

Зоологический журнал, 2019, T. 98, № 1, стр. 37-42
Выработка поведенческого навыка у муравьев Myrmica (Hymenoptera, Formicidae) на характер освещения гнезда
О. Б. Гилева 1, А. В. Гилев 1, *
1 Институт экологии растений и животных УрО РАН,
620144 Екатеринбург, Россия
* E-mail: gilev@ipae.uran.ru
Поступила в редакцию 02.07.2015
После доработки 20.06.2018
Принята к публикации 10.05.2018
Аннотация
Изучали возможность выработки поведенческого навыка по поиску кормушки у муравьев Myrmica ruginodis на характер светового сигнала (непрерывное или прерывистое освещение). В ходе серии опытов выяснилось, что муравьи способны очень быстро, в течение трех предъявлений, обучаться находить кормушку, используя сигналы, которые не связаны с освещением и которые невольно подает исследователь. В частности, муравьи способны ориентироваться по картине условного “неба” – прозрачной арены, расположенной в 30 см над субстратом, и реагируют на запах раствора меда с расстояния в несколько метров. На характер освещения поведенческий навык вырабатывался длительное время, без видимого прогресса, и возник лишь к 20-му предъявлению по типу инсайта.
Муравьи (Hymenoptera, Formicidae) – группа высокоорганизованных общественных перепончатокрылых насекомых, обладающих сложным поведением. У них легко удается выработать условные рефлексы (Плеханов, Кауль, 1976), они хорошо ориентируются по наземным и астрономическим ориентирам (Кауль, 1985; Кауль, Коптева, 1982), обладают развитой системой коммуникации (Резникова, Рябко, 1990; Новгородова, 2006) и способностью к счету (Резникова, Рябко, 1999). Однако это все относится, прежде всего, к муравьям рода Formica, которые являются признанными интеллектуалами и с которыми было проведено большинство работ подобного рода. Более мелкие, но практически вездесущие, муравьи рода Myrmica в этом отношении изучены гораздо слабее. Известно, что они сравнительно легко осваивают многоальтернативный лабиринт (Дашевский и др., 1989; Карась и др., 1986) и способны к переделке навыка (Удалова и др., 1991). Показано, что мирмики способны обучаться друг у друга, достраивая неполный поведенческий стереотип (Резникова и др., 2008). В то же время их эврибионтность, способность обитать в очень разных условиях внешней среды, должна сопровождаться высокой пластичностью поведения, что открывает широкие перспективы для сравнительных исследований.
Характерно, что в большинстве исследований ставится задача изучить способности отдельных особей к научению. При этом для муравьев более характерны взаимодействия с сородичами, формирование группового поведения либо разделения функций среди населения муравейника. Семья муравьев – сложная система из множества однородных элементов, тесно взаимодействующих между собой. Известно, что при формировании систем возникают дополнительные качества или свойства, которые не свойственны отдельным элементам системы. В этой связи представляется интересным изучить реакцию целой семьи муравьев, как некой социальной совокупности, общности, системы, сформированной отдельными особями.
В большинстве опытов пищевое подкрепление обычно располагается рядом с объектом, который имеет сигнальное значение. В этих условиях в первую очередь обучаются отдельные особи, которые затем передают информацию другим муравьям. Можно попробовать смоделировать такую ситуацию, когда семья муравьев будет вынуждена реагировать как целое. Такая ситуация теоретически может складываться, когда сигнал воспринимают одни особи, а реагировать (искать пищевое подкрепление) должны другие. В простейшем случае сигнальный стимул и пищевое подкрепление должны быть пространственно разобщены.
Таким образом, целью нашей работы было изучение возможности выработки поведенческого навыка (определение местоположения кормушки) на стимул, пространственно удаленный от места пищевого подкрепления. Поскольку муравьи воспринимают окружающий мир иначе, чем мы, и могут ориентироваться по малозаметным для нас стимулам, в задачи нашей работы входило также описание различных поведенческих реакций муравьев, как для контроля опыта, так и для выявления новых закономерностей.
МАТЕРИАЛ И МЕТОДИКА
Исследование проводили на семье муравьев Myrmica ruginodis, которую взяли из природы и содержали в лабораторных условиях при комнатной температуре в прозрачном террариуме-формикарии с размерами 50 × 30 см и высотой 30 см (рис. 1). Муравьи были взяты в начале октября, до наступления отрицательных температур, и выдержаны в холодильнике в течение недели. Численность семьи составляла примерно 400–500 особей, было отмечено несколько самок, расплод – зимующие личинки. В формикарий поместили гнездо муравьев целиком, вместе с субстратом. Условия, близкие к природным, были созданы с помощью кусочков гнилой древесины и подушек мха. Для того чтобы мох не высыхал, поддерживалась достаточно высокая влажность. Условия в целом оказались довольно благоприятными для муравьев, и они даже смогли вывести крылатых самок в конце февраля.
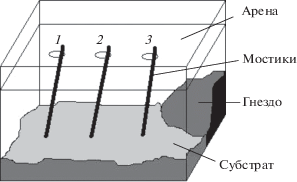
Над формикарием установили прозрачную арену 50 × 30 см с тремя отверстиями на расстоянии 15 см одно от другого. Через эти отверстия на субстрат были опущены деревянные мостики (рис. 1), по которым муравьи могли беспрепятственно выходить на арену и обследовать ее. Муравьев кормили углеводной (раствор сахара 1–2 раза в день) и белковой (мясной фарш или измельченное вареное яйцо 1 раз в 2–3 дня) пищей. Кормушки выставляли на арене, чтобы муравьи привыкали искать здесь пищу.
Опыт начался 15 октября и длился около месяца. Возле одного из мостиков 1–2 раза в день ставили кормушку – ватный тампон на пластиковой подложке с раствором сахара, в который, для усиления привлекательности, добавляли каплю меда. По сигналу муравьи должны были выбрать мостик и подняться по нему. Кормушка оставалась доступной для муравьев в течение часа, после чего ее убирали. Муравьев при этом удаляли с кормушки мягкой кисточкой. Сигнал подавался вручную с помощью настольной лампы (лампа накаливания 60 Вт, 700 лм), освещавшей гнездо через боковую стенку формикария. Лампа располагалась таким образом, чтобы освещать преимущественно область гнезда, однако отсветы могли видеть и рабочие, находящиеся в других частях формикария.
Постоянное освещение в течение минуты означало наличие кормушки у мостика № 1, дальнего от гнезда, прерывистое освещение (включение и выключение лампы с периодичностью 2–3 с в течение 15–30 с) – кормушку у мостика № 3, ближнего к гнезду (мостик № 2 не использовался, но оставался в формикарии). Положение кормушки каждый раз выбирали с помощью датчика случайных чисел (Лакин, 1990), чтобы исключить возможность запоминания последовательности предъявлений муравьями. По ходу выполнения работы условия опыта несколько изменялись в соответствии с реакциями муравьев. Поэтому весь опыт был условно разбит на 3 серии, которые следовали одна за другой без перерыва. Поскольку все изменения вводились по результатам выполнения предыдущей серии, они описываются в разделе “Результаты”.
Критерием правильности выбора служил выход первых муравьев по нужному мостику после подачи светового сигнала. Дополнительно каждые 10 мин проводили подсчет муравьев, вышедших к кормушке. С момента постановки кормушки и в течение всего сеанса также велись наблюдения за перемещениями муравьев в формикарии и на арене, что дало возможность выявить особенности поведения муравьев в условиях изменения местоположения корма и формирования навыка.
РЕЗУЛЬТАТЫ
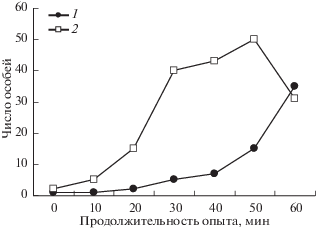
Опыт 1. В первом варианте опыта кормушку помещали на арену возле выбранного случайным образом мостика, затем с помощью лампы муравьям подавали соответствующий сигнал. На рис. 2 показана динамика накопления муравьев на кормушке при втором и третьем предъявлении (оба раза кормушка ставилась к мостику № 1). Видно, что во втором предъявлении рабочие особи скапливались на кормушке сначала медленно (рис. 2, 1). В это время приходили в основном одиночные муравьи, очевидно, разведчики, которые находили кормушку, набирали раствор сахара и уходили в гнездо. Быстрое накопление рабочих наблюдалось лишь в самом конце опыта, когда у муравьев началась мобилизация в гнезде (или на территории). У M. ruginodis мобилизация осуществляется приводом группы (Длусский и др., 1978; Путятина, 2007), что мы могли непосредственно наблюдать: по мостику и арене к кормушке двигались группы рабочих по 3–5 особей.
Рис. 2.
Характер накопления рабочих муравьев на кормушке в первой серии опытов: 1 – предъявление № 2, 2 – предъявление № 3.
Начало движения групп муравьев к кормушке в данном случае служило хорошим маркером того, что муравьи действительно нашли кормушку и знают, по какому мостику надо подниматься. При этом муравьи с самого начала опытов научились выходить к кормушке по тому мостику, около которого она была установлена. Попытки отыскать кормушку с других мостиков были единичны и не приводили к формированию потока мобилизованных групп.
Уже с третьего предъявления муравьи быстро, в течение первых 20 мин, скапливались на кормушке (рис. 2, 2). Движение групп по нужному мостику отмечалось почти сразу же после предъявления светового сигнала. В дальнейшем число муравьев достигало некоторого уровня насыщения и далее не изменялось: число покидающих кормушку фуражиров с сахарным сиропом в зобике было примерно равно числу приходящих к ней. Кривая накопления фуражиров приняла S-образный вид.
Казалось бы, можно сделать вывод, что муравьи научились, и достаточно быстро, всего за три предъявления, распознавать световой сигнал и находить кормушку на соответствующем мостике. Однако дальнейшие наблюдения за поведением муравьев на арене и в формикарии дали неожиданный результат: в последующих опытах движение групп фуражиров на нужном мостике начиналось до подачи светового сигнала. Очевидно было, что муравьи использовали не свет, а какой-то другой сигнал, который мы, не замечая того, невольно подавали, и который они легко распознавали. Причем этот сигнал не вызывал массовой неспецифической активации, выхода на арену множества муравьев по всем мостикам, а точно указывал, по какому мостику нужно подниматься. Этим сигналом оказалась сама кормушка. Дно арены было прозрачным, и муравьи могли видеть, у какого мостика она поставлена.
Этот результат сам по себе оказался достаточно удивительным. Дно арены располагалось в 30 см от субстрата формикария, являясь условным “небом” для муравьев, находящихся в гнезде и на территории. Известно, что муравьи Formica s. str. способны ориентироваться по астрономическим ориентирам и рисунку крон над головой (Кауль, 1985; Кауль, Коптева, 1982), но это относительно крупные муравьи, с развитыми органами зрения и фуражирующие преимущественно на поверхности почвы, как в дневное, так и в ночное время. Мирмики же много времени проводят в толще подстилки и имеют сравнительно небольшие глаза, что не предполагает хорошего зрения. В то же время Каммерс (Cammaerts, 2012) показала, что мирмики способны распознавать объекты различной формы и цвета и, теоретически, ориентироваться по кронам деревьев и звездному небу. В нашем случае муравьи оказались способны увидеть изменения в картине “неба” над головой и, более того, связать их с появлением корма. Примечательно, что муравьи оказались в состоянии обучиться этому всего с трех предъявлений.
Всего в данном варианте опыта было 6 предъявлений.
Опыт 2. В следующей серии опытов мы исключали возможность ориентации муравьев по месту расположения кормушки на арене. Сначала семье предъявлялся световой стимул, гнездо освещалось лампой, затем, после выхода первых рабочих на арену по соответствующему мостику, выставлялась кормушка.
Муравьи также научились быстро находить кормушку, уже с 5-го предъявления (кормушка у мостика № 1) кривая накопления фуражиров на кормушке приняла характерный вид (рис. 2, 2). Однако наблюдения за поведением муравьев на арене и в формикарии показали, что муравьи и в этот раз не используют световой сигнал: движение муравьев вновь начиналось до подачи светового сигнала и одновременно по обоим мостикам. Таким образом, муравьи в обоих случаях появлялись возле нужного мостика и получали кормушку. Они вновь смогли найти стимул, который не значим для нас, но достаточно заметен для них.
Сигналом для начала движения муравьев в этот раз оказался запах раствора меда: перед началом опыта открытую кормушку приносили в помещение, где располагался формикарий. Это было обнаружено нами путем прямых наблюдений за муравьями. Формикарий располагался на рабочем столе, и мы изучали и другие аспекты поведения муравьев, в том числе используя видеосъемку. Муравьи быстро к этому привыкли и не реагировали массовой неспецифической мобилизацией на появление людей. Однако стоило внести в помещении кормушку, как сразу же начиналось движение групп по обоим мостикам. Запах раствора меда муравьи чувствовали с расстояния в несколько метров. При этом они находились в формикарии, закрытом ареной, с тремя небольшими отверстиями. Очевидно, что и в первой серии опытов запах также играл определенную роль в активации муравьев, стимулируя их к поиску пищи.
В этом варианте опыта было всего 7 предъявлений.
Опыт 3. В последней серии опытов мы исключили возможность ориентации муравьев также и по запаховому сигналу. Кормушку вносили в помещение с муравьями в банке с плотно закрытой крышкой. Сначала подавался световой сигнал, затем, после выхода первых рабочих на арену по соответствующему мостику, кормушку выставляли на арену.
При первом предъявлении стимула (освещения) в этой серии опытов муравьи не вышли на мостик вообще. Через 10 мин сигнал повторили. Первый муравей появился на арене только после четвертого повторения сигнала. Таким образом, стало ясно, что у них вообще не сформировалась связь светового сигнала с кормушкой. На световой сигнал они никак не реагировали, первый вышедший муравей был разведчиком и демонстрировал явную поисковую активность, круговые и зигзагообразные движения и т.д. Впрочем, уже к третьему предъявлению муравьи уверенно, группами выходили на арену сразу после подачи светового сигнала.
Но тут выяснилась другая любопытная подробность. Оказалось, муравьи не воспринимали постоянное и прерывистое освещение как разные сигналы. И в первые несколько предъявлений фуражиры выходили группами на оба мостика сразу. Одна группа, естественно, находила кормушку, а вторая начинала обследовать пустую часть арены в поисках корма. При этом муравьи второй группы время от времени натыкались на кормушку у другого мостика.
Далее произошла некоторая оптимизация поведения фуражиров. Муравьи поняли, что кормушку каждый раз выставляют около одного мостика, и стали пытаться угадать следующее местоположение. Это произошло уже к пятому-седьмому предъявлению. Группы муравьев стали дежурить на поверхности субстрата вблизи мостика, на котором было предыдущее предъявление кормушки, а иногда и на самом мостике. Разумеется, одиночные муравьи дежурили и на другом мостике и тоже выходили на арену. Если случалось так, что кормушку выставляли возле того же мостика, что и в прошлый раз, то наблюдалось характерное быстрое нарастание численности фуражиров на кормушке (рис. 3, 1). Если же муравьи не угадывали, и кормушку ставили к другому мостику, то группы фуражиров постепенно перемещались к нему. Нарастание численности фуражиров на кормушке было медленным, почти линейным (рис. 3, 2). Такая реакция – искать корм там, где он был в предыдущих пробах, – характерна для всех животных и проявляется в различных тестах (Зорина, Полетаева, 2001).
Рис. 3.
Характер накопления рабочих муравьев на кормушке в третьей серии опытов: 1 – предъявление № 14 (“угадали”), 2 – предъявление № 19 (“не угадали”).
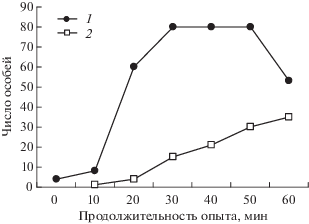
Это продолжалось довольно долго, до 20-го предъявления. После этого, без каких бы то ни было видимых внешних проявлений, муравьи внезапно поняли, что от них требуется. И далее, в 8 последующих предъявлениях всегда массово выходили на нужный мостик после соответствующего сигнала (как на рис. 3, 1). На этом наш опыт был закончен, поскольку муравьи очевидно научились различать непрерывное и прерывистое освещение и выходить на соответствующий мостик по сигналу.
В заключение следует отметить еще один эффект научения, который мы специально не вызывали, но который, тем не менее проявился практически во всех вариантах опыта. На кривых накопления фуражиров на кормушке (рис. 2, 1 и 3, 1) хорошо видно, что на последних 10 мин происходит снижение количества муравьев, иногда довольно значительное. Это означает, что фуражиры сами уходят с кормушки в конце опыта, чтобы не подвергаться процедуре удаления с кормушки кисточкой, безвредной, но, видимо, неприятной для муравьев, судя по их возбуждению и попыткам атаки. Таким образом, муравьи демонстрируют нам эффект научения по времени, достаточно точно отмеряя промежуток времени кормления в 1 ч, и начиная покидать кормушку по его истечении. В других опытах, где мы оставляли кормушку на более длительное время, такого эффекта не наблюдалось.
ОБСУЖДЕНИЕ
Задача, которую мы ставили перед муравьями, относится к дифференцировочным реакциям (Зорина, Полетаева, 2001). В простейшем случае животное должно реагировать на один стимул и не реагировать на другой, в нашем же случае задача была сложнее – муравьям необходимо было реагировать на оба стимула различным образом. Муравьи оказались способны различать непрерывный и прерывистый световые сигналы и реагировать на них соответствующим образом. При этом световой сигнал был пространственно не связан с пищевым подкреплением, что дает нам основания предполагать, что произошла определенная консолидация информации на уровне семьи. Во всяком случае, продолжение исследований в этом направлении представляется весьма перспективным.
Кроме того, муравьи продемонстрировали еще целый ряд интересных поведенческих реакций. Так, выяснилось, что муравьи Myrmica оказались способны ориентироваться по картине условного “неба” над головой, а также воспринимать запахи с расстояния в несколько метров. Были отмечены эффекты оптимизации поиска кормушки, когда муравьи сначала массово выходили на оба мостика, затем пытались искать пищу там, где она появлялась в предыдущий раз. Ранее оптимизацию фуражировки описывал Длусский (1981), который показал возможность глубоких перестроек системы мобилизации у мирмик в зависимости от условий опыта. В нашем случае таких существенных перестроек не наблюдалось, оптимизация заключалась лишь в сокращении времени обнаружения кормушки. Фактически это – одно из проявлений муравьиного алгоритма (Dorigo, 1992), когда система (семья), состоящая из многих однородных элементов (рабочих особей), имеет возможность протестировать все варианты решения задачи (поиск корма) практически одновременно и выбрать оптимальный. Наконец, мы наблюдали явление, сходное с инсайтом, описанным для позвоночных животных. При исключении простых для восприятия муравьев запаховых и зрительных стимулов, после серии проб и ошибок, навык был продемонстрирован в готовом виде, без постепенного нарастания правильных ответов.
В заключение следует отметить, что исследование процесса научения у муравьев позволяет получить обширные сведения об особенностях их восприятия, существенных для них стимулах, когнитивных способностях. Такие исследования представляются весьма перспективными, в том числе и в полевых условиях, где нет ограничений, присущих лабораторным опытам.
Список литературы
Дашевский Б.А., Карась А.Я., Удалова Г.П., 1989. Поведенческая пластичность муравьев Myrmica rubra при обучении в многоальтернативном симметричном лабиринте // Журнал высшей нервной деятельности. Т. 39. № 1. С 81–89.
Длусский Г.М., 1981. Принципы коммуникации у муравьев // Чтения памяти Н.А. Холодковского: Докл. на 33-м ежегодном чтении. Л.: Наука. С. 3–33.
Длусский Г.М., Волцит О.В., Сулханов А.В., 1978. Организация групповой фуражировки у муравьев рода Myrmica // Зоологический журнал. Т. 57. № 1. С. 65–77.
Зорина З.А., Полетаева И.И., 2001. Зоопсихология: Элементарное мышление животных. М.: Аспект Пресс. 320 с.
Кауль Р.М., 1985. Некоторые особенности оптической ориентации Formica rufa (Hymenoptera, Formicidae) // Зоологический журнал. Т. 64. № 12. С. 1823–1828.
Кауль Р.М., Коптева Г.А., 1982. Ночная ориентация муравьев Formica rufa (Hymenoptera, Formicidae) при движении на тропах // Зоологический журнал. Т. 61. № 9. С. 1351–1358.
Карась А.Я., Удалова Г.П., Загораева Е.В., 1986. Роль мотивации при обучении муравья Myrmica rubra в многоальтернативном лабиринте // Вестник ЛГУ. Сер. 3. Вып. 4. С. 43–52.
Лакин Г.Ф., 1990. Биометрия. М.: Высшая школа. 352 с.
Новгородова Т.А., 2006. Экспериментальное исследование передачи информации у лугового муравья (Formica pratensis, Hymenoptera, Formicidae) с помощью лабиринта «бинарное дерево» // Зоологический журнал. Т. 85. № 4. С. 493–499.
Плеханов Г.Ф., Кауль Р.М., 1976. Выработка условного пищевого рефлекса у муравьев Formica rufa на цвет тропы // Зоологический журнал. Т. 55. № 10. С. 1573–1575.
Путятина Т.С., 2007. Выбор стратегии фуражировки как механизм сосуществования видов Myrmica (Hymenoptera, Formicidae) в многовидовой ассоциации муравьев // Зоологический журнал. Т. 86. № 6. С. 701–708.
Резникова Ж.И., Пантелеева С.Н., Яковлев И.К., 2008. Гипотеза распределенного социального обучения и адаптивные возможности популяций: экспериментальные исследования на примере муравьев // Вестник ВОГиС. Т. 12. № 1/2. С. 97–111.
Резникова Ж.И., Рябко Б.Я., 1990. Теоретико-информационный анализ “языка” муравьев // Журнал общей биологии. Т. 51. № 5. С. 601–609.
Резникова Ж.И., Рябко Б.Я., 1999. Экспериментальные исследования способности муравьев к сложению и вычитанию небольших чисел // Журнал высшей нервной деятельности. Т. 49. № 1. С. 12–21.
Удалова Г.П., Жуковская М.И., Карась А.Я., 1991. Способность муравьев Myrmica rubra к множественным переделкам лабиринтного навыка // Журнал высшей нервной деятельности. Т. 41. № 6. С 1154–1162.
Cammaerts M.C., 2012. The visual perception of the ant Myrmica ruginodis (Hymenoptera: Formicidae) // Biologia. V. 67, Iss. 6. P. 1165–1174.
Dorigo M., 1992. Optimization, Learning and Natural Algorithms, PhD thesis, Politecnico di Milano, Italie.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал