

Зоологический журнал, 2019, T. 98, № 10, стр. 1177-1185
О значении находки черепа оленелося (Cervalces sp., Cervidae) со штангами рогов разной длины
П. А. Никольский a, *, А. Э. Басилян a, **, А. М. Анисимов b, c, ***, В. С. Зажигин a, ****, Е. Ю. Павлова c, *****, В. В. Питулько d, ******
a Геологический институт РАН
119017 Москва, Россия
b Институт наук о Земле, С.-Петербургский государственный университет
199034 С.-Петербург, Россия
c ГНЦ РФ Арктический и антарктический научно-исследовательский институт
199397 С.‑Петербург, Россия
d Институт истории материальной культуры РАН
191186 С.-Петербург, Россия
* E-mail: cervalces@rambler.ru
** E-mail: alexandr.basilyan@gmail.com
*** E-mail: m.anisimov@spbu.ru
**** E-mail: zazhvol@gmail.com
***** E-mail: pavlovaelena759@gmail.com
****** E-mail: pitulko.vladimir@gmail.com
Поступила в редакцию 10.03.2019
После доработки 25.03.2019
Принята к публикации 10.04.2019
Аннотация
Изучен череп оленелося Cervalces sp. (Cervidae), имеющий штанги рогов существенно разной длины. Этот череп, вместе с нижней челюстью и четырьмя позвонками, был найден в низовьях р. Яна. Его геологический возраст самый конец эоплейстоцена–первая половина раннего неоплейстоцена. Неравная длина штанг воспроизводилась у данной особи в каждой генерации рогов на протяжении всей жизни, что может указывать на генетическую природу явления, хотя возможна и негенетическая причина такой диспропорции – многие полевые наблюдения и экспериментальные данные показывают, что асимметричные нарушения роста рогов у современных оленей часто происходят из-за травм, особенно повреждений передних конечностей. Если верна генетическая гипотеза, то такая аномалия может быть проявлением интенсивного формообразования, которое могло быть адаптивной реакцией на первое в плейстоцене значительное похолодание климата. Пропорции штанг рогов являются наиболее ценным критерием видовой и подвидовой таксономии и диагностики ископаемых лосей. Описанный нами случай, ввиду его исключительной редкости, вовсе не делает этот признак бесполезным. Однако данная генетическая аномалия может указывать на увеличение изменчивости эволюционных признаков в периоды значительных изменений климата, которое следует иметь в виду при интерпретации морфологии животных, живших в такие эпохи.
Первые представители трибы лосей Alceini (Brookes 1828) появляются в геологической летописи два с половиной (Kahlke, 1990; Breda, Marchetti, 2005, Никольский, 2010) или даже три миллиона лет назад (Vislobokova et al., 1995). Приблизительно 800 тыс. л. н. древние лоси распространились по большей части Палеарктики (кроме самого ее юга) и проникли в Северную Америку (Churcher, Pinsof, 1987; Никольский, 2010). Триба включает до 13 короткоживущих широкоареальных таксонов видового и подвидового ранга, относящихся к вымершему роду Cervalces Scott 1885 (оленелоси) и ныне живущему роду Alces Gray 1821 (лоси).
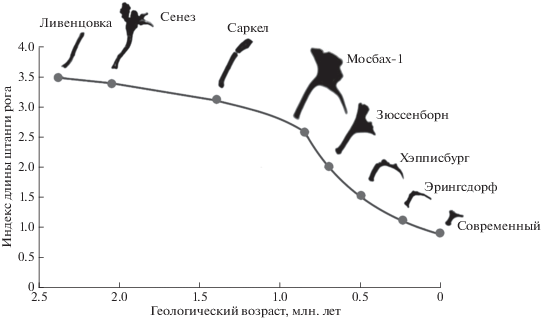
Многие исследователи считают развитие Alceini в Евразии филетическим (Johnson, 1874; Scott, 1885; Heintz, Poplin, 1981; Шер, 1986; Lister, 1993; Breda, Marchetti, 2005; Никольский, 2010). Проследить единую слабоветвящуюся филетическую линию Alceini позволяет доступная для морфометрического выражения эволюционная тенденция, сохраняющаяся на протяжении всей истории трибы. Этой тенденцией является сокращение относительной длины штанги рога при одновременном увеличении площади лопаты (рис. 1).
Однако другие авторы высказывались против филетической модели развития Alceini (Azzaroli, 1985, 1994; Kahlke, 1990; Вислобокова, 1990; Боескоров, 2001). В том числе они указывали на то, что в ряде местонахождений морфотипы Alceini существенно разной эволюционной продвинутости встречаются совместно, следовательно, они геологически одновозрастны. Главными из таких местонахождений в Европе являются Мосбах (Германия) и побережье Норфолка (Англия). В Восточной Сибири к ним относится Улахан-Суллар в низовьях р. Адыча (Республика Саха-Якутия). В качестве возражения можно предположить, что подобное смешение морфотипов может быть результатом аллювиальной или иной тафономической конденсации (палимпсестизации). Иллюзию совместного залегания остатков разного эволюционного уровня также может создавать неточность стратиграфических данных или неверная привязка находок к слоям разреза. Нам показалось полезным исследовать одну удивительную находку, имеющую прямое отношение к этому вопросу. В 2008 г. в низовьях р. Яны (северо-восток Сибири, Республика Саха-Якутия) в ходе исследований геологического строения окрестностей Янской палеолитической стоянки был найден череп, часть нижней челюсти и несколько позвонков древнего оленелося. Штанги рогов этого черепа значительно различаются по длине, одновременно демонстрируя таким образом как архаичные, так и продвинутые признаки на разных рогах. Полностью исключая оба приведенных выше возражения, эта находка прямо указывает на возможность наличия у лосей одного вида (и даже у отдельного индивидуума) признаков разной эволюционной продвинутости. Задачей нашего исследования является ввести в научный оборот эту важную находку и обсудить ее значение для изучения эволюции Alceini и биостратиграфии.
МАТЕРИАЛ И МЕТОДИКА
Материал
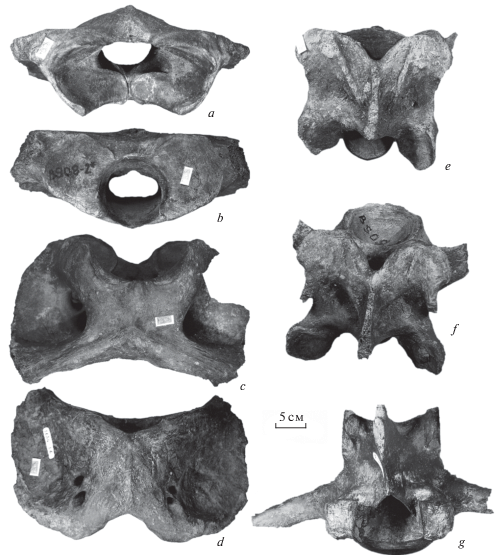
Часть скелета половозрелого самца среднего индивидуального возраста: фрагмент черепа со штангами рогов, обломанными у начала лопат ГИН-1134-1f (рис. 2), левая ветвь нижней челюсти ГИН-1134-1a, атлант ГИН-1134-1b (рис. 3a–3d), III шейный позвонок ГИН-1134-1с (рис. 3e), IV шейный позвонок ГИН-1134-1d (рис. 3f), поясничный позвонок ГИН-1134-1e (рис. 3g). Местонахождение Соп-Хая (нижнее течение р. Яна), криофация 2b слоя 2 – алевриты с псевдоморфозами по повторно-жильным льдам в основании третьей террасы.

Методика
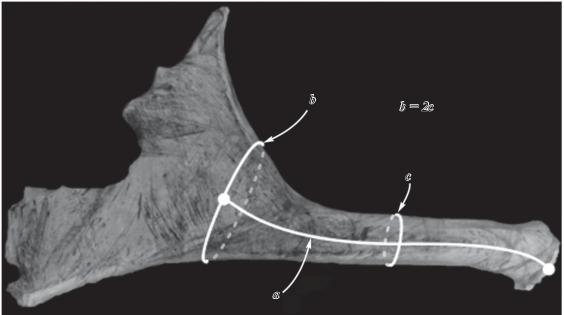
Важнейшим таксономическим признаком древних лосей является относительная длина штанги рога. Для ее определения обычно измеряют обхват или диаметр штанги рога в ее середине, а также длину штанги до начала лопаты. У древних Alceini переход штанги в лопату очень постепенный, что затрудняет однозначное определение места перехода штанги в лопату. У разных авторов разброс измерений длины штанги рога достигает по меньшей мере 25%. Для исключения такой неопределенности на основе принципа, предложенного Чачером и Пинсофом (Churcher, Pinsof, 1987), нами была разработана усовершенствованная методика, согласно которой длина штанги рога (рис. 4a) определяется как расстояние от розетки до того места начала лопаты (рис. 4b), где обхват рога равен двум обхватам в средней части штанги (рис. 4c).
РЕЗУЛЬТАТЫ
Стратиграфия местонахождения Соп-Хая и геологический возраст находки
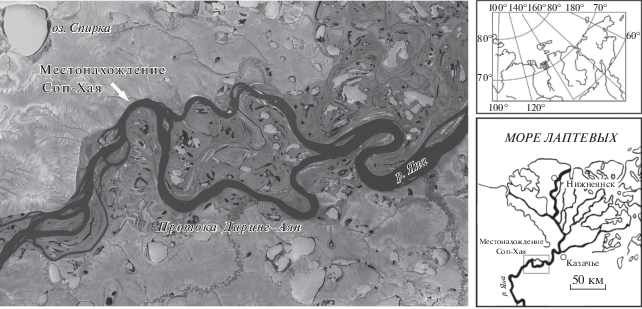
В 2001 г. в низовьях р. Яна была открыта древнейшая в Заполярье палеолитическая стоянка возрастом около 31 000 календарных лет (Pitulko et al., 2004). В ходе многолетнего изучения геологии и стратиграфии окрестностей стоянки в основании разреза четвертичных отложений были выявлены неизвестные здесь ранее досредненеоплейстоценовые слои и найдены разнообразные палеонтологические материалы, в том числе остатки представителя трибы Alceini, обсуждаемые в настоящей статье. Открытое местонахождение древней фауны в соответствии с названием местности получило название Соп-Хая.
Соп-Хая находится на левом берегу р. Яна в 100 км от берега Северного Ледовитого океана (рис. 5). Здесь в береговых обрывах вскрываются разновозрастные четвертичные отложения, слагающие три надпойменные террасы. Высота третьей террасы достигает 40‑45 м, второй – 16–18, первой – 10–12 м.
Разрез III террасы имеет следующее строение снизу вверх (рис. 6):
Рис. 6.
Общий вид и детали строения разреза Соп-Хая: a – гравелиты, b – алевриты песчанистые с повторно-жильными льдами, c – алевриты песчанистые с псевдоморфозами по повторно-жильному льду, d – песок средне-мелкозернистый с линзами гравия, e – прослои торфа, f – алевриты песчанистые эоловые с палеопочвами, g – алевриты песчанистые, h – место находки Cervalces sp., i – находки костей млекопитающих, j – граница между непротаявшей (2a) и протаявшей (2b) фациями слоя 2. На детальном разрезе сеть повторно-жильных льдов не показана. Цифрa в кружкe обозначаeт номер слоя.
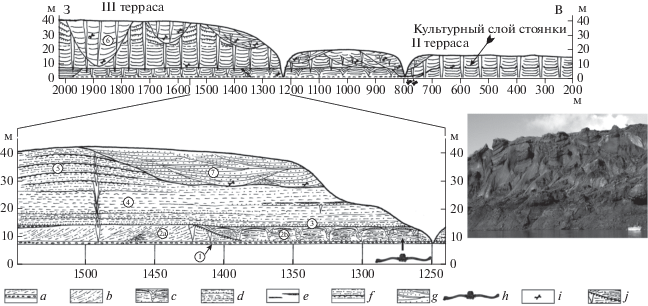
Слой 1. Гравелиты серо-бурые в песчаном матриксе, видимая мощность 1 м.
Слой 2. Геологические тела, обозначенные нами как “слой 2a” и “слой 2b” представляют различные криофации слоя 2. Слой 2a – непротаявший участок слоя 2, вскрывающийся в западной части обнажения: алевриты серые, песчанистые, с сетью сингенетических повторно-жильных льдов. Слой 2b – протаявший в древности участок слоя 2, вскрывающийся в восточной части обнажения: алевриты серые, песчанистые с тонкими прослоями торфа и псевдоморфозами по повторно-жильным льдам. В слое 2b встречены единичные остатки мелких и крупных млекопитающих, в том числе описываемый в настоящей статье экземпляр древнего лося. Мощность слоя 2 достигает 5 м.
Слой 3. Песок серо-желтый, мелко-среднезернистый с мелкой галькой. Встречены многочисленные остатки мелких млекопитающих. Мощность 0.8 м.
Слой 4. Песок серый средне-мелкозернистый, косослоистый с повторно-жильными льдами. Мощность 15 м.
Слой 5. Алеврит серо-бурый песчанистый, косослоистый, эоловый, с повторно‑жильными льдами. Мощность 13 м.
Слой 6. Алеврит серый, песчанистый с повторно-жильными льдами, обогащен органикой, из-за чего имеет почти черный цвет. Встречены остатки крупных млекопитающих. Мощность 30 м.
Слой 7. Алеврит серо-бурый, песчанистый с повторно-жильными льдами, аллювиальный. Встречаются остатки крупных млекопитающих. Мощность 15 м.
Остатки Alceini ГИН-1134-1a-f найдены в алевритах перетаявшей части слоя 2 – в “слое” 2b (рис. 6).
Находки в слое 2 остатков Lemmus sp. указывают на то, что он сформировался не ранее начала эоплейстоцена. Присутствие здесь зубов Dicrostonyx sp. позволяет уточнить нижнюю границу слоя, поместив ее на уровень границы эоплейстоцена и неоплейстоцена, так как Dicrostonyx сменили Predicrostonyx на этом рубеже (Зажигин, 2004). Верхний возрастной предел слоя 2 определяется возрастом фауны из слоя 3, который с размывом перекрывает слой 2:
Rodentia: Clethrionomys ex gr. rutilus Pallas; Lemmus sibiricus Kerr.; Dicrostonyx simplicior Fejfar; Microtus sp.; Mimomys sp. (переотложен);
Lagomorpha: Lepus sp.; Ochotona sp.
Остатки D. simplicior характерны для второй половины раннего неоплейстоцена на уровне окской холодной фазы плейстоцена (Зажигин, 2004). Таким образом, по биостратиграфическим данным возраст слоя 2 находится в интервале между самым концом эоплейстоцена и началом второй половиной нижнего неоплейстоцена, что примерно соответствует геохронологическому диапазону 0.8–0.5 млн лет.
Таксономические признаки, промеры и индексы
Промеры костей, необходимые для определения родовой принадлежности находки приведены в табл. 1.
Таблица 1.
Промеры костей и зубов экземпляра Cervalces sp. ГИН-1134-1 в сравнении с теми же промерами современных и ископаемых Alces
| Промеры и индексы | Экземпляр ГИН-1134-1, мм | Современные и ископаемые Alces, мм |
|---|---|---|
| Межроговая ширина черепа | 262 | 186–206 (n = 44) |
| Высота затылка | 139 | 112–138 (n = 44) |
| Ширина затылка | 196 | 157–189 (n = 44) |
| Длина зубного ряда | 187 | 154–176 (n = 82) |
| Индекс высоты затылка (отношение ширины затылка к его высоте) | 1.41 | 1.20–1.37 (n = 44) |
Промеры штанг рогов (табл. 2) показали, что левый рог длиннее правого на 38%. Индекс левого рога (отношение длины штанги к ее обхвату) равен 3.58, а правого – 2.6 (табл. 2).
Таблица 2.
Промеры штанг рогов ГИН-1134-1f
| Рог | Обхват в середине штанги | Длина штанги | Индекс длины штанги рога (отношение длины штанги рога к ее обхвату) |
|---|---|---|---|
| Правый | 212 | 550 | 2.6 |
| Левый | 212 | 760 | 3.58 |
Изучение костей, зубов и рогов экземпляра ГИН-1134-1 выявило следующие эволюционно-значимые морфологические и морфометрические особенности: штанги рогов длинные (рис. 2, табл. 1); переход штанги в лопату очень постепенный (рис. 2); s‑образный изгиб штанг рогов выражен; череп низкий и широкий (рис. 2, табл. 1), ростральная часть черепа короткая. Хотя сама ростральная часть и не сохранилась, на это указывает небольшая относительная длина нижней челюсти (табл. 1).
ОБСУЖДЕНИЕ
Большие абсолютные размеры скелета (табл. 1), огромная длина штанг рогов (табл. 2), очень постепенный переход штанги в лопату, заметный s-образный изгиб штанг рогов, низкий и широкий череп, короткий рострум (табл. 1) – признаки, совместное наличие которых позволяет уверенно отнести рассматриваемый экземпляр к роду Cervalces и исключить его принадлежность к роду Alces. Видовую принадлежность находки мы тут не рассматриваем, так как имеющиеся противоречивые данные не позволяют решить эту задачу без таксономической ревизии трибы, что выходит за пределы темы настоящего сообщения.
Итак, в отложениях финального эоплейстоцена–начала нижнего неоплейстоцена северо-востока Сибири найден череп оленелося, штанги рогов которого различаются по длине почти на 40%. Такая диспропорции штанг рогов является уникальной. На 210 изученных нами черепах лосей разного геологического возраста из Евразии и Северной Америки разница длин штанг ни разу не превысила 10%. (Никольский, неопубликованные данные).
Асимметрия развития сохранялась у данного индивидуума в каждой генерации рогов, начиная с юного возраста. Об этом свидетельствуют значительные однотипные деформации шейных и поясничных позвонков. Характер этих деформаций указывает на компенсацию скелетно-мышечной системой скручивающего момента, воспринимаемого (в том числе) позвоночником от черепа, правая и левая стороны которого испытывали неодинаковую нагрузку от рогов. Воспроизведение асимметрии развития рогов в каждой их генерации может указывать на генетическую природу этого явления. С другой стороны, большое количество наблюдений и экспериментальные данные показывают, что к односторонней аномалии развития рогов у оленей могут приводить самые разные негенетические причины: травмы и ампутация одной из передних конечностей, серьезные ранения, односторонние повреждения зубов, патологии верхней части пенька рога и даже односторонняя кастрация (Clarke, 1916; Penrose, 1924; Moore, 1931; Seton, 1937; Cowan, 1956; Davis, 1983; Marburger et al., 1972; Karns, Ditchkoff, 2013).
Ввиду отсутствия мягких тканей и большей части скелета мы не можем надежно верифицировать травматическую гипотезу асимметрии рогов у оленелося с Яны, кроме ее части, касающейся патологии пеньков рогов. Пеньки и розетки обоих рогов имеют обычный, не измененный вид – они не были повреждены. Это исключает из числа причин аномалии формирования рогов повреждения пеньков. Серьезные патологии другого типа, особенно отсутствие одной из передних конечностей, ввиду изобилия крупных хищников в начале раннего плейстоцена в Западной Берингии наверняка не позволили бы рассматриваемому оленелосю прожить много лет (стертость зубов и срощенность швов черепа указывают на ранний взрослый возраст особи). Таким образом, более вероятной нам кажется генетическая гипотеза.
Наличие генетической аномалии, выразившейся в обнаруженных нами фенотипических особенностях у лосей именно этого геологического возраста кажется нам не только не удивительным, но и закономерным. В это время, на рубеже эоплейстоцена и неоплейстоцена, началась эпоха наибольшей амплитуды флуктуаций климата (Head, Gibbard, 2005), что стало причиной резких и значительных изменений условий окружающей среды. У лосей такое разрушение привычной среды обитания привело к резкому ускорению эволюции (Никольский, 2010) (рис. 1). Поиск полезных адаптивных новообразований должен был сопровождаться увеличением пределов изменчивости эволюционно-значимых признаков, одним из которых как раз и является укорочение штанги рога.
Заставляет ли обсуждаемый феномен отказаться от применения индексов пропорций штанг рогов для таксономии и филогенетики? Опровергает ли он филетическую модель эволюции Alceini? По всей видимости, нет. Изучение большой серии хорошо датированных черепов с достаточно сохранившимися рогами из всех главных местонахождений ископаемых представителей трибы показывает, что в целом филетический эволюционный тренд сохраняется (Никольский, 2010) (табл. 3, рис. 1). Наши данные показывают, что максимальную величину изменчивости следует ожидать только у лосей, обитавших в промежутке между концом эоплейстоцена и началом среднего неоплейстоцена (Никольский, 2010). На этом рубеже произошла первичная адаптация к новым условиям холодного плейстоцена, после чего наступила относительная стабилизация, хотя заданный в начале плейстоцена (2.5 млн. л. н.) эволюционный тренд сохранился, правда, скорость эволюции увеличилась. Неразличимые при сегодняшней разрешающей способности летописи бифуркации никогда не приводили к закреплению параллельных филетических ветвей (единственное исключение – ответвление американских лосей в раннем неоплейстоцене).
Таблица 3.
Индексы длины штанги рога некоторых важнейших представителей филетической линии Cervalces–Alces (в Европе)
| Местонахождение | Возраст, млн лет | Индекс длины штанги рога (отношение длины штанги к ее окружности) |
|---|---|---|
| Ливенцовка | 2.45 | 3.5 |
| Сенез | 2.05 | 3.4 |
| Саркел | 1.4 | 3.18 |
| Мосбах-1 | 0.85 | 2.6 |
| Зюссенборн | 0.7 | 2.03 |
| Хэпписбург | 0.5 | 1.547 |
| Веймар-Эрингсдорф | 0.2 | 1.35 |
| Моршанск (Современный) | 0.01 | 0.928 |
Рассмотренный в настоящем сообщении случай имеет большее значение для биостратиграфии. Точность биостратиграфических датировок по остаткам лосей для нижнего неоплейстоцена может оказаться низкой из-за больших пределов морфологической изменчивости, связанной с ускорением эволюции, спровоцированным глобальным похолоданием. Если бы рога рассматриваемого в статье экземпляра были бы найдены по отдельности, то по левому рогу, индекс длины штанги рога которого 3.58, можно было бы датировать вмещающие осадки самым началом эоплейстоцена (2.6 млн. лет), а по правому, индекс 2.6, – рубежом эоплейстоцена и неоплейстоцена (0.8 млн. лет) (рис. 1). Возможно в какой-то степени именно с кратковременным возрастанием пределов морфологической изменчивости, а не только, или не столько, с плохой привязкой и тафономической конденсацией связано присутствие в местонахождениях близкого возраста (нижние слои Мосбаха, Кромер Форест Бэд в Норфолке, Улахан-Суллар) “разновозрастных” морфотипов оленелосей.
Получить более надежные данные по обсуждаемым вопросам станет возможным после более точного датирования эволюционно значимых морфотипов лосей.
Список литературы
Боескоров Г.Г., 2001. Систематика и происхождение современных лосей. М.: Наука. 120 с.
Вислобокова И.А., 1990. Ископаемые олени Евразии // Труды Палеонтологического института AH CCCР. Т. 240. С. 1–208.
Зажигин В.С., 2004. О копытных леммингах (Dicrostonyx, Microtinae, Rodentia) Ойгос‑Яра Восточной Сибири и о видовом статусе среднеплейстоценового вида рода Dicrostonyx // Естественная история российской восточной Арктики в плейстоцене и голоцене. М.: ГЕОС. С. 14–31.
Никольский П.А., 2010. Систематика и стратиграфическое значение лосей (Alcini, Cervidae, MAMMALIA) в позднем кайнозое Евразии и Северной Америки. Дис. … канд. геол.-минерал. наук. М.: ГИН РАН. 258 с.
Шер А.В., 1971. Млекопитающие и стратиграфия плейстоцена крайнего Северо-Востока СССР и Северной Америки. М.: Наука. 310 с.
Шер А.В., 1986. История и эволюция лосей. Биология и использование лося. М.: Наука. С. 6–35.
Azzaroli A., 1985. Taxonomy of the Quarternery Alcini (Cervidae, Mammalia) // Acta Zoologica Fennica. V. 170. P. 179–180.
Azzaroli A., 1994. Forest Bed elks and giant deer revisited // Zoological Journal of the Linnean Society. V. 112. № 1–2. P. 119–133.
Breda M, Marchetti, M., 2005. Systematical and biochronological review of Plio-Pleistocene Alceini (Cervidae; Mammalia) from Eurasia // Quaternary Science Review. V. 24. № 5-6. P. 775–805.
Churcher C.S., Pinsof J.D., 1987. Variation in the antlers of North American Cervalces (Mammalia; Cervidae): Review of new and previosly recorded specimens // Journal of Vertebrate Paleontplogy. V. 7. № 4. P. 373–397.
Clarke F.C., 1916. Malformed antlers of deer // California Fish and Game. V. 2. № 2. P. 119–123.
Cowan I.M., 1956. Life and times of the coast black-tailed deer // The Deer of North America. Taylor W.P. (ed.). Harrisburg, Pennsylvania: The Stackpole Co. P 523–617.
Davis T.A., 1983. Antler asymmetry caused by limb amputation and geophysical forces // Antler development in Cervidae. Ed. Brown R.D. Kingsville, Texas, USA: Caesar Kleberg Wildlife Research Institute. P. 223–229.
Head M.J., Gibbard E.L., 2005. Early-Middle Pleistocene Transitions: The Land-Ocean Evidence. London: Geological Society. 326 p.
Heinz E., Poplin F., 1981. Alces carnutorum (Laugel,1862) du Pleistocene de Saint-Prest (France). Systmatique et evolution des Alcines (Cervidae, Mammalia) // Quartarpalaontologie. V. 4. P. 105–122.
Johnson R., 1874. Notice of a new species of deer from the Norfolk Forest Bed // The Annals and Magazine of Natural History. № 73. P. 1–4.
Kahlke H.D., 1990. On the Evolution, Distribution and Taxonomy of Fossil Elk/Moose // Quartarpalaontologie. V. 8. P. 83–106.
Karns G.R., Ditchkoff S.S., 2013. Trauma-Induced Malformed Antler Development in Male White-Tailed Deer // Wildlife Society Bulletin. V. 37. № 4. P. 832–837.
Lister A.M., 1993. The stratigraphical significance of deer species in the Cromer Forest-bed Formation // Journal of Quaternary Science. V. 8. № 2. P. 95–108.
Marburger R.G., Robinson R.M., Thomas J.W., Andregg M.J., Clark. K.A., 1972. Antler malformation produced by leg injury in white-tailed deer // Journal of Wildlife Diseases. V. 8. № 4. P. 311–314.
Moore W.H., 1931. Notes on Antler Growth of Cervidae // Journal of Mammalogy. V. 12. № 2. P. 169–170.
Penrose C.B., 1924. Removal of the testicle in a sika deer followed by deformity of the antler on the opposite side // Journal of Mammalogy. V. 5. № 2. P. 116–118.
Pitulko V.V., Nikolsky P.A., Girya E.Y., Basilyan A.E., Tumskoy V.E., Koulakov S.A., Astakhov S.N., Pavlova E.Y., Anisimov M.A., 2004. The Yana RHS Site: Humans in the Arctic before the Last Glaciation // Science. V. 303. P. 52–56.
Scott W.B., 1885. Cervalces americanus, a Fossil Moose, or Elk, from the Quaternary of New Jersey // Proceedings of the Academy of Natural Sciences of Philadelphia. V. 37. P. 181–202.
Seton E.T., 1937. Hoofed Animals // Lives of game animals. V. 3. New York: Literary Guild of America. 780 p.
Sher A.V., 1986. Olyorian Land Mammal Age of Northeastern Siberia // Palaeontographia Italica. V. 74. P. 97–112.
Sher A.V., 1986a. On the history of mammal fauna of Beringida // Quartärpaläontologie. V. 6. P. 185–193.
Sher A.V., 1987. History and Evolution of Moose in USSR // Proceedings of the Second International Moose Symposium. Swedish Wildlife Research Suppl. 1. Part 1. P. 71–97.
Vislobokova I., Dmitrieva E., Kalmykov N., 1995. Artiodactyls from the Pliocene of Udunga, Western Trans-Baikal, Russia // Journal of Vertebrate Paleontology. V. 15. № 1. P. 146–159.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал