

Зоологический журнал, 2019, T. 98, № 11, стр. 1233-1246
Добыча сов-миофагов (Strix nebulosa, Bubo bubo) как источник избирательного накопления палеотериологических материалов
Н. Г. Смирнов a, *, Ю. Э. Кропачева a, **, С. В. Зыков a, ***
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, Россия
* E-mail: nsmirnov@ipae.uran.ru
** E-mail: kropachevaje@yandex.ru
*** E-mail: svzykov@yandex.ru
Поступила в редакцию 20.02.2019
После доработки 11.03.2019
Принята к публикации 11.03.2019
Аннотация
Исследовано накопление остатков жертв из погадок бородатых неясытей в зимний и гнездовой периоды за ряд лет на контакте тайги и северной лесостепи Зауралья. Из 525 погадок определены остатки 2055 особей животных. Показано, какие из характеристик охотничьей территории и населения мелких млекопитающих в наибольшей степени влияют на состав и соотношение видов добычи. Проведено сравнение с результатами кормодобывающей деятельности филина. Использованы опубликованные данные по материалам из четырех мест многолетнего гнездования на скалах в долинах рек Западного склона Среднего Урала в аналогичных природных условиях. Показана избирательная добыча жертв двух видов сов. Основу рациона составляло небольшое количестве видов основных жертв, преимущественно населяющих открытые местообитания. У обоих видов сопутствующие жертвы – виды закрытых лесных (транзитных) биотопов. Для филина характерен больший размерный диапазон жертв. Особое внимание уделено индикаторной роли сопутствующих жертв при палеореконструкциях.
На основании данных о современном состоянии экосистем, сообществ животных и видов формируются представления о прошлых этапах их существования (Шер, 1999). За последние десятилетия палеонтологи существенно продвинулись в раскрытии механизмов накопления и трансформации костных остатков животных в местонахождениях (Terry, 2010; Andrews, Fernández-Jalvo, 2018; Comay, Dayan, 2018). Именно в тафономии знания о настоящем в наиболее явном виде используются палеонтологами для палеоэкологических реконструкций. Основу этих подходов в отечественной палеозоологии мелких млекопитающих заложил И.М. Громов в середине двадцатого века (1957). Одними из основных накопителей костных остатков мелких млекопитающих являются совы. В данной статье рассмотрены материалы по питанию двух видов сов-миофагов – бородатой неясыти (Strix nebulosa J.R. Forster 1772) и филина (Bubo bubo Linnaeus 1758). Основное внимание уделено изучению избирательного накопления их жертв.
Основными жертвами совы – накопителя материала могут становиться не самые многочисленные виды в природном сообществе. Показано, что у одного и того же вида сов в разных частях ареала в зависимости от доступности тех или иных видов жертв основные и альтернативные виды жертв различаются (Mikkola, 1983; Пукинский, 1993; Korpimäki, Marti, 1995; Zárybnická et al., 2009). Диапазон варьирования рациона наиболее широк у универсальных хищников. Они добывают самых многочисленных и доступных жертв предпочитаемого размера (Korpimäki, 1986; Korpimäki, Marti, 1995; Rutz, Bijlsma, 2006; Zárybnická et al., 2009).
В качестве основного объекта исследования выбрана бородатая неясыть. Благодаря тому, что этот вид является специализированным миофагом, анализ его рациона позволяет получить более четкую картину избирательности, чем при изучении питания универсальных хищников, и описать некоторые механизмы, лежащие в основе накопления костных остатков в орнитогенных отложениях.
Филин является универсальным хищником, на протяжении его ареала отмечены значительные изменения рациона, который включает очень широкий спектр жертв. Для отдельных частей ареала, однако, выделяются наиболее предпочитаемые жертвы, такие как зайцы, водяные полевки, слепушонки, серые полевки (Mikkola, 1983; Шепель, 1992; Пукинский, 1993; Карякин, 1998; Tobajas et al., 2016). Филин является наиболее частым накопителем костного материала по мелким млекопитающим на скальных обнажениях, широко представленных на Урале (Шепель, 1992; Смирнов, Садыкова, 2003).
Цель данной работы – охарактеризовать формирование пищевого спектра бородатой неясыти и филина как источника избирательного накопления палеонтологического материала. Для ее выполнения были поставлены следующие задачи: 1. Оценить различия в питании сов, занимавших разные гнезда, но в пределах участков с одинаковыми биотопическими характеристиками. Оценить межгодовую изменчивость питания. 2. Оценить различия в питании сов на участках с различными биотопическими характеристиками. 3. Оценить избирательность питания бородатой неясыти в изучаемом районе в сравнении с результатами отловов и литературными сведениями по фауне региона. 4. Охарактеризовать состав и структуру жертв филина из отдельных гнезд, расположенных в однотипном ландшафтном окружении, но со специфическим соотношением биотопов. 5. Оценить избирательность питания филина в изучаемом районе в сравнении с литературными данными по составу фауны и структуре населения региона. 6. Оценить сходства и различия избирательного накопления остатков жертв двух видов сов.
МАТЕРИАЛ И МЕТОДИКА
Районы исследования. Работы проведены на основе полевых сборов по питанию бородатой неясыти и филина в двух районах Среднего Урала, находящихся примерно на одной широте 57° с.ш., на расстоянии 200 км друг от друга.
Исследование питания бородатой неясыти проводилось с 2015 по 2018 гг. в Ирбитском р-не Свердловской обл. (восточный склон Урала) на базе научно-практического центра биоразнообразия “Скородум”. Здесь для этих птиц Г.Н. и В.Н. Бачуриными были установлены искусственные гнезда (Смирнов и др., 2015). Изучаемая территория находится на границе лесной и лесостепной зон и отличается разнообразием биотопических условий. Растительность представлена разновозрастными и разнопородными участками леса с вырубками и обширными заболоченными участками, перемежающимися с сельскохозяйственными угодьями (полями, лугами, пастбищами). Подробное описание территории и результаты сравнения состава погадок 1978 и 2015 гг. опубликованы ранее (Смирнов и др., 2015). Заселенные гнезда располагались на территориях с разным соотношением открытых и закрытых местообитаний. В соответствии с биотопическими характеристиками было выделено 4 участка (рис. 1). Каждый участок содержит индивидуальные территории вокруг гнезд диаметром 3 км. Такая площадь выбрана в соответствии с представлением о размере охотничьей территории бородатой неясыти в гнездовой период (Bull et al., 1988; Duncan, Hayward, 1994; van Riper, van Wagtendonk, 2006).
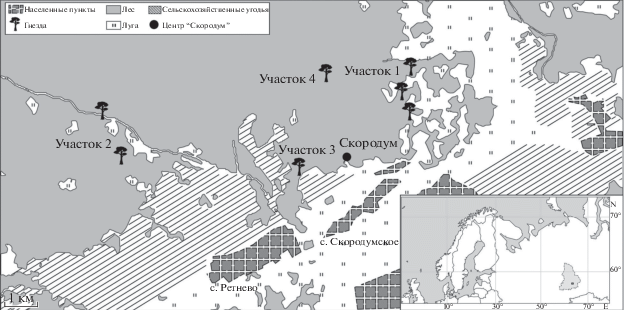
На участке 1 растительность представлена тремя категориями – лугами (47% площади участка) с сосново-березовыми колками (7%) и лесным массивом (46%). Здесь находится 3 гнезда. Далее наряду с номером этот участок будет называться “луговой”. Участок 2 представляет собой лесной массив (88%) с наличием луговых территорий (12%) (2 гнезда). Для него будет использоваться название “лесо-луговой”. Участок 3 – возделываемые и заброшенные поля (47%) и лесной массив (53%) (1 гнездо) – “полевой”. На участке 4 почти вся территория покрыта лесом (98%) с лесными полянами (2%), а ближайшие луга расположены на расстоянии 1.5 км от гнезда (1 гнездо) – “лесной”. Зимние погадки собраны под присадами сов на территории центра “Скородум” и в пределах первых километров вокруг центра (рис. 1).
Материал по питанию филинов собран в местах их многолетнего гнездования (несколько десятков лет) в карстовых навесах и гротах Западного склона Среднего Урала. Здесь проходит граница южной части тайги и островной Красноуфимской лесостепи. Исследованные гнезда и присады находятся на небольших площадках на земле и скальном ложе в верхних частях высоких береговых известняковых обрывов рек Серга (Филин, Старик, Бажуково) и Уфа (Сухореченский) (Смирнов и др., 1992; Смирнов, 1993; Садыкова, 2006; 2011). Все эти местонахождения по структуре окружающих ландшафтов однотипны. На противоположном от скал с гнездом берегу располагается полоса пойменных лугов, на вышележащих террасах смешанные леса разных пород с небольшими лугами. На том или ином удалении от долины реки (от сотен метров до первых километров) находились сельскохозяйственные поля. По противоположному берегу большие пространства (многие километры) заняты сплошными массивами смешанных и хвойных лесов, с небольшими участками открытых пространств в виде вырубок, гарей и верховых болот. Несмотря на то, что структура ландшафта одинакова для всех местонахождений, они отличаются важнейшим параметром – расстоянием до обширных обрабатываемых сельхозугодий. Поля расположены буквально в прямой видимости на расстоянии менее одного километра от гнезда Сухореченского. Гнездо Филин находится вблизи от обширных полей, примыкающих к д. Аракаево. Это расстояние по прямой составляет немного более 1 км. Открытые пространства около гнезд Старик и Бажуково представлены только пойменными естественными лугами, сенокосными полянами в лесу и относительно небольшими выпасами вокруг разъезда Бажуково и д. Половинка на расстоянии в несколько километров.
Анализ пищевого спектра бородатой неясыти. Свежие погадки собирали под присадами возле гнезд в период гнездования, пока птицы не покинут гнездовой участок. В 2016 и 2018 гг. совы покинули гнезда в конце мая, не завершив гнездования из-за депрессии численности грызунов. В 2015 и 2017 гг. гнездование было успешным. Всего данные были получены на основе анализа погадок, собранных в течение тринадцати гнездований и четырех зим. Годы заселения каждого гнезда приведены в табл. 1. Грызуны идентифицированы до вида по коренным зубам (Бородин, 2009), насекомоядные – до рода (Большаков и др., 2000). Число особей определено по максимальному количеству одноименных остатков (коренных зубов). Из 525 погадок определено 5080 остатков от 2055 особей животных. Виды жертв сгруппированы по биотопическому и систематическому принципам (Громов, Поляков, 1977; Башенина, 1981; Большаков и др., 2000; Маркова и др., 2017) с учетом данных по их роли в кормодобывающей деятельности сов. Таких групп образовано шесть: 1. Серые полевки, обитающие в открытых биотопах (луга) – обыкновенная (Microtus arvalis sensu lato) и узкочерепная полевки (M. gregalis); 2. Серые полевки, обитающие в относительно открытых лесных биотопах – полевка-экономка (M. oeconomus) и темная полевка (M. agrestis); 3. Лесные полевки – красная (Clethrionomys rutilus), рыжая (C. glareolus) и красно-серая (C. rufocanus) и 4. Малая лесная мышь (Sylvaemus uralensis). В две отдельные группы выделены землеройки (Soricidae) (5) и другие жертвы (6), остатки которых в погадках бородатой неясыти единичны или малочисленны: ондатра (Ondatra zibethicus), лесной лемминг (Myopus schisticolor), водяная полевка (Arvicola terrestris), обыкновенная белка (Sciurus vulgaris); большой суслик (Spermophilus major); лесная мышовка (Sicista betulina), ласка (Mustela nivalis), лягушки (Rana sp.).
Таблица 1.
Доли жертв в погадках бородатых неясытей, заселявших разные участки в гнездовой период (1–4) и в зимнем питании, %
| Группа жертв | Таксон/Гнездо, № | Луговой (1) | Лесо-луговой (2) | Полевой (3) | Лесной (4) | Зимнее питание | |||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |||
| 1 | Microtus arvalis s. l. | 26.3 | 17.5 | 20.4 | 65.4 | 48.1 | 0 | 5.2 | 14.1 |
| M. gregalis | 42.0 | 35.9 | 24.1 | 0.4 | 0 | 34.2 | 1.3 | 41.1 | |
| 2 | M. oeconomus | 21.7 | 17.9 | 33.6 | 11.9 | 22.2 | 9.8 | 26.1 | 12.4 |
| M. agrestis | 6.2 | 12.7 | 14.1 | 10.4 | 18.0 | 2.4 | 25.4 | 6.9 | |
| 3 | Clethrionomys rutilus | 0.4 | 3.3 | 2.6 | 1.7 | 0.8 | 4.9 | 0.4 | 4.9 |
| C. glareolus | 0.8 | 2.1 | 1.3 | 1.7 | 1.9 | 7.3 | 1.7 | 2.6 | |
| C. rufocanus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.9 | |
| 4 | Sylvaemus uralensis | 0.4 | 0.9 | 0.7 | 0 | 0 | 0 | 1.7 | 2.3 |
| 5 | Sorex sp. | 2.0 | 6.9 | 2.3 | 7.9 | 4.1 | 39.0 | 36.0 | 12.4 |
| 6 | Arvicola terrestris | 0 | 0.7 | 0.1 | 0 | 0 | 0 | 0.9 | 0 |
| Myopus schisticolor | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1.7 | |
| Sciurus vulgaris | 0 | 0 | 0.3 | 0 | 0 | 0 | 0 | 0.6 | |
| Spermophilus major | 0 | 0 | 0 | 0 | 0 | 2.4 | 0 | 0 | |
| Sicista betulina | 0.5 | 0.8 | 0.3 | 0.7 | 4.5 | 0 | 0.4 | 0 | |
| Ondatra zibethicus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.3 | |
| Mustela nivalis | 0.4 | 0.7 | 0.1 | 0 | 0.4 | 0 | 0 | 0 | |
| Rana sp. | 0 | 0.5 | 0.1 | 0 | 0 | 0 | 0.8 | 0 | |
| Годы сбора погадок | 2016, 2018 | 2016–2018 | 2015, 2017, 2018 | 2017, 2018 | 2017 | 2017 | 2016, 2017 | 2015–2018 | |
| Количество погадок | 61 | 61 | 165 | 43 | 63 | 10 | 60 | 62 | |
| Количество особей жертв | 250 | 243 | 534 | 194 | 266 | 41 | 179 | 348 | |
Деление жертв на основные, альтернативные и сопутствующие проводится на основе взаимодействия характеристик хищника и жертв. Основными жертвами являются устойчиво доминирующие размерно-биотопические группы добычи. Альтернативные жертвы при снижении численности основных жертв дополняют рацион (Korpimäki, Norrdahl 1991; Rutz, Bijlsma 2006; Zárybnická et al., 2009). Сопутствующие жертвы оказываются в добыче эпизодически и представлены в погадочном материале единичными экземплярами.
Оценка численности мелких млекопитающих на охотничьей территории бородатых неясытей. Синхронно со сбором погадок в 2016–2018 гг. проведены отловы мелких млекопитающих живоловушками с возвратом в среду обитания. Ловушки были расставлены на 2–4 суток в пределах 1 км2 вокруг заселенных гнезд. Отловами были охвачены сухие и заболоченные участки леса, опушки, заросли кустарников и луга. Ловушки были установлены в линии на расстоянии 10 м друг от друга. Их проверяли дважды в сутки – утром и вечером. Грызунов определяли до вида, насекомоядных – до рода (Большаков и др., 2000; Бородин, 2009). Подсчитано количество животных на 100 ловушко-суток (л/с), отдельно для лесных и луговых линий отловов. Всего отловлено и идентифицировано 186 особей, которые при описании материала объединены в те же группы, что и при анализе погадок.
Для сравнения рационов сов, а также сравнения питания сов и отловов использован метод анализа соответствий. В сопоставление включены группы видов, составляющие не менее 10% костных остатков и пойманных животных. Из сравнений питания неясытей были исключены мыши, т.к. их остатки были единичными и присутствовали не во всех выборках. В сравнительный анализ питания бородатой неясыти и филина была включена водяная полевка и хомяк, т.к. в питании филина эти виды составляли значительную долю, но исключены бурозубки, т.к. не во всех опубликованных работах по филину содержится информация о данной группе (Смирнов и др., 1992; Смирнов, 1993; Садыкова, 2006; Садыкова, 2011). Отношения между долями отдельных групп жертв в рационе и долей луговых территорий на участке обитания оценены с помощью коэффициента корреляции Спирмена. Данные обработаны в программном пакете Statistica 8.0 (StatSoft 2007).
РЕЗУЛЬТАТЫ
Общий таксономический состав добычи и соотношение долей разных видов в питании неясытей. На исследованной территории в целом за все годы сборов в погадках были найдены остатки 16 таксонов млекопитающих (табл. 1). Жертвы, принадлежащие к другим таксонам (птицы, амфибии, насекомые), были исключительно редки. Подавляющее большинство видов принадлежало полевкам. Единичными экземплярами были представлены мыши. Среди насекомоядных были обнаружены несколько видов. Как следует из перечня добычи, почти все виды следует отнести к мелким млекопитающим с размером тела не более 150 мм. Этот небольшой диапазон варьирования размера добычи и ее таксономического состава подтверждает общепризнанный вывод о принадлежности бородатой неясыти к категории специализированных хищников-миофагов.
По доле особей наиболее важной добычей бородатых неясытей являются серые полевки (78%), которых можно считать основными жертвами. Среди них доля видов, характерных для луговых биотопов, составляет 47% (M. arvalis 25% и M. gregalis 22%), для лесных местообитаний – 31% (M. oeconomus 19% и M. agrestis 12%). Далее следуют землеройки (14%), которые, очевидно, являются альтернативными жертвами. На сопутствующие жертвы (12 таксонов) приходится 8%. Лесные полевки составляют 5% (C. glareolus 2.5%, C. rutilus 2.5%, C. rufocanus < 1%), мыши – 1%, остальные виды – 2%. Ранее нами были описаны различия рациона по годам, периодам гнездования и при разной численности жертв в специальной работе и здесь подробно не рассматриваются (Кропачева и др., 2019).
Различия между отдельными гнездами в пределах участков с одинаковыми биотопическими характеристиками. Межгодовая изменчивость питания. На луговом участке (1) зафиксированы значимые межгодовые изменения рациона (χ2 = 54.22, df = 9, p < 0.001). В 2015, 2016 и 2018 гг. во всех выборках доминировали полевки, обитающие в луговых биотопах (M. arvalis и M. gregalis), в 2017 г. преобладали полевки, населяющие лесные местообитания (M. oeconomus и M. agrestis). Если сравнивать материал, накопленный за один и тот же временной интервал, различия между выборками оказываются значимыми только для 2018 г. (χ2 = 23.94, df = 6, p = 0.05). Если суммировать материал за три года для двух гнезд и за два года для одного гнезда, то различия между выборками оказываются значимыми (χ2 = 44.42, df = 6, p < < 0.001). В питании сов из двух гнезд доминировали полевки, обитающие в луговых биотопах (M. arvalis и M. gregalis), в питании сов из третьего гнезда преобладали серые полевки, населяющих лесные местообитания (M. oeconomus и M. agrestis) (табл. 1, рис. 2). На лесо-луговом участке (2) в питании сов доминировали серые полевки, обитающие в луговых биотопах (M. arvalis), далее следовали серые полевки, населяющие лесные местообитания (M. oeconomus и M. agrestis). Различия выборок из двух гнезд значимы (χ2 = 18.72, df = 3, p = 0.005), в основном они заключаются в различных долях этих биотопических групп серых полевок (рис. 2). Межгодовые различия рациона не значимы (χ2 = 7.81, df = 3, p = 0.09). На полевом участке (3) в питании сов примерно равные доли составляли серые полевки, обитающие в открытых биотопах (M. greralis) и бурозубки. На лесном участке (4) зафиксированы значимые межгодовые различия питания (χ2 = 47.35, df = 3, p < < 0.001). В 2016 г. в питании сов доминировали бурозубки, а в 2017 году – серые полевки, населяющие лесные местообитания (M. oeconomus и M. agrestis). В зимнем питании абсолютными доминантами были серые полевки, обитающие в открытых биотопах (M. arvalis и M. gregalis), далее следовали серые полевки, населяющие лесные местообитания (M. oeconomus и M. agrestis), в среднем по 10% приходится на лесных полевок и бурозубок (рис. 2).
Рис. 2.
Средние значения долей групп жертв в рационе бородатых неясытей в гнездовой и зимний периоды.
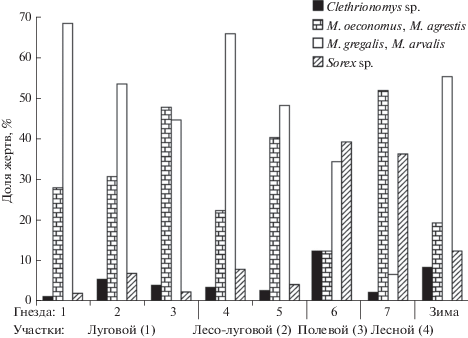
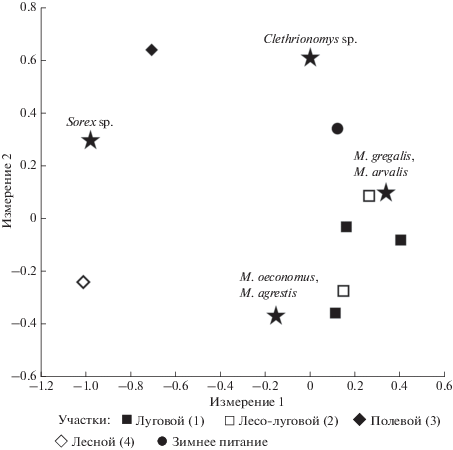
Различия рациона бородатых неясытей на разных участках в период гнездования и зимой значимы (χ2 = 428.29, df = 12, p < 0.001). Наибольшим своеобразием обладал рацион сов с лесного и полевого участков. Наиболее сходными оказались рационы сов, обитающих на луговом и лесо-луговом участках и зимнее питание (табл. 1; рис. 2, 3). Доля серых полевок открытых местообитаний в рационе не коррелирует с долей луговых местообитаний на участке (Spearman rank correlations, Sr = 0.14, P < 0.05).
Рис. 3.
Распределение долей групп видов жертв по гнездам на разных участках по результатам анализа соответствий.
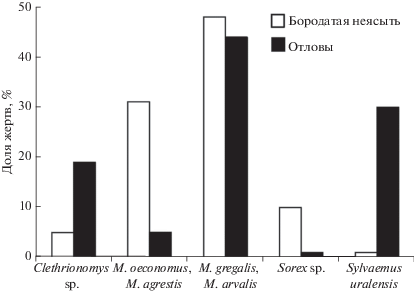
Оценки численности жертв бородатой неясыти путем отловов живоловками. В проведенных отловах, если оценивать их суммарно, преобладали серые полевки (49%), из которых большая доля приходилась на полевок, обитающих в луговых биотопах – 44% (M. arvalis 33% и M. gregalis 11%) и значительно меньшая – на полевок, населяющие лесные местообитания – 5% (M. oeconomus < 1% и M. agrestis 5%). Далее следовали мыши – 31% (Sylvaemus uralensis 31%, Apodemus agrarius < 1%), лесные полевки – 19% (C. rutilus 14%, C. glareolus 5%), и бурозубки – 1%. Доля групп добычи в отловах значимо отличалась от их пропорций в погадках (χ2 = 593.02, df = 4, p < 0.001) (рис. 4). В годы исследований наблюдалась разная численность мелких млекопитающих. В 2016 и 2018 гг. зафиксирована депрессия численности грызунов. В 2017 г. в луговых биотопах отмечена численность 5.0 особей/100л/с, в лесных биотопах – 6.0 особей/100л/с.
Рис. 4.
Средние значения долей групп видов в рационе бородатой неясыти и групп тех же видов в отловах.
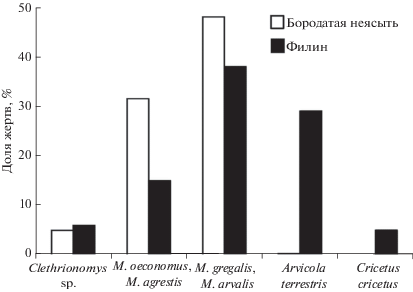
Соотношение долей видов жертв в питании филина. В питании филина отмечено 15 видов грызунов (табл. 2). Преобладают полевки, которые обитают в луговых биотопах изучаемого района – M. arvalis – 39%, далее следует A. terrestris – 29%. Эти виды следует считать основными жертвами, на них приходится 68% рациона. К альтернативным жертвам можно отнести серых полевок, населяющих лесные местообитания (в основном M. oeconomus) и обыкновенного хомяка (Cricetus cricetus), т.к. в разных гнездах их доля колеблется от единиц процентов до 25%. На сопутствующие виды (11 таксонов) приходится в сумме 16%. Рацион филина и бородатой неясыти значимо различались (χ2 = 585.66, df = 4, p < 0.001) (рис. 5).
Таблица 2.
Доля (%) жертв в питании филина
| Таксон | Гнездо | |||
|---|---|---|---|---|
| Бажуково | Филин | Старик | Сухореченский | |
| Microtus arvalis s. l. | 19.0 | 56.7 | 33.8 | 44.7 |
| M. oeconomus | 10.0 | 10.0 | 24.9 | 0.5 |
| M. agrestis | 6.0 | 2.4 | 4.6 | 2.8 |
| Clethrionomys rutilus | 0 | 0.9 | 2.1 | 0 |
| Cl. glareolus | 0 | 1.8 | 3.2 | 2.5 |
| Cl. rufocanus | 0 | 0 | 0.4 | 1.0 |
| Clethrionomys sp. | 7.0 | 4.3 | 0 | 0 |
| Apodemus sp. | 0.2 | 2.2 | 0.4 | 6.7 |
| Arvicola terrestris | 50.3 | 12.6 | 26.0 | 27.2 |
| Cricetus cricetus | 2.0 | 2.4 | 2.5 | 13.7 |
| Myopus schisticolor | 0.2 | 0.1 | 0 | 0 |
| Ondatra zibethicus | 0 | 1.9 | 0.4 | 0 |
| Sciurus vulgaris | 0.3 | 0.4 | 0.7 | 0.2 |
| Rattus norvegicus | 0 | 2.8 | 0 | + |
| Pteromys volans | 0.1 | 0.1 | 0 | 0.4 |
| Sicista betulina | 0.3 | 1.2 | 1.1 | 0 |
| Количество зубов | 23 031 | 672 | 1376 | 1704 |
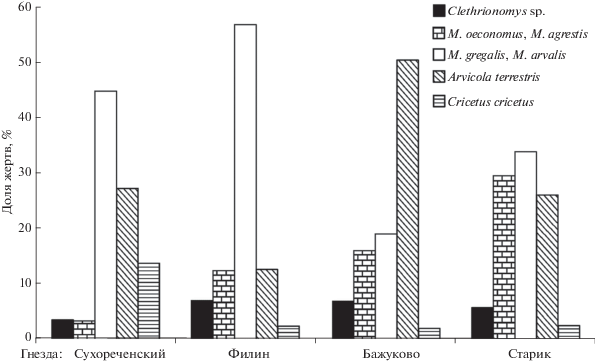
Состав и структура жертв филина из отдельных гнезд, расположенных в однотипном ландшафтном окружении, но со специфическим соотношением биотопов. Различия между выборками из местонахождений, сформированных погадками филина, значимы (χ2 = 88.19, df = 12, p < 0.001). В отложениях трех изученных местонахождений доминировала M. arvalis . В гнезде Филин данный вид был абсолютным доминантом, в местонахождении Сухореченский многочисленны остатки A. terrestris и C. cricetus, в гнезде Старик доминирование M. arvalis наименее выражено, сопоставимые доли принадлежали A. terrestris и M. oeconomus. В местонахождении Бажуково абсолютный доминант – A. terrestris, значительно меньшие доли принадлежали M. arvalis и M. oeconomus (рис. 6).
ОБСУЖДЕНИЕ
Специфика рациона сов определялась площадью охотничьей территории и тем, какие из предпочитаемых охотничьих биотопов и населяющих их жертв на ней представлены.
Различия между отдельными гнездами, но в пределах участков с одинаковыми биотопическими характеристиками. Межгодовая изменчивость питания. При суммировании материалов всех лет для каждого гнезда различия между ними, по-видимому, определялись комплексом факторов: локальными биотопическими условиями охотничьих участков, наличием межгодовой динамики численности мелких млекопитающих, а также чередованием в заселении птицами разных гнезд по годам. Межгодовая динамика, прослеженная на луговом участке (1), заключалась в колебании долей основных жертв при устойчиво малых долях альтернативной добычи. На лесном участке (4) в разные годы наблюдалось чередование доминирования основных и альтернативных жертв. Таким образом, на участках с наличием биотопов, населяемых двумя группами основных жертв, перераспределение в доминировании происходило между ними. При наличии на участке биотопов, населяемых только одной группой основных жертв, перераспределение доминантов происходило между основными и альтернативными жертвами.
Существует ряд факторов, которые не позволяют результаты анализа состава и структуры остатков мелких млекопитающих из многослойных отложений напрямую использовать для реконструкции межгодовой динамики их численности. Накопление погадочного материала происходит неравномерно в разные годы. Захоронение костей жертв и преобразование их в субфоссильные остатки сопровождается их частичной потерей. При формировании отложений может происходить перемешивание остатков, в результате чего правильная стратиграфическая последовательность нарушается. Эти процессы приводят к усреднению данных не только за отдельные годы, но за десятки и сотни лет.
Можно предположить, что при наличии на участке двух и, возможно, более предпочитаемых видов или биотопически сходных групп жертв их доли выравниваются в результате межгодовой динамики. В случае, если на участке обитания в годы с низкой численностью основных жертв происходит переключение на альтернативные жертвы, их доля в отложениях возрастает за счет накопления в течение ряда таких лет.
Различия между питанием сов на участках с различными биотопическими характеристиками. При наличии лугов на участке обитания (участки 1–3), вне зависимости от их площади и числа населяющих их видов, в рационе бородатой неясыти доминировали обитатели лугов. На луговом участке (1) доминировали два вида – M. arvalis и M. gregalis, на лесо-луговом участке (2) в добыче неясыти доминировал один вид – M. arvalis, на полевом участке (3) – M. gregalis. Бородатая неясыть предпочитала охотиться на полевок, населяющих открытые биотопы, так же и в зимний сезон, когда размеры охотничьей территории сов намного больше, чем в гнездовой период (Bull et al., 1988; van Riper, van Wagtendonk, 2006). Следующей по численности группой жертв на участках 1 и 2 были серые полевки, населяющие относительно открытые лесные местообитания (M. oeconomus, M. agrestis). На участке 3 в питании неясыти сопоставимую долю с полевками имели бурозубки. Мы имеем только одну небольшую выборку с данного участка, поэтому его своеобразие может быть связано как с объемом материала, так и с биотопическими характеристиками участка.
Лесной участок (4) отличался от других низкой долей луговых видов, несмотря на его незначительное удаление от лугов (1.5 км). Это характеризует охват территории, которая может отражаться в питании данного вида в гнездовой период. На данном участке, при меньшей доступности луговых видов, в питании доминировали серые полевки, населяющие относительно открытые лесные местообитания (M. oeconomus, M. agrestis). Второй по численности группой жертв на участке 4 являются альтернативные жертвы – бурозубки.
Избирательность питания бородатой неясыти. Специфика рациона в сравнении с другими частями ареала. В погадках неясыти зафиксировано 15 видов грызунов. Все они отмечены в литературе для данной территории (Большаков и др., 2000). Отсутствовали в добыче летяга, бурундук, мышь-малютка, обыкновенный хомяк – виды здесь и повсеместно редкие в Свердловской обл. (Большаков и др., 2000). Кроме того, размеры зверьков этих видов находятся за пределами диапазона размеров предпочитаемой добычи. Из особенно редких, но находящихся в пределах основной размерной группы жертв, в погадках отмечены лесной лемминг и красно-серая полевка. Кроме грызунов в погадках также содержатся остатки бурозубок, ласки и лягушек. Таким образом, фауна грызунов региона достаточно полно отражена в питании неясыти. Во многих работах показано, что с помощью анализа погадок сов можно эффективнее оценить фауну, чем с помощью отловов (Andrews, 1990; Terry, 2010).
В питании неясыти доминировали виды, местообитания которых птицы предпочитали для охоты (Bull et al., 1989; Duncan, Hayward, 1994; van Riper, van Wagtendonk, 2006): серые полевки (M. arvalis и M. gregalis) луговых местообитаний. В проведенных нами отловах также доминировала данная группа мелких млекопитающих. Второй по обилию группой жертв были серые полевки, населяющие относительно открытые лесные биотопы (M. oeconomus, M. agrestis). Доля этих видов в погадках значительно превосходит их долю в отловах. Экономка предпочитает селиться в заболоченных и прочих увлажненных биотопах. Темная полевка обитает на вырубках и гарях, на лесных полянах, болотах. На исследуемой территории много биотопов, подходящих для этих видов. Перечисленные виды являются основными жертвами бородатой неясыти и на других частях ее ареала. Так, в Уральском регионе в питании многочисленна M. arvalis sensu lato, населяющая преимущественно агроландшафты (Карякин, 1998; Шепель, 2011). Темная полевка и, в меньшей степени, полевка-экономка являются основными жертвами бородатой неясыти в Фенноскандии и Белоруссии (Mikkola, Sulkawa, 1970; Mikkola, 1981; Sulkava, Huhtala, 1997; Tishechkin, 1997). Полевка-экономка на Северо-Востоке Европы, так же как на Урале, населяет увлажненные лесные и пойменные местообитания, тогда как темная полевка в Восточной Фенноскандии занимает широкий спектр местообитаний, наиболее обычными из которых являются сельскохозяйственные угодья и окраины населенных пунктов (Ивантер и др., 2013). Таким образом, темная полевка населяет ряд биотопов, характерных на Урале для обыкновенной полевки.
Доля бурозубок в рационе колебалась от нуля до доминирования, а в наших отловах была стабильно низкой. На всем ареале бородатой неясыти бурозубки являются постоянным элементом добычи (Mikkola, Sulkawa, 1970; Mikkola, 1981; Duncan, Hayward, 1994; Sulkava, Huhtala, 1997; Tishechkin, 1997; Карякин, 1998; Пукинский, 2005).
Лесные полевки были редкими жертвами неясытей в период гнездования, их доля повышалась зимой. В проведенных нами отловах лесные полевки были второй по обилию группой. Среди них преобладали Cl. rutilus. Литературные данные свидетельствуют, что в других частях ареала лесные полевки достигали существенной доли в питании бородатой неясыти, однако данная группа была представлена Cl. glareolus (Пермский Край, Башкирия (Карякин, 1998), ряд локалитетов Фенноскандии (Mikkola, 1981)) и Cl. rufocanus (Мурманская область (Mikkola, 1981)). Cl. rutilus в сравнении с Cl. glareolus и Cl. rufocanus занимает более закрытые местообитания (Башенина, 1981). Вероятно, низкая доля лесных полевок в питании бородатой неясыти в исследуемом районе связана с невысокой численностью Cl. glareolus и, в особенности, Cl. rufocanus, а также с меньшей доступностью Cl. rutilus, по сравнению с этими видами. В пользу этого предположения свидетельствует более высокая доля лесных полевок в зимнем питании, когда, из-за отсутствия листвы, они становятся более доступной жертвой, чем в весенне-летний период.
Мыши были единичны в погадках на всех участках, несмотря на их наличие в природе. Они были доминирующей по численности группой в наших отловах. Доля мышей в рационе бородатой неясыти никогда не бывает высокой и в других частях ареала (Mikkola, 1981; Korpimäki, 1986; Tishechkin, 1997). Вероятно, эта группа жертв недоступна для бородатой неясыти, т.к. предпочитает закрытые местообитания и характеризуется высокой подвижностью.
Таким образом, в питании неясыти в сравнении с данными проведенных отловов мелких млекопитающих в изучаемом районе наблюдалась сопоставимая доля серых полевок открытых местообитаний, большая доля серых полевок относительно открытых лесных местообитаний и бурозубок, меньшая доля лесных полевок и мышей.
Состав и структура жертв филина из отдельных гнезд, расположенных в однотипном ландшафтном окружении, но на территориях с разным соотношением биотопов. Общий состав добычи каждого местонахождения полностью отражает состав региональной фауны. Доля остатков трех видов полевок рода Clethrionomys, по которым можно судить о присутствии в жертв питании, добытых филинами в лесных закрытых биотопах, не достигает 10%. Другие виды (обитатели лесных биотопов – белка, летяга, лесные мыши, лесной лемминг) – существенно не меняют представления о резком преобладании в добыче видов открытых биотопов. Различия рациона филинов в четырех местонахождениях, использованных в данной работе, определяются главным образом составом видов-доминантов. В двух гнездах – Сухореченский и Филин – резко доминируют обитатели полей и лугов – обыкновенные полевки, в Бажуково более 50% остатков принадлежит водяной полевке, тогда как в гнезде Старик явного доминанта нет, а примерно по 30% приходится на остатки водяной и обыкновенной полевок и группе, в которую объединены темная полевка и экономка. Такая специфика легко объясняется различиями биотопических характеристик участков, окружающих гнезда. Главное различие, как было указано выше при описании района исследований, состоит в степени удаленности от сельскохозяйственных угодий. Наиболее показателен в этом грот Сухореченский. Он находится на берегу р. Уфа, правый берег который занят таежными лесами Уфимского плато, а левый – Красноуфимской лесостепью, практически полностью освоенной сельскохозяйственными угодьями и населенными пунктами. Судя по основной добыче филина в этом гнезде его основные жертвы были добыты на открытых пространствах лесостепи, а виды лесных местообитаний составляли менее 7%. Размерный диапазон добычи филинов во всех гнездах был одинаково широк (от 50 до 350 мм), но размеры основных жертв существенно различались в соответствии с тем, кто из доминантов – водяная или обыкновенная полевка – играли в добыче основную роль. По доле добываемых особей размерная группа, в которую входят относительно крупные грызуны – ондатра, обыкновенный хомяк, водяная полевка, серая крыса, белка, летяга составляют в гнезде Бажуково 52.4%, в гнезде Старик 29.6, в гнезде Филин, в Сухореченском 43.5. Массу добычи этой группы видов точно оценить трудно, но ясно, что в процентном отношении этот показатель в несколько раз превышает долю мелких жертв.
Избирательность питания филина в изучаемом районе в сравнении с литературными данными по составу фауны и структуре населения региона. Корректная количественная оценка избирательности добычи хищника в сравнении с фауной и структурой населения жертв возможна при строгом соотнесении масштабов исследования территории (локальный, региональный, зональный) и площади охотничьего участка. К сожалению, мы не располагаем такими сведениями, поэтому вынуждены применить только качественный подход и ограничиться предварительными результатами.
Таксономический состав мелких млекопитающих из добычи филинов почти полностью соответствует фауне региона, что исключает этот аспект из обсуждения избирательности. Такое положение заставляет считать филина данного региона универсальным хищником. Эту характеристику усиливает присутствие остатков птиц в изученных нами отложениях (Смирнов и др., 1992; Смирнов, 1993; Садыкова, 2006; 2011).
Размерный диапазон добычи весьма широк – от зайца до мелких землероек и мышовок. Доля добываемых особей из группы мелких жертв составляет от половины до двух третей. Такие оценки, с одной стороны, еще раз характеризуют филина как универсального, а не специализированного хищника, но, с другой стороны, требуют характеристики избирательности в выборе добычи по ее размерам. Представляется оправданным рассматривать во взаимосвязи состав размерных и биотопических групп жертв филина. Два вида составляют основу питания – водяная и обыкновенная полевки. Первый вид составляет основу населения лугов в поймах рек и в то же время он самый массовый вид в размерной группе крупных грызунов. Обыкновенная полевка – массовый вид биотопа сельскохозяйственных лугов и полей, и он же фактически определяет численность группы жертв мелкой размерной группы. В литературе существует представление о том, что хищные птицы промышляют любой доступный по размеру объект (из соответствующего спектра среды обитания), оказавшийся в пределах досягаемости (Pyke, 1984). Этот взгляд определяет возможность выбора хищником только времени и места (биотоп) для охоты в соответствии с его физическими возможностями и поведенческими навыками. Применяя такой подход, следует признать, что избирательность добычи основных жертв филинов в рассматриваемом районе заключается в выборе между охотой в поймах рек на водяных полевок или охотой на открытых пространствах сельскохозяйственных угодий на обыкновенных полевок.
Сходства и отличия избирательного накопления остатков жертв двух видов сов. Филина и бородатую неясыть объединяет ряд общих или близких черт, но другие признаки являются специфическими. Оба вида имеют довольно крупные размеры, хотя бородатая неясыть уступает филину и по длине тела, и по массе. Их объединяет то, что при выборе охотничьих участков они тяготеют к открытым биотопам в виде лугов, полей, пустошей. Если это лесные участки, то относительно открытые, такие как болота, гари, вырубки (Mikkola, 1981; Bull et al., 1989; Duncan, Hayward, 1994; Пукинский, 1993, 2005; Van Riper, van Wagtendonk, 2006).
Рацион филина полнее рациона неясыти отражает фауну мелких млекопитающих за счет добывания крупных жертв – обыкновенного хомяка, белки-летяги, серой крысы. Наиболее частые жертвы у обоих видов – серые полевки, обитающие на открытых пространствах. В рационе бородатой неясыти второй по обилию в рационе группой являются серые полевки относительно открытых лесных биотопов. В питании филина многочисленной жертвой является водяная полевка. В питании бородатой неясыти изучаемого района в 1978 г. отмечалась существенная доля полевок этого вида (Смирнов и др., 2015), следовательно, они являются доступными жертвами и не добываются в настоящее время, вероятно, из-за сократившейся численности. В питании филина заметную долю составляет обыкновенный хомяк, в рационе бородатой неясыти он отсутствует. В исследуемом районе данный вид обитает в настоящее время, но численность его низка.
Анализ состава сопутствующих жертв показывает, что большая их часть представлена обитателями лесных биотопов. Это существенным образом дополняет характеристику территории, на которой расположены изучаемые гнезда.
Зависимость результатов оценки добычи от масштаба рассматриваемой территории. Территория, население мелких млекопитающих которой отражалось в питании неясыти, охватывала по нашим наблюдениям небольшую площадь – 7 км2. Луга как основные охотничьи биотопы определяли населяющие их группы основных жертв. Если эти луга находились в относительной близости от гнезда, то связи между их площадью и долей видов открытых местообитаний не наблюдалось. Нельзя отличить сообщество мелких млекопитающих лесного участка с наличием лугов, составляющих его незначительную часть, от сообщества участка, где луга составляют почти половину площади. В том и в другом случае основу добычи составили виды открытых биотопов. В случае, когда луга располагались на удалении 1.5 км от гнезда, о их наличии свидетельствовали лишь единичные остатки видов луговых местообитаний.
Площадь охотничьей территории филина в гнездовой период значительно больше, чем у бородатой неясыти (Mikkola, 1981, 1983). Кроме того, есть местонахождения, сформированные не только на местах гнезд, но и на местах присад. В таких местонахождениях накопление погадок происходит в течение всех сезонов. Вне гнездового периода охотничья территория существенно расширяется, следовательно, в таких локалитетах территория, отраженная в питании, значительно больше.
Основные жертвы служат индикаторами предпочитаемых охотничьих биотопов, тогда как сопутствующие виды добываются хищниками не только в этих биотопах, но и при транзитном пересечении птицами разных неудобных для охоты местообитаний. С точки зрения получения данных о питании хищников, сопутствующие виды не играют большой роли, тогда как для палеореконструкций они имеют важное индикаторное значение. Выявление данной группы жертв зависит как от пространственного, так и временного масштаба рассмотрения.
ЗАКЛЮЧЕНИЕ
Рассмотрение добычи сов-миофагов как источника накопления палеотериологических материалов приводит к выводу о том, что важнейшим аспектом избирательности служит выбор охотничьих биотопов. Именно в этих биотопах птицы добывают основных и альтернативных жертв. Эта добыча доминирует по числу особей, но принадлежит, как правило, к ограниченному количеству видов. Большая часть из списка добычи принадлежит к сопутствующим жертвам, которых хищники добывают не только на охотничьих, но и на транзитных территориях. Именно эти виды выступают в качестве индикаторов присутствия в окружающем ландшафте таких местообитаний, которые важны для палеореконструкций, но являются второстепенными для кормодобывающей деятельности хищников. Оценка надежности использования сопутствующих жертв как индикаторов тех или иных свойств реконструируемых сообществ с методологических позиций представляется вполне приемлемой. Залогом успешности применения разрабатываемого подхода служит то обстоятельство, что в основе разделения видов жертв на основные, альтернативные и сопутствующие лежит объективная характеристика – доля в питании хищника. Этот динамичный во времени и в пространстве признак относится к той категории, которая не вступает в противоречие с принципом дисконформизма в отличие от целой серии признаков – индикаторов средовых характеристик, основанной на экологических свойствах мелких млекопитающих (Маркова и др., 2017).
Различия в составе добычи бородатых неясытей, занимавших гнезда в пределах участков с наличием двух предпочитаемых для охоты типов биотопов, были незначительными и проявлялись в колебаниях долей основных жертв. Те же проявления имела межгодовая изменчивость питания. При наличии на участке биотопов, населяемых только одной группой основных жертв, перераспределение долей происходило между основными и альтернативными жертвами. Питание сов на разных участках определялось наличием открытых местообитаний. Доля серых полевок этих местообитаний в рационе не находится в прямой зависимости от доли луговых биотопов на участке.
В питании неясытей в сравнении с проведенными отловами наблюдалась сопоставимая доля серых полевок открытых местообитаний, большая доля серых полевок относительно открытых лесных местообитаний и бурозубок, меньшая доля лесных полевок и мышей. Состав и структура жертв филина достаточно полно отражала фауну региона. Питание птиц из отдельных гнезд по составу основных жертв отражало население грызунов, обитающих на предпочитаемых охотничьих биотопах. Избирательность питания филина заключалась в выборе между охотой в пойменных лугах на водяную полевку и экономку и охотой в сельскохозяйственных угодьях на обыкновенную полевку. Сходство в избирательности добычи жертв у двух видов сов заключалось в том, что основу рациона составляло небольшое количестве видов основных жертв, населяющих преимущественно открытые местообитания. У обоих видов сопутствующие жертвы – виды закрытых лесных (транзитных) биотопов. Различия заключаются в большем размерном диапазоне жертв у филина.
Список литературы
Большаков В.Н., Бердюгин К.И., Васильева И.А., Кузнецова И.А., 2000. Млекопитающие Свердловской области: Справочник-определитель. Екатеринбург: изд-во Екатеринбург. 240 с.
Бородин А.В., 2009. Определитель зубов полевок Урала и Западной Сибири (поздний плейстоцен–современность). Екатеринбург: Изд-во УрО РАН. 100 с.
Громов И.М., 1957. Некоторые итоги и перспективы изучения ископаемых четвертичных грызунов фауны СССР // Труды Зоологического Института АН СССР. Т. 22. С. 90–99.
Громов И.М., Поляков И.Я., 1977. Млекопитающие. Фауна СССР; Т. 3, Вып. 8. Л.: Наука. 502 с.
Башенина Н.В. (ред.), 1981. Европейская рыжая полевка. М.: Наука. 351 с.
Ивантер Э.В., Курхинен Ю.П., Соколов А.В., 2013. Экология темной полевки (Microtus agrestis L.) в коренных и антропогенных ландшафтах Восточной Фенноскандии // Экология. № 3. С. 189–189.
Карякин И.В., 1998. Пернатые хищники Уральского региона: Соколообразные (Falconiformes) и Совообразные (Strigiformes). Пермь: Центр полевых исследований Союза охраны животных Урала. С. 385–395.
Кропачева Ю.Э., Смирнов Н.Г., Зыков С.В., Чепраков М.И., Садыкова Н.О., Бачурин Г.Н., 2019. Питание бородатой неясыти Strix nebulosa при разной численности жертв в период гнездования // Экология. № 1. С. 40–46.
Маркова Е.А., Струкова Т.В., Бородин А.В., 2017. Полевки (Arvicolinae, Rodentia) как объект палеоэкологических исследований: классификация видов центральной части Северной Евразии по экологическим предпочтениям современных форм // Зоологический журнал. Т. 96. № 10. С. 1254–1266.
Пукинский Ю.Б., 1993. Филин // Птицы России и сопредельных регионов: Совообразные, Козодоеобразные, Стрижеобразные, Ракшеобразные, Удодообразные, Дятлообразные. М.: Наука. С. 270–289.
Пукинский Ю.Б., 2005. Бородатая неясыть // Птицы России и сопредельных регионов: Совообразные, Козодоеобразные, Стрижеобразные, Ракшеобразные, Удодообразные, Дятлообразные. М.: Товарищество научных изданий КМК. С. 86–98.
Садыкова Н.О., 2006. Неонтологические подходы к изучению механизмов формирования ископаемых локальных фаун грызунов // Современная палеонтология: классические и новейшие методы. М.: ПИН РАН. С. 109–120.
Садыкова Н.О., 2011. Изучение динамики сообществ грызунов на основе субфоссильного материала (на примере серии зоогенных скоплений в таежных районах Северного и Среднего Урала). Автореф. дис. …канд. биол. наук. Екатеринбург. 20 с.
Смирнов Н.Г., 1993. Мелкие млекопитающие Среднего Урала в позднем плейстоцене и голоцене. Екатеринбург: Наука, Уральское Отделение. 62 с.
Смирнов Н.Г., Ерохин Н.Г., Быкова Г.В., Лобанова А.В., Корона О.М., Широков В.Н., Некрасов А.Е., 1992. Грот Сухореченский – памятник истории природы и культуры в Красноуфимской лесостепи // История современной фауны Южного Урала. Свердловск: ИЭРиЖ УРО РАН. С. 20–43
Смирнов Н.Г., Кропачева Ю.Э., Бачурин Г.Н., 2015. Динамика современной фауны грызунов предлесостепных лесов Зауралья // Фауна Урала и Сибири. № 1. С. 167–175.
Смирнов Н.Г., Садыкова Н.О., 2003 Источники погрешностей при фаунистической реконструкции в четвертичной палеозоологии // Четвертичная палеозоология на Урале. Сборник науч. трудов. Екатеринбург: Издательство Уральского университета. С. 98–115.
Шепель А.И., 1992. Хищные птицы и совы Пермского Прикамья. Иркутск: Издательство Иркутского университета. 296 с.
Шепель А.И., 2011. Бородатая неясыть Strix nebulosa Forster, 1772 в Волжско-Камском крае // Вестник Удмуртского университета. Серия Биология. Науки о Земле. Т. 4. С. 85–89.
Шер А.А., 1999. Актуализм и дисконформизм в изучении экологии плейстоценовых млекопитающих // Журнал общей биологии. Т. 51. № 2. С. 163–177.
Andrews P., 1990. Owls, caves and fossils: predation, preservation and accumulation of small mammal bones in caves, with an analysis of the Pleistocene cave faunas from Westbury-sub-Mendip, Somerset, UK. Chicago: University of Chicago Press. 231 p.
Andrews P., Fernández-Jalvo Y., 2018. Seasonal variation in prey composition and digestion in small mammal predator assemblages // International Journal of Osteoarchaeology. V. 28. № 3. P. 318-331.
Bull E.L, Henjum M.G, Rohweder R.S., 1988. Home range and dispersal of great gray owls in northeastern Oregon // Journal of Raptor Research. V. 22. P. 101–106.
Bull E.L., Henjum, M.G., Rohweder, R.S., 1989. Diet and optimal foraging of great gray owls // The Journal of Wildlife Management. V. 53. P. 47–50.
Comay O., Dayan T., 2018. From micromammals to paleoenvironments // Archaeological and Anthropological Sciences. V. 10. № 8. P. 2159–2171.
Duncan J., Hayward P.H., 1994. Review of technical knowledge: great gray owls // Flammulated, Boreal, and Great Gray Owls in the United States: a technical conservation assessment. V. 253. P. 1–159.
Korpimäki E., 1986. Niche relationships and life-history tactics of three sympatric Strix owl species in Finland // Ornis Scandinavica. V. 17. P. 126–132.
Korpimäki E., Marti C.D., 1995. Geographical trends in trophic characteristics of mammal-eating and bird-eating raptors in Europe and North America // The Auk. V. 112. № 4. P. 1004–1023.
Korpimäki E., Norrdahl K., 1991. Numerical and functional responses of kestrels, short-eared owls and long-eared owls to vole densities // Ecology. V. 72. № 3. P. 814–826.
Mikkola H., 1981. Der Bartkauz Strix nebulosa. Wittenberg-Lutherstadt: Ziemsen. 124 p.
Mikkola H., 1983. Owls of Europe. Berkhamsted: T. & AD Poyser. 397 p.
Mikkola H., Sulkava S., 1970. Food of great grey owls in Fenno-Scandia // British Birds. V. 62. P. 23–27.
Pyke G. H., 1984. Optimal foraging theory: a critical review // Annual review of ecology and systematics. V. 15. № 1. P. 523–575.
Rutz C., Bijlsma, R. G., 2006. Food limitation in a generalist predator // Proceedings of the Royal Society of London. Series B: Biological Sciences. V. 273. P. 2069–2076.
Sulkava S., Huhtala K., 1997. The Great Grey Owl (Strix nebulosa) in the changing forest environment of northern Europe // Journal of Raptor Research. V. 31. P. 151–159.
StatSoft Inc, 2007. STATISTICA for Windows (software system for data analysis), version 8.0, http://www.statsoft.pl
Terry R.C., 2010. The dead do not lie: using skeletal remains for rapid assessment of historical small-mammal community baselines // Proceedings of the Royal Society. Series B: Biological Sciences. V. 277. № 1685. P. 1193–1201.
Tishechkin, A.K., 1997. Comparative food niche analysis of Strix owls in Belarus // General Techical Reports, North Central Forest Experiment Station. V. 190. P. 456–460.
Tobajas J., Fernandez-de-Simon J., Díaz-Ruiz F., Villafuerte R., Ferreras P., 2016. Functional responses to changes in rabbit abundance: is the eagle owl a generalist or a specialist predator? // European Journal of Wildlife Research. V. 62. № 1. P. 85–92.
Van Riper C., van Wagtendonk J., 2006. Home range characteristics of Great Gray Owls in Yosemite National Park, California // Journal of Raptor Research. V. 40. № 2. P. 130–141.
Zárybnická M., Sedláček O., Korpimäki E., 2009. Do Tengmalm’s Owls alter parental feeding effort under varying conditions of main prey availability? // Journal of Ornithology. V. 150. № 1. P. 231–237.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал