

Зоологический журнал, 2019, T. 98, № 1, стр. 89-96
Развитие основных типов социальных взаимодействий детенышей евразийской рыси (Lynx lynx) в период раннего постнатального онтогенеза
А. А. Глухова 1, *, С. В. Найденко 2, **
1 Московский зоопарк,
123242 Москва, Россия
2 Институт проблем экологии и эволюции им. А.Н. Северцова РАН,
119071 Москва, Россия
* E-mail: allachagaeva@yandex.ru
** E-mail: snaidenko@mail.ru
Поступила в редакцию 01.03.2016
После доработки 25.04.2018
Принята к публикации 9.03.2018
Аннотация
Рассматриваются формирование и развитие конкурентных и игровых взаимодействий детенышей евразийской рыси в течение первого месяца их жизни, а также связь этих взаимодействий с размером выводка и полом котят. Установлено время возникновения данных типов социальных взаимодействий. Обнаружена отрицательная связь возраста, когда проявляется конкуренция, с размером выводка, в котором растут рысята. Показано, что игровое поведение в рассматриваемый период онтогенеза не зависит от размера выводка и пола детенышей.
Для незрелорождающихся видов млекопитающих характерно появление потомства в замкнутом пространстве убежища. Для детеныша, находящегося в таких условиях, существует только два возможных направления для осуществления первых социальных взаимодействий. В первую очередь, это взаимодействия с матерью, которые являются первым и самым ранним социальным фактором, действующим на детеныша (Крученкова, 2009; Maestripieri, Mateo, 2009). Другим возможным направлением ранних социальных контактов является взаимодействие с однопометниками. По мере взросления детеныши одного выводка начинают все более активно взаимодействовать между собой, постепенно формируя иерархическую структуру группы. Социальные контакты между детенышами одного возраста в раннем онтогенезе млекопитающих представлены, прежде всего, конкурентными (обзор Drummond, 2006; Hudson, Trillmich, 2008) и игровыми (обзор Spinka et al., 2001) взаимодействиями.
Преимущество особи в конкурентных взаимодействиях является показателем ее иерархического статуса. Для экспериментального установления иерархической структуры группы исследователи часто ориентируются на конкурентные взаимодействия за ограниченные ресурсы (пища, вода, территория), а в качестве показателя используется частота агрессивных взаимодействий (Lanctot, Best, 2000; Drummond, 2006).
Детеныши млекопитающих вступают в конкурентные отношения друг с другом с самого раннего возраста. Они конкурируют за молоко матери – единственный источник пищи в данном возрасте. Стратегии этой конкуренции могут быть различны у разных видов млекопитающих и зависеть от особенностей поведения самок (McBride, 1963; Hartsock et al., 1976; Hudson et al., 1995, 2002). Уже на самых ранних этапах развития потомства можно наблюдать постепенное становление иерархических отношений в выводке, которое у многих видов млекопитающих выражается в выборе (захвате) наиболее предпочитаемой пары сосков матери (Ewer, 1959; McBride, 1963; McGuire, 1998). Детеныши способны защищать предпочитаемый, лучший с их точки зрения сосок (домашняя кошка (Felis catus) (Ewer, 1959), домашняя свинья (Sus scrofa) (Scheel et al.,1977; Fraser, Jones, 1975), даманы (Procavia johnstoni и Heterohyrax brucei) (Hoeck 1977)). Для количественной оценки данного явления, как правило, используются число драк за определенный сосок или пару сосков и число побед в них детеныша, предпочитающего этот сосок (пару сосков) (домашняя кошка (Ewer, 1959), домашняя свинья (Scheel et al., 1977), пума (Puma concolor) (Pfeifer, 1980)).
Вторым типом ранних социальных контактов детенышей являются игровые взаимодействия. Они занимают очень важное место в онтогенезе детенышей млекопитающих и привлекают внимание ученых не одно десятилетие (Baldwin, Baldwin, 1974; Bekoff 1974; Caro, 1995; Spinka et al., 2001; Way, 2007; Paulos et al., 2010). Несмотря на различия во взглядах на функциональное значение и социальные аспекты игры, все авторы, так или иначе, сходятся во мнении о важности данных взаимодействий для выживания и нормального развития детенышей. Игровое поведение может быть механизмом развития двигательных и когнитивных навыков. Возможно, в процессе игры происходит “двигательное обучение”, которое положительно сказывается на моторном поведении: развитие выносливости и силы, увеличение двигательных функций организма, развитие мускулатуры, костей, нервной и сердечно-сосудистой систем (Byers, Walker, 1995). Было показано, что наличие опыта социальной игры у детенышей приводит к увеличению гибкости индивидуального поведенческого репертуара (Miller, 1973), а также к более тонкому восприятию социальных сигналов, на которые животное может соответствующим образом реагировать (Baldwin, Baldwin, 1974).
