

Водные ресурсы, 2022, T. 49, № 3, стр. 286-297
Изотопы радия и биогенные вещества в эстуарии реки Раздольной (Амурский залив, Японское море) в период летнего паводка
П. Ю. Семкин a, *, П. Я. Тищенко a, П. П. Тищенко a, Г. Ю. Павлова a, Е. В. Анисимова a, Ю. А. Барабанщиков a, Т. А. Михайлик b, А. Н. Чаркин a, М. Г. Швецова a
a Тихоокеанский океанологический институт им. В.И. Ильичева ДВО РАН
690041 Владивосток, Россия
b Санкт-Петербургский филиал ВНИРО (“ГосНИОРХ” им. Л.С. Берга)
199053 Санкт-Петербург, Россия
* E-mail: pahno@list.ru
Поступила в редакцию 19.08.2021
После доработки 19.08.2021
Принята к публикации 18.11.2021
- EDN: XREREH
- DOI: 10.31857/S0321059622030130
Аннотация
Изучена активность растворенных изотопов радия: 223Ra, 224Ra, 228Ra; концентрация биогенных веществ: аммония, нитратов, нитриты, фосфатов, силикатов, общего азота и общего фосфора в эстуарии р. Раздольной при содержании взвеси в реке >1000 мг/л. В паводок над устьевым взморьем наблюдалась классическая двухслойная эстуарная циркуляция, сформированная стоковым течением и компенсационным потоком с глубины 20 м до глубины 8 м. Основной процесс поступления 228Ra в эстуарий – десорбция из речной взвеси в начале зоны смешения, в районе устьевого бара, где его активность достигала 163 ± 0.03 dpm 100 л–1, что в 139 раз выше, чем в речной воде. В районе устьевого бара наблюдалось увеличение активности 224Ra и 223Ra (соответственно в 4 и 17 раз выше, чем в речной воде) и повышение концентрации общего фосфора, ${\text{NH}}_{{\text{4}}}^{ + }$, DSi, ${\text{NO}}_{{\text{3}}}^{ - }$ и общего азота. Экстремум активности 224Ra (65.41 ± 0.68 dpm 100 л–1), а также повышение 223Ra (1.97 ± 0.11 dpm 100 л–1) зафиксированы в придонных водах устьевого взморья, поэтому источник данных изотопов – в основном донные осадки. Отмечена прямая корреляция в соотношениях “${\text{PO}}_{4}^{{3 - }}$ –224Ra” и “${\text{NH}}_{{\text{4}}}^{ + }$ –224Ra” и зафиксирован одновременный экстремум 224Ra, ${\text{PO}}_{4}^{{3 - }}$ и ${\text{NH}}_{{\text{4}}}^{ + }$ в придонных водах устьевого взморья, который пространственно совпадает с районом наибольшей плотности поселений полихет – активных биоирригаторов поровых вод.
ВВЕДЕНИЕ
Радий – щелочноземельный элемент, который в речной воде преимущественно адсорбируется, но существует в виде растворенного Ra2+ в морской воде из-за ее высокой ионной силы [25, 49]. Десорбция Ra из речной взвеси в эстуариях происходит на ранней стадии зоны смешения (ЗС) речных и морских вод до солености 10‰ [20, 22]. Поэтому растворенные изотопы 224Ra, 223Ra, 228Ra с периодом полураспада соответственно 3.6 и 11.5 сут и 5.7 года демонстрируют неконсервативную зависимость от солености воды в эстуариях при наличии речной взвеси [19, 20, 26, 30, 40]. Второй источник растворенного радия для эстуария – поток воды из осадков, который предполагает обмен поровыми водами (“pore water exchange” (PWE) [43]) и в более широком масштабе подземного устья – разгрузку субмаринных грунтовых вод (“submarine groundwater discharge” (SGD)) [32]. SGD включает в себя разгрузку одновременно пресных материковых и рециркулирующих морских вод в прибрежных водоносных горизонтах (“recirculated submarine groundwater discharge” (RSGD)) [18, 31]. Разгрузка пресных SGD в глобальном водном балансе составляет только 4.5 ± 3.2% от потоков речной воды в океан [15], но именно RSGD вносит 90% потока воды от общего SGD [24]. В ряде работ показано, что PWE может формировать одновременно потоки изотопов радия и биогенных веществ, конкурирующие с их потоками при SGD в небольших прибрежно-морских акваториях [27, 34, 41, 42]. Это во многом происходит из-за того, что донные отложения (ДО) обогащены биогенными веществами вследствие диагенеза в них органического вещества [14]. Поэтому поверхностный сток и поток PWE вносят доминирующий вклад в потоки Ra и биогенных веществ в эстуариях по отношению к пресным водам SGD [21, 23, 37, 38, 42 ]. Обмен поровой воды в прибрежно-морских бассейнах во многом формируется за счет биотурбации (построение нор инфауной и сифональное питание) и биорригации (вентиляция нор) [17, 48, 50]. Однако только относительно недавно было показано, что именно инфауна может быть определяющим звеном в потоках Ra и биогенных веществ при PWE в небольших заливах с выраженной сезонной изменчивостью температуры воды [42]. Морские воды, находящиеся под влиянием источника Ra в суточном и синоптическом диапазонах времени, обогащаются только короткоживущими изотопами 224Ra и 223Ra, в то время как на накопление долгоживущих изотопов 228Ra требуются годы [40]. Это позволяет устанавливать временны́е масштабы динамики вод и биогеохимических процессов в прибрежно-морских акваториях под влиянием PWE, SGD и речного стока.
Обобщение публикаций, связанных с изучением биогенных веществ в эстуарии р. Раздольной, можно найти в работах В.И. Звалинского с соавторами [1–3]; однако публикации, нацеленные на исследование изотопов Ra для данного водного объекта, авторами настоящей статьи не найдены.
Цель данной работы – выявить доминирующие источники и обсудить пути миграции изотопов Ra совместно с основными биогенными веществами на примере эстуария высокомутной р. Раздольной в паводок. В период паводка скорость обмена вод интенсифицирована из-за высоких скоростей стокового и компенсационного потоков воды, что представляет частое, но относительно краткосрочное явление во многих районах Мирового океана, находящихся под влиянием рек. Поэтому результаты данной статьи отражают ситуацию, характерную для процессов, имеющих короткопериодный (суточный и синоптический) диапазон изменчивости, требующий оперативных полевых исследований активности изотопов Ra и концентрации биогенных веществ.
ОБЪЕКТ И МЕТОДЫ ИССЛЕДОВАНИЙ
Трансграничная р. Раздольная (Китай – Приморский край Российской Федерации) впадает в северную часть Амурского залива (зал. Петра Великого, Японское море) (рис. 1). Эстуарий, по [6] и по уточненной терминологии в [9], имеет протяженность ~50 км и расположен в пределах заболоченной Раздольненской депрессии и северной части Амурского залива (рис. 1). Площадь водосбора р. Раздольной составляет 16 800 км2.
Рис. 1.
Район исследований, схема расположения станций и плотность поселения полихет, экз/м2, указанная на вставке в виде изолиний согласно [9]. РВ – речные воды, ВЭ – вершина эстуария, УБ – устьевой бар.
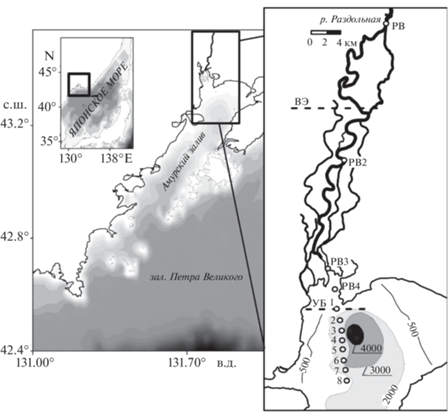
Средний расход реки на посту в с. Тереховка при осреднении за период 10 лет с 2008 г. составляет 97.8 м3/с. В водном режиме реки выделяется устойчивая зимняя межень со среднемесячным расходом 2–3 м3/с в январе и феврале (рис. 2), абсолютный минимумом расхода 0.3 м3/с случается в феврале. Весеннее половодье наблюдается в мае (рис. 2). Во время летних и осенних паводков расход регулярно >400 м3/с, а абсолютный максимум расхода на пике паводков в отдельные годы >3000 м3/с (рис. 2).
Рис. 2.
Среднемесячные, максимальные и минимальные расходы Q, м3/с, р. Раздольной за период 2008–2018 гг.
Динамика ЗС в эстуарии р. Раздольной в наибольшей степени определяется водным режимом реки [2]. При расходе >400 м3/с ЗС вытесняется за пределы устьевого бара и представляет собой линзу распресненных вод в Амурском заливе [2]. Относительно небольшие приливы высотой ~40 см не влияют на дальность проникновения ЗС, но определяют внутрисуточную изменчивость скорости течений в эстуарии [4].
В свою очередь положение ЗС в теплый период года определяет положение района наибольшей интенсивности биогеохимических процессов в эстуарии [13, 44]. В паводок река становится доминирующим источником биогенных веществ для Амурского залива [8]. На пике паводков концентрация взвеси достигает 2800 мг/л при среднегодовой концентрации 154 мг/л [6]. Резкое и многократное увеличение потоков биогенных веществ и взвеси приводит к импульсной эвтрофикации Амурского залива и вызывает формирование гипоксии придонных вод в летний сезон [12, 47]. Сгонные ветры в конце лета – начале осени приводят к затоку в залив в ходе сезонного апвеллинга промежуточных Япономорских вод, обогащенных кислородом относительно придонных гипоксийных вод Амурского залива [12, 47]. При этом в летний сезон наблюдается преимущественно циркуляция “даунвеллингового” типа, способствующая развитию гипоксии [46].
Относительно высокие содержания органического углерода в донных осадках северной части Амурского залива [14] способствуют развитию поселений полихет в данном районе до 6360 экз/м2 при средней плотности 1480 экз/м2 (рис. 1) [10].
Исследовали поверхностный и придонный слои воды толщиной 1 м. Пробы воды были отобраны с помощью 5-литрового батометра Нискина: 30 июня 2020 г. за <4 ч на восьми станциях, расположенных в ЗС (рис. 1); 1 июля 2020 г. на четырех станциях с речными водами (РВ). Полевым работам предшествовали выпадение осадков за 4 сут в количестве 194 мм в районе нижнего водосбора реки и умеренные юго-восточные ветры. Пик уровня воды в реке пришелся на 28 июня и составил 400 м3/с, после чего 1 июля последовал спад расхода до 231 м3/с. В ходе работ погода была безветренная. На каждой станции проводили зондирование водной толщи зондом “SBE 19Plus” (“Sea Beard Electronics”, США) для получения профилей электропроводности, температуры, мутности (в единицах “formazine turbidity unit” (FTU)). Для получения профиля О2 использовали дополнительный оптический датчик О2 “ARO2-Infinity” (“JFE Advantech Co., Ltd.”, Япония) с точностью 2% в диапазоне 0–200% насыщения вод О2. На каждой станции определяли глубину видимости диска Секки. Пробы воды, отобранные для определения растворенных в воде радионуклидов (223Ra, 224Ra, 228Ra), были отфильтрованы в лаборатории в день отбора, биогенные вещества измерены на следующий день, а соленость S – в течение недели. Соленость измерена с использованием солемера “Guildline Autosal 8400B” (“Guildline Instruments”, США) с погрешностью 0.002‰.
Для анализа 223Ra, 224Ra, 228Ra образцы воды объемом 22 л в день отбора отфильтровали через полипропиленовый картридж и вату MnO2 со скоростью ≤0.3 л мин−1 для получения эффективной экстракции 97% Ra [29]. C помощью системы RaDeCC (Radium Delayed Coincidence Counter) определены радиоизотопы 224Ra и 223Ra [33]. Для поправки на поддерживаемый 224Ra проводилась вторая серия измерений через 2–6 недель, чтобы позволить начальной избыточной активности 224Ra достичь векового равновесия с 228Th, который также абсорбировался на Mn-волокне [33, 36]. Активность 228Ra измерена через 6–12 мес. с поправкой на распад 228Th [29]. Для калибровки RaDeCC при измерениях 224Ra, 223Ra и 228Ra использовали стандарт 232Th, как предложено в работе [35].
Общий фосфор Total P и общий азот Total N определяли на автоматическом анализаторе “Skalar San++” (“Skalar”, Нидерланды). Концентрацию аммония (${\text{NH}}_{{\text{4}}}^{ + }$) определяли методом индофенолов. Нитраты ${\text{NO}}_{{\text{3}}}^{ - }$, нитриты ${\text{NO}}_{{\text{2}}}^{ - }$, растворенные силикаты DSi и растворенный неорганический фосфор DIP измеряли стандартными колориметрическими методами. Подробная информация о методах, используемых для анализа биогенных веществ, приведена в [7].
РЕЗУЛЬТАТЫ
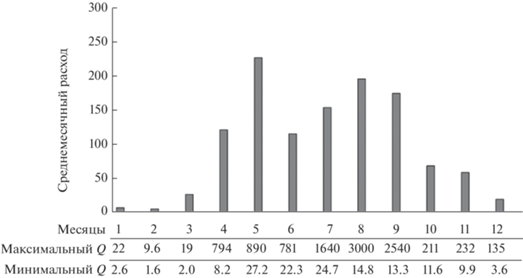
Речная (верхняя) граница ЗС, выделяемая по изогалине 1‰, примыкала к устьевому бару в районе ст. 1 (рис. 3а). На баре у дна соленость воды была 13.2‰, в то время как на поверхности ≤1‰. На относительно небольшом расстоянии от бара в направлении реки на ст. RW 4 (рис. 1) придонный слой был занят речной водой. Стоковое течение формировало линзу высокомутных вод с соленостью <6‰ на расстоянии до 6 км от устьевого бара (рис. 3а). Наибольшая соленость воды (32.7‰) наблюдалась в придонном слое толщиной ~2 м на трех мористых станциях (ст. 6–8). Температура воды в поверхностном слое была ~16°С, а у дна снижалась до 11°С (рис. 3б). Судя по поднятию изолиний (рис. 3а, 3б), в мористой части эстуария формировался компенсационный поток относительно холодных морских вод над устьевым взморьем, что характерно для эстуариев [39]. Динамика вод прослеживалась в поле распределения условной плотности воды в виде двухслойной эстуарной циркуляции, вызванной эффектом вовлечения придонных вод стоковым течением (рис. 3в).
Рис. 3.
Профили солености (а), ‰, температуры (б), °С, условной плотности (в). Отсчет расстояния от устьевого бара.
В речных водах активность растворенных изотопов Ra была низкой (табл. 1). Это было ожидаемо, поскольку в речной воде Ra находится в виде химических соединений, адсорбированных взвесью, которые в процессе пробоподготовки удаляются. С ростом солености наблюдается существенное увеличение активности всех трех измеренных изотопов (табл. 1). При этом наибольшая активность изотопов 223Ra и 228Ra, имеющих период полураспада 11.5 сут и 5.7 года соответственно, зафиксирована в наиболее динамичном районе – в начале ЗС на баре (табл. 1). В распределении активности изотопа 224Ra с периодом полураспада 3.6 сут также отмечено повышение на баре, но экстремум был приурочен к придонным водам на ст. 4 при глубине 8.3 м и солености 32.16‰ (табл. 1).
Таблица 1.
Номера станций, глубина отбора проб H, м, соленость воды S, ‰, активность изотопов радия, dpm 100 л–1
| Станция | H | S | 224Ra | 223Ra | 228Ra | 224Ra/223Ra |
|---|---|---|---|---|---|---|
| РВ | 0.3 | 0.06 | 13.17 ± 0.28 | 0.16 ± 0.08 | 1.17 | 84.2 |
| 1 | 0.3 | 0.13 | ||||
| 1 | 0.9 | 13.20 | 53.22 ± 0.58 | 2.64 ± 0.08 | 163.07 ± 0.03 | 20.2 |
| 2 | 0.2 | 1.85 | ||||
| 2 | 4.4 | 26.66 | 15.30 ± 0.34 | 0.70 ± 0.02 | 81.34 ± 0.01 | 21.9 |
| 3 | 0.1 | 6.21 | ||||
| 3 | 5.8 | 26.58 | 20.59 ± 0.36 | 1.14 ± 0.04 | 70.01 ± 0.03 | 18 |
| 4 | 0.4 | 6.51 | ||||
| 4 | 8.3 | 32.16 | 65.41 ± 0.68 | 1.80 ± 0.09 | 55.15 ± 0.04 | 36.3 |
| 5 | 0.4 | 6.65 | 31.52 ± 0.50 | 1.95 ± 0.09 | 66.63 ± 0.02 | 16.1 |
| 5 | 12.5 | 32.64 | 32.68 ± 0.54 | 1.97 ± 0.11 | 46.31 ± 0.04 | 16.6 |
| 6 | 0.2 | 9.04 | ||||
| 6 | 16.2 | 32.87 | 21.97 ± 0.38 | 1.03 ± 0.03 | 40.09 ± 0.03 | 21.4 |
| 7 | 0.2 | 11.79 | 21.24 ± 0.59 | 0.86 ± 0.10 | 79.52 ± 0.01 | 24.6 |
| 7 | 17.9 | 32.93 | 39.84 ± 0.57 | 1.16 ± 0.05 | 57.28 ± 0.03 | 34.4 |
| 8 | 0.3 | 17.04 | 22.42 ± 0.38 | 1.12 ± 0.04 | 54.36 ± 0.03 | 19.9 |
| 8 | 19.2 | 32.98 | 28.09 ± 0.46 | 1.84 ± 0.10 | 54.47 ± 0.03 | 15.2 |
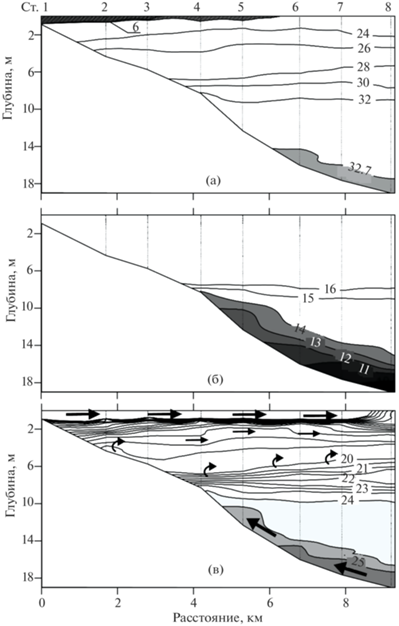
Следует отметить, что повышение активности всех трех изотопов Ra в районе бара при солености 13.2‰ (ст. 1) было зафиксировано в водах с высокой мутностью (рис. 4). В придонном слое воды с соленостью >32‰ также наблюдалась повышенная активность 224Ra с экстремумом на ст. 4, тогда как для 223Ra повышенная активность в этих водах прослеживалась в меньшей степени, а для 228Ra полностью отсутствовала (рис. 4).
Рис. 4.
Зависимость активности изотопов радия от солености и от мутности. 1 – поверхностный слой воды, 2 – придонный слой воды.
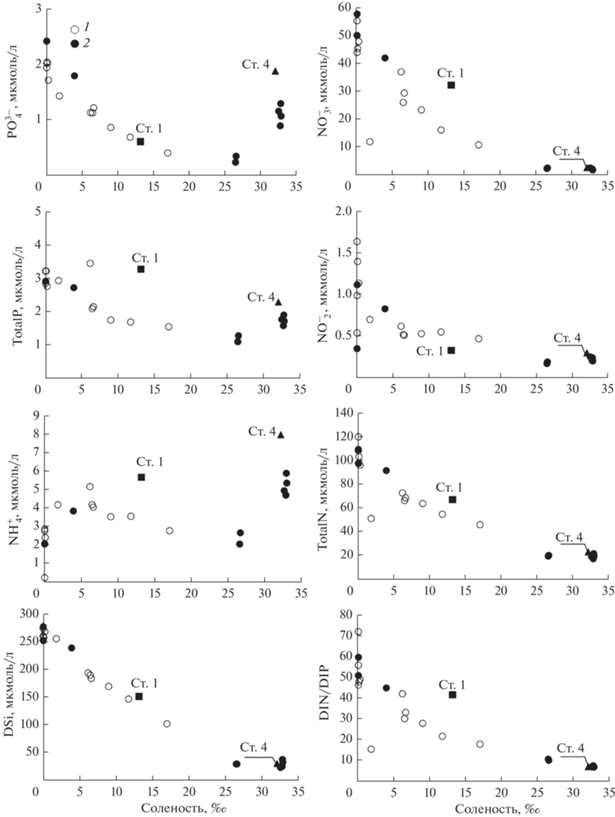
Также зафиксировано повышение концентрации для Total P, ${\text{NH}}_{4}^{ + }$, DSi, ${\text{NO}}_{3}^{ - }$, Total N в районе бара на ст. 1 (рис. 5). Однако для ${\text{NH}}_{4}^{ + }$ и ${\text{PO}}_{4}^{{3 - }}$ максимальные концентрации зафиксированы в водах с наибольшей активностью 224Ra на ст. 4 (рис. 4, 5). Также отмечен общий повышенный уровень концентраций ${\text{NH}}_{4}^{ + }$ и ${\text{PO}}_{4}^{{3 - }}$ в придонных водах с соленостью >32‰, как и для распределения 224Ra.
ОБСУЖДЕНИЕ
Поток биогенных веществ со стоком р. Раздольной был изучен ранее в [8]. Концентрации биогенных веществ в ДО были измерены, а оценки их потоков с участием PWE проведены для мелководных бассейнов зал. Петра Великого [11, 14]. В обсуждении результатов настоящей работы внимание уделяется обнаруженным совместным потокам Ra и биогенных веществ при десорбции из речной взвеси и из донных отложений на устьевом взморье р. Раздольной.
Согласно результатам экспериментов [20], речная взвесь – определяющий фактор для активности десорбированного Ra на начальном этапе зоны смешения. Ее содержание в водах р. Раздольной во многом зависит от расхода реки: минимальная концентрация взвеси составляет ~5 мг/л в зимнюю межень, но может быть >2800 мг/л в период дождей [6, 8]. В ходе настоящих работ видимость диска Секки была <5 см, а вода имела темно-коричневый цвет. Для оценки содержания взвеси использовался датчик мутности, предел показаний которого при имеющейся калибровке зонда (2500 FTU – 1000 мг/л) был превышен, поэтому точных результатов по данному параметру в реке не смогли получить. Интенсивное поступление взвеси в реку было связано с выпадением атмосферных осадков накануне работ непосредственно в районе эстуария, когда за предшествующие четверо суток количество осадков составило 194 мм. Всего же за июнь в районе эстуария выпало 292 мм осадков (архив RP5) при месячной норме 110 мм. Содержание долгоживущих изотопов 228Ra во взвеси водосбора может только увеличиваться, так как их период полураспада существенно превышает возраст вод от истоков, в то время как короткоживущие изотопы подвергаются распаду за период возраста речной воды на водосборе. Таким образом, величина расхода реки может влиять преимущественно на количество растворенных долгоживущих изотопов 228Ra, поставляя их в приемный бассейн при десорбции.
Принимая во внимание то, что накопление изотопов Ra в ДО определяется их константами распада, рециркуляция морской воды в верхнем слое ДО на протяжении суток сопровождается ростом активности только короткоживущих изотопов 223Ra и 224Ra [40], в то время как воды водоносных горизонтов обычно обогащены, помимо 223Ra и 224Ra, долгоживущими изотопами 228Ra [45]. Для изотопов 223Ra и 224Ra разброс на графике “активность–мутность” гораздо больший, особенно для 224Ra, по сравнению с долгоживущим изотопами 228Ra (рис. 4). Экстремум данного радионуклида наблюдался в придонных водах с соленостью >32‰ (табл. 1), которые удалены от источника взвеси в районе устьевого бара. Поэтому поступление радионуклидов 223Ra и 224Ra из ДО оказалось более значимым, а по 224Ra – доминирующим по сравнению с поступлением при десорбции в районе устьевого бара.
Согласно [42], в илистых отложениях прибрежных акваторий активность 224Ra обычно составляет 1100‒1300 dpm 100 л–1. Активность 224Ra для морской воды исследуемого объекта в отсутствии речного стока получена в период ледостава и составляет 6.7 dpm 100 л–1 [5]; т. е. активность 224Ra в поровых водах осадков, принятая из литературных данных (~1000 dpm 100 л–1), может превышать по меньшей мере в 100 раз значение для морской воды на устьевом взморье р. Раздольной.
Возможным механизмом поставки Ra в водную толщу при PWE может быть метаногенез, когда через толщу осадка просачиваются пузырьки метана и увлекают за собой поровые воды [43]. Однако масштаб этого явления не может быть распространен на все устьевое взморье вплоть до глубин 19 м на самой мористой станции настоящих исследований. Поток Ra при повторном взмучивании терригенной взвеси, осевшей на дно, также исключаем, поскольку это не приводит к повышению аммонийного азота и фосфат-ионов. Учитывая, что над устьевым взморьем отсутствует сколь-нибудь значимое неконсервативное повышение 228Ra, возможное влияние SGD на поток Ra в приемный бассейн в данную водную фазу р. Раздольной также исключаем.
PWE зависит от видового состава и численности инфауны, поведение которой определяется условиями окружающей среды – в основном температурой и содержанием растворенного О2 [17, 27, 48, 50]. PWE при участии инфауны достигает глубины ~40 см в ДО за <46 ч [27]. Наиболее активные биоирригаторы – полихеты [49]. Поток PWE при участии полихет будет существенно изменяться в разных частях Амурского залива и, вероятно, достигнет максимума в районе с наибольшей плотностью экземпляров на глубине ~8 м (рис. 1), на которой в наблюдениях был обнаружен экстремум активности 224Ra (табл. 1). Поток PWE при биоирригации, на примере зал. Альфакс Средиземного моря со средней плотностью полихет 10 400 экз/м2, достигает ∼20 л/м2 сут–1 [42]. По аналогии с зал. Альфакс поток PWE при участии полихет в Амурском заливе при плотности поселения 1480 экз/м2 [10] будет, в грубом приближении, в 7 раз меньше, т. е. 2.85 л/м2 сут–1. При этой приближенной оценке поток PWE из ДО площадью 132 км2 в северной части Амурского залива составит 376 × 103 м3/cут, или 4 м3/c. В придонном слое активность 224Ra должна приближаться к таковой в поровых водах (>1000 dpm 100 л–1), однако этому препятствует адвективный перенос придонных вод (рис. 3).
В настоящих исследованиях при расходе реки в день отбора 262.9 м3/с активность 224Ra, десорбированного на баре, в сумме с активностью 224Ra в реке составила 66.39 dpm 100 л–1 (табл. 1). Поток Ra c речным стоком при данных величинах расхода и концентрации взвеси в реке будет превышать поток Ra за счет PWE, несмотря на >100-кратное превышение активности 224Ra в ДО. Вероятно, в межень, когда количество содержащей Ra речной взвеси снизится многократно, как указано выше, поток Ra за счет PWE будет доминирующим. Этому будет способствовать возрастание до 40% [17, 27, 48, 50] потока PWE за счет биоирригации при летнем прогреве вод и понижении концентрации О2 до уровня гипоксии [12]. В зал. Альфакс, например, летом для всей площади поток PWE достигает 13.9, а зимой только 3.1 м3/c [42].
Повышение активности 224Ra совместно с ${\text{PO}}_{4}^{{3 - }}$ и ${\text{NH}}_{4}^{ + }$ (рис. 6), а также повышение активности 223Ra при неизменной солености (рис. 4) указывает на их единый источник. Для биогенных веществ таким источником может быть органическое вещество, подвергающееся микробиологической деструкции как в воде, так и в ДО [14]. Для северной части Амурского залива концентрации ${\text{PO}}_{4}^{{3 - }}$ и ${\text{NH}}_{4}^{ + }$ в летний сезон в поровой воде ДО на горизонте 5 см увеличены в 5 раз и в 34 раза соответственно по сравнению с придонным слоем воды [14]. Относительно малое повышение ${\text{PO}}_{4}^{{3 - }}$ на фоне роста ${\text{NH}}_{4}^{ + }$ в ДО связано с процессами адсорбции–десорбции в поровой воде [14]. Поэтому поровый обмен должен сопровождаться увеличением концентраций ${\text{PO}}_{4}^{{3 - }}$ и ${\text{NH}}_{4}^{ + }$, а также соотношением N/P. Соотношение N/P (рис. 5) в придонных водах с соленостью >32‰, однако, составляет 6.6–6.9, что существенно ниже, чем в соотношении Редфилда, равном 16. Поэтому, в северной части Амурского залива существует дополнительный поток соединений фосфора, вероятно, связанный с жизнедеятельностью полихет.
Рис. 6.
Зависимость концентрации биогенных веществ от активности изотопов 224Ra. 1 – поверхностный слой воды, 2 – придонный слой воды.
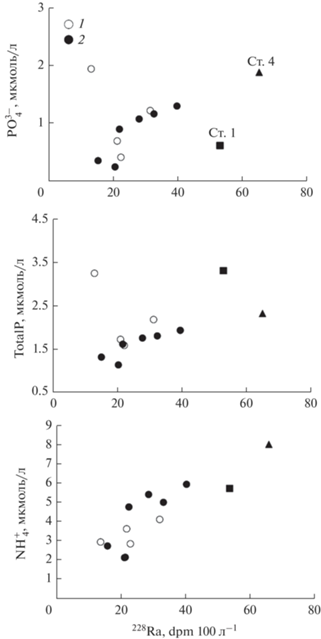
Для Амурского залива деструкция автохтонного органического вещества оказывает основное влияние на концентрации и распределение биогенных веществ в послепаводковый период [12, 47]. В этом случае максимумы биогенных веществ (${\text{PO}}_{4}^{{3 - }}$, ${\text{NH}}_{4}^{ + }$, а также DSI) в придонном слое воды пространственно совпадают с минимальной концентрацией О2, пониженной до состояния гипоксии (<2 мг/л) [12, 47]. Развитию придонной гипоксии при южных ветрах способствует даунвеллинг [46]; в то время как в момент паводка вследствие развития двухслойной эстуарной циркуляции формируется “апвеллинговая” структура распределения плотности (рис. 2). Такой вид циркуляции в Амурском заливе существует регулярно при развитии сгонных ветров и сопровождается адвекцией вод из открытой части залива с относительно высокой концентрацией О2, пониженной концентрацией ${\text{NH}}_{4}^{ + }$, но повышенной концентрацией ${\text{NO}}_{3}^{ - }$ [12, 47]. Однако в компенсационном потоке вод при концентрациях О2 ~ ~5 мг/л по мере его переноса над устьевым взморьем в направлении реки отмечено повышение содержания ${\text{PO}}_{4}^{{3 - }}$ и ${\text{NH}}_{4}^{ + }$, а также фосфора в органической форме и экстремум аквтиности 224Ra (рис. 3, 5, 6, 7 ). Взаимосвязь данных характеристик объясняется влиянием потока поровой воды за счет жизнедеятельности полихет.
Поскольку поток PWE имеет гораздо большую концентрацию веществ в сравнении с вышележащей водой, накопление веществ в придонном слое воды будет определяться интенсивностью адвективного переноса вод вдоль устьевого взморья. В эстуариях сроки адвективного обмена воды могут варьировать от нескольких дней до месяцев и зависят в основном от расхода речной воды [16]. В периоды паводков скорость обмена в эстуарии многократно возрастает за счет усиления стокового течения и интенсификации компенсационного потока в придонном слое вод. Для короткоживущих изотопов, а также биогенных веществ, которые активно задействованы в продукционно-деструкционных процессах, важен временнóй диапазон существования этого потока. Оценка пространственной изменчивости возраста воды в эстуариях с множеством потенциально возможных источников веществ, принимаемых в качестве индикаторов, – сложная задача. Отношения активности 224Ra/223Ra определяют возраст воды в синоптическом диапазоне и позволяют рассчитать “кажущийся возраст радия” (“apparent radium age”) [28]. Потоки растворенных радионуклидов 224Ra из ДО будут существенно увеличивать отношение 224Ra/223Ra в придонных водах в следующих случаях: 1) скорость вертикального перемешивания ниже, чем скорость переноса придонной воды вдоль дна; 2) преодоление придонными водами пути вдоль дна с обогащением 224Ra происходит быстрее, чем скорость распада этих изотопов. При выполнении этих условий происходит накопление 224Ra в придонных водах, что вызывает рост соотношения 224Ra/223Ra, тем самым уменьшая возраст воды, рассчитанный на основе модели с единым источником [28]. В исследуемом случае соотношение 224Ra/223Ra меняется контрастно от станции к станции в интервале от 15.2 до 36.3 при почти неизменной солености компенсационного потока (табл. 1; рис. 3), что указывает на неоднородность поступления 224Ra и 223Ra. Поэтому нет возможности рассчитать точный возраст придонных вод и оценить массовый баланс Ra, не располагая данными по активности 224Ra и 223Ra для поровых вод на каждой станции отбора. Однако само по себе наличие контраста свидетельствует о том, что возраст вод в компенсационном потоке имеет временнóй диапазон, сравнимый с временны́м диапазоном полураспада 224Ra – 3.6 сут, что согласуется с периодом развития наблюдаемой двухслойной циркуляции (рис. 3), сформированной на пике прохождения паводка за два дня до отбора проб воды. Таким образом, полученная картина распределения изотопов и биогенных веществ характерна для синоптического периода от 1 сут до 1–2 недель в зависимости от продолжительности и интенсивности паводка. После ослабления стокового течения и компенсационного потока происходит перестройка динамики вод на противоположную – с преобладанием даунвеллинга при южных ветрах [46].
ВЫВОДЫ
Рассмотрено распределение радионуклидов радия и биогенных веществ над устьевым взморьем р. Раздольной при развитии двухслойной эстуарной циркуляции, период существования которой определяется продолжительностью летнего паводка.
Установлено, что доминирующий процесс поступления в эстуарий растворенных изотопов 228Ra – десорбция из речной взвеси в начале зоны смешения, в районе устьевого бара. Повышение активности изотопов 223Ra и 224Ra в районе бара проявлялось в меньшей степени.
Обмен поровых вод с придонными водами устьевого взморья вызвал возникновение экстремума активности 224Ra и повышение 223Ra. Одновременный экстремум короткоживущих изотопов 224Ra, аммонийного азота и растворенного фосфора в придонном слое устьевого взморья объясняется их поступлением за счет биоирригации и биотурбации полихет.
Разгрузка пресных грунтовых вод на устьевом взморье р. Раздольной в исследуемую фазу водного режима реки на основе распределения изотопов Ra не выявлена.
Список литературы
Звалинский В.И., Марьяш А.А., Тищенко П.Я. и др. Продукционные характеристики эстуария реки Раздольной в период ледостава // Изв. ТИНРО. 2016. Т. 185. С. 155–174.
Звалинский В.И., Недашковский А.П., Сагалаев С.Г. и др. Биогенные элементы и первичная продукция эстуария реки Раздольной // Биология моря. 2005. Т. 31. № 2. С. 107–116.
Звалинский В.И., Тищенко П.П., Михайлик Т.А., Тищенко П.Я. Эвтрофикация залива Петра Великого // Океанологические исследования дальневосточных морей и северо-западной части Тихого океана / Под ред. В.А. Акуличева. Владивосток: Дальнаука, 2013. Кн. 1. С. 260–293.
Семкин П.Ю., Тищенко П.Я., Лобанов В.Б. и др. Обмен вод в эстуарии реки Раздольной (Амурский залив, Японское море) в период ледостава // Изв. ТИНРО. 2019. Т. 196. С. 123–137.
Семкин П.Ю., Тищенко П.Я., Чаркин А.Н. и др. Разгрузка грунтовых осолоненных вод в эстуарии реки Раздольной (Амурский залив) в феврале 2020 г. // Вод. ресурсы. 2021. Т. 48. №. 3. С. 254–259.
Состояние морских экосистем, находящихся под влиянием речного стока. Владивосток: Дальнаука, 2005. 261 с.
Методы гидрохимических исследований основных биогенных элементов. М.: ВНИРО, 1988. 120 с.
Михайлик Т.А., Тищенко П.Я., Колтунов А.М. и др. Влияние реки Раздольной на экологическое состояние вод Амурского залива (Японское море) // Вод. ресурсы. 2011. Т. 38. № 4. С. 474–484.
Михайлов В.Н., Горин С.Л. Новые определения, районирование и типизация устьевых областей рек и их частей–эстуариев // Вод. ресурсы. 2012. Т. 39. № 3. С. 243–257.
Мощенко А.В., Белан Т.А. Полихеты северной части Амурского залива (залив Петра Великого Японского моря): состав, структура и распределение таксоценов // Изв. ТИНРО. 2010. Т. 163. С. 264–285.
Тищенко П.Я., Борисенко Г.В., Барабанщиков Ю.А. и др. Потоки биогенных веществ на границе раздела вода–дно в мелководных бассейнах залива Петра Великого (Японское море) // Геохимия. 2021. В печати.
Тищенко П.Я., Лобанов В.Б., Звалинский В.И. и др. Сезонная гипоксия Амурского залива (Японское море) // Изв. ТИНРО. 2011. Т. 165. С. 108–129.
Тищенко П.Я., Семкин П.Ю., Тищенко П.П. и др. Гипоксия придонных вод эстуария реки Раздольная // ДАН. 2017. Т. 476. № 5. С. 576–580.
Тищенко П.Я., Ходоренко Н.Д., Барабанщиков Ю.А. и др. Диагенез органического вещества в осадках покрытых зарослями зостеры морской (Zostera Marina L.) // Океанология. 2020. Т. 60. № 3. С. 393–406.
Abbott B.W., Bishop K., Zarnetske J.P. et al. Human domination of the global water cycle absent from depictions and perceptions // Nat. Geosci. 2019. V. 12. № 7. P. 533–540.
Alber M., Sheldon J.E. Use of a date-specific method to examine variability in the flushing times of Georgia estuaries // Estuar. Coast. Shelf Sci. 1999 V. 49. Iss. 4. P. 469–482.
Aller R.C., Aller J.Y. Meiofauna and solute transport in marine muds // Limnol. Oceanogr. 1992. V. 37. № 5. P. 1018–1033.
Burnett W.C., Aggarwal P.K., Aureli A. et al. Quantifying submarine groundwater discharge in the coastal zone via multiple methods // Sci. Total En. 2006. V. 367. Iss. 2–3. P. 498–543.
Elsinger R.J., Moore W.S. 226Ra and 228Ra in the mixing zones of the Pee Dee River-Winyah Bay, Yangtze River and Delaware Bay estuaries // Estuar. Coast. Shelf Sci. 1984. V. 18. Iss. 6. P. 601–613.
Hancock G.J, Murray A.S. Source and distribution of dissolved radium in the Bega River estuary, Southeastern Australia // Earth Planet Sci. Lett. 1996. V. 138. Iss. 1–4. P. 145–155.
Gleeson J., Santos I.R., Maher D.T., Golsby-Smith L. Groundwater–surface water exchange in a mangrove tidal creek: Evidence from natural geochemical tracers and implications for nutrient budgets // Mar. Chem. 2013. V. 156. P. 27–37.
Gonneea M.E., Morris P.J., Dulaiova H., Charette M.A. New perspectives on radium behavior within a subterranean estuary // Mar. Chem. 2008. V. 109. P. 250–267.
Krest J.M., Moore W.S., Rama. 226Ra and 228Ra in the mixing zones of the Mississippi and Atchafalaya Rivers: indicators of groundwater input // Mar. Chem. 1999. V. 64. P. 129–154.
Kwon E.Y., Kim G., Primeau F. et al. Global estimate of submarine groundwater discharge based on an bservationally constrained radium isotope model // Geophys. Res. Lett. 2014. V. 41. Iss. 23. P. 8438–8444.
Langmuir D., Riese A.C. The thermodynamic properties of radium // Geochimica et Cosmochimica Acta. 1985. V. 49. Iss. 7. P. 1593–1601.
Li Y-H., Mathieu G., Biscaye P., Simpson H.J. The flux of Ra-226 from estuarine and continental shelf sediments // Earth Planet. Sci. Lett. 1977. V. 37. Iss. 2. P. 237–241.
Martin J.B., Cable J.E., Jaeger J. et al. Thermal and chemical evidence for rapid water exchange across the sediment-water interface by bioirrigation in the Indian River Lagoon, Florida // Limnol. Oceanogr. 2006. V. 51. Iss. 3. P. 1332–1341.
Moore W.S. Determining Coastal Mixing Rates Using Radium Isotopes // Cont. Shelf Res. 2000.V. 20. Iss. 15. P. 1993–2007.
Moore W.S. Fifteen years experience in measuring 224Ra and 223Ra by delayed coincidence counting // Mar. Chem. 2008. V. 109. Iss. 3–4. P. 188–197.
Moore W.S. Radium isotopes in the Chesapeake Bay // Estuar. Coast. Shelf Sci. 1981. V. 12. Iss. 6. P. 713–723.
Moore W.S. The effect of submarine groundwater discharge on the ocean // Annual Rev. Mar. Sci. 2010. V. 2. P. 59–88.
Moore W.S. The subterranean estuary: a reaction zone of ground water and sea water // Mar. Chem. 1999. V. 65. Iss. 1–2. P. 111–126.
Moore W.S., Arnold R. Measurement of 223Ra and 224Ra in coastal waters using delayed coincidence counter // J. Geophys. Res. 1996. V. 101. Iss. C1. P. 1321–1329.
Moore W.S., Beck M., Riedel T. et al. Radium-based pore water fluxes of silica, alkalinity, manganese, DOC, and uranium: A decade of studies in the German Wadden Sea // Geochim. Cosmochim. Acta. 2011. V. 75. Iss. 21. P. 6535–6555.
Moore W.S., Cai P. Calibration of RaDeCC systems for 223Ra measurements // Mar. Chem. 2013. V. 156. P. 130–137.
Moore W.S., de Oliveira J. Determination of residence time and mixing processes of the Ubatuba, Brazil, inner shelf waters using natural Ra isotopes // Estuar. Coast. Shelf Sci. 2008. V. 76. Iss. 3. P. 512–521.
Moore W.S., Krest J. Distribution of 223Ra and 224Ra in the plumes of the Mississippi and Atchafalaya Rivers and the Gulf of Mexico // Mar. Chem. 2004. V. 86. P. 105–119.
Peterson R.N., Burnett W.C., Taniguchi M. et al. Determination of transport rates in the Yellow River-Bohai Sea mixing zone via natural geochemical tracers // Continental Shelf Res. 2008. V. 28. Iss. 19. P. 2700–2707.
Pritchard D.W. What is an estuary: a physical viewpoint // Estuaries. Washington: Am. Ass. Adv. Sci., 1967. Publ. 83. P. 3–5.
Rama Moore W.S. Using the Ra quartet for evaluating groundwater input and water exchange in salt marshes // Geochim. Cosmochim. Acta. 1996. V. 60. Iss. 43. P. 4645–4652.
Rodellas V., Garcia-Orellana J., Masqué P., Font-Muñoz J.S. The influence of sediment sources on radium-derived estimates of Submarine Groundwater Discharge // Mar. Chem. 2015 V. 171. P. 107–117.
Rodellas V., Garcia-Orellana J., Trezzi G. et al. Using the radium quartet to quantify submarine groundwater discharge and porewater exchange // Geochim. Cosmochim. Acta. 2017. V. 196. Iss. P. 58–73.
Santos I.R., Eyre B.D., Huettel M. The driving forces of porewater and groundwater flow in permeable coastal sediments: A review // Estuar. Coast. Shelf Sci. 2012. V. 98. P. 1–15.
Shulkin V., Tishchenko P., Semkin P., Shvetsova M. Influence of river discharge and phytoplankton on the distribution of nutrients and trace metals in Razdolnaya River estuary, Russia // Estuar. Coast. Shelf Sci. 2018. V. 211. P. 166–176.
Szabo Z., de Paul V.T., Fischer J.M. et al. Occurrence and geochemistry of radium in water from principal drinking-water aquifer systems of the United States // Applied Geoch. 2012. V. 27. Iss. 3. P. 729–752.
Tishchenko P.P., Tishchenko P.Ya., Lobanov V.B. et al. Summertime in situ monitoring of oxygen depletion in Amursky Bay (Japan/East Sea) // Continent. Shelf Res. 2016. V. 118. P. 77–87.
Tishchenko P.Ya., Tishchenko P.P., Lobanov V.B. et al. Impact of the transboundary Razdolnaya and Tumannaya Rivers on deoxygenation of the Peter the Great Bay (Sea of Japan) // Estuar. Coast Shelf Sci. 2020. V. 239. https://www.sciencedirect.com/science/article/ pii/S0272771418301689?via%3Dihub
Volkenborn N., Polerecky L., Hedtkamp S.I.C. et al. Bioturbation and bioirrigation extend the open exchange regions in permeable sediments // Limnol. Oceanogr. 2007. V. 52. Iss. 5. P. 1898–1909.
Webster I.T., Hancock G.J., Murray A.S. Modelling the effect of salinity on radium desorption from sediments // Geochim. Cosmochim. Acta. 1995. V. 59. P. 2469–2476.
Woodin S.A., Wethey D.S., Volkenborn N. Infaunal hydraulic ecosystem engineers: cast of characters and impacts // Integr. Comp. Biol. 2010. V. 50. P. 176–187.
Дополнительные материалы отсутствуют.