

Зоологический журнал, 2019, T. 98, № 1, стр. 5-14
Новые виды щетинохвосток рода Allopsontus silv. (Archaeognatha, Machilidae) c Кавказа и из Таджикистана
В. Г. Каплин *
Всероссийский научно-исследовательский институт защиты растений
196608 Санкт-Петербург–Пушкин, Россия
* E-mail: ctenolepisma@mail.ru
Поступила в редакцию 31.12.2017
После доработки 11.07.2018
Принята к публикации 19.02.2018
Аннотация
Описаны два новых вида щетинохвосток семейства Machilidae (Allopsontus tyrnyauzi sp. n. и Allopsontus zinchenkoi sp. n.). Среди известных видов подрода Kaplinilis Mendes 1990, к которому относится A. tyrnyauzi sp. n., лишь у одного вида, A. bifarius (Wygodzinsky 1970), из Западной Монголии и Тувы самцы имеют короткие густые прижатые пигментированные щетинки на пятом–седьмом члениках нижнечелюстных и третьем членике нижнегубных щупиков. A. tyrnyauzi sp. n. отличается от A. bifarius меньшей относительной длиной линии контакта глаз, более расширенными парными глазками, прямым вершинным углом стернитов брюшка, более узкими сенсорными полями на передних бедрах самцов и меньшим числом члеников парамер. Второй описанный вид, A. zinchenkoi sp. n., относится к подроду Anisopsontus Mendes 1990 и отличается от других видов подрода цветом глаз, отношением ширины парного глазка к его длине, относительно более длинными грифельками брюшных кокситов, более крупными размерами сенсорных полей на передних бедрах самцов и сравнительно коротким яйцекладом у самок.
При обработке материалов, собранных в апреле и июле 2017 г. в Кабардино-Балкарии и Варзобском ущелье Таджикистана, выявлены два новых вида рода Allopsontus Silvestri 1911. Их описания приведены ниже. Типы новых видов хранятся в коллекции ЗИН РАН, С.-Петербург.
Allopsontus tyrnyauzi Kaplin sp. n. (рис. 1, 1–8)
Рис. 1.
Allopsontus tyrnyauzi sp. n. (голотип, самец): 1 – глаза и парные глазки, 2 – передняя нога, 3 – нижнечелюстной щупик, 4 – передняя часть верхней челюсти, 5 – нижнегубной щупик, 6 – стернит и кокситы VIII сегмента брюшка, 7 – X тергит брюшка, 8 – IX коксит брюшка и половой аппарат. Масштаб 0.1 мм.
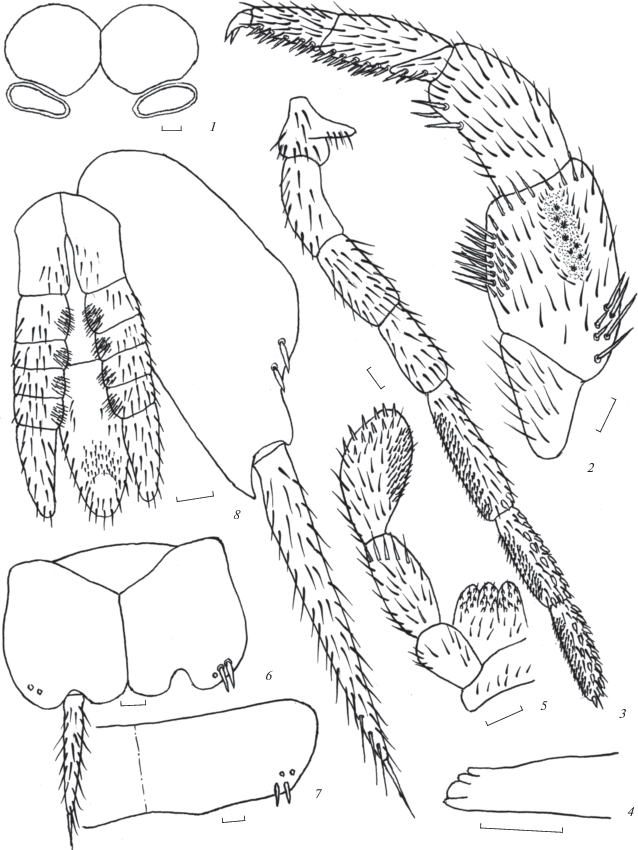
Материал. Голотип, ♂ (в препаратах), Кабардино-Балкария, окрестности г. Тырныауза, кустарниково-злаковая степь с барбарисом, 30.04. 2017. Паратип, 1 ♂, там же (В.Г. Каплин).
Описание. Самец. Длина тела 8.0–8.2; ширина 2.0–2.1 мм. Общая окраска буровато-коричневая, все тело с коричневым гиподермальным пигментом средней и сильной интенсивности. Наиболее интенсивно пигментированы лоб, бока наличника, верхняя губа, нижнечелюстные щупики, участки вокруг глаз, глазков и оснований усиков. Длина усиков 0.7–0.8 длины тела. Цепочки передней части жгутика усиков 5-члениковые с выраженной вторичной сегментацией у третьего и четвертого члеников от основания. Длина церок 3.1–3.2 мм, или 0.38–0.39 длины тела. Вершины церок с одним опорным шипом. Церки содержат 13 члеников. Первые 4–5 члеников церок от основания разделены не отчетливо, границы между остальными члениками четкие. Все членики, за исключением одного или двух вершинных, с двумя–тремя внутренне-боковыми опорными макрохетами в передней части, отсутствующими на хвостовой нити.
Глаза в спирте темные, одноцветные. Общая ширина глаз 0.81–0.85, длина – 0.40–0.42 мм. Отношение длины глаза к его ширине около 1.0. Линия контакта глаз составляет 0.28–0.30 их длины. Парные глазки сублатеральные по отношению к глазам, узкоовальные (рис. 1, 1), светло-коричневые с узким белым ободом. Их размеры около 0.29 × 0.10 мм. Расстояние между внутренними краями глазков 0.46–0.47, наружными – 1.02–1.04 общей ширины глаз.
Длина последнего (7-го) членика нижнечелюстных щупиков составляет 0.90 длины предпоследнего членика, а 5-й членик в 1.6 раза длиннее 4-го. Дорсальная поверхность 6–7-го члеников нижнечелюстных щупиков с 13–14, 5-го – с 3 бесцветными зубьевидными хетами с темными вершинами. 5–7-й членики нижнечелюстных и 3-й членик нижнегубных щупиков с мелкими прижатыми пигментированными щетинками (рис. 1, 3, 5). Нижнечелюстные и нижнегубные щупики без длинных тонких извилистых щетинок. Последний членик нижнегубных щупиков овально-треугольный. Его длина в 1.9 раза больше ширины. Вершины верхних челюстей 4-лопастные (рис. 1, 4).
Передние бедра и голени расширенные (табл. 1). Передние бедра в базальной части с 4–6 сравнительно длинными дорсальными макрохетами. Наиболее короткие ноги – средние. Отношение длины апикального членика задней лапки к ее общей длине около 0.41. Вентральная поверхность лапок и голеней с двумя рядами игловидных пигментированных щетинок, распределение которых приведено в табл. 2. Передние бедра с узкими сенсорными полями, включающими 9–11 розетковидных сенсилл. Сенсиллы сенсорного поля расположены в один неровный ряд (рис. 1, 2). Ширина сенсорного поля составляет 0.15 ширины бедра, его длина – 0.43 длины бедра. Сенсорное поле не доходит до середины заднего основания бедра на 0.47, его переднего основания – на 0.10 длины бедра. Грифельки на тазиках средних и задних ног, их длина около 0.5 мм. Ноги без длинных тонких извилистых щетинок.
Таблица 1.
Отношение длины частей ног к их ширине у самца Allopsontus tyrnyauzi sp. n.
| Часть ноги | Пара ног | ||
|---|---|---|---|
| передняя | средняя | задняя | |
| Бедро | 1.58–1.60 | 1.79–1.80 | 1.90–2.00 |
| Голень | 1.44 | 1.82–1.84 | 2.33–2.58 |
| Лапка | 4.20–4.21 | 4.29–4.30 | 5.08–5.38 |
Таблица 2.
Количество утолщенных игловидных щетинок на ногах самца Allopsontus tyrnyauzi sp. n.
| Сегмент | Пара ног | |||
|---|---|---|---|---|
| передняя | средняя | задняя | ||
| Членик лапки | 1-й | 3–4 | 4 | 6 |
| 2-й | 12 | 12 | 14 | |
| 3-й | 10 | 8 | 10 | |
| Голень | 4 | 12–14 | 16–18 | |
| Бедро | 20–25 | 6–8 | 6 | |
Вершинный угол стернитов брюшка прямой или тупой, на II–V сегментах брюшка самца составляет 90°, на VI сегменте – 100°, VII сегменте – 106°, VIII сегменте – 110°, брюшные кокситы I–VIII сегментов брюшка c 1 + 1 выпячивающимися мешочками (рис. 1, 6). Соотношение длины грифельков, стернитов и кокситов брюшка приведены в табл. 3.
Таблица 3.
Соотношения длин стернитов, кокситов и грифельков брюшка у самца Allopsontus tyrnyauzi sp. n.
| Сегмент брюшка | Стернит : кокситы | Грифельки (без опорных шипов) : кокситы | Опорные шипы : грифельки |
|---|---|---|---|
| II–V | 0.52–0.55 | 0.59–0.62 | 0.28 |
| VI | 0.49 | 0.56 | 0.29 |
| VII | 0.43 | 0.56 | 0.30 |
| VIII | 0.33 | 0.72 | 0.26 |
| IX | – | 0.87 | 0.16 |
Грудные тергиты без макрохет. Распределение сублатеральных макрохет на брюшных тергитах и кокситах приведено в табл. 4, их расположение на X тергите брюшка показано на рис. 1, 7. Кокситы IX сегмента брюшка с наружными макрохетами, внутренние макрохеты на них отсутствуют.
Таблица 4.
Число сублатеральных макрохет на брюшных тергитах и кокситах у самца Allopsontus tyrnyauzi sp. n.
| Сегмент брюшка | Тергит | Коксит |
|---|---|---|
| I–II | 0 | 0 |
| III | 1–2 + 1–2 | 0–1 + 0–1 |
| IV | 3 + 3 | 2 + 2 |
| V | 3 + 3 | 1–2 + 1–2 |
| VI | 3 + 3 | 2 + 2 |
| VII | 3 + 3 | 2 + 2 |
| VIII | 3–4 + 3–4 | 2–3 + 2–3 |
| IX | 5 + 5 | 2–0 – 0–1 |
| X | 4 + 4 | – |
Половой аппарат с парамерами на IX сегменте брюшка (рис. 1, 8). Парамеры 1 + 5-члениковые, пенис немного выступает за вершины парамер, доходит до вершин кокситов IX сегмента брюшка.
Дифференциальный диагноз. Отсутствие длинных тонких извилистых щетинок на ногах, нижнечелюстных и нижнегубных щупиках у описанных самцов позволяет отнести их к роду Allopsontus Silvestri 1911, для самок которого характерен длинный, тонкий членистый яйцеклад первого типа без копательных шипов (Каплин, 1993). Виды с одной парой выпячивающихся мешочков на I–VII кокситах брюшка у обоих полов объединяют в подрод Kaplinilis Mendes 1990, который включает 9 известных видов, а именно A. longistylis (Silvestri 1906), A. bifarius (Wygodzinsky 1970), A. confaratus (Wygodzinsky 1970), A. intergerivus (Wygodzinsky 1970), A. atrans (Kaplin 1982), A. dzhungaricus (Kaplin 1985), A. smelyanskii Kaplin 1999, A. volgensis Kaplin 1999, и A. abkhazicus Kaplin 2017 (Wygodzinsky, 1970; Каплин, 1982, 1985, 1999, 2017; Mendes, 1990; Mendes et. al., 2000). Самцы близкого подрода Allopsontoides Mendes 1990 с одним известным видом, A. simplex (Kaplin 1982), также имеют лишь одну пару выпячивающихся мешочков на I–VII кокситах брюшка, но у самок их две пары на II–IV брюшных кокситах. Среди указанных видов короткие густые прижатые пигментированные щетинки на 5–7-м члениках нижнечелюстных щупиков и на третьем членике нижнегубных щупиков имеются у самцов лишь одного вида – A. bifarius, известного из Западной Монголии и Тувы. Отличительные признаки A. tyrnyauzi sp. n. и A. bifarius приведены в табл. 5.
Таблица 5.
Дифференцирующие морфологические признаки самцов Allopsontus bifarius и A. tyrnyauzi sp. n.
| Признак | A. bifarius | A. tyrnyauzi sp. n. |
|---|---|---|
| Отношение длины глаза к его ширине | 0.9 | 1.0 |
| Отношение длины линии контакта глаз к их длине | 0.42 | 0.28–0.30 |
| Отношение ширины парных глазков к их длине | 2.0 | 2.9 |
| Сенсорные поля на передних бедрах самцов | Занимают 0.67 длины и 0.33 ширины бедра, включают 18 розетковидных сенсилл, расположенных в 2–3 неровных ряда | Занимают 0.43 длины и 0.15 ширины бедра, включают 9–11 розетковидных сенсилл, расположенных в один неровный ряд |
| Вершинный угол V стернита брюшка, градусы | 110 | 90 |
| Число члеников парамер полового аппарата | 1 + 6 | 1 + 5 |
Этимология. Видовой эпитет Allopsontus tyrnyauzi sp. n. образован от названия г. Тырныауз в Кабардино-Балкарии.
Allopsontus zinchenkoi Kaplin sp. n. (рис. 2, 1–9; 3, 1–4)
Рис. 2.
Allopsontus zinchenkoi sp. n.: 1 – глаза и парные глазки, 2 – цепочка передней части жгутика усиков передняя нога; 3, 4 – нижнегубной щупик, 5 – передняя часть верхней челюсти, 6 – передняя нога, 7 – задняя нога, 8 – стернит и кокситы VIII сегмента брюшка, 9 – вершина церки (1–3, 5–9 – голотип, самец; 4 – паратип, самка). Масштаб 0.1 мм.
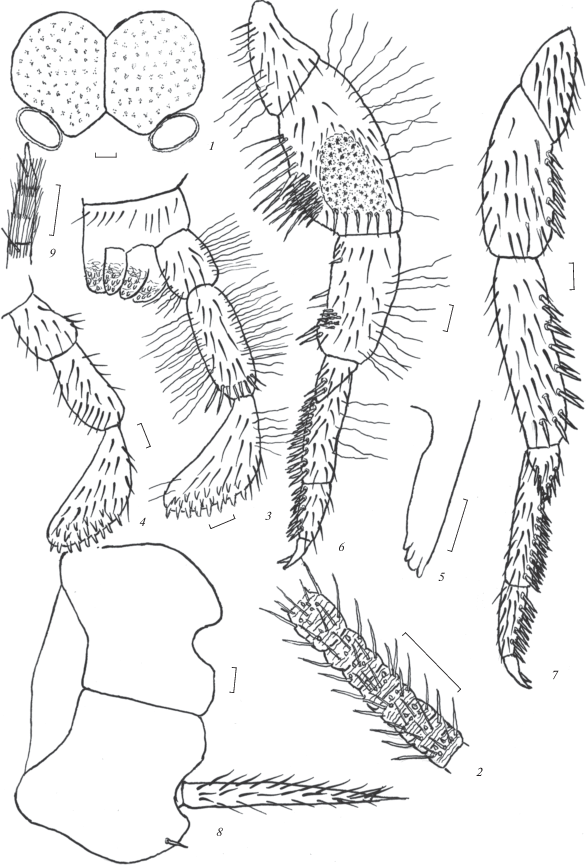
Материал. Голотип, ♂ (в препаратах), Таджикистан, Варзобское ущелье, 4 км ВСВ кишлака Калон, 39°03′ с.ш., 68°53′ в.д., 2474 м над ур. м., почвенные ловушки, 5–7.07. 2017. Паратипы: 1 ♀ (в препаратах), 3 ♂♂ (1 ♂ в препаратах) там же; 3 км СВ кишлака Калон, Сиёхкух, 39°04′ с.ш. 68°52′ в.д., 2440 м над ур. м, 9.07.2017; 1 ♀, там же, у ручья, 11.07. 2017 (В.К. Зинченко).
Описание. Длина тела самцов 7.3–8.5, самок – 10.2–11.0; ширина, соответственно 2.1–2.3 и 2.7–3.1 мм. Общая окраска тела буровато-коричневая, все тело с красновато-коричневым гиподермальным пигментом у самок средней, самцов сильной интенсивности. У самцов чешуйки дорсальной поверхности тела преимущественно черные, X, VII и IV тергиты брюшка с терминальными сравнительно узкими поперечными полосами светлых чешуек, вентральные чешуйки темно-бурые. У самок дорсальные и вентральные чешуйки бурые, коричневато-бурые, тергиты без выраженных полос. У самцов усики немного длиннее, самок – короче тела. Цепочки передней части жгутика усиков 8–9-члениковые (рис. 2, 2). Длина церок 2.7–3.1 мм, или 0.36–0.40 длины тела у самцов, 0.28–0.32 – у самок. Вершины церок с одним опорным шипом (рис. 2, 9). Церки самцов содержат 11–12, самок 13–14 члеников. Членики, за исключением одного или двух вершинных, с 2–3 боковыми опорными макрохетами в их передней части. Подобные макрохеты имеются также в базальной части хвостовой нити.
Глаза в спирте беловато-серые с мелким коричневым крапом, узкой темно-коричневой окантовкой и более широкой, особенно в теменной части, полосой из белых и беловато-светло-коричневых чешуек вокруг глаз позади парных глазков. У самцов впереди на лбу между глазами вблизи линии их контакта пятно из белых чешуек, от которого к непарному глазку идет узкая полоска из белых чешуек. Общая ширина глаз у самцов 0.81–0.87, самок 0.93–0.97, длина, соответственно 0.50–0.58 и 0.57–0.64 мм. Отношение длины глаза к его ширине у самцов 1.3–1.4, самок 1.2–1.3. Линия контакта глаз составляет у самцов 0.56–0.62, самок – около 0.5 их длины. Парные глазки сублатеральные по отношению к глазам, овальные (рис. 2, 1), светло-коричневые с узким белым ободом. Их размеры 0.21–0.23 × 0.13–0.14 мм. Длина глазков в 1.7–1.8 раза меньше их ширины. Расстояние между внутренними краями глазков 0.52–0.60, наружными – 0.88–0.92 общей ширины глаз.
Последний членик нижнечелюстных щупиков самцов с закругленной вершиной, его длина составляет у самцов 0.77–0.78, у самок 0.74–0.75 длины предпоследнего членика, 5-й членик нижнечелюстных щупиков самцов в 1.2–1.3, самок – 1.5–1.6 раза длиннее 4-го членика. Дорсальная поверхность 7-го членика нижнечелюстных щупиков самцов с 12–14, 6-го – с 10–12, 5-го – с 1–2; самок, соответственно с 11–12, 13–14 и 3–4 бесцветными зубьевидными хетами с темными вершинами. 5–7-й членики нижнечелюстных щупиков самцов с мелкими прижатыми пигментированными щетинками. На нижнегубных щупиках таких щетинок нет (рис. 3, 1). Первый и второй членики нижнегубных щупиков самцов с длинными извилистыми щетинками, имеющимися в небольшом количестве также на их третьем членике. Последний членик нижнегубных щупиков овально-треугольный, у самцов заметно изогнутый (рис. 2, 3, 4). Его длина у самцов и самок в 2.4–2.5 раза больше ширины. Вершины верхних челюстей 4-зубчатые (рис. 2, 5).
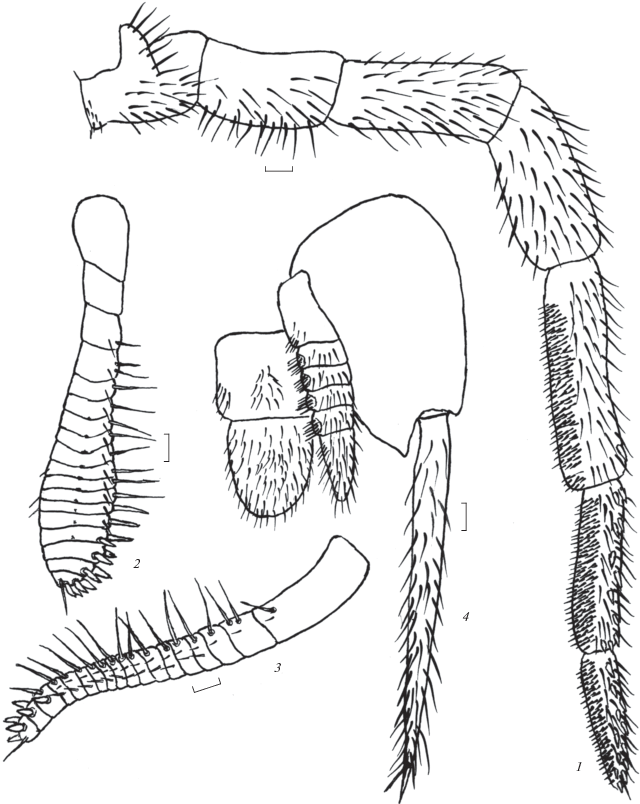
Рис. 3.
Allopsontus zinchenkoi sp. n.: 1 – нижнечелюстной щупик, 2 – передний гонапофиз, 3 – задний гонапофиз, 4 – IX коксит брюшка и половой аппарат (1, 4 – голотип, самец; 2, 3 – паратип, самка). Масштаб 0.1 мм.
Передние бедра самцов и самок расширенные (табл. 6). Отношение длины апикального членика задней лапки к ее общей длине у самцов 0.32–0.34, самок – 0.37–0.38 (рис. 2, 7). Вентральная поверхность лапок и голеней с двумя рядами игловидных пигментированных щетинок, распределение которых приведено в табл. 7. Передние бедра самцов с сравнительно обширными открытыми сенсорными полями, примыкающими к поперечному переднему ряду макрохет и включающими 46–55 розетковидных сенсилл, (рис. 2, 6). Ширина сенсорного поля составляет 0.56–0.60 ширины бедра, его длина – 0.45–0.48 длины бедра. Сенсорное поле не доходит до середины заднего основания бедра на 0.38–0.42 длины бедра. Передние голени самцов заметно изогнутые с полем, включающим 9–14 утолщенных коротких пигментированных игловидных щетинок. Передние бедра самцов с вентральным выступом, на котором расположено 20–24 утолщенных пигментированных игловидных щетинок. Грифельки имеются на тазиках средних и задних ног, их длина у самцов 0.46–0.48, самок около 0.6 мм. Отношение длины грифельков к ширине тазиков у самок 1.4–1.5, самцов – 1.2–1.4. Лапки, голени, бедра, вертлуги и тазики передних ног самцов с длинными тонких пигментированными извилистыми щетинками. Коготки лапок двуцветные с поперечной белой перевязью вблизи вершины и более размытой вблизи основания.
Таблица 6.
Отношение длины частей ног к их ширине у Allopsontus zinchenkoi sp. n.
| Часть ноги | Пол | Ноги | ||
|---|---|---|---|---|
| передние | средние | задние | ||
| Бедро | Самец | 1.93–2.03 | 2.53–2.66 | 2.74–2.98 |
| Самка | 2.05–2.07 | 2.28–2.34 | 2.46–2.47 | |
| Голень | Самец | 2.20–2.26 | 2.76–3.02 | 4.34–4.58 |
| Самка | 2.19–2.23 | 2.41–2.43 | 3.67–3.68 | |
| Лапка | Самец | 5.54–6.76 | 6.29–4.30 | 8.60–9.34 |
| Самка | 5.58–5.62 | 5.32–5.34 | 7.03–7.14 | |
Таблица 7.
Количество утолщенных игловидных щетинок на ногах самца Allopsontus zinchenkoi sp. n.
| Часть ноги | Самец | Самка | |||||
|---|---|---|---|---|---|---|---|
| пара ног | |||||||
| передняя | средняя | задняя | передняя | средняя | задняя | ||
| Членик лапки | 1-й | 2–3 | 5–6 | 6–7 | 3–4 | 4–6 | 7 |
| 2-й | 16–19 | 12–16 | 16–20 | 10–14 | 10–12 | 10–12 | |
| 3-й | 8–10 | 8–10 | 10 | 8–10 | 8 | 8 | |
| Голень | 2–3 + 9–14 | 10–12 | 14–16 | 12–13 | 13 | 18 | |
| Бедро | 20–24 | 4–7 | 8–9 | 2–3 | 2–3 | 8–9 | |
Вершинный угол брюшных стернитов тупой, на II–VI сегментах брюшка самцов составляет 103°–110°, на VII сегменте – 121°–123°, самки – соответственно 103°–115° и около 145°. VIII стернит брюшка у самцов трапециевидный с закругленными передними боковыми углами, сравнительно ровным передним и выпуклым задним (базальным) основанием (рис. 2, 8). Переднее основание трапеции в 2.0 раза короче базального основания. Базальные углы стернита около 37°. Брюшные кокситы I–VII сегментов брюшка самцов c 1 + 1, у самок кокситы I, VI и VII сегментов с 1 + 1, II–V – с 2 + 2 выпячивающимися мешочками. Соотношение длины грифельков, стернитов и кокситов брюшка приведены в табл. 8.
Таблица 8.
Соотношения длин стернитов, кокситов и грифельков брюшка у Allopsontus zinchenkoi sp. n.
| Сегмент брюшка | Стернит : кокситы | Грифельки (без опорных шипов) : кокситы | Опорные шипы : грифельки | |||
|---|---|---|---|---|---|---|
| самец | самка | самец | самка | самец | самка | |
| II | 0.63 | 0.62 | 0.95 | 0.81 | 0.22 | 0.19 |
| III–V | 0.67–0.68 | 0.64 | 085–0.92 | 0.74–0.76 | 0.22–0.24 | 0.19–0.20 |
| VI | 0.59 | 0.59 | 0.83 | 0.72 | 0.24 | 0.20 |
| VII | 0.51 | 0.39 | 0.94 | 0.86 | 0.20 | 0.17 |
| VIII | 0.28 | – | 1.05 | 1.01 | 0.20 | 0.15 |
| IX | – | – | 1.45 | 1.13 | 0.12 | 0.10 |
Грудные, I–III брюшные тергиты и брюшные стерниты без макро- и мезохет. Распределение макро- и мезохет на прочих брюшных тергитах и кокситах приведено в табл. 9.
Таблица 9.
Число сублатеральных макро- и мезохет на брюшных тергитах и кокситах Allopsontus zinchenkoi sp. n.
| Сегмент брюшка | Тергит | Коксит | ||
|---|---|---|---|---|
| самец | самка | самец | самка | |
| I | 0 | 0 | (5–7) – (3–5) + (3–5) – (5–7) | 9 – 8 + 9 – 8 |
| II | 0 | 0 | 0 | 3 – 0 + 0 – 3 |
| III | 1 + 1 | 0 | 1 – 0 + 0 – 1 | 0 |
| IV | 1–2 + 1–2 | 0 | (1–2) – 0 + 0 – (1–2) | 1 – 0 + 0 – 1 |
| V | 2–4 + 2–4 | 1 + 1 | (1–2) – 0 + 0 – (1–2) | 1 – 0 + 0 – 1 |
| VI | 3–4 + 3–4 | 1 + 1 | 1 – 0 + 0 – 1 | 1 – 0 + 0 – 1 |
| VII | 2–4 + 2–4 | 2–3 + 2–3 | (1–2) – 0 + 0 – (1–2) | 0 |
| VIII | 4–5 + 4–5 | 3 + 3 | (0–1) – 0 + 0 – (0–1) | 0 |
| IX | 3–5 + 3–5 | 3–4 + 3–4 | 0 | 14 – 1 + 1 – 15 |
| X | 1–2 + 1–2 | 1 + 1 | – | – |
Яйцеклад самки короткий, утолщенный, членистый с копательными шипами, незначительно выходит за вершины кокситов IX сегмента брюшка. Длина яйцеклада 1.5–1.6 мм. Передние и задние гонапофизы 18-члениковые (рис. 3, 2, 3). Шесть вершинных члеников передних гонапофизов с копательными шипами. На первом членике от вершины их 3–4, 2–4-м – по 2, 5-м и 6-м по одному шипу. Три вершинных членика задних гонапофизов с короткими толстыми копательными шипами. На первом членике от вершины их 3, на втором – 2, третьем – один шип. Вершина задних гонапофизов с копательными шипами заметно изогнута. Все членики задних гонапофизов и 17 члеников передних гонапофизов, за исключением базального членика, со щетинками. 4–16-й членики задних гонапофизов с одной, редко двумя длинными боковыми щетинками, длина которых в 1.5 раза больше ширины члеников. Апикальные членики передних и задних гонапофизов с сравнительно короткими бесцветными конечными иглами, длина которых примерно равна длине двух терминальных члеников, взятых вместе.
Половой аппарат самца с парамерами на IX сегменте брюшка (рис. 3, 4). Парамеры 1 + 5-члениковые, пенис широкий, почти не выступает за вершины парамер. Пенис и парамеры немного выступают за вершины кокситов IX сегмента брюшка. Апикальный членик пениса длиннее базального в 1.2–1.3 раза. Отношение ширины апикального членика пениса к его длине составляет около 0.66.
Дифференциальный диагноз. Allopsontus zinchenkoi sp. n. может быть отнесен к подроду Anisopsontus Mendes 1990, представители которого характеризуются наличием одной пары выпячивающихся мешочков на кокситах I–VII сегментов брюшка у самцов, а самки имеют одну пару мешочков на I, VI и VII, две пары на II–V сегментах брюшка и яйцеклад с копательными шипами. Подрод включает три описанных вида (Wygodzinsky, 1970; Каплин, 1982, 1987): A. ciliatus (Wygodzinsky 1970) (Монголия, Бурятия, Тыва, Казахстан, Киргизия, Таджикистан), A. hissaricus (Kaplin 1987) (Таджикистан, Гиссарский хребет, ущелье Кондара) и A. pulchellus (Kaplin 1982) (Юго-Восточный Казахстан). A. zinchenkoi sp. n. отличается от известных подрода Anisopsontus цветом глаз, отношением ширины парного глазка к его длине, более длинными грифельками брюшных кокситов, более крупными размерами сенсорных полей на передних бедрах самцов, сравнительно коротким яйцекладом, включающим 18 члеников (табл. 10).
Таблица 10.
Дифференцирующие морфологические признаки видов подрода Allopsontus (Anisopsontus) Mendes 1990
| Признак | A. ciliatus | A. pulchellus | A. hissaricus (самка не известна) | A. zinchenkoi sp. n. |
|---|---|---|---|---|
| Длина тела, мм | 8–11 | 10–14 | 9 | 7–11 |
| Цвет глаз | Сероватые, двуцветные с мелким темным крапом | Темные с голубоватым оттенком | Голубоватые | Беловато-серые с мелким коричневым крапом |
| Отношение длины глаза к его ширине | 1.1–1.2 | 1.3 | 1.4 | 1.2–1.4 |
| Отношение длины линии контакта глаз к их длине | 0.65 | 0.60 | 0.57–0.58 | 0.5–0.6 |
| Отношение длины парного глазка к его ширине | 1.9–2.0 | 1.2–1.3 | 1.5 | 1.7–1.8 |
| Отношение расстояния между внутренними краями парных глазков к общей ширине глаз | 0.55 | 0.55–0.65 | 0.44–0.47 | 0.52–0.60 |
| Мелкие прижатые пигментированные щетинки на члениках нижнечелюстных щупиков самцов | 5–7-м | 2–7-м | Отсутствуют | 5–7-м |
| Отношение длины последнего членика нижнегубных щупиков к его ширине | 2.0–2.1 (♂), 3.3–3.4 (♀) | 2.2–2.4 (♂), 2.6–2.7 (♀) | 3.0–3.3 | 2.4–2.5 |
| Отношение длины сенсорного поля на передних бедра самцов к длине и отношение его ширины к ширине бедра | 0.50, 0.45 |
0.25–0.30, 0.35–0.40 | 0.25, 0.23 |
0.56–0.60, 0.45–0.48 |
| Вершинный угол стернита V сегмента брюшка, градусы | 100–110 | 110–115 | 100–110 | 100–105 |
| Отношение длины грифельков (без опорных шипов) к длине кокситов III–V сегментов брюшка | 0.55–0.65 | 0.65–0.70 | 0.62–0.68 | 0.85–0.92 (♂), 0.74–0.76 ((♀) |
| Отношение длины грифельков (без опорных шипов) к длине кокситов VIII сегмента брюшка | 0.7–0.8 | 0.8–0.9 | 0.9 | 1.0–1.1 |
| Отношение длины грифельков (без опорных шипов) к длине кокситов IX сегмента брюшка самца (самки) | 0.95 (0.7– 0.75) | 1.25 (0.9) | 1.40 | 1.45 (1.13) |
| Число члеников парамер полового аппарата самцов | 1 + 5 | 1 + 7 | 1 + 7 | 1 + 5 |
| Число члеников яйцеклада | 20–22 | 36–37 | ? | 18 |
| Число терминальных члеников задних гонапофизов с копательными шипами | 3 | 19 | ? | 3 |
Этимология. Вид назван именем известного энтомолога В.К. Зинченко.
Список литературы
Каплин В.Г., 1982. Новые данные по фауне щетинохвосток (Thysanura) Монголии, Казахстана и Средней Азии // Насекомые Монголии. Вып. 8. Л.: Наука. С. 16–61.
Каплин В.Г., 1985. Новые виды щетинохвосток рода Machilanus (Thysanura, Machilidae) с Джунгарского Алатау // Зоологический журнал. Т. 64. № 3. С. 459–461.
Каплин В.Г., 1987. Новые виды щетинохвосток рода Machilanus (Thysanura, Machilidae) с Гиссарского хребта // Зоологический журнал. Т. 66. № 7. С. 1099–1103.
Каплин В.Г., 1993. Щетинохвостки рода Allopsontus Silv. (Thysanura, Machilidae) мировой фауны // Зоологический журнал. Т. 72. № 3. С. 53–67.
Каплин В.Г., 1999. Новые виды щетинохвосток семейств Machilidae и Lepismatidae (Thysanura) из Европейской части России и Узбекистана // Зоологический журнал. Т. 78. № 1. С. 16–30. (Entomological Review. V. 79. № 3. P. 310–324).
Каплин В.Г., 2017. Новые виды щетинохвосток рода Allopsontus Silv. (Microcoryphia, Machilidae) из Юго-Восточного Казахстана и Абхазии // Зоологический журнал. Т. 96. № 6. С. 641–650. (Entomological Review. V. 97. № 5. P. 602–611).
Mendes L.F., 1990. An annotated list of generic and specific names of Machilidae (Microcoryphia, Insecta) with identification keys for the genera and geographical notes // Estudos, Ensaios e Documentos. Instituto de Investigaçȃo Cientifica Tropical. Lisboa. V. 155. P. 1–127.
Mendes L.F., Gaju-Ricart M., Bach de Roca C., Molero-Baltanás R., 2000. On some Silvestri species of Machilidae (Microcoryphia, Insecta) which types are in the Muséum National d’Histoire Naturelle, Paris // Pedobiologia. V. 44. № 3–4. P. 268–284.
Wygodzinsky P., 1970. Results of the zoological explorations of Dr. E. Kaszab in Mongolia. 202. Thysanura and Microcoryphia (Insecta) // American Museum novitates. № 2401. P. 1–25.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал