

Водные ресурсы, 2021, T. 48, № 1, стр. 52-60
Динамика морфофункциональных групп фитопланктона Рыбинского водохранилища и оценка качества его вод по индексу сообществ
Л. Г. Корнева a, *, В. В. Соловьева a
a Институт биологии внутренних вод им. И.Д. Папанина РАН
152742 Ярославская обл., пос. Борок, Россия
* E-mail: korneva@ibiw.ru
Поступила в редакцию 14.08.2019
После доработки 17.03.2020
Принята к публикации 09.06.2020
Аннотация
На основе результатов исследования фитопланктона разных плесов Рыбинского водохранилища в 2000–2005 гг. проведен анализ изменения соотношения видов, представляющих различные морфофункциональные группы, выделенные по физиологическим, морфологическим и экологическим показателям [29]. Дана оценка качества вод по индексу сообществ Q, разработанного с учетом пропорций этих групп, и проведено сопоставление результатов этой оценки с полученной по значениям индекса сапробности Пантле–Букк и биомассы фитопланктона.
В современную эпоху, когда из-за увеличения населения, урбанизации, индустриализации и химизации сельского хозяйства наблюдается резкое ухудшение качества пресных вод, необходим регулярный мониторинг качества водных ресурсов, необходимый для сохранения их удовлетворительного состояния и рационального использования. Для этого наряду с физико-химическими показателями широко используют биологические. Фитопланктон как первичный продуцент органического вещества и источник кислорода в воде – ключевой индикатор состояния водной среды. До сих пор оценка качества вод Рыбинского водохранилища (Верхняя Волга) проводилась с помощью индекса сапробности Пантле–Букк в модификации Сладечека [1, 5, 9, 16] и по значениям биомассы фитопланктона [5, 7]. Однако эти показатели позволяют оценить только степень органического загрязнения вод и их уровень трофии. Для реализации Европейской Водной рамочной директивы с целью сохранения и восстановления водных экосистем предложено оценивать экологический статус водоемов по структурным показателям фитопланктона [20]. Для этого с учетом экологических преференций отдельных видов планктонных водорослей в Норвегии, Австрии и Словении используют индекс BI (Brettum Index), в Германии – PTSI (Phytoplankton Taxa Seen Index), а в Швеции и Италии – TPI (Trofiskt planktonindex) [24]. К наиболее простым и удобным показателям экологического состояния водоемов можно отнести “венгерский” индекс сообществ (Q) [25], расчет которого основан на изменении соотношения функциональных групп фитопланктона, различающихся по физиологическим, морфологическим и экологическим признакам [23, 29]. В разработанной классификации этих групп каждая имеет буквенно-цифровой код. Помимо определенной совокупности видов водорослей, код включает в себя характеристики среды обитания, формирующей этот комплекс. Авторы классификации исходили из того, что сообщество в целом может быть более показательным параметром состояния вод, чем отдельные индикаторные виды. Данная система создавалась в ходе исследований, проводимых на озерах в Англии, и была успешно применена и для рек [19, 27]. Использование индекса Q рекомендовано для разных регионов мира. Его преимущество состоит в том, что он не предназначен для оценки последствий какой-либо одной формы антропогенного воздействия [25]. К настоящему времени число выделенных функциональных групп водорослей приближается к 40, они различаются по количеству входящих в них таксонов [26].
Цель настоящего исследования – проследить динамику морфофункциональных групп фитопланктона различных плесов Рыбинского водохранилища, дать оценку качества вод Рыбинского водохранилища с помощью Q и сравнить ее результаты с полученными по значениям индекса сапробности и биомассы фитопланктона.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Для этой цели проведен анализ таксономического состава и биомассы фитопланктона в 203-х пробах, собранных на 30-ти станциях (рис. 1), расположенных в различных плесах Рыбинского водохранилища с мая по октябрь 2000–2005 гг. В водохранилище (площадь 4550 км2, средняя глубина 5.6 м, коэффициент условного водообмена 1.9) условно выделяют центральное озеровидное расширение и участки, расположенные по бывшим руслам рек Волги, Мологи и Шексны. Воды различных плесов различаются по гидрологическим и физико-химическим показателям [17, 18].
Рис. 1.
Картосхема территории расположения станций в Волжском (I), Моложском (II), Шекснинском (III) и Главном (IV) плесах Рыбинского водохранилища в 2000–2005 гг.
Отбор проб проводили пластмассовым метровым батометром системы Элгморка объемом 4 л последовательно с каждого метрового горизонта от поверхности до дна, смешивая затем в одну интегральную. Пробы воды концентрировали путем прямой фильтрации поочередно через мембранные фильтры с диаметром пор 5 и 1.2 мкм под давлением не более 20.3–30.4 кПа. Консервацию живого фитопланктона проводили раствором Люголя с добавлением формалина и ледяной уксусной кислоты. Численность водорослей оценивали в камере “Учинская-2” объемом 0.02 мл при 450-кратном увеличении, биомассу определяли обычным счетно-объемным методом [11]. Виды, составлявшие ≥10% общей биомассы фитопланктона, относили к доминирующим. Оценку качества воды проводили с помощью индекса сообществ Q:
где Pi – доля биомассы i-й функциональной группы в общей биомассе фитопланктона; F – коэффициент (фактор F), установленный для i-й функциональной группы для определенного типа водоема по [25].Поскольку доминирующие виды в 81% случаев в сумме составляли >85% общей биомассы, авторы посчитали возможным использовать для расчета индекса Q соотношение биомассы только ведущих, входящих в состав отдельных функциональных видов групп фитопланктона в соответствии с [25, 26, 29]. Правомерность такого подхода подтверждалась и высоким значением коэффициентa корреляции (0.9) между индексами, рассчитанными по биомассе доминирующих видов и совокупности всех видов в пробе. Расчет Q проводили для каждой пробы с последующим осреднением по плесам и водохранилища в целом. Соответствие видов кодам различных функциональных групп проводили согласно предложенным классификациям [25, 26, 29]. Диапазон значений коэффициентов F (номера фактора) для выявленных функциональных ассоциаций водорослей в планктоне Рыбинского водохранилища заимствовали из типологии, разработанной для европейских озер 2-го типа – щелочных, где преобладают гидрокарбонатные ионы, с площадью акватории >100 км2 [25]. Величина фактора F в таких водоемах варьирует от 0 до 5. Его наибольшее значение (5) присваивают комплексам видов, обитающих в стабильных природных условиях, неизмененных при внешнем воздействии, а самое низкое (0) – тем, что обитают в сильно нарушенных экосистемах. При плохом качестве поверхностных вод Q приобретает наименьшие значения – 0–1, при удовлетворительном – 1–2, при умеренном – 2–3, при хорошем – 3–4 и при высоком – 4–5 [25].
Оценку сходства состава доминирующих видов проводили с помощью индекса Сёренсена [12]. Сапробность оценивали с помощью индекса Пантле–Букк в модификации Сладечека [32]. При этом индикаторную значимость видов определяли по спискам [37]. Оценку качества воды проводили по биомассе фитопланктона и сапробности согласно критериям, разработанным для комплексной экологической классификации качества поверхностных вод суши [14]. При оценке связи Q с другими параметрами качества воды использовали его значения, рассчитанные по совокупности всех видов, обнаруженных в пробе, имеющих код отдельных функциональных групп. Оценку связи биотических и абиотических параметров осуществляли с помощью коэффициента детерминации для линейной регрессии и коэффициента ранговой корреляции Спирмена.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В 2000–2005 гг. состав доминирующих комплексов фитопланктона Рыбинского водохранилища был сформирован цианобактериями (Cyanobacteria) – 9 видoв водорослей, диатомовыми (Bacillariophyta) – 19, золотистыми (Chrysophyta) – 1, криптофитовыми (Cryptophyta) – 4, динофитовыми (Dinophyta) – 2 и зелеными (Chlorophyta) – 5. Их максимальное количество отмечено в планктоне Волжского плеса (28 видов), которое плавно снижалось в Главном (23), Шекснинском (18) и Моложском (14) плесах. Наибольшим сходством (по коэффициенту Сёренсена) доминирующего состава характеризовался фитопланктон северных плесов – Шекснинского и Моложского (71%), наименьшим – Главного и Шекснинского (14%). Большинство доминантов соответствовало определенным функциональным группам водорослей (табл. 1), каждая из которых имеет буквенный код в разработанной шкале [26, 29], за исключением диатомовых: Cyclostephanos invisitatus (M.H. Hohn, Heller.) E.C. Ther., Stoermer and Håk., который входил в состав доминирующего комплекса с 1978 г. [4], и Acanthoceras zachariasii (Brun) Sim., который лидировал в 2005 г. [7], а также из динофлагеллят Peridiniopsis kevei Grigorszky et Vasas (среди доминантов с 2005 г.). В соответствии с результатами, полученными на р. Нарве [27], Stephanodiscus binderianus (Kütz.) Krieg. отнесли к группе B.
Таблица 1.
Представленность различных функциональных групп (код) фитопланктона в плесах Рыбинского водохранилища в 2000–2005 гг. (здесь и в табл. 2, на рис. 2: В – Волжский, М – Моложский, Ш – Шекснинский, Г – Главный плесы; F – значение фактора F; 1 – доминирование, 0 – отсутствие доминирования; LO – крупноклеточные динофлагелляты, некоторые хроококковые цианобактерии, обитатели мелководных и глубоководных, олиго-эвтрофных средних и больших озер; Н2 – азотфиксирующие цианобактерии (Anabaena lemmermannii P.G. Richter, Gloeotrichia echinulata P.G. Richter), толерантные к низкому содержанию биогенных веществ и чувствительные к перемешиванию и световому лимитированию, обитатели олиго-мезотрофных стратифицированных озер или мезотрофных мелководных озер с хорошими световыми условиями; М – виды рода Microcyctis, предпочитающие эвтрофные-гипертрофные небольшие водоемы, толерантные к высокой световой инсоляции и чувствительные к проточности и низкой освещенности; H1 – азотфиксирующие цианобактерии эвтрофных, стратифицированных и мелководных озер с низким содержанием азота, чувствительные к перемешиванию воды, световому лимитированию, низкому содержанию фосфора; Е – золотистые водоросли (Dinobryon, Mallomonas, Synura), предпочитающие мелководные олиготрофные водоемы, толерантные к низкому содержанию биогенных веществ в воде, способные переходить к миксотрофии, чувствительные к дефициту CO2; В – диатомовые водоросли, обитатели перемешиваемых мезотрофных малых и средних озер, толерантные к световому лимитированию, чувствительные к стратификации и повышению рН; D – диатомовые, предпочитающие мелководные хорошо перемешиваемые воды, включая реки, чувствительные к снижению биогенов; С – виды, предпочитающие перемешиваемые малые и средние эвтрофные озера, толерантные к световому и углеродному лимитированию, чувствительные к стратификации и снижению концентрации кремния; P – виды, обитающие в мелководных эвтрофных озерах со средней глубиной 2–3 м и более, а также в перемешиваемом эпилимнионе стратифицированных озер; Тв – обитатели лотических экосистем (реки, ручьи); N – виды, обитающие в мелководных мезотрофных озерах со средней глубиной 2–3 м и более, а также в перемешиваемом эпилимнионе стратифицированных озер, чувствительные к повышению рН; МР – обитатели перемешиваемых водоемов с высоким содержанием минеральной взвеси; Y – фитофлагеляты, обитатели мелководных богатых биогенами водоемов, толерантные к световому лимитированию, способные жить в разнообразных лентических экосистемах при низком прессе хищников; X2 – жгутиковые, обитатели мезо-эвтрофных водоемов, толерантные к стратификации, чувствительные к перемешиванию и выеданию фильтраторами; G – зеленые жгутиковые, обитающие в стратифицированных эвтрофных небольших озерах и крупных водохранилищах в период стагнации; J – хлорококковые зеленые водоросли, обитатели мелководных богатых питательными веществами водоемов, чувствительные к световому лимитированию; F – колониальные зеленые водоросли, обитатели глубоко перешиваемых мезотрофных вод с высокой прозрачностью, чувствительные к дефициту CO2 и толерантные к содержанию биогенов и высокой мутности)
| Код | F | В | М | Ш | Г |
|---|---|---|---|---|---|
| LО | 3 | 0 | 0 | 0 | 1 |
| H2 | 3 | 0 | 0 | 1 | 0 |
| M | 0 | 1 | 1 | 1 | 1 |
| H1 | 1 | 1 | 1 | 1 | 1 |
| E | 2 | 1 | 0 | 0 | 1 |
| B | 2 | 1 | 1 | 1 | 1 |
| D | 4 | 1 | 1 | 1 | 1 |
| C | 4 | 1 | 0 | 1 | 1 |
| P | 5 | 1 | 1 | 1 | 1 |
| TВ | 5 | 1 | 0 | 1 | 1 |
| N | 2 | 1 | 1 | 1 | 1 |
| MP | 5 | 1 | 0 | 0 | 1 |
| Y | 3 | 1 | 1 | 0 | 1 |
| X2 | 3.5 | 1 | 0 | 1 | 0 |
| G | 0 | 1 | 0 | 1 | 0 |
| J | 2 | 0 | 0 | 0 | 1 |
| F | 5 | 0 | 0 | 0 | 1 |
В весенний период в составе фитопланктона водохранилища преобладали криптофитовые и диатомовые водоросли. В Волжском плесе преимущественно вегетировали представители групп: D (Cyclostephanos invisitatus и Stephanodiscus hantzschii Grun.), B (S. minutulus (Kütz.) Cleve & Möll. и S. binderanus) и Y (Komma caudata (L.Geitler) D.R.A. Hill, Cryptomonas ovata Ehr. и C. curvata Ehr.). Наряду с ними лидировала Aulacoseira subarctica (O. Müll.) Haworth (код В), основной доминант Моложского плеса. В Главном и Шекснинском плесах преимущественно развивались виды из группы В (Aulacoseira islandica (O. Müll.) Sim. и Stephanodiscus binderanus).
На спаде развития весеннего комплекса фитопланктона в Волжском плесе состав доминантов практически не менялся, увеличивалась лишь доля (%) биомассы видов из функциональной группы В. В центральной части водоема основную биомассу создавали диатомовые из группы: C (Asterionella formosa Hass.), P (Fragilaria crotonensis Kitt.) и B (Stephanodiscus neoastraea (Håk. & Hickel) emend. Casper, Scheffler et Augsten). В Моложском плесе господствовали виды из группы P – Aulacoseira granulata (Ehr.) Sim. и A. subarctica. Состав доминирующего комплекса водорослей Шекснинского плеса был более разнообразным. Наряду с диатомовыми Tabellaria fenestrata (Lyngb.) Kütz. (код N) и видами из родов Aulacoseira Thw. и Stephanodiscus Ehr. в его верхнем участке развивались зеленые водоросли из групп Х2 (Chlamydomonas reinhardtii P.A. Dang.) и G (Pandorina morum (O.F. Müll.) Bory de Saint-Vincent). С 2004 г. Actinocyclus normanii (Greg.) Hust. (код D), ранее входивший в число ведущих видов только в Шекснинском плесе, отмечен в Главном и нижних участках Волжского и Моложского плесов.
В летний период и до конца осени по всей акватории водохранилища активно вегетировали цианобактерии из групп H1 (Aphanizomenon flos-aquae Ralfs ex Bornet & Flahault, виды из рода Dolichospermum (Ralfs ex Bornet & Flahault) P. Wacklin, L. Hoffmann & Kom.) и M (Microcystis aeruginosa (Kütz.) Kütz., M. wesenbergii (Kom.) Kom. in Kondr.). Безгетероцистные цианобактерии из групп LO (из рода Gloeocapsa Kütz.), Н2 (из рода Gloeotrichia Ag.) и М (из рода Microcystis (Kütz.) Elenk.) преимущественно развивались в восточной части водоема (от Шекснинского плеса до плотины Рыбинской ГЭС (рис. 1)), диазотрофы из рода Dolichospermum (группа Н1) – в западной части, начиная с Моложского плеса. В Волжском плесе вклад группы Н1 (Aphanizomenon flos-aquae) в общую биомассу фитопланктона был наименьшим при их преимущественной вегетации в нижней расширенной части, прилегающей к акватории Главного плеса. На данном участке отмечалась высокая представленность фитофлагеллят из групп Y (динофлагеляты из рода Glenodinium Ehr.) и X2 (зеленые из рода Chlamydomonas Ehr.).
В период позднеe летo – осень на всей акватории водоема совместно с цианобактериями из групп H1 и М продолжали развиваться диатомовые из групп Р, В и D (Aulacoseira granulata, A. subarctica, Skeletonema subsalsum (Cleve-Euler) Bethge, Stephanodiscus binderanus, S. hantzschii, S. neoastraea). В конце сентября – начале октября в состав доминирующего комплекса входила Aulacoseira islandica.
Таким образом, доминирующий состав фитопланктона Рыбинского водохранилища был в основном представлен шестью функциональными группами: В, Н1, P, D, M и С. Относительный вклад этих групп в общую биомассу фитопланктона значительно варьировал: В – 31 ± 4, Н1 – 20 ± 3, Р – 13 ± 3, D – 9 ± 3, М – 10 ± 3, С – 4 ± 1%. Биомассы доминантов из других категорий составляли ≤1% каждая.
В Волжском плесе развивались преимущественно водоросли из групп В, D, Н1; в Моложском – В, Р, Н1; в Шекснинском – В, М, H1, D, P; в Главном – Н1, В, М, С, Р (табл. 2).
Таблица 2.
Относительный вклад, %, основных функциональных групп в общую биомассу фитопланктона в различных плесах Рыбинского водохранилища в 2000–2005 гг.
| Функциональные группы | В | М | Ш | Г |
|---|---|---|---|---|
| M | 7 ± 2 | 6 ± 3 | 14 ± 3 | 16 ± 2 |
| H1 | 14 ± 4 | 15 ± 5 | 11 ± 2 | 25 ± 3 |
| D | 16 ± 3 | 7 ± 2 | 12 ± 3 | 5 ± 1 |
| C | 4 ± 1 | 3 ± 1 | 9 ± 2 | 11 ± 2 |
| B | 23 ± 4 | 34 ± 7 | 27 ± 3 | 19 ± 3 |
| P | 8 ± 2 | 19 ± 5 | 10 ± 2 | 10 ± 1 |
| Прочие | 28 ± 4 | 16 ± 5 | 17 ± 2 | 14 ± 1 |
Индекс Q варьировал от 0.4 до 4.3. Его среднемесячная величина постепенно снижалась от поздней весны (2.0 ± 0.0 – 2.9 ± 0.3) к лету–осени (1.3 ± 0.1 – 2.2 ± 0.8) на всех исследуемых участках (рис. 2).
Рис. 2.
Сезонная динамика среднего индекса сообществ Q различных плесов Рыбинского водохранилища в 2000–2005 гг.
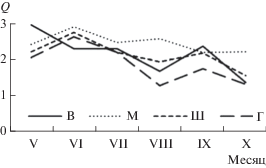
Наибольшая сезонная вариабельность Q прослеживалась в Главном плесе (коэффициент вариации 46%). Она снижалась в Шекснинском и Волжском плесах (37 и 40% соответственно) и принимала наименьшее значение в Моложском (23%). Максимальные средние значения Q получены в весенний период в Волжском плесе (2.9 ± 0.3) и минимальные – в августе–октябре в Главном плесе (1.3 ± 0.1).
Статистический анализ показал, что Q положительно коррелировал с вкладом (%) в общую численность и биомассу диатомовых водорослей и концентрацией общего азота, а отрицательно – с индексом сапробности, общей численностью и долей цианобактерий (табл. 3).
Таблица 3.
Статистические параметры связи индекса Q c отдельными биотическими и абиотическими показателями (R – коэффициент детерминации, F – критерий Фишера, p – уровень значимости)
| Показатель | Коэффициент корреляции Спирмена | Параметры линейной регрессии | ||
|---|---|---|---|---|
| R | F | p< | ||
| Общая численность, тыс. кл/л | –0.30 | –0.32 | 23.46 | 0.000 |
| Общая биомасса, г/м3 | –0.05* | –0.09* | 1.62 | 0.203 |
| Индекс сапробности | –0.17 | –0.18 | 6.84 | 0.009 |
| Численность цианобактерий, % | –0.66 | –0.54 | 82.44 | 0.0000 |
| Биомасса цианобактерий, % | –0.74 | –0.77 | 286.1 | 0.000 |
| Общий азот, мг/л | 0.35 | 0.40 | 7.84 | 0.008 |
| Численность диатомовых, % | 0.76 | 0.61 | 116.7 | 0.000 |
| Биомасса диатомовых, % | 0.78 | 0.78 | 314.3 | 0.000 |
В среднем за исследованный период наибольшее значение Q получено для вод Моложского плеса (2.5 ± 0.1), что означает их “умеренное качество”. Значение Q снижалось в Шекснинском (2.2 ± 0.2) и Волжском (2.1 ± 0.1) плесах и достигало наименьших величин в Главном (1.8 ± 0.1), что свидетельствовало о низком качестве его вод.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Проведенный анализ показал, что выявленные доминирующие виды фитопланктона, принадлежащие к различным группам (кодам) в соответствии с разработанной морфофункциональной классификацией [29], чаще присутствуют на различных участках водохранилища совместно. Большая их часть встречалась и до строительства плотины Рыбинской ГЭС в реках Волге, Шексне и Мологе [2, 3]. После создания водохранилища доминирующий комплекс стал пополняться новыми видами.
C 1954 г. в группе В, представленной таксонами рода Aulacoseira, регистрируются водоросли из рода Stephanodiscus (Stephanodiscus neoastraea, с 1978 г. – S. minutulus) и с 2000 г. – Lindavia radiosa (Grun.) De Toni and Forti [4, 8]. В 1980-х гг. в состав доминирующего комплекса включена Cyclotella meneginiana Kütz. (группа С) [12]. В 1960-е гг. группу D дополнили Stephanodiscus hantzschii и Skeletonema subsalsum и в 1980–1990-е гг. – Actinocyclus normanii [4, 8]. Виды из родов Peridiniopsis Lemm. и Cryptomonas Ehr. (группа Y) отмечались в числе ведущих с 1950-х гг. [4]. В 1970-е гг. к ним присоедились Komma caudata и Cryptomonas ovata, в 1980-е гг. – Cryptomonas marssonii Skuja, Peridiniopsis quadridens (Stein) Bour., в 1990-е гг. – P. penardii (Lemm.) Bour., в 2000 г. – P. kevei и Cryptomonas curvata [4, 8, 12]. Зеленые – Carteria multifilis (Fres.) O. Dill и Chlamydomonas reinhardii, а также криптофитовые – Rhodomonas lens Pascher (групп Х2) вошли в доминирующий комплекс фитопланктона в период настоящих исследований.
Состав ведущих видов из других функциональных групп с течением времени практически остался неизменным. Наиболее часто существенный вклад в общую биомассу вносили диатомовые (Aulacoseira granulata, Diatoma tenuis Ag. и Fragilaria crotonensis) из группы P и цианобактерии из ассоциаций H1 (диазотрофы Dolichospermum planctonicum (Brunnth.) Wacklin et al., D. flos-aquae (Lyngb.) Wacklin et al., D. spiroides (Klebahn) Wacklin et al. и Aphanizomenon flos-aquae) и М (Microcystis aeruginosa, M. viridis (A. Br. in Rabenh.) Lemm. и M. wesenbergii). До создания Рыбинского водохранилища цианобактерии преимущественно вегетировали в озерах междуречья и на подпертых участках устьев рек [3]. Золотистые водоросли из родов Dinobryon Ehr. и Mallomonas Perty из группы Е регистрировались в составе фитопланктона до заполнения водохранилища [2, 3]. В период настоящих исследований только Dinobryon divergens O.E. Imhof был выявлен в числе ведущих в Волжском и Главном плесах. В последнем фиксировались таксоны из ассоциаций LO, J и F. Доминанты из группы LO наиболее часто отмечались в 1950–1960-е гг. [4, 8, 15] и в основном были представлены, как и в настоящих наблюдениях, жгутиковыми крупноклеточными видами из рода Peridinium Ehr. и цианобактериями из порядка Chroococcales. В группе J, сформированной из зеленых водорослей, образующих ценобии, преобладал Pediastrum boryanum (Turp.) Menegh. [8, 12]. Колониальные зеленые из группы F разнообразно были представлены в начале 1960-х гг., а c середины 1990-х гг. преимущественно развивался Sphaerocystis planctonica (Korsh.) Bourr. [8].
В числе ведущих видов на акватории водохранилища не зарегистрированы: Aphanothece clatrata West & G.S. West (группа К) и Planktolyngbya limnetica (Lemm.) Kom.-Legn. et Cronb. (группа S1), Ceratium hirundinella (O.F. Müll.) Dujardin (группа Lm), Trachelomonas volvocina (Ehr.) Ehr. (группа W2), превалирующие в конце 1950-х – начале 1960-х гг. и в 1980-е гг., и Schroederia setigera (Schröd.) Lemm. (группа Х3) – в конце 1990-х гг. [4, 8]. Количество доминантов в группе Тв и МР уменьшилось.
Выявленные с учетом предшествующих результатов [6] закономерности сезонной сукцессии развития фитопланктона показали, что в весенний период преобладали виды, свойственные мезотрофным (группа В, Моложский плес), мезо-эвтрофным (группы В и С, Шекснинский и Главный плесы) и эвтрофным (группа D, Волжский плес) водоемам. Летом основную биомассу фитопланктона водохранилища создавали ассоциации, предпочитающие эвтрофные воды, – D, Р, M и H1. Осенью совместно с диатомовыми из групп В (Моложский и Главный плесы) и Р (Волжский и Шекснинский плесы) продолжали вегетацию цианобактерии из групп Н1 и М по всей акватории водохранилища. Таким образом, разнообразие альгоценозов с общими абиотическими предпочтениями увеличивалось от весны к осени. Это хорошо согласуется с физико-химическими данными. Наибольшие различия между участками Рыбинского водохранилища по химическому составу и физическим свойствам водных масс наблюдаются весной, а летом и осенью они незначительны [17].
В целом, согласно морфофункциональной классификации [29], в 2000-е гг. в Рыбинском водохранилище развивались преимущественно водоросли, характерные для мелководных мезотрофных и эвтрофных вод. Это виды, толерантные к световому лимитированию (функциональные группы B, C, P, Y, G, J), дефициту углерода (С, P, H1), биогенных веществ (N, E, F, H1, Н2, Lo), чувствительные к перемешиванию (X2, M, H1, H2, Lo) и стратификации (B, C, N, P), реагирующие на повышение рН (B, N), дефицит кремния (группы B, C, P), биогенных веществ (D, G) и углерода (E, F).
Качество воды исследованных участков Рыбинского водохранилища (табл. 4) по индексу сапробности и общей биомассе фитопланктона в среднем соответствовало 3-му классу, удовлетворительной чистоте [14], а по индексу Q – 3-му классу, умеренному загрязнению [25], что вполне сопоставимо между ними. Результаты статистического анализа показали, что состояние экосистемы Рыбинского водохранилища по Q ухудшалось по мере увеличения численности и вклада (доли) цианобактерий, а также уровня органического загрязнения (сапробности). Степень связи Q с индексом сапробности была самая низкая (табл. 3), поскольку сапробность выражает только уровень органического загрязнения, а индекс сообществ – общее состояние вод. При этом улучшение качества воды сопровождалось увеличением концентрации общего азота. К концу 1970-х гг. среди гидробиологов сформировалось устойчивое мнение о прямой связи эвтрофирования и ухудшения качества воды с увеличением концентрации фосфора в озерах умеренных широт [30, 31, 35, 36]. В последние годы разрабатывается новая парадигма N- и P-лимитирования в озерах [21, 22]. Исходя из соотношения А. Редфилда [28], в эвтрофных и гипертрофных водах соотношение N : P низкое, что благоприятно и для роста цианобактерий [33, 34]. В олиго-мезотрофных озерах число N : P увеличивается. Экспериментально показано, что его увеличение благоприятно для развития зеленых водорослей [10].
Таблица 4.
Изменение индекса сообществ Q, сапробности S, общей биомассы фитопланктона Вобщ, г/м3, доли биомассы цианобактерий Вциано, %, диатомовых водорослей Вдиат, %, в различных плесах Рыбинского водохранилища в 2000–2005 гг. (n – число проб)
| Плес | Q | S | Вобщ | Вциано | Вдиат | n |
|---|---|---|---|---|---|---|
| Моложский | 2.5 ± 0.1 | 2.08 ± 0.07 | 3.6 ± 0.9 | 21 ± 5 | 66 ± 6 | 17 |
| Шекснинский | 2.2 ± 0.2 | 1.97 ± 0.03 | 2.2 ± 0.3 | 26 ± 5 | 64 ± 5 | 31 |
| Волжский | 2.1 ± 0.1 | 2.07 ± 0.04 | 1.8 ± 0.3 | 20 ± 5 | 64 ± 5 | 37 |
| Главный | 1.8 ± 0.1 | 1.92 ± 0.04 | 2.1 ± 0.2 | 40 ± 3 | 52 ± 3 | 118 |
| Водохранилище | 2.5 ± 0.1 | 1.98 ± 0.01 | 2.3 ± 0.1 | 33 ± 2 | 57 ± 2 | 203 |
Из этого следует, что азота больше в низкопродуктивных водах, а поскольку индекс сообществ увеличивается при хорошем состоянии вод, то положительная связь Q с концентрацией общего азота вполне правомерна.
ВЫВОДЫ
В начале 2000-х гг. в фитопланктоне Рыбинского водохранилища развивались преимущественно ассоциации, чувствительные к перемешиванию (группы M, H1) и стратификации (B, C, P), толерантные к дефициту света (B, C, P, Y), углерода (С, P, H1), предпочитающие достаточное содержание кремния (B, C, P) и других биогенных элементов (D). Разнообразие функциональных групп, обладающих общими абиотическими требованиями, возрастало от весны к осени. Индекс сообществ Q менялся от 0.4 до 4.3, а его среднемесячная величина постепенно понижалась от поздней весны к осени. Снижение его значений, что свидетельствовало об ухудшении качества вод Рыбинского водохранилища, прослеживалось по мере увеличения суммарной численности фитопланктона, доли численности и биомассы цианобактерий и сапробности. Анализ качества вод по индексу сообществ, биомассе фитопланктона и индексу сапробности дал сходные результаты, однако для получения адекватной оценки качества вод необходимо использовать комплекс различных показателей.
Авторы выражают искреннюю благодарность команде экспедиционного флота ИБВВ РАН и Т.П. Зайкиной за помощь в сборе материала.
Список литературы
Волга и ее жизнь / Под ред. Н.В. Буторина, Ф.Д. Мордухай-Болтовского. Л.: Наука, 1978. 348 с.
Грезе Б.С. О планктоне Шексны, Мологи и Сити // Тр. Ярослав. естеств.-ист. и краевед. общ-ва. 1928. Т. IV. Вып. 2. С. 11–17.
Есырева В.И. Флора водорослей р. Волги от Рыбинска до г. Горького // Тр. Ботан. сада МГУ. 1945. Вып. 82. С. 10–90.
Корнева Л.Г. Сукцессия фитопланктона // Экология фитопланктона Рыбинского водохранилища. Тольятти, 1999. С. 89–148.
Корнева Л.Г. Фитопланктон Рыбинского водохранилища: состав, особенности распределения, последствия эвтрофирования // Современное состояние экосистемы Рыбинского водохранилища. СПб.: Гидрометеоиздат, 1993. С. 50–113.
Корнева Л.Г., Соловьева В.В. Опыт использования морфофункциональной классификации пресноводных водорослей для оценки динамики и пространственного распределения ассоциаций фитопланктона Рыбинского водохранилища // Ярославский педагогич. вестн. 2012. Т. 3. № 3. С. 110–114.
Корнева Л.Г., Соловьева В.В., Макарова О.С. Разнообразие и динамика планктонных альгоценозов водохранилищ Верхней и Средней Волги (Рыбинское, Горьковское, Чебоксарское) в условиях эвтрофирования и изменения климата // Экология, морфология и систематика водных растений. Ярославль: Филигрань, 2016. С. 35–45.
Корнева Л.Г., Соловьева В.В., Митропольская И.В. и др. Сообщества фитопланктона водохранилищ Верхней Волги // Экологические проблемы Верхней Волги. Ярославль: ЯГТУ, 2001. С. 87–93.
Кузьмин Г.В., Охапкин А.Г., Ильинский А.Л. Фитопланктон как индикатор сапробности вод Главного плеса Рыбинского водохранилища // Биология низших организмов. Тр. ИБВВ РАН. 1978. Вып. 40 (43). С. 36–52.
Левич А.П., Булгаков Н.Г. О возможности регулирования видовой структуры лабораторного альгоценоза // Изв. РАН. Сер. биол. 1993. № 4. С. 569–578.
Методика изучения биогеоценозов внутренних водоемов / Под. ред. Ф.Д. Мордухай-Болтовского. М.: Наука, 1975. 240 с.
Митропольская И.В. Структура и динамика фитопланктона Рыбинского водохранилища. Автореф. дис. … канд. биол. наук. М: МГУ, 2010. 27 с.
Мэгарран Э. Экологическое разнообразие и его измерение. М.: Мир, 1992. 184 с.
Оксиюк О.П., Жукинский В.Н., Брагинский Л.П. и др. Комплексная экологическая классификация качества поверхностных вод суши // Гидробиол. журн. 1993. Т. 29. № 4. С. 62–77.
Рыбинское водохранилище и его жизнь. Л.: Наука, 1972. 364 с.
Соловьева В.В., Корнева Л.Г. Современная характеристика сапробности Рыбинского водохранилища по фитопланктону // Вода: химия и экология. 2012. № 5. С. 18–23.
Степанова И.Э., Бикбулатов Э.С., Бикбулатова Е.М. Закономерности динамики содержания биогенных элементов в водах Рыбинского водохранилища за годы его существования // Вода: химия и экология. 2013. № 1. С. 15–28.
Фортунатов М.А. Цветность и прозрачность вод Рыбинского водохранилища как показатель его режима // Тр. Ин-та биол. водохранилищ. М.; Л.: АН СССР, 1959. Вып. 2 (5). С. 246–357.
Devercelli M. Changes in phytoplankton morpho-functional groups induced by extreme hydroclimatic events in the Middle Paraná River (Argentina) // Hydrobiologia. 2010. V. 639. P. 5–19.
Directive of the European Parliament and of the Council 2000/60EC Establishing a Framework for Community Action in the Field of Water Policy. European Commission PE-CONS 3639/1/100. Rev. 1. Luxembourg: EC Parliament and Council, 2000.
Guildford S.J., Hecky R.E. Total nitrogen, total phosphorus, and nutrient limitation in lakes and oceans: Is there a common relationship? // Limnol. Oceanogr. 2000. V. 45 (6). P. 1213–1223.
James J. Elser, Matthew E.S. Bracken, Elsa E. Cleland, Daniel S. Gruner, W. Stanley Harpole, Helmut Hillebrand, Jacqueline T. Ngai, Eric W. Seabloom, Jonathan B. Shurin, Jennifer E. Smith. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems // Ecol. Letters. 2007. V. 10 P. 1135–1142.
Kruk C., Mazzeo N., Lacerot G., Reynolds C.S. Classification schemes for phytoplankton: A local validation of a functional approach to the analysis of species temporal replacement // J. Plankton Res. 2002. V. 24. № 9. P. 901–912.
Marchetto A., Mariani M.A., Luglie A., Sechi N. A numerical index for evaluating phytoplankton response to changes in nutrient levels in deep mediterranean reservoirs // J. Limnol. 2009. V. 68. № 1. P. 106–121.
Padisák J., Borics G., Grigorszky I., Soróczki-Pintér É. Use of phytoplankton assemblages for monitoring ecological status of lakes within the Water Framework Directive: the assemblage index // Hydrobiologia. 2006. V. 553. P. 1–14.
Padisák J., Crossetti L.O., Naselli-Flores L. Use and misuse in the application of the phytoplankton functional classification: a critical review with updates // Hydrobiologia. 2009. V. 621. P. 1–19.
Piirsoo K., Pall P., Tuvikene A., Viik M., Vilbaste S. Assessment of water quality in a large lowland river (Narva, Estonia/Russia) using a new Hungarian potamoplanktic method // Estonian J. Ecol. 2010. V. 59. № 4. P. 243–258.
Redfield A.C. On the proportion of organic derivatives in sea water and their relation to the composition of plankton // James Johnstone Memorial Volume. Liverpool: Univ. Press of Liverpool, 1934. P. 176–192.
Reynolds C.S., Huszar V., Kruk C., Naselli-Flores L., Melo S. Towards a functional classification of the freshwater phytoplankton // J. Plankton Res. 2002. V. 24. № 5. P. 417–428.
Schindler D.W. Eutrophication and recovery in experimental lakes: Implications for lake management // Sci. 1974. V. 184. № 4139. P. 897–899.
Schindler D.W., Fee E.J., Ruszcynski T. Phosphorus input and its consequences for phytoplankton standing crop and production in the Experimental Lakes Area and in similar lakes // J. Fish. Res. Board Can. 1978. V. 35. P. 190–196.
Sládeček V. System of Water Quality from the Biological Point of View // Arch. Hydrobiol. 1973. Beih. 7. Ergebnisse der Limnologie. Heft 7. P. 1–218.
Smith V.H. Low N to P favor dominance by blue-green algae in lake phytoplankton // Sci. 1983. V. 221. P. 669–671.
Suttle C., Cochlan W.P., Stockner J.G. Size-dependent ammonium and phosphate uptake, and N : P supply ratios in an oligotrophic lake // Can. J. Fish. and Aquat. Sci. 1991. V. 48. P. 1226–1234.
Vollenweider R.A. Advances in defining critical loading levels of phosphorus in lake eutrophication // Mem. Inst. Ital. Idrobiol. 1976. V. 33. P. 53–83.
Vollenweider R.A. Input-output models // Schweiz. Z. Hydrol. 1975. V. 37. P. 53–84.
Wegl R. Index für die Limnosaprobität // Wasser und Abwasser. 1983. Band 26. 175 p.
Дополнительные материалы отсутствуют.