Игровая активность животных может зависеть от разных факторов. Так, интенсивность и частота игровых взаимодействий существенно меняются с возрастом. По мере физического развития детенышей частота игровых взаимодействий постепенно увеличивается (Найденко, 1997; Алексеева и др., 2014). Однако далее происходит постепенное снижение игровой активности (Gomendio, 1988; Panksepp, 1981). В ряде работ были обнаружены различия в интенсивности игры между самцами и самками (Berger, 1980; Dobao et al., 1985; Ward et al., 2008).
Модельным видом для исследования поведения и развития детенышей у представителей семейства кошачьих (Felidae) традиционно является домашняя кошка (Ewer, 1959, 1961; Martin, 1984, 1986; Deag et al., 1987; Hudson et al., 2009). Данный вид наиболее удобен для изучения в связи с доступностью животных и, как правило, не агрессивным их отношением к людям. Однако распространение результатов, полученных для домашней кошки, на всех представителей семейства невозможно. Это связно с большим количеством различий в образе жизни, социальной структуре и условиях обитания разных видов. О поведении детенышей диких представителей семейства кошачьих в природе, особенно в первые недели после родов, известно мало, что объясняется скрытным образом жизни этих хищников и, соответственно, невозможностью их подробного наблюдения в естественных условиях. Для евразийской рыси (Lynx lynx L.) на сегодняшний день нет данных, касающихся особенностей становления социальных отношений в пределах выводка в первые дни жизни потомства, мало изучены пути формирования сложного поведения этих животных, факторы, влияющие на социальное развитие котят. Вероятно, иерархический статус котенка в выводке в раннем онтогенезе может быть крайне важным фактором в формировании дальнейшего жизненного пути детеныша.
Цель настоящей работы – описать динамику развития ранних социальных взаимодействий детенышей евразийской рыси в течение первого месяца их жизни. Для этого было необходимо исследовать развитие конкурентных и игровых взаимодействий детенышей в течение первых четырех недель их жизни и оценить связь ранних типов взаимодействий с размером выводка и полом котят.
МАТЕРИАЛЫ И МЕТОДЫ
Условия содержания животных
Работа была проведена на НЭБ “Черноголовка” ИПЭЭ РАН в период с 2005 по 2010 гг. Объектом исследования были 16 выводков от 6 самок европейского подвида рыси L. l. lynx (2 выводка с 4 котятами, 7 выводков с 3 котятами, 6 выводков с 2 котятами и один выводок с одним котенком). Выводки, родившиеся на НЭБ “Черноголовка” в 2005–2010 гг. содержали в вольерах 74 м2. Каждый выводок с самкой находился в отдельной вольере, лишенной прямостоячих деревьев, а на почве произрастали различные злаки. Каждая вольера имела высоту 4.5 м, верхняя часть стенки (1 м) была покрыта листами оцинкованного железа для предотвращения побегов животных. Кроме того, каждая самка имела доступ к примыкающей к вольере закрытой клетке площадью 8 м2. Беременных самок кормили один раз в день, шесть дней в неделю, один день – голодный. Основу рациона составляли цельные тушки кур. Ежедневно животные получали около 1 кг мяса, водой же были обеспечены ad libitum. При появлении детенышей суточный рацион увеличивали примерно в 1.5 раза, зверей кормили 7 раз в неделю. Самки рожали в деревянных домиках (100–130 × 80 × 80 см), которые они использовали в качестве укрытий для себя и своего выводка в первые недели после родов.
Условия проведения наблюдений
В течение первых 3–4 недель жизни котят самки с детенышами практически не покидали убежища. Поэтому внутри каждого выводкового домика устанавливали камеру наблюдения (KPC-190SWX, Корея), при помощи которой осуществляли видеосъемку животных. С 2005 по 2007 год видеозапись вели на видеомагнитофон (JVC HR-J870), в 2008–2009 гг. на 4-хканальный видеорегистратор (AT-200 4W + VGA), в 2010 г. запись вели на компьютер. Для подсветки внутри выводковых домиков были установлены лампы мощностью 15 Вт. Каждого детеныша до проведения видеосъемки метили индивидуально, начиная с возраста 1–4 дня, выстригая шерсть на отдельных частях тела (круп, спина, загривок, голова).
Регистрация поведения животных
В работе были использованы данные 1221 ч видеонаблюдений. Видеозапись поведения животных проводили один раз в неделю в течение 24 ч в первые четыре недели жизни котят (за начальную точку (0) были приняты 1–4-й дни жизни детенышей, точка 1 – возраст одна неделя (6–8 дней)…, 4 – возраст четыре недели). Для анализа полученного видеоизображения использовали метод непрерывной регистрации данных (Martin, Bateson, 1993). Все время наблюдений было разделено на часовые интервалы. Мы регистрировали число форм поведения, осуществляемых рысятами внутри выводкового домика. При обработке видеоматериала учитывали только время, когда животные находились в пределах обзора видеокамеры.
В работе были использованы данные наблюдений за выводками с разным числом детенышей. Средний размер выводка евразийской рыси составляет по разным источникам от 2 до 2.5 детенышей (Стоянова, Сваринская, 1964; Гептнер, Слудский, 1972; Kvam, 1991; Найденко, 2005). Мендл (Mendl, 1988) обнаружил существенные различия в онтогенезе детенышей млекопитающих из выводков, где число детенышей было больше среднего для вида, и выводков, где это число было меньше среднего для вида. Мы также разделили выводки на “маленькие” с 1–2 котятами (число котят меньше среднего) и “большие” с 3–4 детенышами (число детенышей больше среднего).
Пол детенышей определяли в первые 4 дня их жизни. Если несколько котят в выводке были одного пола, то в качестве сравниваемого значения при анализе половых различий сначала рассчитывали средние показатели для этих котят. Выводки, где все котята были одного пола, а также выводок с одним детенышем не были включены в анализ половых различий в развитии детенышей. Всего для сравнений детенышей по полу были использованы данные 471 ч наблюдений за 11 котятами-самцами и 11 котятами-самками из семи выводков (шесть выводков с тремя котятами, один – с четырьмя детенышами).
При исследовании конкурентных взаимодействий детенышей мы оценивали количество актов борьбы за сосок матери, осуществляемые котятами у разных пар сосков. При этом регистрировали общее количество элементов борьбы, инициированных каждым котенком, количество побед каждого котенка в борьбе за сосок (независимо от того, кто был инициатором), пару сосков, на которой отмечены взаимодействия котят. Рысь имеет четыре пары сосков, но пара грудных сосков не используется детенышами (Чагаева, 2012), поэтому выделяли взаимодействия на передней (2-я пара), средней (3-я пара) и задней (4-я паховая пара) парах сосков.
Регистрируемые элементы поведения
Борьба за сосок (число отмеченных случаев борьбы в выводке) – детеныши пытаются оттолкнуть друг друга от определенного соска. В каждом случае отмечали котенка, инициировавшего борьбу за сосок.
Победа в борьбе за сосок (количество).
Социальная игра (количество взаимодействий).
Мы выделяли три типа игры игра с матерью, игра с однопометниками (оба типа – социальные игры) и самостоятельная игра (Найденко, 1997). Игровые взаимодействия с партнером включают у рысей приглашение, напрыгивание, обхват, борьбу, игру с частями тела партнера (лапами, хвостом), погоню, скрадывание, затаивание (Найденко, 1997; Алексеева и др., 2014), кусание (один котенок кусает другого или самку; этот игровой элемент наблюдается, как правило, уже на второй–третьей неделе жизни детенышей, когда активное игровое взаимодействие еще невозможно из-за ограниченности моторных функций организма). Интенсивное развитие игрового поведения наблюдается уже после месячного возраста котят, достигая пика примерно в трехмесячном возрасте (Алексеева и др., 2014). В настоящей работе этот период онтогенеза рысят не рассматривался. Все социальные игровые взаимодействия в первые четыре недели развития котят сводились к покусываниям однопометников или матери.
Самостоятельная игра – игра с предметом (редко наблюдается в течение первого месяца жизни котят), валяние (находясь на спине, котенок энергично “перекатывается” с боку на бок, мышцы конечностей при этом находятся в тонусе), лазание, бег.
Игровые взаимодействия регистрировали по принципу наличия/отсутствия без учета их длительности. В парных взаимодействиях животных различали инициатора (того, кто начинал взаимодействия) и объект (того, на кого были направлены взаимодействия) действия.
Статистический анализ
Чтобы избежать влияния фактора “выводок” на результаты исследований и псевдорепликации данных, связанных с размером выводка, каждый показатель (кроме оговоренных ниже) рассчитывали сначала как средний для всех котят выводка и лишь затем его использовали в анализе. При анализе половых различий внутри выводка в частотах поведения сравнивали критерием Вилкоксона для сопряженных пар значения для самок и самцов в этом выводке, рассчитанные как средние значения для детенышей каждого пола (то есть если в выводке был один самец и две самки, то сначала рассчитывали средние значения для самок, а затем этот показатель использовали в анализе).
Для проведения статистического анализа использовали программы Microsoft Excel и Statistica 6.0. Данные на графиках представлены в виде средних значений и стандартной ошибки среднего (M + SE). При статистической обработке данных мы использовали только непараметрические критерии.
Для анализа изменений параметров с возрастом детенышей мы использовали Критерий Вилкоксона для сопряженных пар (Wilcoxon Matched Pairs Test). Данный критерий был также использован для статистических сравнений сопряженных измерений параметров развития (пол детенышей).
Для сравнения оцениваемых параметров в неравных и несвязанных выборках (в выводках разного размера) мы использовали U-критерий Манна–Уитни (Mann–Whitney U test).
РЕЗУЛЬТАТЫ
Конкурентные взаимодействия
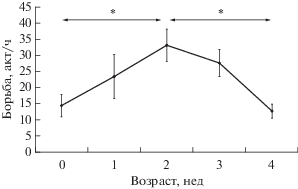
Было прослежено количество инициированных актов борьбы за сосок в течение первого месяца жизни детенышей (рис. 1). Уже в первые дни жизни котята вступают в конкурентную борьбу за соски матери, однако число таких взаимодействий не слишком велико (в среднем 14.4 ± 3.5 актов на час для котенка). К двухнедельному возрасту котят число элементов борьбы достоверно растет (тест Вилкоксона для сопряженных пар: Z = 2.5, n = 12, p < 0.05), достигая своего максимального значения (в среднем 33.1 ± 5.0). Далее к концу первого месяца жизни детенышей происходит постепенное уменьшение числа подобных взаимодействия (Z = 3.7, n = 24, p < 0.05). В возрасте четырех недель наблюдается наиболее низкая для первого месяца развития котят частота борьбы за соски (в среднем 12.7 ± 2.2).
Рис. 1.
Количество инициированных детенышами актов борьбы за соски в разном возрасте. Тест Вилкоксона для сопряженных пар: * p < 0.05.
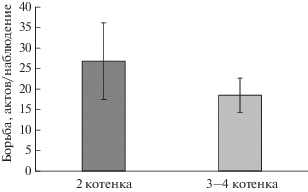
Мы проанализировали связь числа инициированных актов борьбы за соски и размера выводка (рис. 2). Детеныши из маленьких выводков несколько более активно инициировали борьбу за соски матери в течение первого месяца жизни, однако эти различия были недостоверны (U-критерий Манна–Уитни: Z = 0.35, ns, n1 = 7, n2 = 8).
Рис. 2.
Количество инициированных детенышами актов борьбы за соски в выводках разного размера. U-критерий Манна–Уитни: * p < 0.05.
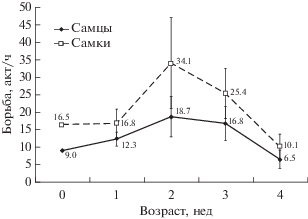
Не было выявлено никаких различий в частоте инициирования актов борьбы за соски матери детенышами разного пола (тест Вилкоксона для сопряженных пар: Z = 1.2, n = 7, ns), несмотря на наличие некоторой тенденции детенышей-самок более активно инициировать борьбу за соски матери (рис. 3).
Игровые взаимодействия
Хотя проявление игровых взаимодействий на ранних этапах онтогенеза наблюдается достаточно редко, впервые элементы игры начинают появляться уже в возрасте 1–2 недель жизни котят (рис. 4). При этом в возрасте одной недели элементы игры единичны.
Рис. 4.
Игровое поведение детенышей в течение первого месяца жизни. Тест Вилкоксона для сопряженных пар: * p < 0.05.
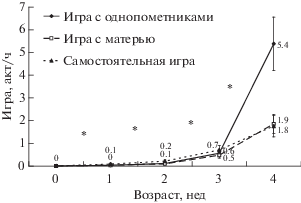
В первые дни жизни котят элементы игры в их поведении полностью отсутствуют. Элементы социальной игры (с однопометниками и матерью) начинают появляться к двухнедельному возрасту (рис. 4). Далее в течение первого месяца жизни котят число игровых элементов быстро растет, увеличиваясь достоверно каждую неделю (для игры с однопометниками: Z = 2.7–2.8, n = 9–11, p < 0.05; для игры с матерью: Z = 2.4–3.0, n = 9–12, p < 0.05). Элементы самостоятельной игры отмечали уже в возрасте одной-двух недель (рис. 4), хотя они и были представлены только одной разновидностью, характерной для раннего периода развития. Количество наблюдаемых элементов самостоятельной игры также достоверно увеличивалось каждую неделю в течение первого месяца жизни котят (Z = 2.0–2.5, n = 9–12, p < 0.05).
Можно было бы ожидать, что в выводках большого размера, при наличии большего числа потенциальных партнеров для игры, детеныши будут проявлять элементы игрового поведения чаще, чем в более маленьких выводках. Для проверки наличия связи между игровым поведением котят и размером выводка мы сравнили среднее числоэлементов игры, проявляемых детенышами из маленьких (1–2 котенка, для игры с однопометниками только выводки с двумя котятами) и больших (3–4 котенка) выводков в разном возрасте. Не выявлено различий в проявлении игрового поведения ни по одному из рассмотренных типов игры между детенышами из выводков разного размера (U-критерий Манна-Уитни: для игры с однопометниками: Z = 0.1, ns, n1 = 7, n2 = 9; для игры с матерью: Z = 0.3, ns, n1 = 7, n2 = 9; для самостоятельной игры: Z = 0.7, ns, n1 = 7, n2 = 9).
Также не было обнаружено различий в игровой активности детенышей разного пола ни по одному типу игрового поведения в течение всего рассматриваемого периода (тест Вилкоксона для сопряженных пар: для игры с однопометниками: Z = 0.3–0.7, n = 5–6, ns; для игры с матерью: Z = = 1.1–1.8, n = 5–6, ns; для самостоятельной игры: Z = 1.2–1.8, n = 5–6, ns).
ОБСУЖДЕНИЕ
Конкурентные взаимодействия
С первых часов жизни детеныши многих млекопитающих вынуждены вступать в конкурентные взаимодействия с однопометниками, целью которых является получение единственного доступного им в этом возрасте энергетического ресурса – материнского молока. У видов, у которых время кормления не разбито на короткие периоды, определяемые самкой (кролики (Hudson et al., 1995, 2002)), то есть не имеет жестких ограничений, эти взаимодействия, тем не менее, могут носить достаточно агрессивный характер (свиньи (Algers, 1993)). Исследования особенностей ранних конкурентных взаимодействий были проведены для многих видов млекопитающих (Tsai, 1931; Ewer, 1959; Fraser, Thompson, 1986; Ota et al., 1991; Tomaszycki et al., 1998; McGuire, 1998; McGuire, Sullivan, 2001). Для ряда видов было обнаружено неравномерное использование детенышами разных пар сосков матери, иными словами, было установлено наличие определенного порядка сосания внутри выводков. Это было показано, например, для сосновых (Microtus pinetorum) и прерийных полевок (M. ochrogaster) (McGuire, Sullivan, 2001; McGuire, 1998), свиней (Hemsworth et al., 1976; Scheel et al., 1977), японских макак (Macaca fuscata) (Ota et al., 1991), макак-резусов (Macaca mulatta) (Tomaszycki et al., 1998), домашних кошек (Ewer, 1959; Hudson et al., 2009), снежного барса (Uncia uncia) (McVittie, 1978), пумы (Pfeifer, 1980) и евразийской рыси (Чагаева, 2012; Glukhova, Naidenko, 2014).
Нами было обнаружено, что общее количество актов борьбы за соски в течение первых двух недель жизни котят постепенно увеличивалось, достигая максимума в возрасте двух недель, и постепенно уменьшалось к четвертой неделе развития. Увеличение частоты борьбы к возрасту двух недель мы связываем с постепенным взрослением котят, формированием некоторых локомоторных навыков. В возрасте двух недель у рысят открываются глаза и слуховые проходы, они становятся более самостоятельными (Найденко, 2005). Ранее было показано неравномерное использование рысятами разных пар сосков матери (Чагаева, Найденко, 2012; Glukhova, Naidenko, 2014). Предпочтения формировались уже в первые дни жизни детенышей, и к четвертой неделе жизни происходило усиление данных предпочтений. Уменьшение частоты борьбы к концу первого месяца жизни детенышей мы связываем с постепенным усилением предпочтений определенных пар сосков. Таким образом, в течение первых четырех недель развития у рысят происходит постепенное упорядочивание использования определенных пар сосков матери, в выводке формируется определенный порядок сосания, что снижает необходимость в конкурентных взаимодействиях. Данный результат хорошо согласуется с предположением о том, что распределение детенышей по соскам матери позволяет снизить агрессивные взаимодействия однопометников при поиске соска (Ewer, 1959; McBride, 1963; Hartsock, Graves, 1976).
Нами было также обнаружено, что детеныши из больших выводков несколько реже инициируют борьбу за соски, чем котята из маленьких выводков. Можно предположить, что, несмотря на наличие наиболее предпочтительной пары сосков для рысят, ее доступность для конкретного детеныша определяется его силой и иерархическим статусом в выводке. В маленьких выводках (2 котенка) перед детенышами не встает проблемы нехватки молока и в дальнейшем возможного голода, что подтверждают и несколько более высокие темпы роста детеныша в двойнях (Naidenko, 2006), поэтому котята могут попытаться занять наиболее предпочтительное место, чаще инициируя борьбу за “лучшую” пару и не рискуя остаться голодными в случае поражения. В больших выводках (3–4 котенка) для детенышей оказывается более важным получить доступ к пище, независимо от пары сосков. В этом может играть роль, во-первых, ограниченное количество молока у матери; во-вторых, детенышей много, и они физически мешают друг другу получить доступ к соскам.
Игровые взаимодействия
Игровые элементы начинают проявляться в поведении детенышей евразийской рыси, начиная с возраста двух недель. Ранее наблюдения за развитием игрового поведения рысят проводили только после выхода детенышей из убежища (в возрасте 3–4 недель) (Найденко, 1997, 2005), что не позволяло обнаружить наличие игровых элементов в их поведении в более раннем возрасте.
Было выдвинуто предположение, что число элементов игры с однопометниками будет больше в выводках, где наблюдаются более быстрые темпы физического развития (Elwood, Broom, 1978). Так как физическое развитие обычно связано с числом детенышей в выводках (Mendl, 1988; Naidenko, 2006), вероятно существует непрямая связь размера выводка и интенсивности игры с однопометниками. Нами, однако, не было обнаружено никаких различий в проявлении элементов игрового поведения детенышами в течение первого месяца жизни в выводках разного размера. Ранее подобных различий также не было обнаружено для детенышей более старшего возраста (Найденко, 2005).
Детеныши разного пола у рысей не демонстрировали различий в проявлении игрового поведения, что показано и для домашней кошки (Caro, 1981). Вместе с тем, Каро (Caro, 1981) выявил достоверные половые различия в игре с однопометниками при сравнении выводков, состоящих только из детенышей одного пола (только самцов или только самок). В нашей работе выборка, к сожалению, не позволила нам провести такое сравнение (слишком мало выводков с котятами одного пола).
Таким образом, конкурентные взаимодействия у рысят наблюдаются уже в первые дни жизни. Частота их проявления непосредственно связана с возрастом: максимальна в двухнедельном возрасте и достоверно снижается в четвертой неделе развития. Степень проявления конкурентных взаимодействий у детенышей при кормлении молоком зависит от размера выводка: конкуренция выше в маленьких выводках. В возрасте одной-двух недель в поведении детенышей евразийской рыси начинают проявляться игровые элементы, однако, частота проявления игрового поведения в первый месяц жизни не связана с размером выводка и полом котят.
Список литературы
Алексеева Г.С., Антоневич А.Л., Ерофеева М.Н., Найденко С.В., 2014. Социальная игра в онтогенезе внутривыводковых отношений евразийской рыси (Lynx lynx) // Известия РАН. Серия биологическая. № 4. С. 382–390.
Гептнер В.Г., Слудский А.А., 1972. Млекопитающие Советского Союза. Т.2. Ч. 2. Хищные (Гиены и кошки). М.: Высшая школа. 551 с.
Крученкова Е.П., 2009. Материнское поведение млекопитающих. М.: КРАСАНД. 208 с.
Найденко С.В., 1997. Социальное поведение рыси и особенности его формирования в онтогенезе. Дис. … канд. биол. наук. ИПЭЭ им. А.Н. Северцова. М. 252 с.
Найденко С.В., 2005. Особенности размножения и постнатального развития Евразийской рыси. М.: Товарищество научных изданий КМК. 111 с.
Стоянова В.К., Сваринская В.Г., 1964. Наблюдения за рысями в Рижском зоосаде // Материалы Прибалтийской конф. Рига. С. 21–25
Чагаева А.А., 2012. Особенности внутривыводковых отношений евразийской рыси (Lynx lynx L.) в период раннего постнатального онтогенеза. Дис. … канд. биол. наук. ИПЭЭ им. А.Н. Северцова. Москва. 139 с.
Чагаева А.А., Найденко С.В., 2012. Материнское поведение евразийской рыси в период раннего постнатального онтогенеза детенышей // Известия РАН. Серия биологическая. № 1. С. 54–60.
Algers B., 1993. Nursing in pigs: communicating needs and distributing resources // Journal of Animal Science. V. 71. P. 2826–2831.
Baldwin J.D., Baldwin I.B., 1974. Exploration and Social Play in squirrel Monkeys (Saimiri) // American Zoologist. V. 14. P. 303–315.
Bekoff M., 1974. Social play and play-soliciting by infant Canids // American Zoologist. V. 14. P. 323–340.
Berger J., 1980. The ecology, structure and function of social play in Bighorn sheep (Ovis canadensis) // Journal of Zoology. V. 192. P. 531–542.
Byers J.A., Walker C., 1995. Refining the motor training hypotheses for the evolution of play // American Naturalist. V. 146. P. 25–40.
Caro T. M., 1981. Sex differences in the termination of social play in cats // Animal Behaviour. V. 29. № 1. P. 271–279.
Caro T.M., 1995. Short-term costs and correlates of play in cheetahs // Animal Behavior. V. 49. P. 333–345.
Deag J.M., Lawrence C.E., Manning A., 1987. The consequences of differences in litter size for the nursing cat and her kittens // Journal of Zoology (London). V. 213. P. 153–179.
Dobao M.T., Rodriganez J., Silio L., 1985. Choice of companions in social play in piglets // Applied Animal Behavior Sciences. V. 13. P. 259–266.
Drummond H., 2006. Dominance in vertebrate broods and litters // Quarterly Review of Biology. V. 81. № 1. P. 3–32.
Elwood R.W., Broom D.M., 1978. The influence of litter size and parental behaviour on the development of Mongolian gerbil pups // Animal Behaviour. V. 26. P. 438–454.
Ewer R.F., 1959. Suckling behaviour in kittens // Behaviour. V. 15. P. 147–160.
Ewer R.F., 1961. Further observations on suckling behaviour in kittens, together with some general considerations of the interrelations of innate and acquired responses // Behaviour. V. 17. № 4. P. 247–260.
Fraser D., Jones R.M., 1975. The “teat order” of suckling pigs. I. Relation to birth weight and subsequent growth // Journal of Agricultural Science. V. 84. P. 387–391.
Fraser D., Thompson B.K., 1986. Variation in piglet weights: relationship to suckling behaviour, parity number and farrowing crate design // Canadian Journal of Animal Science. V. 66. P. 33–46.
Glukhova A., Naidenko S., 2014. Suckling behavior in Eurasian lynx (Lynx lynx L.) cubs: characteristics and correlation with competitive interactions // Zoo biology. V. 33. P. 388–393.
Gomendio M., 1988. The development of different types of play in gazelles: implications for the nature and functions of play // Animal Behaviour. V. 36. P. 825–837.
Hartsock T.G., Graves H.B., 1976. Neonatal behaviour and utrition-related mortality in domestic swine // Journal of Animal Science. V. 42. P. 235–241.
Hemsworth P.H., Winfield C.G., Mullaney P.D., 1976. A study of the development of the teat order in piglets // Applied Animal Ethology. V. 2. P. 225–233.
Hoeck H.N., 1977. “Teat order” in Hyrax (Procavia johnstoni and Heterohyrax brucei) // Zeitschrift fur Säugetierkunde. V. 42. P. 112–115.
Hudson R., Muller A., Kennedy G., 1995. Parturition in the rabbit is compromised by daytime nursing: the role of oxitocin // Biology of reproduction. V. 53. P. 519–524.
Hudson R., Labra-Cardero D., Mendoza-Soylovna A., 2002. Sucking, not milk, is important for the rapid learning of nipple-search orders in newborn rabbits // Developmental Psychobiology. V. 41. № 3. P. 226–235.
Hudson R., Trillmich F., 2008. Sibling competition and cooperation in mammals: challenges, developments and prospects // Behavioral Ecology and Sociobiology. V. 62. P. 299–307.
Hudson R., Raihani G., Gonza’lez D., Bautista A., Distel H., 2009. Nipple preferences and contests in suckling kittens of the domestic cat are unrelated to presumed nipple quality // Developmental Psychobiology. V. 51. P. 322–332.
Kvam T., 1991. Reproduction in the European lynx, Lynx lynx // Zeitschrift fur Säugetierkunde. V. 56. P. 146–158.
Lanctot R.B., Best L.B., 2000. Comparison of methods for determinig dominance rank in male and female prarie voles (Microtus ochrogaster) // Journal of Mammology. V. 81. № 3. P. 734–745.
Maestripieri D., Mateo J.M., 2009. Maternal effects in mammals. Chicago: The University of Chicago Press. 352 P.
Martin P., 1984. The time and energy costs of play behaviour in the cat // Zeitschrift fur Tierpsychologie. V. 64. P. 298–312.
Martin P., 1986. An experimental study of weaning in the domestic cat // Behaviour. V. 99. № 3-4. P. 221–229.
Martin P., Bateson P., 1993. Measuring behaviour: an introductory guide. 2nd ed. Cambridge, England: Cambridge Univ. Press. 238 p.
McBride G., 1963. The “teat order” and communication in young pigs // Animal Behaviour. V. 11. P. 53–56.
McGuire B., 1998. Suckling behavior of Prairie voles (Microtus ochrogaster) // Journal of Mammalogy. V. 79. № 4. P. 1184–1190.
McGuire B., Sullivan S., 2001. Suckling behavior of Pine voles (Microtus pinetorum) // Journal of Mammalogy. V. 82. № 3. P. 690–699.
McVittie R., 1978. Nursing behavior of snow leopard cubs // Appiied Animal Ethology. V. 4. P. 159–168.
Mendl M., 1988. The effects of litter size variation on mother-offspring relationships and behavioural and physical development in several mammalian species (principally rodents) // Journal of Zoology (London). V. 215. P. 15–34.
Miller S., 1973. Ends, means, and galumphing: some leitmotifs of play // American Anthropologist. V. 75. P. 87–98.
Naidenko S.V., 2006. Body mass dynamic in Eurasian lynx Lynx lynx kittens during lactation // Acta theriologica. V. 51. № 1. P. 91–98.
Ota K., Makino Y., Kimura M., Suzuki J., 1991. Lactation in the Japaneses monkey (Macaca fuscata): yield and composition of milk and nipple preference of young // Primates. V. 32. № 1. P. 35–48.
Panksepp J., 1981. The ontogeny of play in rats // Developmental Psychobiology. V. 14. № 4. P. 327–332.
Paulos R.D., Trone M., Kuczaj II S.A., 2010. Play in wild and captive Cetaceans // International Journal of Comparative Psychology. V. 23. P. 701–722.
Pfeifer Sh., 1980. Role of the nursing order in social development of mountain lion kittens // Developmental Psychobiology. V. 13. № 1. P. 47–53.
Scheel D.E., Graves H.B., Sherritt G.W., 1977. Nursing order, social dominance and growth in swine // Journal of Animal Science. V. 45. № 2. P. 210–229.
Spinka M., Newberry R.C., Bekoff M., 2001. Mammalian play: training for the unexpected // The Quarterly Review of Biology. V. 76. № 2. P. 141–168.
Tomaszycki M., Cline Ch., Griffin B., Maestripieri D., Hopkins W., 1998. Maternal cradling and infant nipple preferences in Rhesus monkeys (Macaca mulatta) // Developmental Psychobiology. V. 32. P. 305–312.
Tsai L.S., 1931. Sucking preference in nursing young rats // Journal of Comparative Psychology. V. 12. P. 251–256.
Ward C., Bauer E.B., Smuts B.B., 2008. Partner preferences and asymmetries in social play among domestic dog, Canis lupus familiaris, littermates // Animal Behaviour. V. 76. № 4. P. 1187–1199.
Way J., 2007. Social and play behavior in a wild eastern coyote, Canis latrans, pack // The Canadian Field-Naturalist. V. 121. P. 397–401.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал