

Зоологический журнал, 2019, T. 98, № 10, стр. 1156-1167
Морфологическая дифференциация лошадей (Equus ovodovi, Equus hemionus) и границы их обитания в Западной Сибири в позднем плейстоцене
Н. А. Пластеева a, *, С. К. Васильев b, **, А. М. Клементьев c, ***, П. А. Косинцев a, d, ****
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, Россия
b Институт археологии и этнографии СО РАН
630090 Новосибирск, Россия
c Институт земной коры СО РАН
664033 Иркутск, Россия
d Уральский федеральный университет
620002 Екатеринбург, Россия
* E-mail: natalya-plasteeva@yandex.ru
** E-mail: svasiliev@archaeology.nsc.ru
*** E-mail: klem-al@yandex.ru
**** E-mail: kpa@ipae.uran.ru
Поступила в редакцию 01.03.2019
После доработки 15.04.2019
Принята к публикации 20.04.2019
Аннотация
Исследованы изменчивость размеров костей конечностей и особенности распространения двух видов мелкой формы лошади (род Equus L.) – лошади Оводова (E. ovodovi Eisenmann et Vasiliev) и кулана (E. hemionus Pallas), обитавших в Западной Сибири в позднем плейстоцене. Показано, что метрические признаки во многих случаях позволяют достаточно надежно различать эти два вида. Анализ ископаемого материала свидетельствует, что ареалы двух видов в прошлом не перекрывались. Ареал лошади Оводова охватывал территорию юго-востока Западной Сибири – Приобское плато, Алтай и Кузнецкий Алатау. Находки кулана происходят из юго-западных и юго-восточных районов Западной Сибири, что указывает на узкое распространение вида в регионе в позднем плейстоцене.
Для позднего плейстоцена Северной Евразии характерно совместное обитание двух форм лошадей, крупной и мелкой, которые принадлежали к разным подродам рода Equus L. 1758. Крупная форма лошадей – это представители подрода Equus s. str., или Настоящие лошади. Этот подрод в позднем плейстоцене включал вид Equus ferus Boddaert 1785 (Kahlke, 1999; Пластеева и др., 2012; van Asperen et al., 2012; Пластеева, Клементьев, 2017). Мелкая форма лошади в разных частях Северной Евразии была представлена тремя разными видами: европейским плейстоценовым ослом (E. (Asinus) hydruntinus Regalia 1904), куланом (E. (Hemionus) hemionus Pallas 1775) и лошадью Оводова (E. (Sussemionus) ovodovi Eisenmann et Vasiliev 2011). Кулан иногда включается в подрод Asinus (Groves, Grubb, 2011). В настоящее время кулан обитает в Передней, Средней и Восточной Азии (Moehlman, 2002), плейстоценовый осел и лошадь Оводова вымерли в разное время в прошлом.
Ареал E. hydruntinus занимал южные районы Западной и Восточной Европы (Громова, 1965; Uerpmann, 1976; van Asperen et al., 2012), а также Переднюю Азию (Orlando et al., 2006), тогда как два других вида мелкой формы лошади были характерны для азиатской части субконтинента. Кулан в позднем плейстоцене обитал в Средней Сибири (Мотузко, Орешников, 2005), Прибайкалье и Забайкалье (Вангенгейм 1977; Каспаров, 1986; Иметхенов, Калмыков, 1988; Клементьев, 2008), в Монголии и Китае (Boule et al., 1928; Dong et al., 1999; Оводов, 2001).
Лошадь Оводова – реликтовый вид, доживший на юго-востоке Западной Сибири до второй половины позднего плейстоцена. Костные остатки этого вида довольно многочисленны в отложениях Приобского плато, Алтая и Кузнецкого Алатау (Orlando et al., 2009; Eisenmann, Vasiliev, 2011; Plasteeva et al., 2015). По молекулярно-генетическим данным лошадь Оводова принадлежит к группе некабаллоидных лошадей, однако не является родственной формой ныне живущим ее представителям (Vilstrup et al., 2013; Druzhkova et al., 2017).
Видовая дифференциация лошадей по морфологическим признакам затруднена, поэтому точные границы ареалов отдельных видов в прошлом до сих пор остаются малоизученными. Например, ранее для территории Западной Сибири описаны находки плейстоценового осла (Форонова, 1990; Деревянко и др., 2003), кулана (Форонова, 1990) и лошади Оводова (Eisenmann, Vasiliev, 2011; Plasteeva et al., 2015). Присутствие E. hydruntinus в Сибири позднее не подтвердилось (Форонова, 2001; Foronova, 2002; Druzhkova et al., 2017). В настоящее время остается неясным, где проходила восточная граница распространения лошади Оводова в позднем плейстоцене и перекрывались ли ареалы E. ovodovi и E. hemionus в это время.
В данной работе изучен новый ископаемый материал по мелким лошадям из местонахождений Западной Сибири. Нами проведен сравнительный анализ морфологических признаков на костях конечностей E. ovodovi и E. hemionus, обобщены диагностические признаки, в том числе на элементах скелета, которые прежде не рассматривались. Полученные данные позволили провести видовую диагностику костей позднеплейстоценовых лошадей и, как результат, оценить границы обитания кулана и лошади Оводова в Западной Сибири.
МАТЕРИАЛЫ И МЕТОДЫ
Район исследования включает юг Западно-Сибирской равнины, горные районы Алтая и Кузнецкого Алатау и, таким образом, ограничен с запада и востока бассейнами рек Тобол и Енисей.
Костные остатки лошади Оводова в настоящее время описаны из девяти местонахождений на юго-востоке региона (рис. 1), в которых они представлены совместно с крупной лошадью E. ferus. Все известные точки находок позднеплейстоценового кулана расположены на территории Приенисейской Сибири (Foronova, 2002; Мотузко, Орешников, 2005), тогда как в остальной части исследуемого региона достоверных данных о присутствии вида нет.
Рис. 1.
Карта местонахождений с позднеплейстоценовыми находками лошадей Западной Сибири. Equus sp.: 1 – Афонтова Гора II, 2 – Красный Яр, 3 – Куртак, 4 – Омск, 5 – Рахимовское I, 6 – Чумыш; E. ovodovi: 7 – Проскурякова грот, 8 – Логово Гиены пещера, 9 – Окладникова пещера, 10 – Тараданово, 11 – Чик, 12 – Денисова пещера, 13 – Каминная пещера, 14 – Чагырская пещера, 15 – Страшная пещера.
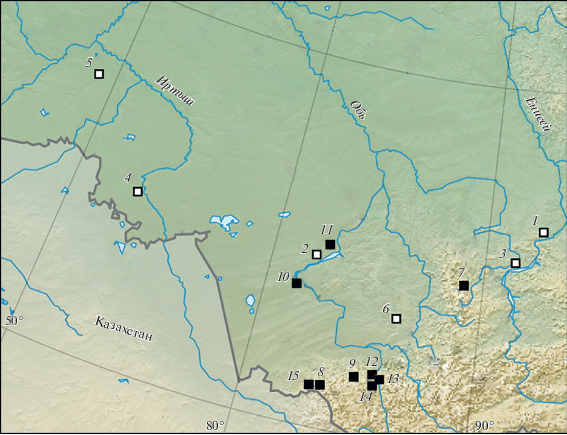
Материалом настоящей работы послужили выборки костей мелкой лошади из 6 новых и уже известных местонахождений региона: Афонтова Гора II, Красный Яр, Куртак, Рахимовское I, Омск и Чумыш (рис. 1, табл. 1). Предыдущие находки мелкой формы лошади из Куртака были отнесены к E. hemionus (Foronova, 2002), а новые материалы из Куртака, Афонтовой Горы II, Красного Яра, Рахимовского I, Омска и Чумыша не изучены или не были определены до вида (Бондарев, Пластеева, 2016).
Таблица 1.
Общее число элементов скелета Equus sp., E. ovodovi, E. ferus и E. hemionus, использованных в работе
| Элемент скелета | Equus sp. | E. ovodovi | E. ferus | E. hemionus | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7–11 | 7–11 | П | Г | Р | |
| Astragalus | 1 | 1 | 1 | – | 1 | 1 | 53 | 96 | 9 | 117 | 15 |
| Mc III | – | 1 | 24 | – | – | – | 29 | 141 | 10 | 32 | 16 |
| Mt III | 2 | – | 18 | – | – | 1 | 24 | 115 | 14 | 47 | 16 |
| Phalanx I | 2 | 2 | 11 | 1 | – | 1 | 19 | 92 | 9 | 169 | 30 |
Костный материал из всех местонахождений, за исключением Афонтовой Горы II, происходит из аллювиальных или субаэральных отложений и поэтому имеет широкий хронологический интервал. Радиоуглеродные даты по костям животных из большинства местонахождений попадают в пределы изотопно-кислородных стадий (ИКС) 3 и 2, т.е. 58–11.7 тыс. л.н. (Кузьмин и др., 2011; Лобачев и др., 2012). Стоянка Афонтова Гора II датируется 14–12 тыс. л.н. (Кузьмин и др., 2011; Артемьев и др., 2013), что соответствует второй половине ИКС 2 (17–11.7 тыс. л.н., позднеледниковье).
Для видовой диагностики костных остатков проанализированы морфологические признаки на костях конечностей всех видов лошадей – E. ferus, E. ovodovi и E. hemionus, которые могли обитать в регионе в позднем плейстоцене. В сравнительном анализе использованы выборки позднеплейстоценовых E. ferus и E. ovodovi из пяти местонахождений: грота Проскурякова, пещер Логово Гиены и Окладникова, Тараданово и Чика (табл. 1, рис. 1). Костные остатки E. ovodovi из других пещер Алтая (рис. 1) в большинстве своем сильно раздроблены и малопригодны для морфометрического анализа (Васильев, 2013; Васильев, Зенин, 2009; Васильев и др., 2006; Васильев и др., 2010).
Коллекции E. hemionus происходят из местонахождений Южной Сибири второй половины позднего плейстоцена (Клементьев, 2008; 2013; Щетников и др., 2015), из археологических стоянок Прикаспийской низменности, датируемых от неолита до раннего железного века, 7–3 тыс. л.н., а также рецентных сборов с территории Монголии и Туркмении. Монгольских и туркменских куланов относят к разным подвидам – E. h. hemionus и E. h. kulan соответственно. Таким образом, выборки кулана включают экземпляры с большой части ареала и разного возраста, что позволяет уменьшить влияние внутривидовой изменчивости.
Объекты исследования – таранная (astragalus), пястная (metacarpale III, Mc III), плюсневая (metatarsale III, Mt III) кости, передние и задние первые фаланги (phalanx I, ph I ant. et post.).
Весь ископаемый материал хранится в фондах Института археологии и этнографии СО РАН, Новосибирск; Института земной коры СО РАН, Иркутск; Зоологического института (ЗИН) РАН, Санкт-Петербург; Музея Института экологии растений и животных (ИЭРЖ) УрО РАН, Екатеринбург; Музея Бурятского научного центра СО РАН, Улан-Удэ, и Омского регионального отделения РГО, Омск; субфоссильный и рецентный материал – в фондах ИЭРЖ УрО РАН и ЗИН РАН.
При проведении сравнительного анализа морфологические различия между E. ferus, E. ovodovi и E. hemionus оценивались по комплексу метрических и неметрических признаков. В работе рассматривались целые и фрагментированные кости с приросшими эпифизами, принадлежавшие половозрелым животным. Половой диморфизм не учитывался, т.к. он не проявляется на костях дистальных отделов конечностей лошадей (van Asperen, 2013). Ввиду фрагментированности части костного материала размеры выборок для разных метрических признаков различаются.
Измерение морфометрических признаков костей выполнено по общепринятой методике (Eisenmann et al., 1988). С каждой пястной и плюсневой кости были сняты 9 промеров: 1 – наибольшая длина, 3 – ширина диафиза, 4 – поперечник диафиза, 5 – ширина проксимального конца, 6 – поперечник проксимального конца, 10 – ширина дистального конца в буграх, 11 – ширина дистального конца в суставе, 12 – поперечник дистального конца на гребне, 14 – наибольший поперечник медиального мыщелка дистального конца. На таранной кости были измерены 6 признаков: 1 – наибольшая длина, 2 – длина медиального гребня, 3 – ширина между гребнями, 4 – наибольшая ширина кости, 5 – ширина дистального сустава, 6 – поперечник дистального сустава. На фалангах измеряли 6 признаков: 1 – наибольшая длина, 2 – сагиттальная длина, 3 – ширина диафиза, 4 – ширина проксимального конца, 5 – поперечник проксимального конца, 6 – ширина дистального конца. Относительные размеры костей (индексы) высчитывались по стандартным формулам (Громова, 1949). Все измерения костей выполнены авторами, за исключением выборки рецентного монгольского кулана, размеры которого приведены по данным V. Eisenmann [vera-eisenmann.com].
Анализ морфометрических признаков костей проводился методами описательной и многомерной (метод главных компонент, ГК) статистики, а также с помощью двумерных диаграмм. Диаграммы были построены для всех вариантов сочетаний метрических признаков, из них выбраны и представлены те, что демонстрируют наибольшее разделение разных видов. Результаты анализа ГК в тексте представлены лишь для целых пястных и таранных костей, на которых представлены изучаемые признаки (№ 1, 3–6, 11–12 для mc III, № 1–6 для astragalus). Анализ данных выполнен в программах Statistica 5.0 (StatSoft Inc., USA) и Microsoft Excel.
РЕЗУЛЬТАТЫ
Основные морфологические признаки на костях конечностей у E. ovodovi и E. hemionus являются для них общими, что отражает их принадлежность к группе некабаллоидных лошадей, и хорошо отличают эти два вида от E. ferus (табл. 2). Кости конечностей у двух видов мелкой формы лошади грацильные и характеризуются низкими значениями индекса ширины диафиза; пястная и плюсневая кости со слабо выступающим сагиттальным гребнем, их ширина в дистальном суставе не превышает ширину в надсуставных буграх.
Таблица 2.
Основные диагностические признаки на костях конечностей лошадей
| Признак | E. ferus | E. ovodovi | E. hemionus |
|---|---|---|---|
| Пястная и плюсневая кости грацильные (M/Lim 3 : 1, %) | Нет mc: 16.6/14.7–18.7 mt: 13.1/10.6–15.7 |
Да mc: 13.9/12.8–14.9 mt: 11.4/10.5–12.5 |
Да mc: 12.3/10.9–13.6 mt: 10.1/8.8–11.2 |
| Ширина Mc III/Mt III в надсуставных буграх | < ширины дистального конца | > или = ширине дистального конца | > или = ширине дистального конца |
| Задненаружный отросток на проксимальном конце Mt III | Развит сильно | Развит слабо | Развит слабо |
| Выступание сагиттального гребня на дистальном конце Mc III/Mt III (M/Lim 14 : 12, %) | Сильное mc: 82.8/77.1–87.0 mt: 82.9/73.8–87.3 |
Слабое mc: 88.7/84.7–92.8 mt: 86.8/84.4–90.6 |
Слабое mc: 88.4/81.8–93.8 mt: 87.7/81.8–93.2 |
| Фаланги I грацильные (Lim 3 : 1, %) |
Нет ant.: 43.6/38.5–47.1 post.: 43.9/38.5–49.0 |
Да ant.: 34.3/32.2–36.8 post.: 34.5/32.9–35.5 |
Да ant.: 32.0/29.2–37.2 post.: 33.0/29.5–37.6 |
| Диафиз фаланг I в сечении | Овальный | Округлый | Округлый |
Пределы изменчивости абсолютных значений признаков костей E. ovodovi и E. hemionus в значительной степени совпадают (табл. 3), однако для костей E. ovodovi в среднем характерны более крупные размеры и большая массивность. Если абсолютные размеры костей подвержены изменчивости, то их относительные размеры достаточно стабильны и могут служить диагностирующим признаком. Так, значениe индекса ширины диафиза пястной кости у изученных экземпляров E. ovodovi варьирует от 12.8 до 14.9%, а в выборке E. hemionus – от 10.9 до 13.6% (табл. 2).
Таблица 3.
Абсолютные (мм) размеры костей конечностей E. ferus, E. ovodovi и E. hemionus (N, M/Lim)
| Признак | E. ferus | E. ovodovi | E. hemionus | ||
|---|---|---|---|---|---|
| П | Г | Р | |||
| Пястная кость Mc III | |||||
| 1 | $\frac{{134,228.5}}{{208.2{\kern 1pt} --{\kern 1pt} 250.7}}$ | $\frac{{16,{\text{ }}231.5}}{{219.0{\kern 1pt} --{\kern 1pt} 250.3}}$ | $\frac{{8,{\text{ }}227.2}}{{217.6{\kern 1pt} --{\kern 1pt} 236.5}}$ | $\frac{{3,{\text{ }}233.8}}{{225.5{\kern 1pt} --{\kern 1pt} 240.9}}$ | $\frac{{16,{\text{ }}226.2}}{{212.1{\kern 1pt} --{\kern 1pt} 240.0}}$ |
| 3 | $\frac{{132,{\text{ }}38.0}}{{32.8{\kern 1pt} --{\kern 1pt} 44.8}}$ | $\frac{{17,{\text{ }}32.1}}{{28.8{\kern 1pt} --{\kern 1pt} 36.0}}$ | $\frac{{8,{\text{ }}29.6}}{{28.7{\kern 1pt} --{\kern 1pt} 31.0}}$ | $\frac{{3,{\text{ }}29.6}}{{27.3{\kern 1pt} --{\kern 1pt} 30.9}}$ | $\frac{{16,{\text{ }}27.0}}{{24.2{\kern 1pt} --{\kern 1pt} 29.0}}$ |
| 11 | $\frac{{137,{\text{ }}52.5}}{{47.5{\kern 1pt} --{\kern 1pt} 58.2}}$ | $\frac{{23,{\text{ }}43.8}}{{40.6{\kern 1pt} --{\kern 1pt} 48.0}}$ | $\frac{{10,{\text{ }}40.6}}{{37.6{\kern 1pt} --{\kern 1pt} 42.9}}$ | $\frac{{22,{\text{ }}41.0}}{{38.2{\kern 1pt} --{\kern 1pt} 43.9}}$ | $\frac{{16,{\text{ }}39.7}}{{36.7{\kern 1pt} --{\kern 1pt} 44.0}}$ |
| 12 | $\frac{{135,{\text{ }}38.7}}{{33.2{\kern 1pt} --{\kern 1pt} 42.2}}$ | $\frac{{22,{\text{ }}33.3}}{{30.2{\kern 1pt} --{\kern 1pt} 36.6}}$ | $\frac{{9,{\text{ }}30.9}}{{28.2{\kern 1pt} --{\kern 1pt} 33.2}}$ | $\frac{{22,{\text{ }}31.3}}{{29.7{\kern 1pt} --{\kern 1pt} 34.1}}$ | $\frac{{16,{\text{ }}31.1}}{{29.2{\kern 1pt} --{\kern 1pt} 33.5}}$ |
| Плюсневая кость Mt III | |||||
| 1 | $\frac{{109,{\text{ }}272.0}}{{246.6{\kern 1pt} --{\kern 1pt} 301.2}}$ | $\frac{{14,{\text{ }}268.9}}{{257.2{\kern 1pt} --{\kern 1pt} 276.5}}$ | $\frac{{10,{\text{ }}268.5}}{{249.8{\kern 1pt} --{\kern 1pt} 284.6}}$ | $\frac{{3,{\text{ }}267.0}}{{258.6{\kern 1pt} --{\kern 1pt} 277.9}}$ | $\frac{{15,{\text{ }}263.4}}{{248.8{\kern 1pt} --{\kern 1pt} 277.0}}$ |
| 3 | $\frac{{109,{\text{ }}35.7}}{{29.0{\kern 1pt} --{\kern 1pt} 43.2}}$ | $\frac{{14,{\text{ }}30.6}}{{28.2{\kern 1pt} --{\kern 1pt} 33.2}}$ | $\frac{{11,{\text{ }}28.4}}{{26.2{\kern 1pt} --{\kern 1pt} 31.2}}$ | $\frac{{3,{\text{ }}27.5}}{{24.4{\kern 1pt} --{\kern 1pt} 29.5}}$ | $\frac{{16,{\text{ }}25.7}}{{22.9{\kern 1pt} --{\kern 1pt} 28.0}}$ |
| 11 | $\frac{{112,{\text{ }}53.3}}{{45.0{\kern 1pt} --{\kern 1pt} 64.2}}$ | $\frac{{23,{\text{ }}43.4}}{{37.5{\kern 1pt} --{\kern 1pt} 47.4}}$ | $\frac{{13,{\text{ }}40.8}}{{37.1{\kern 1pt} --{\kern 1pt} 45.0}}$ | $\frac{{46,{\text{ }}39.9}}{{36.2{\kern 1pt} --{\kern 1pt} 43.8}}$ | $\frac{{16,{\text{ }}38.0}}{{34.9{\kern 1pt} --{\kern 1pt} 41.1}}$ |
| 12 | $\frac{{113,{\text{ }}39.9}}{{32.2{\kern 1pt} --{\kern 1pt} 45.8}}$ | $\frac{{22,{\text{ }}33.7}}{{31.4{\kern 1pt} --{\kern 1pt} 36.6}}$ | $\frac{{14,{\text{ }}31.6}}{{28.8{\kern 1pt} --{\kern 1pt} 33.7}}$ | $\frac{{44,{\text{ }}31.1}}{{28.2{\kern 1pt} --{\kern 1pt} 33.6}}$ | $\frac{{16,{\text{ }}31.5}}{{29.6{\kern 1pt} --{\kern 1pt} 34.0}}$ |
| Таранная кость Astragalus | |||||
| 1 | $\frac{{96,{\text{ }}62.5}}{{52.7{\kern 1pt} --{\kern 1pt} 77.7}}$ | $\frac{{53,{\text{ }}57.0}}{{50.5{\kern 1pt} --{\kern 1pt} 61.5}}$ | $\frac{{8,{\text{ }}53.7}}{{50.9{\kern 1pt} --{\kern 1pt} 56.6}}$ | $\frac{{114,{\text{ }}54.0}}{{46.8{\kern 1pt} --{\kern 1pt} 59.1}}$ | $\frac{{15,{\text{ }}53.3}}{{47.0{\kern 1pt} --{\kern 1pt} 58.0}}$ |
| 3 | $\frac{{96,{\text{ }}32.2}}{{27.6{\kern 1pt} --{\kern 1pt} 38.1}}$ | $\frac{{52,{\text{ }}28.7}}{{24.9{\kern 1pt} --{\kern 1pt} 32.8}}$ | $\frac{{8,{\text{ }}26.3}}{{23.6{\kern 1pt} --{\kern 1pt} 28.7}}$ | $\frac{{116,{\text{ }}25.4}}{{21.8{\kern 1pt} --{\kern 1pt} 31.4}}$ | $\frac{{15,{\text{ }}24.8}}{{23.0{\kern 1pt} --{\kern 1pt} 28.0}}$ |
| Фаланга I передняя Ph I ant. | |||||
| 1 | $\frac{{45,{\text{ }}90.1}}{{81.7{\kern 1pt} --{\kern 1pt} 100.5}}$ | $\frac{{12,{\text{ }}86.4}}{{79.6{\kern 1pt} --{\kern 1pt} 95.5}}$ | $\frac{{2,{\text{ }}--}}{{83.1{\kern 1pt} --{\kern 1pt} 94.7}}$ | 1, 89.8 | $\frac{{15,{\text{ }}80.9}}{{68.8{\kern 1pt} --{\kern 1pt} 90.0}}$ |
| 3 | $\frac{{46,{\text{ }}39.3}}{{34.5{\kern 1pt} --{\kern 1pt} 43.7}}$ | $\frac{{13,{\text{ }}29.5}}{{27.9{\kern 1pt} --{\kern 1pt} 32.4}}$ | $\frac{{2,{\text{ }}--}}{{25.4{\kern 1pt} --{\kern 1pt} 28.6}}$ | $\frac{{88,{\text{ }}27.0}}{{22.9{\kern 1pt} --{\kern 1pt} 30.9}}$ | $\frac{{13,{\text{ }}26.0}}{{24.0{\kern 1pt} --{\kern 1pt} 29.0}}$ |
| 4 | $\frac{{44,{\text{ }}59.1}}{{53.0{\kern 1pt} --{\kern 1pt} 66.0}}$ | $\frac{{12,{\text{ }}46.9}}{{44.0{\kern 1pt} --{\kern 1pt} 51.9}}$ | $\frac{{2,{\text{ }}--}}{{45.2{\kern 1pt} --{\kern 1pt} 46.9}}$ | $\frac{{88,{\text{ }}42.2}}{{38.1{\kern 1pt} --{\kern 1pt} 45.7}}$ | $\frac{{15,{\text{ }}41.8}}{{39.1{\kern 1pt} --{\kern 1pt} 45.0}}$ |
| 6 | $\frac{{45,{\text{ }}48.4}}{{42.9{\kern 1pt} --{\kern 1pt} 53.6}}$ | $\frac{{13,{\text{ }}39.7}}{{36.4{\kern 1pt} --{\kern 1pt} 43.8}}$ | $\frac{{2,{\text{ }} - }}{{37.3{\kern 1pt} --{\kern 1pt} 39.8}}$ | $\frac{{68,{\text{ }}35.8}}{{32.7{\kern 1pt} --{\kern 1pt} 39.2}}$ | $\frac{{15,{\text{ }}35.3}}{{33.7{\kern 1pt} --{\kern 1pt} 37.5}}$ |
| Фаланга I задняя Ph I post. | |||||
| 1 | $\frac{{43,{\text{ }}86.0}}{{78.0{\kern 1pt} --{\kern 1pt} 91.6}}$ | $\frac{{6,{\text{ }}83.2}}{{80.0{\kern 1pt} --{\kern 1pt} 87.1}}$ | $\frac{{7,{\text{ }}82.5}}{{73.4{\kern 1pt} --{\kern 1pt} 94.7}}$ | 1, 89.3 | $\frac{{15,{\text{ }}74.1}}{{65.2{\kern 1pt} --{\kern 1pt} 82.0}}$ |
| 3 | $\frac{{45,{\text{ }}37.8}}{{32.7{\kern 1pt} --{\kern 1pt} 43.2}}$ | $\frac{{6,{\text{ }}28.7}}{{28.0{\kern 1pt} --{\kern 1pt} 30.1}}$ | $\frac{{7,{\text{ }}26.2}}{{22.9{\kern 1pt} --{\kern 1pt} 28.7}}$ | $\frac{{70,{\text{ }}25.2}}{{21.5{\kern 1pt} --{\kern 1pt} 28.4}}$ | $\frac{{15,{\text{ }}24.8}}{{22.5{\kern 1pt} --{\kern 1pt} 27.0}}$ |
| 4 | $\frac{{46,{\text{ }}59.5}}{{51.9{\kern 1pt} --{\kern 1pt} 66.3}}$ | $\frac{{6,{\text{ }}48.2}}{{45.3{\kern 1pt} --{\kern 1pt} 52.3}}$ | $\frac{{7,{\text{ }}42.8}}{{38.0{\kern 1pt} --{\kern 1pt} 47.0}}$ | $\frac{{67,{\text{ }}42.2}}{{37.8{\kern 1pt} --{\kern 1pt} 46.6}}$ | $\frac{{15,{\text{ }}42.1}}{{38.0{\kern 1pt} --{\kern 1pt} 45.0}}$ |
| 6 | $\frac{{47,{\text{ }}45.7}}{{35.6{\kern 1pt} --{\kern 1pt} 50.5}}$ | $\frac{{6,{\text{ }}38.7}}{{36.9{\kern 1pt} --{\kern 1pt} 41.7}}$ | $\frac{{7,{\text{ }}35.6}}{{31.7{\kern 1pt} --{\kern 1pt} 41.2}}$ | $\frac{{53,{\text{ }}33.7}}{{30.3{\kern 1pt} --{\kern 1pt} 36.9}}$ | $\frac{{15,{\text{ }}32.5}}{{30.4{\kern 1pt} --{\kern 1pt} 35.1}}$ |
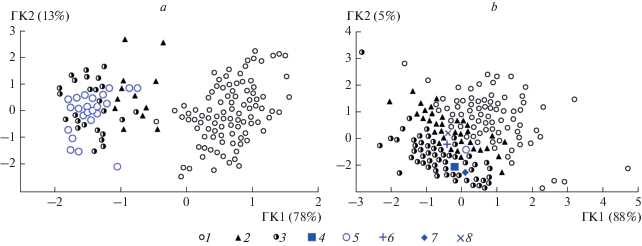
Результаты многомерного анализа не позволяют различать кости E. ovodovi и E. hemionus со 100% вероятностью (рис. 2). По совокупности размерных признаков костей конечностей изученные выборки образуют две хорошо очерченные группы: кабаллоидные (E. ferus) и некабаллоидные (E. ovodovi, E. hemionus) лошади. Кости мелких лошадей из новых местонахождений по результатам анализа попадают в группу некабаллоидных лошадей. Анализ для разных элементов скелета демонстрирует сходные результаты, отличия наблюдаются лишь в степени перекрывания значений признаков у двух групп. Наибольшее перекрывание облаков выражено для таранной кости, что отражает ее меньшую изменчивость по сравнению с другими элементами скелета. Различия между двумя группами проявляются по оси ГК 1, которая связана со всеми признаками пястной кости, за исключением ее длины, и всеми признаками таранной кости, кроме ее ширины.
Рис. 2.
Результаты анализа пястных (a) и таранных (b) костей методом главных компонент: 1 – E. ferus, 2 – E. ovodovi, 3 – E. hemionus, 4 – Equus sp. (Афонтова Гора II), 5 – Equus sp. (Куртак), 6 – Equus sp. (Рахимовское I), 7 – Equus sp. (Чумыш), 8 – Equus sp. (Красный Яр).
При сходном характере распределения в осях ГК экземпляры, принадлежащие кулану, отличаются от лошади Оводова более мелкими размерами. Различие в размерах костей E. hemionus и E. ovodovi хорошо видно на двумерных диаграммах. Несмотря на существование зоны перекрывания значений признаков у двух видов, большинство экземпляров надежно дифференцируется (рис. 3). Большая часть пястных и плюсневых костей из Куртака, а также плюсневая кость из Афонтовой Горы II располагаются в области значений размерных признаков E. hemionus. Несколько пястных и плюсневых костей из Куртака попадают в зону перекрывания значений признаков E. ovodovi и E. hemionus и могут представлять собой крайние варианты изменчивости признаков этих видов. Лишь две плюсневые кости из Куртака и плюсневая кость из Чумыша по размерам и пропорциям соответствуют E. ovodovi (табл. 4). Пястная кость из Красного Яра по своим абсолютным и относительным размерам (табл. 4) может быть отнесена к E. ovodovi.
Рис. 3.
Двумерный график рассеяния экземпляров пястных (a), плюсневых (b), таранных (c) костей и первых фаланг (d) лошадей относительно среднего значения (M ± σ): 1 – E. ovodovi, 2 – E. hemionus, 3 – Equus sp. (Куртак), 4 – Equus sp. (Афонтова Гора II), 5 – Equus sp. (Чумыш), 6 – Equus sp. (Омск), 7 – Equus sp. (Рахимовское I), 8 – Equus sp. (Красный Яр).
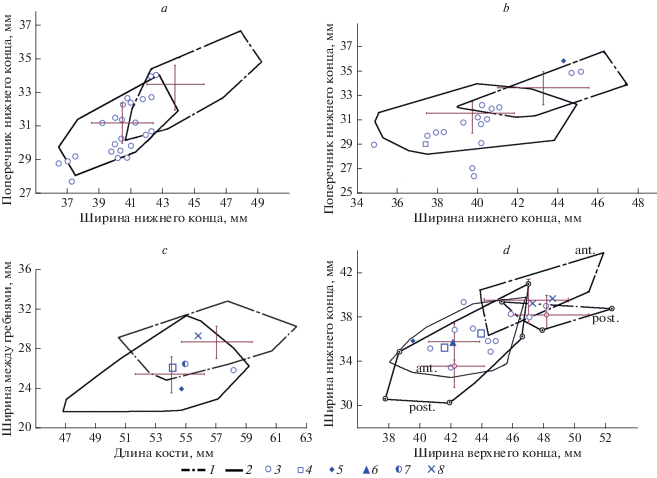
Таблица 4.
Абсолютные (мм) и относительные (%) размеры костей конечностей лошадей Западной Сибири (N, M/Lim)
| Выборка | Признак | ||||
|---|---|---|---|---|---|
| Mc III | 1 | 3 | 11 | 12 | 3 : 1 |
| E. hemionus Куртак | $\frac{{17,{\text{ }}223.9}}{{208.1{\kern 1pt} --{\kern 1pt} 231.8}}$ | $\frac{{17,{\text{ }}29.0}}{{27.0{\kern 1pt} --{\kern 1pt} 31.3}}$ | $\frac{{17,{\text{ }}39.7}}{{36.6{\kern 1pt} --{\kern 1pt} 41.1}}$ | $\frac{{17,{\text{ }}30.3}}{{27.8{\kern 1pt} --{\kern 1pt} 32.5}}$ | $\frac{{17,{\text{ }}12.9}}{{11.9{\kern 1pt} --{\kern 1pt} 14.3}}$ |
| Equus sp. Куртак | $\frac{{7,{\text{ }}230.6}}{{226.2{\kern 1pt} --{\kern 1pt} 234.5}}$ | $\frac{{7,{\text{ }}30.0}}{{28.2{\kern 1pt} --{\kern 1pt} 31.6}}$ | $\frac{{7,{\text{ }}42.0}}{{41.1{\kern 1pt} --{\kern 1pt} 42.8}}$ | $\frac{{7,{\text{ }}32.4}}{{30.5{\kern 1pt} --{\kern 1pt} 34.2}}$ | $\frac{{7,{\text{ }}13.0}}{{12.5{\kern 1pt} --{\kern 1pt} 13.6}}$ |
| E. ovodovi Красный Яр | 1, 217.6 | 1, 31.3 | 1, 45ca | – | 1, 14.4 |
| Mt III | 1 | 3 | 11 | 12 | 3 : 1 |
| E. hemionus Афонтова Гора II | – | 2, 23.6–26.2 | 1, 37.4 | 1, 29.0 | – |
| E. hemionus Куртак | $\frac{{13,{\text{ }}262.4}}{{208.1{\kern 1pt} --{\kern 1pt} 274.7}}$ | $\frac{{13,{\text{ }}27.3}}{{25.3{\kern 1pt} --{\kern 1pt} 29.0}}$ | $\frac{{13,{\text{ }}38.7}}{{34.9{\kern 1pt} --{\kern 1pt} 40.3}}$ | $\frac{{11,{\text{ }}29.5}}{{26.5{\kern 1pt} --{\kern 1pt} 31.3}}$ | $\frac{{13,{\text{ }}10.4}}{{9.6{\kern 1pt} --{\kern 1pt} 11.2}}$ |
| E. ovodovi Куртак | 2, 267.5–269.4 | 2, 32.2–32.6 | 2, 44.8–45.1 | 2, 35.0–35.1 | 2, 12.0–12.2 |
| Equus sp. Куртак | $\frac{{3,{\text{ }}259.3}}{{252.7{\kern 1pt} --{\kern 1pt} 271.6}}$ | $\frac{{3,{\text{ }}26.3}}{{25.4{\kern 1pt} --{\kern 1pt} 27.5}}$ | $\frac{{3,{\text{ }}40.6}}{{40.2{\kern 1pt} --{\kern 1pt} 41.0}}$ | $\frac{{3,{\text{ }}32.0}}{{31.8{\kern 1pt} --{\kern 1pt} 32.1}}$ | $\frac{{3,{\text{ }}10.2}}{{10.1{\kern 1pt} --{\kern 1pt} 10.3}}$ |
| E. ovodovi Чумыш | 1, 265.0 | 1, 29.1 | 1, 44.3 | 1, 36.3 | 1, 11.0 |
| Astragalus | 1 | 2 | 3 | 4 | 4 : 1 |
| E. hemionus Афонтова Гора II | 1, 54.1 | 1, 54.8 | 1, 26.1 | 1, 49.1 | 1, 90.8 |
| E. hemionus Куртак | 1, 58.3 | 1, 58.2 | 1, 26.1 | 1, 52.7 | 1, 90.4 |
| E. ovodovi Красный Яр | 1, 55.7 | 1, 56.6 | 1, 29.0 | 1, 56.0 | 1, 100.5 |
| E. hemionus Рахимовское I | 1, 54.9 | 1, 54.5 | 1, 26.6 | 1, 52.4 | 1, 95.4 |
| E. hemionus Чумыш | 1, 54.7 | 1, 55.1 | 1, 24.3 | 1, 54.0 | 1, 98.7 |
| Ph I ant. | 1 | 3 | 4 | 6 | 3 : 1 |
| E. hemionus Афонтова Гора II | 1, 85.9 | 1, 27.3 | 1, 42.3 | 1, 35.3 | 1, 31.8 |
| E. hemionus Куртак | 2, 84.9–88.2 | 2, 27.0–28.4 | 2, 40.9–43.2 | 2, 35.5–38.7 | 2, 31.8–32.2 |
| E. hemionus Омск | 1, 82ca | 1, 26.5 | 1, 42.2 | 1, 35.7 | 1, 32.3 |
| E. ovodovi Красный Яр | 1, 87.8 | 1, 30.2 | 1, 47.3 | 1, 40.0 | 1, 34.4 |
| Ph I post. | 1 | 3 | 4 | 6 | 3:1 |
| E. hemionus Афонтова Гора II | 1, 89.0 | 1, 26.6 | 1, 44.0 | 1, 36.7 | 1, 29.9 |
| E. hemionus Куртак | $\frac{{6,{\text{ }}77.1}}{{71.4{\kern 1pt} --{\kern 1pt} 82.7}}$ | $\frac{{6,{\text{ }}26.9}}{{26.5{\kern 1pt} --{\kern 1pt} 27.6}}$ | $\frac{{6,{\text{ }}43.7}}{{42.2{\kern 1pt} --{\kern 1pt} 44.6}}$ | $\frac{{6,{\text{ }}35.9}}{{33.7{\kern 1pt} --{\kern 1pt} 37.3}}$ | $\frac{{6,{\text{ }}35.0}}{{32.5{\kern 1pt} --{\kern 1pt} 37.1}}$ |
| Equus sp. Куртак | $\frac{{3,{\text{ }}81.9}}{{80.5{\kern 1pt} --{\kern 1pt} 82.9}}$ | $\frac{{3,{\text{ }}27.4}}{{26.6{\kern 1pt} --{\kern 1pt} 28.0}}$ | $\frac{{3,{\text{ }}47.3}}{{46.3{\kern 1pt} --{\kern 1pt} 48.3}}$ | $\frac{{3,{\text{ }}38.4}}{{38.0{\kern 1pt} --{\kern 1pt} 38.6}}$ | $\frac{{3,{\text{ }}33.5}}{{32.1{\kern 1pt} --{\kern 1pt} 34.8}}$ |
| E. hemionus Чумыш | 1, 81.5 | 1, 26.6 | 1, 39.5 | 1, 35.4 | 1, 32.6 |
| E. ovodovi Красный Яр | 1, 83.3 | 1, 27.9 | 1, 48.4 | 1, 38.9 | 1, 33.5 |
Таранные кости из Афонтовой Горы II и Рахимовского I располагаются в области перекрывания признаков двух видов, однако имеют минимальные для E. ovodovi размеры и в среднем более близки E. hemionus. С наибольшей вероятностью эти экземпляры могут быть отнесены к кулану (табл. 4). Таранная кость из Красного Яра по своим морфометрическим признакам более соответствует E. ovodovi.
Передние и задние фаланги из Афонтовой Горы II, Омска, Чумыша, а также большая часть фаланг из Куртака по своим размерам и пропорциям соответствуют кулану. Лишь две задние фаланги из Куртака, передняя и задняя фаланги из Красного Яра попадают в область значений E. ovodovi, еще одна фаланга из Куртака располагается в области перекрывания значений двух видов. Вместе с тем, по признаку относительной ширины диафиза один экземпляр из Куртака соответствует значениям E. hemionus, два других – попадают в область перекрывания двух видов. Необходимо также отметить, что эти кости относятся к задней конечности. Как уже было показано, пястные и плюсневые кости демонстрируют морфологические различия, связанные с функциональной ролью передней и задней конечностей лошадей (Громова, 1949; Eisenmann, Beckouche, 1986), и поэтому имеют разное значение для видовой диагностики. Можно предположить сходные проявления и на других элементах скелета. Вышеуказанные экземпляры фаланг из Куртака отнесены к Equus sp. (табл. 4) и их видовая принадлежность нуждается в определении методами молекулярно-генетического анализа.
Таким образом, большая часть костей из Куртака, Чумыша, все находки из Афонтовой Горы II, Омска и Рахимовского I с высокой вероятностью принадлежит кулану (рис. 4). К лошади Оводова отнесены две плюсневые кости из Куртака, плюсневая кость из Чумыша, и все экземпляры костей из Красного Яра. Пястные, плюсневые кости и задние фаланги лошадей из Куртака, попавших в область перекрывания признаков двух видов, по морфологическим показателям не могут быть определенно отнесены к конкретному виду.
Рис. 4.
Пястные (a), плюсневые (b–d), таранные (e–g) кости и первые фаланги (h–k) лошадей из позднеплейстоценовых отложений Западной Сибири: a – E. hemionus Куртак, b–c – E. hemionus Афонтова Гора II, d – E. ovodovi Куртак, e – E. hemionus Куртак, f – E. hemionus Рахимовское I, g – E. hemionus Чумыш, h–i – E. hemionus Афонтова Гора II, j – E. hemionus Омск, k – E. hemionus Чумыш.

ОБСУЖДЕНИЕ
Проведенный сравнительный анализ показал, что различия между двумя видами некабаллоидных лошадей – E. ovodovi и E. hemionus – проявляются в средних значениях и пределах изменчивости абсолютных и относительных размеров. Для костей конечностей E. ovodovi в среднем характерны более крупные размеры и большая массивность. Несмотря на то, что кулан и лошадь Оводова генетически заметно удалены друг от друга (Orlando et al., 2009; Vilstrup et al., 2013), морфологические различия между этими видами на костях конечностей являются размерными.
Все известные позднеплейстоценовые находки мелкой формы лошади Западной Сибири принадлежат двум видам – лошади Оводова и кулану. Костные остатки E. ovodovi многочисленны в местонахождениях юго-востока региона – на территории Приобского плато, Алтая и Кузнецкого Алатау и отсутствуют в юго-западной его части (Plasteeva et al., 2015). На остальной территории Северной Азии вид не известен. К востоку от изучаемого региона, в Южной Сибири, представлены ископаемые находки кулана (Вангенгейм 1977; Каспаров, 1986; Иметхенов, Калмыков, 1988; Клементьев, 2008), а на северо-востоке Сибири на протяжении позднего плейстоцена обитали только кабаллоидные лошади (Шер, 1971; Лазарев, 1980).
Данные о встречаемости костных остатков лошади Оводова только в южных районах (южнее 56-й параллели), а также данные о составе сопутствующих видов млекопитающих (Пластеева, Васильев, 2017) позволяют заключить, что этот вымерший вид был достаточно теплолюбивым и тяготел к открытым остепненным ландшафтам. В конце периода ИКС 3 лошадь Оводова исчезает из фауны Западной Сибири и в отложениях более позднего времени ее находки отсутствуют. Радиоуглеродные даты, полученные по костям E. ovodovi, попадают в интервал 45–23 тыс. л.н. (Orlando et al., 2009; Plasteeva et al., 2015), что указывает на вымирание вида до наступления последнего максимума похолодания.
Костные остатки кулана из изученных местонахождений Прииртышья и Предалтайской равнины немногочисленны. Наиболее представительный материал происходит из крайней восточной части исследуемого региона (Куртак, Афонтова Гора II), а также из других местонахождений Красноярского водохранилища (Мотузко, Орешников, 2005; Kuzmin, 2011). Все эти местонахождения, за исключением Куртака, датируются позднее 17 тыс. л.н. (Кузьмин и др., 2011; Артемьев и др., 2013).
Таким образом, две морфологически сходные формы лошадей – E. ovodovi и E. hemionus – не обитали симпатрично в Западной Сибири. В датированных отложениях совместные находки двух видов неизвестны. Лишь в двух местонахождениях (Куртак и Чумыш) обнаружены кости, морфологически соответствующие как кулану, так и лошади Оводова. Из этого следует, что оба вида могли обитать только на востоке изучаемого региона, но не известно – синхронно или в разные периоды позднего плейстоцена.
ЗАКЛЮЧЕНИЕ
В позднем плейстоцене на юге Западной Сибири кроме настоящих лошадей (подрод Equus) были представлены два вида мелкой формы лошади – лошадь Оводова (E. ovodovi) и кулан (E. hemionus). Ареалы этих двух видов не перекрывались. На юго-востоке Западной Сибири (Приобское плато, Алтай и Кузнецкий Алатау) до начала последнего максимума похолодания обитала лошадь Оводова. Находки кулана происходят из юго-западных и юго-восточных районов Западной Сибири, что указывает на узкое распространение вида в регионе в течение позднего плейстоцена. Оба вида принадлежат к группе некабаллоидных лошадей, морфологические различия между которыми проявляются в абсолютных и относительных размерах костей конечностей.
Список литературы
Артемьев Е.В., Дроздов Н.И., Макулов В.И., 2013. Основные этапы археологического изучения стоянки Афонтова Гора II // Известия Иркутского Государственного Университета. Серия Геоархеология. Этнология. Антропология. № 1. С. 58–71.
Бондарев А.А., Пластеева Н.А., 2016. Плейстоценовые лошади Прииртышья // Проблемы и перспективы палеонтологических исследований: Материалы LXII сессии Палеонтологического Обшества при РАН. СПб.: ВСЕГЕИ. С. 226–227.
Вангенгейм Э.А., 1977. Палеонтологическое обоснование стратиграфии антропогена Северной Азии (по млекопитающим). М.: Наука. 172 с.
Васильев С.К., 2013. Фауна крупных млекопитающих из плейстоценовых отложений Чагырской пещеры (Северо-Западный Алтай) по материалам раскопок 2007–2011 годов // Археология, этнография и антропология Евразии. № 1(53). С. 28–44.
Васильев С.К., Деревянко А.П., Маркин С.В., 2006. Фауна крупных млекопитающих финала сартанского времени Северо-Западного Алтая (по материалам пещеры Каминной) // Археология, этнография и антропология Евразии. № 2 (26). С. 2–22.
Васильев С.К., Зенин А.Н., 2009. Фаунистические остатки из пещеры Страшная (северо-западный Алтай) по материалам раскопок в 1988–2008 годах // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Т. 15. С. 56–62.
Васильев С.К., Шуньков М.В., Цыбанков А.А., 2010. Остатки крупных млекопитающих из плейстоценовых отложений в восточной галерее Денисовой пещеры (материалы 2009-2010 годов) // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Т. 16. С. 28–32.
Громова В., 1949. История лошадей (род Equus) в Старом Свете // Труды Зоологического института АН СССР. Т. 17. Вып. 1. С. 1–375.
Громова В.И., 1965. Краткий обзор четвертичных млекопитающих Европы (опыт сопоставления). М.: Наука. 142 с.
Деревянко А.П., Шуньков М.В., Агаджанян А.К., Барышников Г.Ф., Малаева Е.М., Ульянов В.А., Кулик Н.А., Постнов А.В., Анойкин А.А., 2003. Природная среда и человек в палеолите Горного Алтая. Новосибирск: Издательство ИАЭТ СО РАН. С. 214–219.
Иметхенов А.Б., Калмыков Н.П., 1988. Фауна и флора кайнозоя Бурятии. Улан-Удэ: Издательство БФ СО АН СССР. 181 с.
Каспаров А.К., 1986. Остатки млекопитающих из позднепалеолитического поселения Сухотино-4 в Забайкалье // Труды Зоологического института АН СССР. Т. 149. С. 98–106.
Клементьев А.М., 2008. Кулан (E. cf. hemionus) из позднего плейстоцена Забайкалья // Фауна и флора Северной Евразии в позднем кайнозое. Челябинск: Рифей. С. 186–198.
Клементьев А.М., 2013. Фауны позднекаргинского времени Иркутского амфитеатра // Известия Иркутского гос. ун-та. Серия Геоархеология. Этнология. Антропология. Т. 1. С. 30–43.
Кузьмин Я.В., Орлова Л.А., Зенин В.Н., Лбова Л.В., Дементьев В.Н., 2011. Радиоуглеродное датирование палеолита Сибири и Дальнего Востока России: материалы к каталогу 14C дат // Stratum Plus. V. 1. С. 171–200.
Лазарев П.А., 1980. Антропогеновые лошади Якутии. М.: Наука. 189 с.
Лобачев Ю. В., Васильев С. К., Орлова Л. А., 2012. Позднеплейстоценовая териофауна с р. Чумыш (Алтайский край) и новые данные по местонахождению на р. Чик (Новосибирская область) // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Новосибирск. Т. 18. С. 106–110.
Мотузко А.Н., Орешников И.А., 2005. Фауна крупных млекопитающих Дербинского археологического района // Древности Приенисейской Сибири. Вып. 4. Красноярск: РИО ГОУ ВПО ЛГПУ им. В.П. Астафьева. С. 23–31.
Оводов Н.Д., 2001. Плейстоценовая фауна пещеры Цаган Агуй (МНР) // Природные условия, история и культура Западной Монголии и сопредельных регионов: Сборник материалов конференции. Томск: Издательство ТГУ. С. 146–147.
Пластеева Н.А., Бурова Н.Д., Косинцев П.А., 2012. Лошади (Equus ferus Boddaert, 1785) Восточной Европы конца позднего плейстоцена // Бюллетень Московского общества испытателей природы. Отдел Биологический. Т. 117. Вып. 5. С. 3–11.
Пластеева Н.А., Васильев С.К., 2017. Фаунистическое окружение лошади Оводова в позднем плейстоцене // Фауна Урала и Сибири. Т. 2. С. 204–214.
Пластеева Н.А., Клементьев А.М., 2017. Позднеплейстоценовая лошадь Equus (Equus) ferus (Perissodactyla, Equidae) Прибайкалья и Забайкалья // Труды Зоологического института РАН. Т. 321. Вып. 2. С. 180–198.
Форонова И.В., 1990. Ископаемые лошади Кузнецкой котловины. Новосибирск: Институт геологии и геофизики СО АН СССР. 131 с.
Форонова И.В., 2001. Четвертичные млекопитающие юго-востока Западной Сибири (Кузнецкая котловина): филогения, биостратиграфия, палеоэкология. Новосибирск: СО РАН. С. 92–113.
Шер А.В., 1971. Млекопитающие и стратиграфия плейстоцена крайнего северо-востока СССР и Северной Америки. М.: Наука. С. 145–195.
Щетников А.А., Клементьев А.М., Филинов И.А., Семеней Е.Ю., 2015. Крупные млекопитающие опорных разрезов неоплейстоцена Тункинской рифтовой долины, юго-западное Прибайкалье // Стратиграфия. Геологическая корреляция. Т. 23. № 2. С. 104–128.
Boule M., Breuil H., Licent E., Teilhard de Chardin P., 1928. Le Paleolithique de la Chine // Archives de’l Institut de Paleontologie Humaine. Mem. 4. P. 1–102.
Dong W., Xu Q., Jin Ch., Liu J., 1999. Quaternary herbivore fauna in northeastern China: evolution under climatic change // Chinese Science Bulletin. V. 44. P. 129–132.
Druzhkova A. S., Makunin A.I., Vorobieva N.V., Vasiliev S.K., Ovodov N.D., Shunkov M.V., Trifonov V.A., Graphodatsky A.S., 2017. Complete mitochondrial genome of an extinct Equus (Sussemionus) ovodovi specimen from Denisova cave (Altai, Russia) // Mitochondrial DNA Part B. V. 2. P. 79–81.
Eisenmann V., 2010. Sussemionus, a new subgenus of Equus (Perissodactyla, Mammalia) // Comptes Rendus Biologies. V. 333. P. 235–240.
Eisenmann V., Alberdi M.T., de Giuli G., Staeche U., 1988. Studying fossil horses. V. 1: Methodology // Collected papers after the “New York International Hipparion Conference, 1981”. Leiden; New York; København; Köln. 71 p.
Eisenmann V., Beckouche S., 1986. Identification and Discrimination of metapodials from Pleistocene and modern Equus, wild and domestic // Equids in the ancient world. Eds Meadow R.H., Uerpmann H.-P. Wies-Baden: Reichert. P. 117–163.
Eisenmann V., Vasiliev S.K., 2011. Unexpected finding of a new Equus species (Perissodactyla, Mammalia) belonging to a supposedly extinct sub-genus in Late Pleistocene deposits of Khakassia (southwestern Siberia) // Geodiverisitas. V. 33. № 3. P. 519–530.
Equus actuels et récemment éteints / V. Eisenmann: URL: http://www.vera-eisenmann.com/equidae-equus-actuels-et-recemment-eteints-rubrique17.html (дата обращения 1.12.2017)
Foronova I.V., 2002. Late Quaternary equids (genus Equus) of south-western and south-central Siberia // Equids in time and space: 9th ICAZ conference. Durham. P. 20–30.
Groves C., Grubb P., 2011. Ungulate taxonomy. Baltimore, MD: John Hopkins University Press. 310 p.
Kahlke R.D., 1999. The history of the origin, evolution and dispersal of the Late Pleistocene Mammuthus-Coelodonta faunal complex in Eurasia (large mammals). Hot Springs: Mammoth Site of Hot Springs South Dakota Inc. 219 p.
Kuzmin Y.V., 2011. Mammalian fauna from Palaeolithic sites in the Upper Yenisei river basin (Southern Siberia): review of the current zooarchaeological evidence // International Journal of Osteoarchaeology. V. 21. P. 218–228.
Moehlman P. (ed.), 2002. Equids: Zebras, asses and horses. Status survey and conservation action plan. IUCN, Gland, Switzerland and Cambridge. P. 62–64.
Orlando L., Mashkour M., Burke A., Douady C.J., Eisenmann V., Hänni C., 2006. Geographic distribution of an extinct equid (Equus hydruntinus: Mammalia, Equidae) revealed by morphological and genetical analyses of fossils // Molecular Ecology. V. 15. P. 2083–2093.
Orlando L., Metcalf J.L, Alberdi M.T., Telles-Antunes M., Bonjean D., Otte M., Martin F., Eisenmann V., Mashkour M., Morello F., Prado J.L., Salas-Gismondi R., Shockey B.J., Wrinn P.J., Vasil’ev S.K. et al., 2009. Revising the recent evolutionary history of equids using ancient DNA // PNAS. V. 106. P. 21754–21759.
Plasteeva N.A., Vasiliev S.K., Kosintsev P.A., 2015. Equus (Sussemionus) ovodovi Eisenmann et Vasiliev, 2011 from the Late Pleistocene of Western Siberia // Russian Journal of Theriology. V. 14. P. 187–200.
Uerpmann H.-P., 1976. Equus (Equus) caballus und Equus (Asinus) hydruntinus im Postpleistozän der Iberischen Halbinsel (Perissodactyla, Mammalia) // Säugetierkundliche Mitteilungen. V. 24. P. 206–218.
van Asperen E.N., 2013. Implications of age variation and sexual dimorphism in modern Equids for middle Pleistocene Equid taxonomy // International Journal of Osteoarchaeology. V. 23. P. 1–12.
van Asperen, E.N., Stefaniak K., Proskurnyak I., Ridush B., 2012. Equids from Emine-Bair-Khosar Cave (Crimea, Ukraine): co-occurrence of the stenonid Equus hydruntinus and the caballoid E. ferus latipes based on skull and postcranial remains // Palaeontologia Electronica. V. 15. P. 5A.
Vilstrup J.T., Seguin-Orlando A., Stiller M., Ginolhac A., Raghavan M., Nielsen S.C.A., Weinstock J., Froese D., Vasiliev S.K., Ovodov N.D., Clary J., Helgen K.M., Fleischer R.C., Cooper A., Shapiro B., Orlando L., 2013. Mitochondrial phylogenomics of modern and ancient Equids // PLoS ONE. V. 8 (2): e55950.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал