

Водные ресурсы, 2022, T. 49, № 1, стр. 103-111
Комплексный анализ распределения компонентов экосистемы в Черном море с учетом гидрохимических и гидрометеорологических факторов
С. Б. Крашенинникова a, *, Н. И. Минкина a, И. Г. Шокурова b, Э. З. Самышев a
a ФИЦ Институт биологии южных морей им. А.О. Ковалевского РАН
299011 Севастополь, Россия
b ФИЦ Морской гидрофизический институт РАН
299011 Севастополь, Россия
* E-mail: svetlanabk@mail.ru
Поступила в редакцию 27.01.2021
После доработки 29.06.2021
Принята к публикации 29.06.2021
- EDN: PQZVSB
- DOI: 10.31857/S0321059622010096
Аннотация
Проведен анализ распределения компонентов экосистемы (фитопланктона, ракообразного эпи-, батипланктона и желетелого планктона) с учетом гидрохимических и гидрометеорологических условий на основе данных комплексных съемок в Черном море. Учитывались следующие факторы среды: гидрохимические – биогенные элементы (нитраты, нитриты, фосфаты, кремний) как лимитирующие факторы роста биомассы фитопланктона и гидрометеорологические (скорость и направление ветра, экмановская накачка, апвеллинги, плотностные фронты), влияющие на распределение и концентрацию биогенов. Получено, что пространственное распределение биогенных элементов и фитопланктона хорошо согласуется с сезонными изменениями вертикального подъема вод – экмановской накачки, связанной с неоднородностью поля ветра над морем. Анализ межгодовой изменчивости экмановской накачки за период 1979–2016 гг. показал, что зимой над всей глубоководной частью моря преобладает циклоническая завихренность в поле ветра, сопровождающаяся вертикальным подъемом вод, что создает благоприятные условия для обогащения вод верхнего слоя моря биогенными элементами. В летний сезон циклоническая завихренность ветра в основном преобладает над восточной частью моря, внося вклад в подъем глубинных вод. Над западной частью моря летом преобладает в среднем антициклоническая завихренность, препятствуя подъему глубинных вод.
ВВЕДЕНИЕ
При исследовании функционирования экосистемы Черного моря актуально изучение пространственно-временнóй изменчивости компонентов экосистемы и их трофических связей в верхнем фотическом слое [5, 10, 17, 32]. Кроме того, остается важным вопрос о зависимости численности и биомассы компонентов экосистемы от условий их среды обитания. Известно, что биомасса фито- и зоопланктона меняется по сезонам и неоднородно распределена по акватории Черного моря, что связано с действием на экосистему различных факторов [16, 18, 22, 32]. К числу таких факторов относятся: гидрологические (температура, соленость, градиенты плотности морской воды), гидрометеорологические (потоки тепла на поверхности, скорость ветра, экмановская накачка, прибрежный апвеллинг), гидрохимические (биогены, поллютанты), оптические (освещенность). Кроме того, трофические связи внутри экосистемы и сезонные вариации гидрометеорологических условий приводят к изменениям не только численности, но и видов фитопланктона в течение года [1, 21].
Анализ согласованности пространственно-временнóй изменчивости компонентов экосистемы и факторов среды – наиболее информативный способ выявления связей между ними. В настоящее время имеются непрерывные ряды данных, позволяющие анализировать изменчивость гидрометеорологических факторов. Однако анализ изменчивости компонентов экосистемы за счет влияния факторов среды затруднен из-за отсутствия регулярных данных о численности и биомассе фито- и зоопланктона, а также о концентрации биогенных элементов [12, 16, 18, 32]. Поэтому для таких целей часто используют численные модели [6, 26]. В последнее время изменчивость отдельных компонентов экосистемы также изучают на основе спутниковых наблюдений, имеющих регулярный характер [11, 13]. Тем не менее натурные данные – основной индикатор состояния экосистемы моря. Анализ межгодовой изменчивости компонент экосистемы по натурным наблюдениям показывает важность влияния на нее климатических факторов, метеоусловий и концентрации биогенных элементов [18, 22]. В большинстве ранних исследований рассматривается влияние единичных факторов на изменчивость отдельных компонентов экосистемы. В настоящей работе ставится задача учета комплекса факторов, одновременно влияющих на работу нескольких компонентов экосистемы. Выполненные в разные сезоны 1992 г. уникальные комплексные съемки гидрологических, оптических, биологических и гидрохимических характеристик, включая съемку CoMSBlack'92 [4, 24], а также данные атмосферного реанализа [19] позволяют учесть наибольшее число факторов и наиболее полно проанализировать их влияние на распределение основных компонентов экосистемы.
Цель работы – анализ влияния гидрохимических и гидрометеорологических факторов среды на распределение основных компонентов экосистемы (фито- и зоопланктона) в Черном море.
МАТЕРИАЛЫ И МЕТОДЫ
В работе использовались следующие данные:
биомасса фитопланктона, ракообразного эпи-, батипланктона и желетелых организмов (мг/м3); концентрация биогенных элементов – нитратов (мг/л), нитритов, кремния, общего фосфора (мкг/л); температура (°С) и соленость (psu), полученные в рейсе № 60 научно-исследовательского судна погоды (НИСП) “Г. Ушаков” в марте 1992 г. и рейсах № 57 и 61 научно-исследовательского судна (НИС) “Э. Кренкель” в мае и июле 1992 г. [5]; все съемки выполнены по единой схеме станций во всей северной половине Черного моря;
температура (°С) и соленость (psu) из банка данных Морского гидрофизического института РАН (ФИЦ МГИ РАН) [3];
скорость ветра на высоте 10 м с дискретностью 6 ч по реанализу ERA-Interim (0.75° × 0.75°, 6 ч) [19].
Кроме того, для сравнения и анализа полученных результатов привлекались оценки, представленные в работах [1, 8, 12, 18, 20, 23, 24, 25].
Для анализа изменчивости пространственно-временнóго распределения компонентов экосистемы предварительно рассчитывалась средняя концентрация биомассы фито- и зоопланктона и биогенов в четырех слоях (поверхностном микрослое, верхнем перемешанном слое, слое термоклина и ниже термоклина до 50 м для фитопланктона и до 100 м для зоопланктона) по методике, изложенной в работе [23]. С учетом суточных вертикальных перемещений фито-, мезозоопланктона и желетелого планктона рассматривалась также средняя концентрация биомассы на каждой станции в слое от поверхности до верхней границы галоклина. Для концентрации биогенов также находились средневзвешенные величины.
Анализировались следующие гидрометеорологические факторы, оказывающие влияние на распределение биогенных элементов: экмановская вертикальная скорость, возникающая в результате неоднородности ветрового поля (завихренности), прибрежный апвеллинг, скорость и направление ветра и плотностные фронты в районах стока рек.
В открытом море распределение биогенных элементов анализировалось на основе его сравнения с распределением экмановской вертикальной скорости (экмановской накачки) на нижней границе экмановского слоя. Вертикальная экмановская скорость вычислялась на основе данных о скорости ветра по формуле [31]:
Изменение концентрации биогенных элементов у берегов анализировалось на примере района моря у Южного берега Крыма (ЮБК) в сравнении с изменениями повторяемости ветров, вызывающих апвеллинг. Наиболее вероятные условия возникновения апвеллинга в прибрежье ЮБК – западные и юго-западные ветры [9]. Для ветров указанных направлений вычислялась среднемесячная повторяемость (%) по отношению к общему числу ситуаций с разным направлением ветра. Направление ветра определялось как направление вектора скорости, полученного осреднением компонент скорости ветра (u, v) в рассматриваемом районе.
Плотностные фронты определялись путем вычисления горизонтальных градиентов плотности морской воды ${{\rho }_{0}}$ (кг/м4). Плотность рассчитывалась на основе уравнения ЮНЕСКО по данным о температуре и солености, предварительно интерполированных на регулярную сетку методом оптимальной интерполяции [2, 27].
РЕЗУЛЬТАТЫ
Компоненты экосистемы
На рис. 1 представлены пространственные распределения суммарной биомассы фитопланктона, ракообразного эпи-, батипланктона и желетелого планктона в марте, мае и июле 1992 г.
Рис. 1.
Пространственное распределение средней концентрации биомассы фитопланктона (а, б, в), ракообразных эпипланктона (г, д, е) и батипланктона (ж, з, и), желетелого планктона (к, л, м) в марте, мае и июле 1992 г. Величины биомассы выражены в мг/м3.
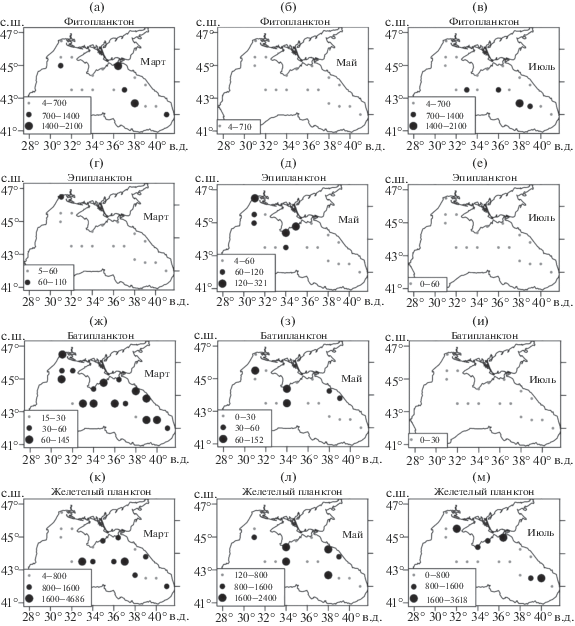
Большие значения биомассы фитопланктона отмечались в марте и июле, малые – в мае. Средняя по анализируемой области концентрация фитопланктона в мае была в 3 раза меньше, а в июле – в 1.5 раза меньше, чем в марте (рис. 1а–1в). Высокие значения биомассы микроводорослей в марте и июле в основном наблюдались в центральной и восточной частях моря. В течение года также менялось вертикальное распределение фитопланктона. Наибольшая биомасса фитопланктона была сконцентрирована в поверхностном слое в марте и мае, в подповерхностном слое (в сезонном термоклине) – в июле.
Суммарная биомасса эпи- и батипланктона уменьшалась от марта к июлю, однако их динамика имела свои особенности. В мае средняя биомасса эпипланктона была в 3 раза выше, чем в марте и июле (рис. 1г–1е). Эпипланктон весной в основном располагался в поверхностном слое моря, а в июле в подповерхностном. Средняя концентрация батипланктона (копепод) была в мае в 2 раза меньше, а в июле в 10 раз меньше, чем в марте.
Средняя концентрация желетелого планктона (преимущественно гребневиков Mnemiopsis leidyi A. Agassiz, 1865 и Pleurobrachia pileus O. F. Müller, 1776) в пелагиали практически не менялась в рассматриваемые месяцы (рис. 1к–1м). Однако она снижалась в поверхностном слое и возрастала в подповерхностном слое от марта к июлю. В марте и мае большие значения биомассы желетелого планктона наблюдались в центре Черного моря. В июле его концентрация увеличилась у берегов Крыма и в районе Батумского антициклонического круговорота, что связано с увеличением загрязнений антропогенного происхождения [28, 30].
Гидрохимические факторы
Пространственное распределение биогенных элементов имеет характер, аналогичный распределению фитопланктона. Большая концентрация нитритов и нитратов отмечалась в марте по всей исследуемой области как в пелагиали, так и в прибрежных районах. К маю она уменьшилась в 2 раза (рис. 2а–2д). В июле концентрации нитритов и нитратов увеличились по сравнению с маем, но в основном в глубоководной восточной части моря и у берегов Крыма.
Рис. 2.
Пространственное распределение биогенов в пелагиали северной части Черного моря: нитриты NO2 (а, б, в), мкг/л, нитраты NO3 (г, д, е), мгк/л, общий фосфор P, (ж, з, и), мкг, для марта, мая и июля 1992 г.
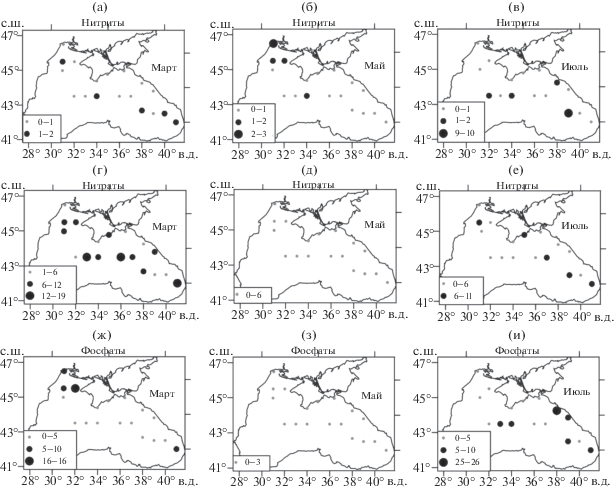
Концентрации фосфатов в среднем по акватории Черного моря в марте и в июле также оказались выше, чем в мае (рис. 2ж–2и). Большие значения отмечались на северо-западном шельфе в марте и в восточной части моря – в июле.
Гидрометеорологические факторы
Рост обилия фитопланктона – первого звена в пищевой цепи – лимитируется концентрацией биогенных элементов. Распределение биогенных элементов зависит от внешних факторов и их пространственно-временнóй неоднородности. К таким факторам относится ветровое воздействие, по-разному проявляющееся в центральных и прибрежных областях, циркуляция вод, сток рек и бытовых отходов, плотностная структура вод.
В центральной части моря основной источник биогенов – их поступление из глубинных вод, так как здесь их концентрация увеличивается с глубиной. Преобладающая в течение всего года циклоническая завихренность ветра над морем обусловливает вертикальный подъем вод и более высокое положение пикноклина в центральной части моря. Наиболее высокие вертикальные скорости подъема вод, обусловленные циклонической завихренностью ветра, отмечаются с января по март (рис. 3а). С этим, по-видимому, связаны высокие концентрации нитритов и нитратов в центральных районах, наблюдаемые в марте (рис. 2а, 2г), и, как следствие, большие величины биомассы фитопланктона (рис. 1а).
Рис. 3.
Сезонный ход экмановской вертикальной скорости (а), м/с, скорости ветра (б), м/с, повторяемости западных и юго-западных ветров, %, у Южного берега Крыма (в); пространственное распределение экмановской вертикальной скорости в марте, мае и июле 1992 г. (г, д, е) соответственно (по данным реанализа ERA-Interim 1979–2016 гг.). Точками на рисунках (г, д, е) обозначены районы, где вертикальная скорость превышала 10–5, м/с; Средние горизонтальные градиенты плотности в марте и мае, вычисленные по данным 1980–1992 гг. [3] (ж, з), и в июле по данным 1992 г. [3] (и).
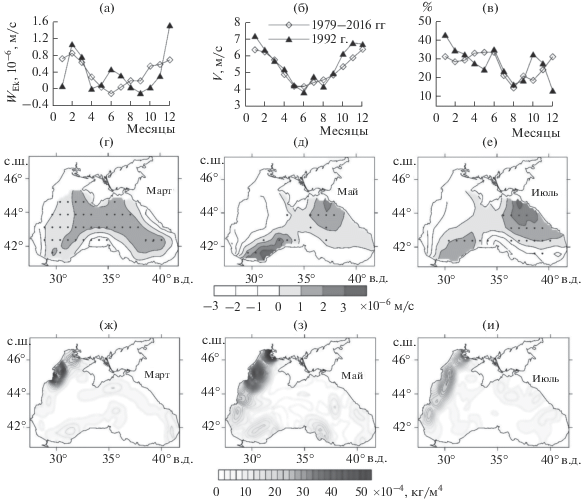
На рис. 3г–3е представлено пространственное распределение средней экмановской скорости для марта, мая и июля 1992 г. Точками обозначены локальные районы подъема вод, обусловленного синоптическими ситуациями с циклонической завихренностью ветра, которые сопровождаются положительной вертикальной скоростью >10–5 м/с. В марте наблюдались большие значения экмановской скорости во всей глубоководной части моря (рис. 3г), в мае и июле – в восточной части Черного моря и у берегов Турции (рис. 3д, 3е). Повышенные концентрации биогенов (нитритов, нитратов и фосфатов) в июле (рис. 2и, 2м) также локализовались в восточной части моря, где отмечены повышенные значения экмановской накачки (точки на рис. 3е). В июле 1992 г. средняя по акватории величина экмановской накачки была в 3 раза выше, чем в мае (рис. 3а). Этим обстоятельством могут объясняться более высокие концентрации нитратов в июле по сравнению с маем в юго-восточном районе (рис. 2з) и, как следствие, наибольшие значения биомассы фитопланктона (рис. 1в).
На рис. 4 приведены средние значения экмановской вертикальной скорости для января–марта и июня–августа для глубоководных районов западной и восточной частей моря в 1979–2016 гг. В январе–марте, когда над морем усиливается циклоническая завихренность ветра (рис. 3а), вертикальный подъем вод преобладает как в западной, так и в восточной частях моря. Отрицательные значения, свидетельствующие об опускании вод, наблюдаются лишь в некоторые годы в западной части моря. Летом, когда циклоническая завихренность ветра в среднем по морю ослабевает (рис. 3а), величина экмановской накачки для двух частей моря различается по знаку. В восточной части моря преобладает циклоническая завихренность ветра, поэтому экмановская скорость в основном имеет положительные значения, т. е. происходит интенсивный подъем вод. В западной части моря преобладает антициклоническая завихренность в поле ветра, поэтому значения экмановской вертикальной скорости в среднем отрицательные и направлены вниз, что ослабляет подъем вод, богатых биогенными элементами в этой части моря.
Рис. 4.
Экмановская вертикальная скорость, м/с, осредненная для восточной и западной глубоководных частей моря в январе–марте (а) и июне–августе (б).
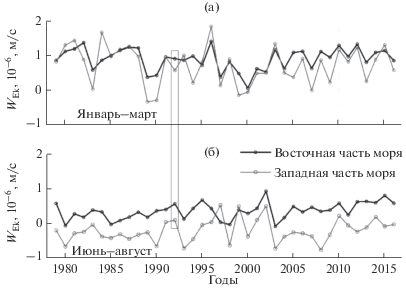
Таким образом, можно предположить, что летом в глубоководных районах восточной части моря значимый источник поступления биогенных элементов в фотический слой – экмановский подъем вод за счет циклонической завихренности ветра над морем.
Высокие концентрации биогенов в центральных районах моря в июле по сравнению с маем также могут быть связаны с большими скоростями ветра. В результате воздействия сильного ветра происходит механическое перемешивание вод, приводящее к обогащению поверхностных вод биогенными элементами [11]. В июле 1992 г. средняя скорость ветра превышала среднемноголетнюю величину за период 1979–2016 гг. и была выше майской (рис. 3б). Высокие значения скорости и завихренности ветра в июле 1992 г. также сопровождались усилением циркуляции вод в море [8, 23].
Концентрация биогенных элементов в прибрежных районах может меняться за счет апвеллингов, а также за счет сезонного изменения стока рек и бытовых стоков [5, 15]. У ЮБК апвеллинг возникает в основном при западных и юго-западных ветрах [9], отгоняющих поверхностные воды в море, взамен которых поднимаются глубинные воды. Расчет показывает, что в марте 1992 г. повторяемость ветров этих направлений составляла 32%, а в мае и июле была приблизительно одинаковой – 24 и 23% соответственно. Отметим, что в мае 1992 г. повторяемость этих ветров была намного меньше среднемноголетней величины за 1979–2016 гг. (рис. 4в). Высокие концентрации нитратов у ЮБК в марте и июле (рис. 2) могут быть связаны с прибрежным апвеллингом, способствующим увеличению концентрации биогенов в прибрежных водах. Высокие концентрации нитритов в мае в районе Северо-западного шельфа связаны, вероятно, с сезонным стоком рек.
Большие горизонтальные градиенты плотности в районах стока рек могут служить барьером для распространения биогенных элементов, поэтому во фронтальных зонах может содержаться повышенная их концентрация [29]. Поступление биогенных элементов через плотностные фронты происходит в основном за счет переноса мезомасштабными вихрями [13]. Плотностной фронт во все месяцы находится вдоль западного побережья моря с наибольшими значениями горизонтального градиента в районах стока рек Дунай и Днепр (рис. 3ж, 3з). В районе фронта в марте наблюдались повышенные концентрации нитритов и фосфора, в мае – нитритов (рис. 2).
ОБСУЖДЕНИЕ
Проведенный анализ пространственно-временнóй изменчивости компонент экосистемы на примере комплексных съемок в 1992 г. показывает влияние всех факторов – гидрометеорологических, гидрохимических и биотических – на продуктивность экосистемы.
Распределение и величина биомассы фитопланктона, по данным всех съемок, хорошо согласуются с изменчивостью биогенных элементов и величиной экмановской вертикальной скорости, возникающей в результате завихренности в поле ветра (рис. 1а–1в; 2; 3г–3е). Максимальные биомассa фитопланктона и концентрации биогенных элементов в фотическом слое отмечается в марте, когда экмановский подъем вод максимальный. В июле в восточной части моря наблюдались высокие концентрации фитопланктона и фосфора (рис. 1в, 2и). Согласно [1, 20], в июле 1992 г. в восточной части моря наблюдалось цветение кокколитофорид, биомасса которых лимитируется содержанием фосфора [14, 20]. Здесь же за счет циклонической завихренности ветра отмечены высокие значения вертикальной экмановской скорости, не уступающие значениям в марте (рис. 3г, 3е).
Согласно [18, 22], биомасса фитопланктона летом 1992 г. – наибольшая за современный период наблюдений с 1960-х гг. По хронологии, введенной в [12], 1992 г. относится к холодному периоду 1984–1996 гг., когда преобладали холодные зимы, за счет зимней конвекции и перемешивания увеличивался поток биогенных элементов из глубинных слоев моря и они накапливались их в верхнем слоя моря. Этот же год относят к периоду экологического кризиса 1989–1992 гг., последовавшего за периодом интенсивной эвтрофикации 1985‒ 1988 гг. за счет антропогенной нагрузки [18, 22]. С антропогенной эвтрофикацией вод связывают и низкую прозрачность морской воды, которая в 1992 г. была наименьшей за многолетний период наблюдений (с 1923 г.) – средняя прозрачность снизилась с 23 до 6.2 м [20, 25].
Сравнение величины экмановской вертикальной скорости в 1992 г. со значениями в другие годы показывает, что зимой указанного года вертикальная экмановская скорость в восточной части моря была незначительно выше среднего значения за 1979–2016 гг., а в западной части – ниже среднего значения (рис. 4а). Летом средняя вертикальная скорость в восточной (0.55 × 10–6 м/с) и в западной (0.08 × 10–6 м/с) частях моря значительно превышала среднемноголетние значения (0.33 × 10-6 м/с и –0.21 × 10-6 м/с соответственно) (рис. 4б). Таким образом, летом 1992 г. метеорологические условия сопровождались более сильным подъемом вод, чем в среднем за 1979–2016 гг., что может быть одной из причин высокой продуктивности в этот год. В июле максимумы биомассы фитопланктона и концентрации биогенных элементов были в термоклине и под ним. Сильная стратификация в термоклине препятствует проникновению биогенных элементов в верхние слои. Потребление фитопланктоном биогенов, поступающих из нижележащих слоев путем перемешивания, возможно, осуществляется за счет его заглубления.
Трофические связи в экосистеме также вносят вклад во временну́ю и пространственную изменчивость компонент экосистемы. По данным съемок, уменьшение биомассы фитопланктона в мае происходило на фоне больших величин биомассы ракообразного эпипланктона, а его увеличение в июле – при уменьшении суммарной биомассы эпи- и батипланктона. Несмотря на особенности 1992 г., изменение биомассы фитопланктона и зоопланктона от марта к июлю хорошо согласуется с современными представлениями об их сезонном ходе [1, 7, 21, 26].
ЗАКЛЮЧЕНИЕ
Черноморская пелагическая экосистема находится под воздействием большого числа факторов среды, что требует комплексного учета их влияния. В настоящей работе на примере данных комплексных съемок, выполненных в марте, мае и июле 1992 г., предпринята попытка комплексного подхода к учету факторов среды, влияющих на основные компоненты экосистемы, таких как фитопланктон, ракообразный эпи-, батипланктон и желетелые организмы. Проведено сопоставление гидрометеорологических факторов (экмановской накачки, скорости ветра, апвеллинга, гидрологических фронтов) и распределения концентраций биогенных элементов и биомассы фитопланктона.
Данные съемок показали, что распределения биогенных элементов и биомассы фитопланктона хорошо согласуются с пространственно-временнóй изменчивостью вертикальной экмановской скорости. Усиление экмановского подъема вод в марте сопровождалось высокой концентрацией биогенных элементов и, как следствие, высокими величинами биомассы фитопланктона. Высокие концентрации биогенных элементов в июле наблюдались в основном в центральной и восточной частях моря, где отмечены высокие значения экмановской вертикальной скорости.
По расчетам на основе данных атмосферного реанализа ERA-Interim, летом в восточной глубоководной части моря, в отличие от западной, преобладает подъем вод, обусловленный неоднородностью поля скорости ветра над морем, что может вносить значимый вклад в увеличение продуктивности в этой части моря. По расчетам на основе гидрологических данных, в западной части моря и на северо-западном шельфе находится плотностной фронт за счет стока рек Дунай и Днепр, в его районе наблюдается высокое содержание биогенных элементов.
Список литературы
Брянцева Ю.В. Оценка обилия фитопланктона в Черном море и его сезонная динамика в 1992 году // Тр. ЮгНИРО. 1996. Т. 42. С. 184–189.
Гандин Л.С. Объективный анализ метеорологических полей. Л.: Гидрометеоиздат, 1963. 287 с.
Годин Е.А., Белокопытов В.Н., Ингеров А.В. и др. Свид. о гос. регистрации базы данных № 2 019 621 008 “Черное море: гидрология – 2018”.
Грузов Л.Н., Люмкис П.В., Нападовский Г.В. Исследования пространственно-временнóй структуры планктонных полей северной половины Черного моря в 1992–93 гг. // Исследование экосистемы Черного моря. Одесса: Министерство охраны окружающей природной среды Украины, Укр. науч. центр экологии моря, 1994. Вып. 1. С. 94–127.
Дзицкий В.С., Минкина Н.И., Орлова И.Г. и др. Комплексная оценка загрязненности вод Черного моря в 90-е годы прошлого столетия // Системы контроля окружающей среды. Средства, модели и мониторинг. Севастополь: Морской гидрофиз. ин-т, 2011. Вып. 15. С. 265–273.
Дорофеев В.Л. Моделирование декадной изменчивости экосистемы Черного моря // Морской гидрофиз. журн. 2009. № 6. С. 71–81.
Дорофеев В.Л., Сухих Л.И. Изучение долговременной изменчивости экосистемы черного моря на основе ассимиляции данных дистанционных измерений в численной модели // Вод. ресурсы. 2019. Т. 46. № 1. С. 58–69.
Иванов В.А., Белокопытов В.Н. Океанография Черного моря. Севастополь, 2011. с. 212.
Иванов В.А., Михайлова Э.Н. Апвеллинг в Черном море. С.: ЭКОСИ-Гидрофизика, 2008. 92 с.
Крашенинникова С.Б., Минкина Н.И., Самышев Э.З. и др. Влияние комплекса факторов среды на биомассу фитопланктона и зоопланктона в Черном море в весенний период // Экология и строительство. 2019. № 4. С. 14–21.
Кубряков А.А., Микаэлян А.С., Станичный С.В. Вертикальная эволюция цветений кокколитофорид и ее влияние на растворенную органику в Черном море по данным буев БИО-АРГО // Современные методы и средства океанологических исследований (МСОИ-2019). 2019. С. 214–217.
Микаэлян А.С. Временнáя динамика фитопланктона глубоководного бассейна Черного моря. Дис. … докт. биол. наук. М.: ИО РАН, 2018. 266 с.
Микаэлян А.С., Зацепин А.Г., Кубряков А.А. Воздействие мезомасштабной вихревой динамики на биопродуктивность морских экосистем (обзор) // Морской гидрофиз. журн. 2020. Т. 36. № 6. С. 646–675.
Микаэлян А.С., Силкин В.А., Паутова Л.А. Развитие кокколитофорид в Черном море: межгодовые и многолетние изменения // Океанология. 2011. Т. 51. № 1. С. 45–53.
Совга Е.Е., Кондратьев С.И., Годин Е.А. и др. Сезонная динамика содержания и локальные источники биогенных элементов в водах прибрежной акватории Гераклейского полуострова // Морской гидрофиз. журн. 2017. № 1. С. 56–65.
Стельмах Л.В., Куфтаркова Е.А., Бабич И.И. Сезонная изменчивость скорости роста фитопланктона в прибрежных водах Черного моря (район Севастополя) // Морской экол. журн. 2009. Т. 8. № 1. С. 67–80.
Финенко Г.А., Романова З.А., Аболмасова Г.И., и др. Трофические взаимоотношения в планктонном сообществе Черного моря на современном этапе // Экология моря. 2006. Т. 71. С. 50–54.
Юнев О.А., Коновалов С.К., Великова В. Антропогенная эвтрофикация в пелагической зоне Черного моря: долговременные тренды, механизмы, последствия. М.: ГЕОС, 2019. 164 с. https://doi.org/10.34756/GEOS/2019.16.37827
Dee D.P., Uppala S.M., Simmons A.J. et al. The ERA-Interim Reanalysis: Configuration and Performance of the Data Assimilation System // Quarterly J. Royal Meteorol. Society. 2011. V. 137. № 656. P. 553–597.
Mankovsky V.I., Vladimirov V.L., Solovev M.V., Besiktepe S. Optical properties of the Black Sea: results of the CoMSBlack and TU-Black Sea Programs // NATO Sci. Ser. 2 Environ. security. 1998. V. 47. № 2. P. 145–162.
Mikaelyan A.S., Kubryakov A.A., Silkin V. et al. Regional climate and patterns of phytoplankton annual succession in the open waters of the Black Sea // Deep Sea Res. Pt I. Oceanogr. Res. Papers. 2018. V. 142. P. 44–57.
Mikaelyan A.S., Zatsepin A.G., Chasovnikov V.K. Long-term changes in nutrient supply of phytoplankton growth in the Black Sea // J. Mar. Sys. 2013. V. 117–118. P. 53–64.
Newhose Y. Daily euphotic zone primary productivity “Pionner” cruise of the international Indian Ocean expedition // Arch. Oceanogr. Limnol. 1968. V. 16. P. 1–43.
Oguz T., Besiktepe S., Basturk O. et al. CoMSBlack'92 Physical and Chemical Intercalibration Workshop // Workshop rep. № 98. Erdemli, Turkey: Unesco, 1993.
Oguz T., Dippner J.W., Kaymaz Z. Climatic regulation of the Black Sea hydro-meteorological and ecological properties at interannual-to-decadal time scales // J. Mar. Systems. 2006. V. 60. № 3–4. P. 235–254.
Oguz T., Ducklow H.W., Malanotte-Rizzoli P. Modeling distinct vertical biogeochemical structure of the Black Sea: Dynamical coupling of the oxic, suboxic, and anoxic layers // Global Biogeochem. Cycles. 2000. V. 14. № 4. P. 1331–1352.
Polonskii A.B., Shokurova I.G. Statistical structure of the large-scale fields of temperature and salinity in the Black Sea // Physical Oceanogr. 2008. V. 18. № 1. P. 38–51.
Purcell J.E., Uye S., Lo W.T. Anthropogenic causes of jellyfish blooms and their direct consequences for humans: a review // Mar. Ecol. Progress Ser. 2007. V. 350. P. 153–174.
Ragueneau O., Lancelot C., Egorov V. et al. Biogeochemical transformations of inorganic nutrients in the mixing zone between the Danube River and the north-western Black Sea // Estuarine, Coastal Shelf Sci. 2002. V. 54. № 3. P. 321–336.
Richardson A.J., Bakun A., Hays G.C. et al. The jellyfish joyride: causes, consequences and management responses to a more gelatinous future // Trends Ecol. Evolution. 2009. V. 24. № 6. P. 312–322.
Tomczak M., Godfrey J.S. Regional oceanography: an introduction. Delhi: Daya Publ. House, 2003. 401 p.
Yunev O.A., Moncheva S., Carstensen J. Long-term variability of vertical chlorophyll a and nitrate profiles in the open Black Sea: eutrophication and climate change // Mar. Ecol. Prog. Ser. 2005. V. 294. P. 95–107.
Дополнительные материалы отсутствуют.