.
Madeira D., Araújo J.E., Vitorino R., Capelo J.L., Vinagre C., Diniz M.S. Ocean warming alters cellular metabolism and induces mortality in fish early life stages: A proteomic approach // Environ. Res. 2016. V. 148. P. 164–176.
Mitchell S., Boateng I., Couceiro F. Influence of flushing and other characteristics of coastal lagoons using data from Ghana // Ocean and Coastal Management. 2017. V. 143. Is. 1. P. 26–37.
Sánchez M.I., Paredes I., Lebouvier M., Green A.J. Functional role of native and invasive filter-feeders, and the effect of parasites: Learning from hypersaline ecosystems // PLoS ONE. 2016. V. 11. Is. 8. Article number e0161478.
Schiedeck D., Sundelin B., Readman J.W., Macdonald R.W. Interactions between climate change and contaminants // Marine Pollut. Bull. 2007. V. 54. P. 1845–1856.
Shadkam S., Ludwig F., van Vliet T.H., Pastor A., Kabat P. Preserving the world second largest hypersaline lake under future irrigation and climate change. // Sci. Total. Environ. 2016. V. 559. P. 317–325.
Sirota I., Arnon A., Lensky N.G. Seasonal variations of halite saturation in the Dead Sea // Water Resour. Res. 2016. V. 52. Is. 9. P. 7151–7162.
Van Stappen G. Use of cysts // FAO Fisheries Technical Paper. 1996. № 36. P. 102–123.
Wooldridge T.H., Adams J.B., Fernandes M. Biotic responses to extreme hypersalinity in an arid zone estuary // South Africa J. Botany. 2016. V. 107. P. 160–169.
Дополнительные материалы отсутствуют.