

Зоологический журнал, 2019, T. 98, № 12, стр. 1373-1391
Понятие “вид” у птиц: теория и практика
Е. А. Коблик a, *, Я. А. Редькин a, С. В. Волков a, b, А. А. Мосалов c
a Научно-исследовательский зоологический музей Московского государственного университета
125009 Москва, Россия
b Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Россия
c Институт биологии и химии, Московский государственный педагогический университет
129164 Москва, Россия
* E-mail: koblik@zmmu.msu.ru
Поступила в редакцию 07.02.2019
После доработки 12.02.2019
Принята к публикации 15.02.2019
Аннотация
В настоящее время стало очевидным, что репродуктивная изоляция не может оставаться ведущим критерием для понимания границ видов, хотя этот критерий по-прежнему применим для оценки статуса большинства форм. Наличие/отсутствие гибридизации между особями разных форм не связано напрямую со степенью эволюционной обособленности последних. Гибридизация способна приводить к возникновению новых таксонов, в том числе таксонов, имеющих видовой статус. При реконструкции родственных связей в сложных группах, включающих формы разного эволюционного уровня (от географических рас до “хороших” видов), взаимоотношения между которыми осложнены наличием гибридогенного полиморфизма и/или гибридогенных популяций, оправдано применение филогенетического метода. С учетом новых данных и концепций мы предлагаем по отношению к птицам использовать оригинальные взаимосвязанные определения понятий вид, подвид и полувид. Определения основаны на двух основных критериях: биологическом, состоящeм в оценке репродуктивных взаимоотношений конкретных форм друг с другом, и филогенетическом, состоящeм в оценке их эволюционного возраста и степени родства. Основным признаком вида (species) как эволюционной общности следует считать его стабильность во времени и пространстве, даже в случаях периодических нарушений репродуктивной изоляции. Географические внутривидовые расы имеют устойчивые различия разной степени, но не обладают репродуктивной изоляцией и при контактах скрещиваются, образуя зоны интерградации. Таксономически их обозначают как подвиды (subspecies). Описание новых подвидов продуктивно до такой степени дробности, которая наиболее полно отражает географическую структуру вида. Для молодых форм, приблизившихся в процессе эволюции к уровню видовой обособленности, целесообразно возродить категорию “полувид” (semispecies). Полувиды, как правило, обладают значительными морфологическими различиями и выраженной экологической спецификой, но связаны с близкородственными формами потоками генов в зонах контакта. Выделение этой категории не регламентировано “Кодексом зоологической номенклатуры”. При превалировании “дробительских” тенденций в современной систематике полувиды чаще приравниваются к видам, т.е. фигурируют под биноминальными названиями. Мы предлагаем дополнительно указывать принадлежность полувида к конкретной видовой группе (надвиду), имя которой приводить в круглых скобках между родовым и видовым названиями. Это позволит очертить естественные комплексы, избегая необоснованного повышения таксономического статуса форм, находящихся в процессе становления. В качестве полувидов не должны рассматриваться представители удаленных филогенетических линий (исторически давно обособившихся форм), даже в случаях нарушения их репродуктивной изоляции и наличия устойчивой гибридизации между ними.
Понятие вида – одно из ключевых понятий в биологии и основа биологической систематики живых организмов. Число признаваемых видов птиц в мировых сводках периодически изменяется. После принятия концепции политипического вида это число сократилось с 18 939 видов в 1890–1909 гг. до 8616 видов в 1946 г. (Haffer, 1992, 1997), но с тех пор растет вновь, достигая сейчас почти 11 000 видов (Dickinson, Remsen, 2013; Dickinson, Christidis, 2014; del Hoyo, Collar, 2014, 2016). В посвященном отряду Passeriformes томе Illustrated Checklist of the Birds of the World (del Hoyo, Collar, 2016) на 628 случаев дробления видов приходится всего 41 случай их объединения. Сходные “дробительские” тенденции (касающиеся и таксонов высокого ранга), сопровождающиеся усложнением структуры ранжирования, превалируют в систематике практически всех царств живой природы (Zhang, 2013; Аверьянов, Лопатин, 2014). Действительно, в ходе революции 1930-х годов в систематике и укрупнения “чрезмерно узко понимаемых” видов многие таксоны потеряли видовой ранг незаслуженно. С накоплением и осмыслением данных о специфике ряда форм такие ошибки были постепенно исправлены (см. обзор Степанян, 1983). Вместе с тем, видовой статус все чаще стали получать внешне слаборазличимые, обычно аллопатричные формы, ранее считавшиеся конспецифичными. В качестве критериев их выделения приводят генетические дистанции, факты отсутствия обмена генами, различия в вокализации, экологии, деталях окраски и т.п. В каком-то смысле можно говорить о “таксономической инфляции” и “девальвации понятия вид” (del Hoyo, Collar, 2014).
Недавно развернувшаяся полемика о “таксономической дисциплине” и “таксономической анархии” (Garnett, Christidis, 2017; Thompson et al., 2018) в очередной раз обозначила и проблему прикладного характера: в зависимости от того, какая концепция вида используется систематиками, представители одного класса животных могут оказаться в более угрожаемом положении, чем представители другого класса, поскольку получают меньшее финансирование на сохранение. Например, в 2012 г., согласно Закону США об исчезающих видах, примерно одинаковые суммы средств были выделены на защиту генетического разнообразия птиц и более дробно разделенных млекопитающих. Американские орнитосистематики придерживаются биологической концепции вида (далее БКВ), которую большинство таксономистов-териологов считают “консервативной”. С принятием филогенетической концепции вида (далее ФКВ) число видов птиц увеличилось бы в два раза, соответственно увеличилось бы и финансирование на их охрану (Garnett, Christidis, 2017). Существует и противоположное мнение, что увеличение числа признаваемых видов не зависит от перехода на ФКВ (Sangster, 2009).
В русскоязычной орнитологической литературе увеличение числа видов последовательно критикуют, выступая, либо с позиции очень узко понимаемой БКВ (Пфандер, 2018), либо с позиций эпигенетической теории эволюции и неприятия всей синтетической теории эволюции (Михайлов, 2003, 2015, 2017, 2018). Несмотря на разные мировоззренческие позиции, оба упомянутых автора выступают за возвращение к широкой трактовке видов, в целом соответствующей понятию “зоогеографического вида” (Haffer, 1997).
Таким образом, птицы, послужив главной моделью в процессе становления БКВ благодаря своей специфике (в первую очередь сложному поведению), становятся и ее “последним оплотом”. Вместе с тем в последние годы было проведено множество исследований, позволяющих взглянуть иначе на некоторые из критериев, традиционно применяемых к понятию “вид” для представителей класса Aves. Не претендуя на философские обобщения, мы сочли полезным суммировать новые данные и в операционном ключе переформулировать некоторые понятия.
Критерии, связанные с гибридизацией
Неопределенность критерия репродуктивной изоляции продолжает оставаться слабым местом “классической” БКВ (Панов, 1989). За это БКВ подвергается справедливой критике со стороны сторонников ФКВ. Очевидно, осмысление категории вида не сводится к его репродуктивной замкнутости (Панов, 1993). Многочисленные современные данные указывают на широкое распространение регулярной межвидовой гибридизации у птиц в природе (в том числе без каких-либо селективных последствий) – она отмечена между более чем 850 видами, т.е. двенадцатой частью состава мировой авифауны (Панов, 1989, 2001; McCarthy, 2006; Brelsford, 2011). В районах симпатрии родственных видов, например восточного (Luscinia luscinia) и южного (L. megarhynchos) соловьев, до 10% пар оказываются смешанными (Becker, 2007), а доля морфологически различимых гибридов – и более высокой (Kováts et al., 2013). Достаточно распространенной оказывается и гибридизация между представителями разных родов и даже семейств (McCarthy, 2006).
В ряде случаев формы видового ранга вступают в гибридизацию, масштабы которой оказываются столь значительными, что приводят к возникновению устойчивого гибридогенного полиморфизма и/или фактическому поглощению части популяций одного вида другим. Примеры таких ситуаций – взаимоотношения представителей групп “больших белоголовых чаек”, “желтых трясогузок” и “серых сорокопутов” в относительно недавнее время (Панов, 1989, 1993; Liebers et al., 2004; Тайкова, Редькин, 2014; Редькин и др., 2015; Red’kin et al., 2016).
Обзор Лавренченко (2013) показывает, что среди млекопитающих только 15 видов имеет предположительно гибридогенное происхождение, с учетом двух полиплоидов и одного межродового гибрида. Среди птиц известно еще меньше случаев, которые можно было бы уверенно трактовать как обособление гибридов в самостоятельный вид с возникновением механизмов, изолирующих от обеих родительских форм. Их трудно выявить с применением одних лишь молекулярно-генетических или морфологических методов, поэтому для обоснования требуется комплексный подход, включающий оценку морфологической (как минимум) специфики и характер репродуктивных взаимоотношений. Гибридогенным видом стали признавать итальянского воробья (Passer italiae), по формальным признакам таковыми могут считаться желтошапочный манакин (Lepidothrix vilasboasi), ареал которого отграничен от соседних форм крупными реками (Barrera-Guzman et al., 2017), и сформировавшийся (с достижением репродуктивной изоляции) буквально в течение нескольких поколений галапагосский вьюрок “Geospiza sp. nova” один из родителей которого – аборигенный вид G. conirostris, а другой – недавний вселенец G. fortis (Lamichhaney et al., 2017).
Несовпадение принадлежности к группировкам по признакам морфологии, вокализации, экологии с результатами филогенетических построений, основанных на молекулярных данных, дает основания предполагать гибридогенное происхождение некоторых видов с неравновесным заимствованием частей генома и столь же неравномерным проявлением фенотипа. В частности, таковой представляется ситуация с кластеризацией ряда видов пеночек (Phylloscopidae), относящихся “не к своим” подродам (Коблик и др., 2017).
В фауне Северной Америки изучен случай возникновения древесницы Одюбона (Dendroica auduboni sensu stricto), которая, как оказалось, представляет собой не просто южную группу подвидов миртовой древесницы (D. сoronata), а продукт ее скрещивания с предположительно другим видом (Brelsford, 2011; Brelsford et al., 2011; Milá, Irwin, 2011). Широко распространенная форма auduboni, действительно имеет черты сходства с миртовой древесницей – северным членом родительской пары. Тем не менее, характер ее отношений с D. сoronata sensu stricto (парапатрия с узкими стабильными зонами гибридизации) не соответствует зонам интерградации, характерной для подвидов (Hubbard, 1969), и это не позволяет относить форму auduboni sensu stricto к тому же виду. По окраске она тяготеет к изолированным узкоареальным расам nigrifrons и goldmani, возможно, представляющим собой “южного члена пары”. В последних сводках (del Hoyo, Collar, 2016) формы auduboni, nigrifrons и goldmani объединяют под старшим названием D. auduboni в таксон видового ранга (и частично гибридогенного происхождения). Некоторые же исследователи (Milá, Irwin, 2011), помимо признания гибридогенного D. auduboni, считают южные формы самостоятельным видом D. nigrifrons или придают видовой ранг каждой из них.
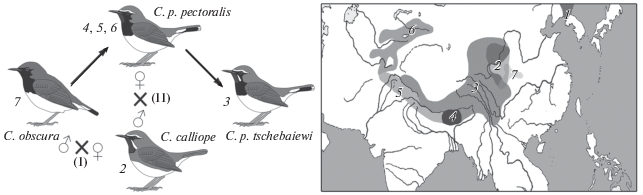
Была предложена гипотеза происхождения черногрудой красношейки (Calliope pectoralis) в результате давней гибридизации соловья-красношейки (C. calliope) и черногорлого соловья (C. obscura) (Спиридонова, Вальчук, 2017; Спиридонова и др., 2019). Происхождение тибетской черногрудой красношейки (C. (с.) tschebaiewi) в ходе более поздней гибридизации C. pectoralis и C. calliope авторы также предполагают, интерпретируя наличие ядерной копии митохондриального генома C. calliope у “дочерней” формы C. (с.) tschebaiewi, и наоборот. Принимались во внимание также фенотипические признаки и ареалогические особенности современных форм (рис. 1).
Рис. 1.
Гипотеза гибридогенного происхождения черногрудой красношейки (Calliope pectoralis), по: Спиридонова и др., 2019 (с изменениями). Слева: схема двух предполагаемых событий межвидовой гибридизации с указанием пола птиц каждого родительского вида, в результате которых возникли подвиды C. pectoralis: I – возникновение C. pectoralis – предка подвидов C. p. confusa, C. p. pectoralis и C. p. ballioni; II – возникновение формы C. p. tschebaiewi. Справа: гнездовые ареалы видов рода Calliope и их подвидов: C. calliope (1 – C. c. calliope, 2 – С. c. beicki), С. pectoralis (3 – С. p. tschebaiewi, 4 – С. p. confuse, 5 – С. p. pectoralis, 6 – С. p. ballioni), 7 – C. obscura.
Из классической БКВ вытекает, что гибридизация животных как явление – это лишь нарушение репродуктивной изоляции (Майр, 1968, 1974), поэтому она не должна рассматриваться как фактор видообразования. Однако на беспозвоночных было показано, что интрогрессия генов может быть источником генетической изменчивости для адаптации в новых условиях среды и может привести к быстрой адаптивной эволюции (Lewontin, Birch, 1966), а счет гибридогенных видов среди животных (исключая высших позвоночных) идет уже на сотни (например, Боркин, Литвинчук, 2013). Роль межвидовой гибридизации в формировании современного многообразия птиц остается по сути неизученной, во многом из-за традиционного неприятия такой возможности сторонниками БКВ.
Молекулярно-генетические и филогенетические критерии
Совершенствование методов оценки филогенетических взаимоотношений форм, позволяет все шире использовать их для реконструкции родственных связей внутри таксономических комплексов. Делаются попытки оценивать примерный эволюционный возраст и относительные сроки обособления конкретных таксонов и их группировок. Практически вся современная версия ФКВ по существу основана на данных молекулярных исследований. Теоретически предсказывают, что не позже третьего десятилетия 21 века молекулярные исследования надежно обоснуют систематику птиц до видового уровня (Harr, Price 2012). Но к возможности использования различных молекулярно-генетических методов, а также методик построения филограмм для оценки родства различных форм пока нужно относиться с большой осторожностью (например, Павлинов, 2005; Mayr, 2008; Wiley, Liberman, 2011; Павлинов, Любарский, 2011; Зеленков, 2015).
В частности, широкое использование генов мтДНК с целью видовой идентификации (ДНК-кодирования) привело к появлению в последние десятилетия ряда “ложных видов”, поскольку оказывалось связано с ядерными копиями митохондриальных генов, коамплифицирующимися вместе с ортологичными генами мтДНК (Song et al., 2008). В ряде случаев значимые различия по используемым митохондриальным маркерам нивелируются по причине текущей гибридизации между таксонами либо являются следствием обмена генами, происходившего в прошлом, например как у обыкновенной (Emberiza citrinella) и белошапочной (E. leucocephalos) овсянок (Irvin et al., 2009).
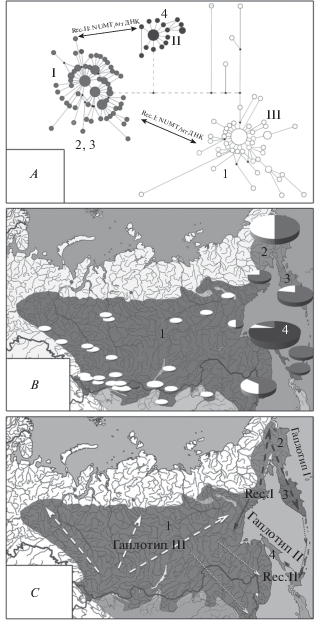
Гипотеза о возможности появления новых гаплотипов путем обратного переноса модифицированных ядерных копий в митохондриальный геном (Спиридонова и др., 2016) хорошо объясняет случаи быстрого возникновения резких молекулярно-генетических отличий у совсем молодых популяционных группировок, в который раз показав несостоятельность использования маркеров только митохондриального генома. Так у соловья-красношейки (C. calliope) на северо-востоке Азии было установлено существование двух сильно обособленных гаплотипических группировок, соответствующих дальневосточным подвидам явно недавнего происхождения (Спиридонова и др., 2017). В процессе расселения вида на северо-восток Азии и на Сахалин вероятно дважды происходило событие гомологичной рекомбинации, повлекшее обмен части митохондриального генома на соответствующую ядерную копию, существенно отличающуюся от исходного варианта, и закрепившуюся на колонизированных территориях по типу эффекта основателя (рис. 2). Значительная дивергенция митохондриальных гаплотипов далеко не всегда указывает на древность группировок, а в соответствии с упомянутой выше гипотезой, может быть следствием эффекта рекомбинации между частями ядерного генома и митоходриальным геномом. Как и в случае с происхождением черногрудой красношейки, возможность подобной рекомбинации пока выглядит дискуссионной, однако данный феномен известен или предполагается и для других групп живых организмов (Потокина и др., 2017; Гребельный и др., 2018).
Рис. 2.
Вероятные пути появления новых гаплотипов путем обратного переноса модифицированных ядерных копий в митохондриальный геном, по: Спиридонова и др., 2017 (с изменениями). A – Филогенетическая сеть гаплотипов мтДНК гена cyt b C. calliope. Гаплотип I [C. c. anadyrensis (2)–C. c. camtschatkensis (3)]; гаплотип II [C. c. sachalinensis (4)]; гаплотип III [C. c. calliope (1)]. Стрелки между гаплогруппами обозначают прошедшее рекомбинационное событие между ядерными копиями мтДНК (NUMT) и мтДНК, в результате которого возник новый гаплотип. B – Географическое распределение митохондриальных гаплотипов C. calliope по ареалу. Размер круга отображает количество образцов. Сектор в круге отображает представленность гаплогрупп: серый цвет – гаплогруппа I, черный – II, белый – III. C – Гипотетическая схема демографической истории подвидов C. calliope, построенная по данным последовательностей гена cyt b. Большие стрелки обозначают первый этап распространения, маленькие стрелки – второй. Белые стрелки указывают гаплогруппу C.c. calliope, серые стрелки – гаплогруппу C.c. anadyrensis–C.c. camtschatkensis. Rec I и Rec II – события рекомбинации.
Наконец, важным моментом стало осознание систематиками факта, что глубокие или слабые молекулярные различия далеко не всегда соответствуют большим или малым фенотипическим различиям. Известны случаи, когда симпатрично обитающие биологические виды оказываются генетически неразличимыми по традиционно используемым маркерам. Например, у ряда морфологически близких, но надежно различимых видов, для которых в условиях симпатричного распространения широкой гибридизации не происходит и/или доказана строгая ассортативность спаривания, различия по маркерам мтДНК могут быть минимальными или вовсе отсутствовать. Таковы случаи с представителями комплекса северопалеарктических снегирей (Pyrrhula pyrrhula, P. griseiventris, P. cineracea) (Arnaiz-Villena et al., 2001; Töpfer et al., 2011; Ивушкин, 2015) и тремя видами клестов (Loxia curvirostra, L. scotica, L. pytyopsittacus) на Британских о-вах (Piertney et al., 2001; Summers et al., 2002; 2007). В свою очередь, популяции (даже не географические расы) одного вида могут обладать генетическими различиями, соответствующими обычным дистанциям между “хорошими” видами. Таковы, например, различия в мтДНК двух популяций горихвостки-лысушки (Phoenicurus phoenicurus) из Германии, составляющие 5% (del Hoyo, Collar, 2014). Один и тот же вид в разных частях ареала может иметь различный набор мтДНК (в районах симпатрии с близким видом – нередко чужой, заимствованный в результате гибридизации), что, как давно известно, никак не сказывается на его фенотипе и экологических характеристиках (Vijay et al., 2016).
Практически все дискуссионные, с точки зрения классической систематики, таксономические решения оказываются следствием попыток реконструировать филогенетические связи внутри тех или иных групп на основе анализа одного или нескольких отдельно взятых митохондриальных или ядерных маркеров. Такой анализ, как правило, проводится без соотнесения результатов молекулярно-генетических исследований с элементарными представлениями о морфологическом облике объектов, их экологической специфике, а также без учета сведений о характере репродуктивных и пространственных взаимоотношений между ними.
Нужно отметить, что при систематизации сложных околовидовых групп, включающих формы разного эволюционного уровня (от географических рас до хороших видов), взаимоотношения которых осложнены случаями гибридизации (и, как следствие, возникновением гибридогенного полиморфизма), применение филогенетического метода остается единственным средством реконструкции родственных связей, необходимых для правильной таксономической оценки форм (Редькин и др., 2015). Самые противоречивые результаты филогенетических построений, в конечном счете, удается правильно обосновать благодаря комплексному подходу с обязательным включением фенотипических, эколого-морфологических (в самом широком плане) и молекулярно-генетических данных с перекрестной проверкой результатов (Абрамсон, 2013). Именно такой подход называют “интегративной таксономией” (см. обзор Sangster, 2018).
Фенотипические и другие “классические” критерии
Традиционные критерии оценки уровня дивергенции спорных таксонов, несмотря на известную субъективность, применяются орнитосистематиками. Их набор даже расширяется за счет вовлечения в анализ не только морфологических, но и поведенческих, экологических различий, особенностей вокализации, фенологии, гнездовой биологии, географического размещения и прочих аспектов (Rasmussen, Anderton, 2005). Для изолированных форм с разным успехом используется метод, когда уровень различий в паре аллопатрических таксонов оценивается путем сравнения с уровнем различий близких симпатричных таксонов с доказанным видовым статусом (Helbig et al., 2002).
В рамках недавно предложенной системы количественных критериев разграничения видов (Tobias et al., 2010) фенотипические различия (т.е. различия в оперении, морфологии, морфометрии, вокализации) оцениваются по четырехбалльной шкале в зависимости от степени их выраженности. Взаимозависимые различия (например, более длинное крыло и пропорционально больший клюв) могут быть оценены только один раз. Чтобы избежать преувеличенной оценки незначительных различий, могут оцениваться только три окрасочных, два морфометрических и два акустических различия. Молекулярным различиям между таксонами не даются количественные оценки, поскольку генетические и фенотипические различия не коррелируют. Также принимаются во внимание экологические и поведенческие различия, и, если они существуют, они получают 1 дополнительный балл (или 2 балла для “неперекрывающихся отличий в брачном поведении”). Наконец, учитываются особенности распространения: аллопатрические (независимо от того, как сильно они разделены) ареалы вовсе не дают баллов, парапатрия добавляет 3 балла, узкая зона гибридизации – 2 балла и широкая зона гибридизации – 1 балл. Таксоны, набирающие в сравнении в общей сложности 7 и более баллов, считаются отличимыми достаточно, чтобы получить статус полноценных видов (на основании балльностей, набираемых сходными симпатрическими видами, и по сравнению с более низкими баллами для таксонов, которые общепризнанно считаются подвидами). “Критерии Тобиаса” не применяют в качестве по-настоящему объективного метода, а скорее используют как практический инструмент, помогающий оценить степень различий между несимпатрическими таксонами максимально более последовательным образом.
При кажущейся очевидности и логичности подхода, он выглядит, на наш взгляд, излишне “механистичным”. Его использование на практике, как правило, приводит к дроблению таксонов, случаи укрупнения единичны (del Hoyo, Collar, 2014; 2016). Примененные нами к спорным таксонам птиц фауны Северной Евразии “критерии Тобиаса” практически всегда подтверждали их видовой статус.
Понятие “вид” применительно к птицам
Современные обзоры (например, Крюков, 2003; Coyne, Orr, 2004; Mallet, 2006; Price, 2008; Васильева, 2009; Павлинов, Любарский, 2011; Павлинов, 2017, 2018, 2019; Рубцов 2015; Sangster, 2018) по-разному трактуют и оценивают сильные и слабые стороны основных соперничающих концепций вида, а также эволюционной и филогенетической парадигм, их способность отражать процессы видообразования и применимость в таксономии. В большинстве обзоров признается, что понятие вида не универсально, в любом случае следует разграничивать таксономическую и эволюционную сущности категории «вид». Появляются и новые определения вида. Сторонники БКВ (Johnson et al., 1999) и сторонники ФКВ (Sangster, 2014) сблизили позиции, в результате была предложена совместно разработанная “Комплексная биологическая концепция вида”: “Вид у птиц представляет собой систему популяций, являющуюся по существу монофилетической, генетически связанной и генеалогически согласованной родственной группой особей, которые имеют одинаковую во времени и пространстве систему размножения, представляют собой независимую эволюционную линию и демонстрируют существенную, но не обязательно полную репродуктивную изоляцию от других таких систем” (цит. по: del Hoyo, Collar, 2014). Таким образом, неполная изоляция и наличие гибридов не являются препятствием для придания видового статуса четко различимым формам, ранее считавшимся конспецифичными.
Некоторые отечественные исследователи сомневаются в принципиальной возможности компромисса между БКВ и ФКВ. А.С. Рубцов не без оснований считает ФКВ попыткой возврата к типологической концепции вида. Сам он определяет “Процесс видообразования как установление симпатрии между дивергирующими таксонами, а виды — как морфологически различимые группы организмов, способные сосуществовать в зоне симпатрии в течение длительного времени” (2015). Однако, исходя из этого определения, мы не должны считать видами никакие парапатрические таксоны, даже если невозможность их симпатрии и трансграничных контактов обусловлена иными факторами, а не наличием интрогрессивной гибридизации.
Михайлов (2003, 2015, 2017, 2018) рассматривает вид как наименьшую единицу морфо-генетической устойчивости и канализованную норму реакции, т.е. как результат разнообразных процессов, а не стадию континуума. В кратком выражении это эпигенотип, т.е. динамичный архетип осуществления формы в череде поколений. Михайлов считает маркером истинного видового статуса лишь репродуктивную изоляцию, которая выражается в генетической несовместимости (посткопулятивная изоляция) или обусловлена мешающей спариванию морфологией, несовпадением запаховых сигналов и других неповеденческих параметров. Поведенческую же, прекопулятивную изоляцию (столь характерную именно для птиц и млекопитающих) он низводит до обратимой ассортативности скрещивания, основанной на избирательности предпочтений при образовании пар.
При этом не принимается во внимание, что несовпадение брачного ритуала у птиц представляет собой ничуть не менее (а порой и более) эффективный вариант репродуктивной изоляции, чем, например, фактор сниженной выживаемости гибридов, обусловленный генетическими причинами. Иными словами, упор делается не на степень воздействия (эффективность) изолирующих механизмов, а лишь на их природу. Следуя этой логике, установить видовой статус для симпатричных родственных форм, исходя только из отсутствия видимой гибридизации в природе, мы не можем – нужно каждый раз устанавливать, какие механизмы ей препятствуют. Эксперименты в неволе вряд ли могут считаться решающими, поскольку животные неизбежно находятся под влиянием условий ex situ (вне естественных мест обитания), и причины успеха/неуспеха размножения сложно оценить количественно и качественно (Frankham, 2008). Непонятно также, каким образом происходит дальнейшая дивергенция уже не скрещивающихся “поведенческих” форм, приводящая (в условиях симпатрии!) к возникновению “настоящих” морфовидов, представляют ли собой разные морфовиды парапатрические формы без признаков гибридизации и т.д.
Появление поведенческой прекопулятивной изоляции без сопутствующей посткопулятивной, на наш взгляд, напротив, может считаться эволюционным прорывом – возникновением фактора, который достаточно эффективно (но не специально) удерживает группу от “размывания” и делает излишним существование затратных генетических, биохимических и морфологических преград при условии достижения таксоном высокого уровня нервной деятельности. Однако мы не исключаем, что поведенческая прекопулятивная изоляция лишь маркирует уровень дивергенции, уже сопровождающийся хотя бы неполной и неявной посткопулятивной изоляцией форм.
Исходя из практических задач, мы сочли необходимым сформулировать такое определение вида, чтобы оно в большей степени соответствовало известной на сегодняшний день ситуации, касающейся птиц, и сочеталось бы с определениями таксонов более низкого ранга. Предлагаемое здесь определение базируется на двух ведущих критериях: биологическом, состоящем в оценке репродуктивных взаимоотношений конкретных форм друг с другом, и филогенетическом, состоящем в оценке степени эволюционного родства таксонов как на молекулярно-генетическом, так и на сравнительно-морфологическом материале.
Вид (species) – совокупность популяций, обладающая эволюционной общностью (включая морфологический, генетический, экологический аспекты) и способная поддерживать свою стабильность при контактах с другими видами на протяжении длительного времени. Основным критерием вида, очевидно, должны считаться не существующие или потенциальные ограничения в гибридизации, а его стабильность во времени и пространстве. Следует подчеркнуть, что поддержание стабильности подразумевает не только репродуктивную изоляцию разного уровня и разной природы, но и другие свойства популяций в районах контакта – взаимоизбегание (например, расхождение по экологическим нишам и стациям, сдвиг репродуктивных циклов) и/или прямую конкуренцию за ресурсы с взаимоисключением. Не все виды способны жить симпатрично, даже при отсутствии гибридизации между ними. В соответствии с теорией прерывистого равновесия (“пунктирной эволюции”) Н. Элдриджа/С. Гулда (получившей не так давно очередные подтверждения, см. Millien, 2006; Pagel et al., 2006), стабильность вида должна прослеживаться на значительном отрезке времени (от нескольких тысяч лет), и пока не следует считать полноценными видами гибридогенные формы, эко- и социорасы, возникшие “на наших глазах”, даже при их соответствии критериям классической БКВ (например, как в случае с “новым” галапагосским вьюрком).
Надо отметить, что наше определение в значительной степени перекликается с определением вида в эволюционной концепции, сформулированным Уайли и Мейденом (Wiley, Mayden, 1985) на основании взглядов Дж. Симпсона: “Вид – это биологический объект, состоящий из организмов, сохраняющий свою индивидуальность во времени и пространстве, и имеющий свою собственную эволюционную судьбу и исторические тенденции”. Вместе с тем оно более детализировано и не содержит нечетких утверждений относительно “судьбы” и “тенденций”. Мы полагаем неизбежным использовать критерии ФКВ при анализе таксонов с аллопатрическим распространением, при обнаружении гибридогенного полиморфизма и в других сложных случаях (Редькин и др., 2015; Red’kin et al., 2016).
Геном почти любого вида включает чужеродную ДНК, свидетельствующую, помимо прочего, о периодической гибридизации с другими видами (например, Hailer et al., 2012; Miller et al., 2012; Soubrier et al., 2016). Тем не менее, даже при значительной примеси чужеродного генома вид длительное время сохраняет свою эколого-морфологическую специфику (вероятно, до вызванного разными причинами наступления периода “разбалансированности”).
Понятие “подвид”
В качестве антитезы к понятию “вид”, в значении “таксон, не достигший видовой самостоятельности”, “не-вид”, нередко ошибочно используют термин “подвид”. Понятие “подвид” (subspecies) было окончательно узаконено во второй половине 19 в. для обозначения единиц внутривидовой географической изменчивости (см. обзор: Винарский, 2015) и в настоящее время представляет собой наименьшую таксономическую категорию, регламентируемую Международным кодексом зоологической номенклатуры (2004). Основной подход к изучению географической изменчивости вида исторически сводился к описанию внешних морфологических отличий одной группы особей, географически удаленной от другой группы особей того же вида и ее номенклатурному обозначению.
Для всех подвидов применяется номенклатурное обозначение с иерархическим (род, вид, подвид) триноминальным названием. Критериями для выделения подвида являются устойчивые морфологические отличия, часто при минимальных различиях экологических особенностей, или отсутствии таковых. Как ни странно, термин “подвид” продолжает оставаться весьма расплывчатым, ввиду разных значений, ему придаваемых (Винарский, 2015). Существует даже тенденция избегать слова “подвид”, заменяя его синонимами “раса”, “разновидность”. “Разновидности различаются немногими, чисто поверхностными признаками, а виды – множеством взаимообусловленных свойств” (Панов, 1993).
Проявление географической изменчивости внешних морфологических признаков свойственно подавляющему большинству животных. Чаще всего такая изменчивость выражается в образовании географических рас, в большей или меньшей степени отражающих популяционную структуру видов. Географические расы могут быть как резко дифференцированными (но в меньшей степени, чем виды внутри данного рода или подрода), так и относительно слабо (но статистически достоверно) различимыми (например, Red’kin, Konovalova, 2006). В составе политипического вида могут оказаться расы разного возраста: от недавно изолированных, но имеющих достоверные отличия (закрепившиеся благодаря эффекту основателя), до реликтовых, длительное время существовавших в условиях надежной пространственной изоляции, но в ряде случаев, вступающих в интерградацию. В местах контакта (если он имеет место) соответствующих морфологически обособленных популяций (или их группировок) образуются переходные зоны – зоны интерградации (или районы неограниченного смешивания признаков). Ширина таких зон существенно варьирует, но она всегда меньше площади областей, занятых “фенотипически чистыми” популяциями, т.е. областей, в пределах которых морфологические признаки соответствующих подвидов стабильны.
Клинальная (географическая) изменчивость принимается нами, вслед за многими авторитетами, как постепенное изменение одного или нескольких морфологических признаков на значительном расстоянии в пределах географически непрерывного континуума популяций данного вида. В пределах этого континуума практически невозможно выделение зон, в которых данные признаки становятся стабильными. Среди птиц палеарктической фауны случаи клинальной изменчивости объективно редки и мало изучены. В качестве примера можно привести изменчивость окраски оперения у дроздовидной камышевки (Acrocephalus arundinaceus), выраженную в постепенном осветлении общей окраски с запада на восток. Восточные популяции этого вида ранее пытались выделить в подвид “A. a. zarudnyi” (Дементьев, 1937; Портенко, 1960; Vaurie, 1959). Однако такая точка зрения не находит подтверждения вследствие невозможности наметить области постоянства проявления окрасочных отличий (Опаев, 2010). В некоторых случаях, в пределах очень широко распространенного подвида, например северопалеарктического ополовника (Aegithalos caudatus caudatus), изменчивость ряда размерных показателей на пространстве от Скандинавии до побережья Охотского моря может иметь флуктуирующий характер, без достоверных различий между соседними географическими выборками (Лукьянчук и др., 2017).
Во многих случаях исследователям, проводившим ревизии подвидовой систематики, не удавалось достигнуть консенсуса о числе географических рас вида. Часто таксономические решения принимались экспертами на основе изучения коллекционных материалов, “имеющихся у них под рукой”. Однако материалы распределены по музейным коллекциям крайне неравномерно и порой не отражают реальной картины географической изменчивости. Одна из причин возникающих разночтений – исследователи, ревизующие виды, не имеют возможности обработать многочисленные, но разрозненные материалы с одинаковой степенью подробности (Редькин, 2015).
В настоящее время филогеографические исследования позиционируются как одно из основных направлений в познании популяционно-генетической структуры вида и влияния гибридизации в зонах контакта родственных таксонов (Политов, Мудрик, 2018). Однако картина, выявленная в результате таких исследований политипических видов птиц, в большинстве случаев, не совпадает с традиционными представлениями о географической изменчивости и внутривидовой систематике этих группировок.
Обычно прямая взаимосвязь между гаплогруппами и географическими расами прослеживается у видов, чрезвычайно слабо подверженных географической изменчивости. Таков, например, грач (Corvus frugilegus), обособленность двух хорошо выраженных подвидов которого четко поддерживается филогеографическими построениями. Вообще же филогеографическая структура вида, выявленная методами анализа мтДНК, крайне редко отражает географическую изменчивость морфологических признаков птиц (Haring et al., 2007), что дает филогеографам основания обвинять морфологов в “фантомности” ряда подвидов (Zink, 2004).
Для всех широко распространенных политипических видов наличие филогеографических структур, маркирующее группировки или расы, удается проследить только в отдельных частях гнездового ареала, тогда как на большей его части обособленность местных форм совершенно не обнаруживается посредством таких методик. Так, надежно географически изолированная камчатская раса сороки (Pica pica camtschatica) резко отличима по молекулярным маркерам, от 6 рас “западной” группировки, контактирующих друг с другом, и не имеющих филогеографических различий (Kryukov et al., 2017).
Возникновение географических рас может произойти буквально за несколько десятков лет. Например, в популяциях интродуцированного красноухого бюльбюля (Pycnonotus jocosus), образовавшихся на островах Реюньон, Маврикий и Оаху, возникли заметные морфологические отличия от предковых популяций. При этом на о. Реюньон за 30 лет сформировались 2 популяции, имеющие и генетические различия между собой (Le Gros et al., 2016). Для серебряных белоглазок (Zosterops lateralis), быстро расселившихся на островах близ Австралии, показано, что увеличение размеров тела островных форм произошло в течение всего 200 лет. При этом размеры формы Z. l. chlorocephalus, возникшей за это время на о. Херон, почти на треть превышают размеры ее материкового предка Z. l. familiaris (Clegg et al., 2002).
В некоторых случаях причиной проявления признаков, маркирующих различия популяционных группировок, становится происходившая в прошлом или происходящая в настоящее время межвидовая гибридизация. Яркими примерами таких форм являются восточная обыкновенная овсянка (Emberiza citrinella erythrogenys) (Панов 1989, 2001), североевропейская желтая трясогузка (Motacilla flava thunbergi) (Редькин, 2001а,б; Сотников, 2006), равнинный туркестанский жулан (Lanius phoenicuroides karelini) (Панов, 2008).
В качестве подвидов как таксономической категории по нашему мнению, следует квалифицировать любые географические формы вида, статистически достоверно отличающиеся по морфологическим признакам, вне зависимости от их эволюционного возраста. Описание новых подвидов (с их номенклатурным обозначением) представляется целесообразным до той степени дробности, при которой они будут наиболее полно отражать географическую (популяционную) структуру данного вида. Популяции/формы, имеющие гибридогенное происхождение (доказанное или предположительное), исследователи обычно либо игнорируют в плане номенклатурного обозначения (тогда их названия даются в кавычках), либо никак не выделяют среди прочих подвидовых названий. На наш взгляд, такие формы, если они отвечают критериям выделения географических рас, целесообразно обозначать триноминальным именем, сопровождаемым каким-либо значком (например, астериском – Emberiza citrinella erythrogenys*), который не регламентируется Кодексом, но который прямо указывает на гибридогенное происхождение таксона.
Считать любой подвид потенциальным видом неверно. Географическая раса способна достигнуть видового уровня дивергенции лишь при условии длительной изоляции в рефугиуме. Тем не менее, в классической ФКВ для подвидов БКВ просто “не осталось места” – эти таксоны получают ранг видов (см. обзор: Винарский, 2015, 2015а).
Определение подвида мы формулируем так: Подвид (=географическая раса) – популяция или группа популяций вида, которая имеет устойчивые морфологические отличия от соседних популяций, но может свободно скрещиваться с ними в районах контакта (при наличии таковых) с образованием переходных зон (зон интерградации). Симпатрия подвидов без образования зон интерградации у птиц практически невозможна. Отсутствие репродуктивной изоляции ожидаемо и между представителями географически изолированных подвидов, причисляемых к общему виду на основании морфологических (иногда и генетических) данных.
Интересным исключением (в отношении симпатрии конспецифичных форм) являются ситуации с так называемыми экологическими расами, изученные на примере клеста-еловика (Loxia curvirostra). В североамериканских популяциях этого вида выделяют 9 экологических рас (Parchman et al., 2006), еще 7 таких рас известно для западноевропейских популяций (Edelaar et al., 2008). Эти расы сходны по структуре песен, но хорошо различимы по позывкам и, будучи адаптированы к питанию на разных видах хвойных, различаются по нескольким морфометрическим признакам, в первую очередь по строению клюва. Считается, что дивергенция экологических рас клеста началась не ранее 11 тыс. лет назад, с наступлением современной постледниковой эпохи (Benkman et al., 2010). Этот вывод согласуется с данными молекулярных исследований: экологические расы еловика не различаются по мтДНК, но демонстрируют устойчивые различия на сериях по мультилокусным маркерам. Вне периода размножения клесты формируют кочующие стаи, каждая стая состоит из особей только одной экологической расы (при этом все члены стаи издают только один тип позывок), а брачные пары формируются между членами одной стаи. Если строго следовать критериям БКВ, экорасы клеста-еловика можно трактовать как “самостоятельные виды на ранних этапах дивергенции” (Edelaar, 2008) и расценивать в качестве возможного доказательства симпатрического видообразования у птиц. Однако, учитывая номадный характер гнездования и кочевок клестов (типичный для многих вьюрковых), мы предполагаем, что экорасы формировались аллопатрично, а их симпатрия возникала как вторичное явление, в результате номадных переселений, обусловленных урожаями и неурожаями семян конкретных хвойных пород. На наш взгляд, они могут считаться эквивалентом нормальных географических рас, и номенклатурно обозначаться как подвиды, поскольку в условиях периодического сокращения пищевых ресурсов репродуктивная изоляция между ними может нарушаться, приводя к обмену генами и, возможному слиянию друг с другом.
Понятие “полувид”
Термин “полувид” (semispecies) был введен Эрнстом Майром в 1942 г. (Mayr, 1942). Согласно его определению “Формы, находящиеся на заключительных стадиях дивергенции, обозначаются в рамках БКВ термином “полувиды”, под которыми понимают близкородственные таксоны, достигшие видового уровня дивергенции, но видовой статус которых не может быть подтвержден из-за их аллопатричного распространения или в силу неполной репродуктивной изоляции. В отличие от подвидов, полувиды имеют частично сформированные изолирующие механизмы, поэтому гибридизация между ними не носит характер интрогрессии и не приводит к их слиянию” (Майр, 1968).
Во времена становления БКВ отсутствовали многие возможности оценки уровня обособленности таксонов, каковыми мы располагаем сейчас, поэтому в понятие полувидов автоматически попадали практически все гибридизирующие и все морфологически сходные аллопатрично распространенные сколько-нибудь родственные виды. Разнобой оценок феномена полувидов наблюдается и поныне – от предложения считать полувид одной из обязательных ступеней таксонов низшего ранга (Пфандер, 2018) до полного отрицания биологического смысла этого понятия и его трактовки как буферной категории (“мусорной корзины”) для форм с неизученными родственными связями и репродуктивными отношениями (Михайлов, 2018). Понятием “полувиды” (semispecies, allospecies) активно оперировал Степанян (1983), разделив надвидовые комплексы на категории superspecies и ex-conspecies. Использует понятие semispecies и Рубцов (2015). Териологи (например, Павлинов, 2019) в настоящее время предпочитают термин алловид (allospecies).
Выделение пограничной таксономической категории “полувид” специально не регламентировано “Кодексом зоологической номенклатуры” (2004). Таким образом, любые формы, ранг которых признан более высоким, чем подвидовой, могут приравниваться к видам и фигурировать далее под биноминальными латинскими названиями, например, в нормативных документах природоохранной направленности. Однако, по нашему мнению, при составлении списков авифаун, а также в специальных работах посвященных эволюции и систематике таксономических групп, применение данной категории представляется вполне оправданным, поскольку это все же не виды в полном смысле слова.
В нашей трактовке “Полувид” – это популяция или совокупность популяций, которая не по всем критериям соответствует понятию вида, но имеет более значимые, чем у подвидов, различия с соседними популяциями и способна образовывать с ними зоны симпатрии без интерградации в них. Во всех случаях полувиды – это молодые, но уже сильно обособившиеся формы (или группировки форм), демонстрирующие неполную репродуктивную изоляцию при контактах с близкими формами.
Для полувидов, составляющих один надвид (superspecies), наиболее типично парапатрическое распространение. Если сближенные границы их ареалов не заданы естественными линейными барьерами (руслами рек, хребтами и т.д.), то парапатрия таких форм часто выглядит следующим образом: узкие зоны пространственного контакта перемежаются лакунами, где оба члена пары отсутствуют, или имеют мозаичное очаговое распространение с низкой численностью. Пространственные отношения такого рода характерны, например, для зарничек (Phylloscopus (inornatus) inornatus/Ph. (inornatus) humei) (Редькин, Коновалова, 2003; Red’kin, Konovalova, 2004) или черноголовых чеканов (Saxicola (torguata) maura/S. (torguata) stejnegeri) (Opaev et al., 2018). В районах непосредственного контакта полувидов не образуются зоны интерградации, характерные для географических рас одного вида (Крюков, Блинов, 1981; Блинов и др., 1993; Марова, Шипилина, 2015). Наличие лакун и очагового характера распространения также объяснимо: расходясь по требованиям к среде обитания сильнее, чем географические расы, полувиды на краях ареалов могут находитьтся в пессимальных (субоптимальных) условиях. Это обстоятельство не позволяет полувидам заселить “спорное” пространство с нормальной плотностью, характерной для популяций единого вида.
Пример парапатрического распространения – два политипических комплекса щеглов: собственно щегол (Carduelis (carduelis) carduelis) и седоголовый щегол (C. (carduelis) caniceps). Для них свойственна узкая зона пространственного контакта, в которой существует постоянная гибридизация, но она не приводит к формированию популяций с переходными признаками (Заблоцкая, 1975). Филогенетические различия этих форм также минимальны (Arnaiz-Villena et al., 2001). Морфологические различия представителей этих комплексов глубоки и затрагивают множество окрасочных и пластических признаков. Давно известны также устойчивые различия по голосу, в т.ч. различия в структуре позывок. В отечественной орнитологической литературе комплексы традиционно трактуют как виды, в зарубежной “постмайровской” литературе – как конспецифичные формы, однако в последнем издании Illustrated Checklist of the Birds of the World (del Hoyo, Collar, 2016) они уже фигурируют как самостоятельные виды. В предлагаемой здесь трактовке щегол и седоголовый щегол – несомненные полувиды.
Известно немало случаев, когда спорные формы образуют обширные зоны симпатрии. В качестве примера можно привести комплекс северопалеарктических снегирей, в котором представлены две географически разобщенные формы: обыкновенный (Pyrrhula (pyrrhula) pyrrhula) и дальневосточный (P. (pyrrhula) griseiventris) снегири, а также серый снегирь (P. (pyrrhula) cineracea), симпатричный двум другим формам. В зарубежной литературе все эти формы считаются конспецифичными лишь по той причине, что генетически они чрезвычайно близки (Arnaiz-Villena et al., 2001). Однако факт симпатрии с крайне редкой гибридизацией противоречит такой трактовке. Помимо окраски, птицы трех форм отличаются друг от друга размерами и пропорциями, имеются и вполне уловимые на слух отличия голоса (Ивушкин, 2015). Таким образом, при объективной генетической близости эти формы не могут быть приняты лишь как географические расы. В аналогичных случаях с таксономическими парами краснозобый/чернозобый (Turdus (ruficollis) ruficollis/T. (ruficollis) atrogularis) и рыжий/бурый дрозды (T. (naumanni) naumanni/T. (naumanni) eunomus), члены которых тоже долгое время считались конспецифичными географическими расами при наличии широкой симпатрии, сейчас принято рассматривать как самостоятельные виды (Степанян, 2003; Dickinson, Christidis, 2014; del Hoyo, Collar, 2016; Clements, 2017 и др.), несмотря на регулярную гибридизацию (Портенко, 1981; Вальчук и др., 2013). Исходя из наших взглядов (Редькин и др., 2015), и упомянутые снегири, и упомянутые дрозды – полувиды.
Желтолобая трясогузка (Motacilla (flava) lutea) – форма, которую в зарубежных сводках считают лишь подвидом желтой трясогузки (Motacilla flava sensu stricto), распространена симпатрично с конспецифичными M. f. flava и M. f. beema почти на всем протяжении своего ареала, от Поволжья до юга Западной Сибири и севера Казахстана. Это противоречит общепринятым взглядам на географические расы. Кроме того, на юге часть области ее гнездования перекрывается с ареалом черноголовой трясогузки (M. (flava) feldegg). От симпатрично распространенных рас желтой и черноголовой трясогузок желтолобая резко отличается окраской самцов, биологическими особенностями и экологическими предпочтениями (Бахтадзе, Казаков, 1985; Бахтадзе, 1987; Муравьев, 1993, 1996, 1997). На основании наличия симпатрии при отсутствии видимой интрогрессивной гибридизации Степанян (1978, 1983, 2003) придавал этой форме статус монотипического вида (правда, включив в него и аллопатричную североатлантическую форму flavissima). На наш взгляд, исходя из нынешней ситуации, форму lutea (но без фенотипически сходной flavissima) правильнее квалифицировать как полувид, так как ни устойчивым видом, ни, тем более, географической расой она не является.
Аллопатрически распространенные родственные формы квалифицировать как полувиды (allospecies в узком смысле) довольно сложно. Без экспериментов в неволе (порой дающих неоднозначные результаты) мы не можем судить о степени/отсутствии репродуктивной изоляции в сравниваемых популяциях, известные же в настоящее время генетические дистанции, как и фенотипические различия, не являются универсальным критерием (см. выше). Тем не менее, в случаях, когда рассматривается весь надвидовой комплекс (включая симпатрические и парапатрические формы), можно делать выводы и о статусе его аллопатрических членов. В частности, это показано в отношении уже упоминавшегося надвида черноголовых чеканов (Saxicola torquata sensu lato) в том числе его слабоизученной изолированной формы (полувида) – чекана Пржевальского (S. (torquata) przewalskii) (Калинин и др., 2018).
В составе одних и тех же надвидовых комплексов в статусе полувидов могут быть приняты как формы, легко диагностируемые при помощи молекулярно-генетических маркеров, так и формы, генетические различия которых нивелируются благодаря происходящей или происходившей в прошлом гибридизации. Примером, может служить группа Remiz pendulinus sensu lato, в составе которой венценосный (R. (pendulinus) coronatus) и китайский (R. (pendulinus) consobrinus) ремезы генетически хорошо дифференцированы, тогда как, резко обособленные друг от друга морфологически и экологически обыкновенный (R. (pendulinus) pendulinus) и тростниковый (R. (pendulinus) macronyx) ремезы оказываются практически неразличимыми генетически (Barani-Beiranvand et al., 2017).
В качестве полувидов не могут рассматриваться фенотипически сходные, но филогенетически удаленные, не близкородственные формы или группировки форм даже в тех случаях, когда между их представителями имеет место регулярная гибридизация, как например, между фенотипически похожими дроздовидной (Acrocephalus arundinaceus) и голосистой (A. stentoreus) камышевками или между серебристой чайкой (Larus argentatus) и хохотуньей (L. cachinnans).
Мы допускаем, что, не приобретя достаточно эффективной репродуктивной изоляции, но уже выйдя из рефугиумов, полувиды с течением времени могут вновь слиться друг с другом вследствие интрогрессивной гибридизации. “Разбалансированная” форма в перспективе может быть целиком поглощена более многочисленной и устойчивой одной или несколькими родственными формами. Такое развитие событий, вероятно, имеет место в случае с желтолобой трясогузкой (Motacilla (flava) lutea), уже исчезнувшей на большей части гнездового ареала в Азии в течение буквально 100 последних лет наблюдений, где на месте прежней области ее гнездования в настоящее время встречаются лишь немногочисленные особи со следами скрещивания с желтой трясогузкой (Редькин, 2012; Рябицев, 2014).
Большинство форм, отвечающих определению полувида, на сегодняшний момент не обнаруживают явных тенденций к вторичному слиянию или “растворению” в другой форме. Чтобы с определенностью характеризовать ход и результат этих процессов, необходимы сотни (возможно, тысячи) лет наблюдений.
До сих пор не решен вопрос номенклатурного обозначения полувидов, в том числе ввиду расплывчатости самого понятия, не признаваемого многими систематиками. В идеале континуум стадий процесса видообразования от “хорошо обособленная раса/подвидовая группа” до “почти вид” (ex-conspecies в трактовке Степаняна, 1983) требует дробной иерархии с несколькими дополнительными градациями между рангами подвид и вид. Представляется полезным хотя бы введение единого обозначения полувидов как особой дополнительной категории. В орнитологических сводках последних лет полувиды, фигурирующие под биноминальными названиями, составляют “львиную долю” тех самых сотен “новых видов”, явившихся причиной раздражения, обвинений орнитосистематиков в чрезмерном дробительстве и отказе от БКВ. Но такие же полувиды оказываются “замаскированными” в сводках и в качестве групп подвидов, для удобства снабженных метками разных цветов (del Hoyo, Collar, 2014, 2016). Мы полагаем наиболее корректным (Коблик и др., 2006; Коблик, Архипов, 2014; Редькин и др., 2015 и др.) применять для всех полувидов биноминальные латинские названия с обязательным обозначением их принадлежности к конкретной видовой группе (надвиду). Международным кодексом зоологической номенклатуры (2004) допускается использование старшего названия в круглых скобках между названием родовой группы и видовым именем для обозначения видовой группы (статья 6 “Интеркалярные названия”, пункт 2). Такое обозначение, например Parus (major) bokharensis, четко показывает отличие таксона и от вида, и от географической расы.
Считая, вслед за Майром (1968, 1971) и Степаняном (1983) географическую изменчивость и видообразование несколько разными явлениями, мы полагаем, что полувиды (даже находящиеся на ранних стадиях становления) должны фигурировать в сводках и каталогах в видовых списках (с указанием надвида в скобках), а не в перечнях подвидов внутри вида “в широком понимании”.
* * *
На современном уровне наших знаний ни одна из групп критериев вида не может претендовать на универсальность. При неизбежной эклектичности и субъективности комплексного подхода (“интегративной таксономии”) пока только такая оценка разнообразных признаков позволяет принимать адекватные решения в пограничных ситуациях видообразования у птиц.
Признавая проблему “неуниверсальности” трактовок, мы, тем не менее, впервые попытались дать во взаимосвязи три понятия – вида, подвида и полувида у птиц, сформулировав их с учетом практического применения появившихся в последние десятилетия данных и концепций. Рубцов (2015) в концептуально близкой нашим взглядам теоретической работе “Репродуктивная изоляция и понятие вида у птиц” приводит в виде таблицы стадии видообразования с характеристиками ситуаций в зонах контакта. Выделяемые им этапы и соответствующие варианты придания статуса формам, находящимся на этих этапах, в общих чертах соответствуют предлагаемым нами понятиям, в том числе понятию полувида, как таксона, находящегося на пути эволюции в самостоятельный вид (табл. 1).
Таблица 1.
Стадии видообразования и обоснование реальности видов (Рубцов, 2015)
| № | Таксономический статус | Экологическая сегрегация/ Ассортативность спаривания | Развитие ситуации в зоне вторичного контакта |
|---|---|---|---|
| 1а | Экологические расы | Есть/есть | При резком сокращении ресурсов нарушаются механизмы прекопуляционной изоляции. Массовая гибридизация при отсутствии пространственной изоляции между дивергирующими таксонами приводит к их полному слиянию |
| 1б | Подвиды | Нет/нет | Массовая гибридизация в постепенно расширяющейся зоне контакта приведет к слиянию таксонов с течением времени |
| 1в | Подвиды | Есть/нет | То же, что и 1б, но расширение зоны контакта и слияние таксонов происходит быстрее (Грабовский, 1995) |
| Нижняя граница вида | |||
| 2а | Полувиды | Нет/есть | Образование узкой и стабильной во времени зоны вторичного контакта |
| 2б | Полувиды | Есть/есть | Образование широкой зоны контакта, случайная гибридизация. Если гибриды более конкурентноспособны при выборе местообитаний, чем родительские формы, то уровень гибридизации постепенно нарастает и экологическая сегрегация между дивергирующими таксонами исчезает, что приводит к сокращению зоны контакта и установлению узкой и стабильной во времени гибридной зоны |
| Верхняя граница вида | |||
| 3 | Сестринские виды | Есть/есть | Если гибриды при выборе местообитаний менее конкурентноспособны, чем родительские формы, то уровень гибридизации остается низким, образуется широкая зона симпатрии |
По нашим оценкам, в составе авифауны России и сопредельных территорий присутствуют не менее 40 полувидов, составляющих ядро спорных таксономических случаев в этом регионе (Редькин и др., 2015). Предлагаемое нами решение – регламентация понятия “полувид” в рамках широко трактуемой БКВ и введение полувидов как дополнительной категории, позволяет, не выходя за пределы требований таксономической номенклатуры, обозначить непрерывность процесса эволюции видов. Именно к полувидам, видимо, должны относиться большинство слабо дифференцированных “видов”, выделение которых в последние десятилетия вызывает обвинения в чрезмерном дробительстве.
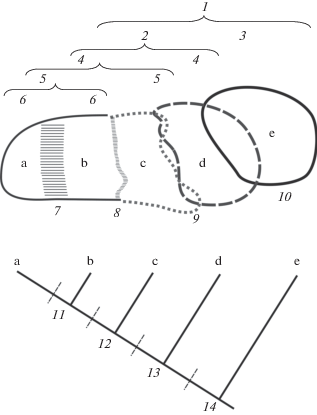
Предложение ввести в нормальный таксономический обиход понятие “полувид” в качестве промежуточной категории, отражающей незавершенные стадии процесса видообразования, с нашей точки зрения, не противоречит биологической и эволюционной концепциям вида (полувиды заполняют хиатус между ними). С позиций типологической и филогенетической концепций вида полувиды должны считаться полноценными видами, а с позиций зоогеографической концепции вида – не должны считаться таковыми (рис. 3).
Рис. 3.
Названия низших таксонов в биологической концепции вида. Верхняя часть схемы показывает пространственный контакт между таксонами, нижняя – границы соответствующих видов в рамках разных концепций. 1 – видовая группа; 2 – зоогеографический вид – надвид; 3 – зоогеографический вид – изовид (самостоятельный вид); 4 – парапатрический вид; 5 – группа подвидов, мегаподвид; 6 – подвид; 7 – широкая (свыше 100 км) зона интерградации; 8 – узкая гибридная зона; 9 – зона парапатрического контакта; 10 – широкое перекрытие ареалов; 11 – филогенетический вид (ФКВ); 12 – эволюционный вид; 13 – биологический вид (БКВ); 14 – зоогеографический вид. В этом примере 5, 4, 3 или 2 таксономических вида признаются в зависимости от того, какая концепция вида применяется и где установлена соответствующая граница вида (по: Haffer, 1997).
Мы отдаем себе отчет, что решение “проблемы вида” в философском ключе может осуществляться самыми разными способами, включая гибкий, фактически “безранговый” филогеографический подход (Павлинов, 2018). Возможно, предлагаемые нами определения и решения могут показаться кому-то неоптимальными, частными и консервативными. Тем не менее, по нашему опыту, они наилучшим образом работают на практике при оценке спорных и пограничных ситуаций среди птиц в рамках общепринятых подходов в таксономии и научной номенклатуре.
Список литературы
Абрамсон Н.И., 2013. Молекулярная и традиционная филогенетика. На пути к взаимопониманию // Современные проблемы биологической систематики. Труды ЗИН РАН. Прил. 2. С. 219–229.
Аверьянов А.О., Лопатин А.В., 2014. Макросистематика плацентарных млекопитающих: современное состояние проблемы // Зоологический журнал. Т. 97. № 7. С. 798–813.
Бахтадзе Г.Б., 1987. Распределение пигментов в перьях желтых трясогузок (Motacilla flava L., M. feldegg Mich., M. lutea Gmel.) и окраска их оперения // Известия Северо-Кавказского науч. центра высш. шк. Сер. естеств. науки. Ростов-на-Дону: Деп. в ВИНИТИ, 18. 08 1987. № 1920. 5 с.
Бахтадзе Г.Б., Казаков Б.А., 1985. Сравнительный анализ мест гнездования сероголовой, черноголовой и желтолобой желтых трясогузок на юге Европейской части СССР // Вестник зоологии. № 5. С. 55–59.
Блинов В.Н., Блинова Т.К., Крюков А.П., 1993. Взаимодействие серой и черной ворон (Corvus cornix L., С. corone L.) в зоне симпатрии и гибридизации: структура зоны и возможные факторы изоляции // Гибридизация и проблема вида у позвоночных. Труды Зоол. музея МГУ. Т. 30. С. 97–117.
Боркин Л.Я., Литвинчук С.Н., 2013. Гибридизация, видообразование и систематика животных // Современные проблемы биологической систематики. Труды ЗИН РАН. Прил. 2. С. 83–139.
Вальчук О.П., Редькин Я.А., Сотников В.Н., 2013. Первая встреча краснозобого дрозда Turdus ruficollis и новые находки птиц с фенотипами группы темнозобых дроздов T. ruficollis и T. atrogularis в Приморье // Русский орнитологический журнал. Т. 22. № 947. С. 3315–3320.
Васильева Л.Н., 2009. Иерархия и комбинаторика признаков в эволюционной систематике // Труды ЗИН РАН. Прил. 1. С. 235–249.
Винарский М.В., 2015. Судьба категории подвида в зоологической систематике. 1. История // Журнал общей биологии. Т. 76. № 1. С. 3–14.
Винарский М.В., 2015б. Судьба категории подвида в зоологической систематике. 2. Современность // Журнал общей биологии. Т. 76. № 2. С. 99–110.
Гребельный С.Д., Иванова Н.Ю., Нефедова Е.А., 2018. Обмен между ядерным и митохондриальным геномами (по результатам анализа ядерных и цитоплазматических копий митохондриальных генов — numts и cymts) // Цитология. Т. 60. № 11. С. 899–902.
Дементьев Г.П., 1937. Полный определитель птиц СССР: Воробьиные. Т. 4. М.-Л. 334 с.
Заблоцкая М.М., 1975. Голосовые реакции седоголовых и черноголовых щеглов (Passeres, Carduelinae): Форма и функциональное значение позывов // Бюллетень Московского общества испытателей природы. Отд. биол. Т. 80. № 3. С. 22–38.
Зеленков Н.В., 2015. Методы филогенетики и эволюционной биологии: достижения и ограничения // XIV Междунар. орнитол. конф. Сев. Евразии (Алматы, 18–24 августа 2015 г.). Т. 2. Доклады. Алматы. С. 138–165.
Ивушкин В.Е., 2015. Род Pyrrhula Brisson, 1760: состав, распространение и особенности экологии // Русский орнитологический журнал. Т. 24. № 1143. С. 1679–1738.
Калинин Е.Д., Опаев А.С., Соловьёва Е.Н., Головина М.В., Марова И.М., Редькин Я.А., 2018. Комплексный анализ изменчивости палеарктических форм черноголовых чеканов // Всеросс. орнитол. конгр. (г. Тверь, Россия, 29 января – 4 февраля 2018). Тезисы докладов. Тверь. С. 139–140.
Коблик Е.А., Архипов В.Ю., 2014. Фауна птиц стран Северной Евразии в границах бывшего СССР: списки видов // Зоологические исследования. № 14. М. 171 с.
Коблик Е.А., Архипов В.Ю., Волков С.В., Мосалов А.А., Редькин Я.А., 2017. Гималаи – “ключ” к пониманию таксономического разнообразия азиатских пеночек (Phylloscopidae, Aves) // Российские гималайские исследования: вчера, сегодня, завтра. СПб. С. 173–178.
Коблик Е.А., Редькин Я.А., Архипов В.Ю., 2006. Список птиц Российской Федерации. М. 256 с.
Крюков А.П., 2003. Современные концепции вида и роль российских биологов в их разработке // Проблемы эволюции. Т. V. С. 31–39.
Крюков А.П., Блинов В.Н., 1981. Взаимодействие серой и черной ворон (Corvus cornix L., С. corone L.) в зоне симпатрии и гибридизации: есть ли отбор против гибридов? // Журнал общей биологии. Т. 42. № 1. С. 128–135.
Лавренченко Л.А., 2013. Гибридогенное видообразование у млекопитающих: иллюзия или реальность? // Журнал общей биологии. Т. 74. № 4. С. 253–267.
Лукьянчук О.А., Марова И.М., Редькин Я.А., 2017. Географическая изменчивость морфологических и акустических признаков северных популяций ополовника Aegithalos caudatus (Passeriformes, Aegithalidae) // Зоологический журнал. Т. 96. № 4. С. 429–438.
Майр Э., 1968. Зоологический вид и эволюция. М.: Мир. 597 с.
Майр Э., 1971. Принципы зоологической систематики. М.: Мир. 454 с.
Майр Э., 1974. Популяции, виды, эволюция. М.: Мир. 464 с.
Марова И.М., Шипилина Д.А., 2015. Гибридные зоны у птиц: морфологические, биоакустические и генетические аспекты. // XIV Междунар. орнитол. конф. Северной Евразии (Алматы, 18–24 августа 2015 г.) Т. 2. Доклады. Алматы. С. 373–395.
Международный кодекс зоологической номенклатуры. 2004. Издание четвертое. М. 223 с.
Михайлов К.Е., 2003. Типологическое осмысление “биологического вида” и пути стабилизации околовидовой таксономии у птиц // Орнитология. Т. 30. С. 9–24.
Михайлов К.Е., 2015. Морфогенез и экогенез в эволюции птиц: их нетождественность и ее последствия // XIV Междунар. орнитол. конф. Северной Евразии (Алматы, 18−24 августа 2015 г.). Т. 2. Доклады. Алматы. С. 181–204.
Михайлов К.Е., 2017. Сложность дивергенции и структура вида у птиц // Труды Мензбировского орнитологического общества Вып. 3. М. С. 40–67.
Михайлов К.Е., 2018. Экологические расы и двойственность природы репродуктивной изоляции у птиц: может ли околовидовая морфо-гено-таксономия адекватно отобразить разноплановость дивергенции популяций у нейросложных организмов? // Русский орнитологический журнал. Т. 27 № 1695. С. 5581–5601.
Муравьев И.В., 1996. К вопросу о биологии группы желтых трясогузок в Пензенской области // Краевед. исслед. и проблема экол. образ. Пенза. С. 51–52.
Муравьев И.В., 1997. Сравнительная экология близкородственных видов на примере рода Motacilla L. Автореф. дис. … канд. биол. наук. М. 20 с.
Муравьев И.В., 1993. Гнездование группы “желтых” трясогузок в Пензенской области // Экология и охрана окружающей среды. Рязань. С. 87–88.
Опаев А.С., 2010. Основные направления дивергенции криптических видов птиц (на примере комплекса “дроздовидная камышевка”). Автореф. дис. … канд. биол. наук. М. 24 с.
Павлинов И.Я., 2005. Введение в современную филогенетику (кладогенетический аспект). М. 391 с.
Павлинов И.Я., 2017. Проблема вида в биологии: истоки и современность // Концепции вида у грибов: новый взгляд на старые проблемы (Материалы VIII всероссийской микологической школы-конференции). М.: Изд. МГУ. С. 5–19.
Павлинов И.Я., 2018. Основания биологической систематики: история и теория. М.: Товарищество научных изданий КМК. 786 с.
Павлинов И.Я., 2019. Звери России: справочник-определитель. (в 2-х томах). Определители по флоре и фауне России. Вып. 14. М.: Товарищество научных изданий КМК. 702 с.
Павлинов И.Я., Любарский Г.Ю., 2011. Биологическая систематика: эволюция идей. М. 667 с.
Панов Е.Н., 1989. Гибридизация и этологическая изоляция у птиц. М.: Наука. 512 с.
Панов Е.Н., 1993. Граница вида и гибридизация у птиц // Гибридизация и проблема вида у позвоночных. Сб. трудов Зоол. музея МГУ. Т. 30. С. 53–96.
Панов Е.Н., 2001. Межвидовая гибридизация у птиц: эволюция в действии // Природа. № 6. С. 51–59.
Панов Е.Н., 2008. Сорокопуты (семейство Laniidae) мировой фауны. Экология, поведение, эволюция. М.: Товарищество научных изданий КМК. 650 с. + 64 цв. вкл.
Политов Д.В., Мудрик Е.А., 2018. Молекулярно-генетические подходы к анализу внутривидовой структуры, микросистематики и микроэволюции птиц // Зоологические исследования. № 20. М. С. 115–118.
Портенко Л.А., 1960. Птицы СССР. Ч. 4. М.-Л.: Изд-во АН СССР. 417 с.
Портенко Л.А., 1981. Географическая изменчивость темнозобых дроздов (Turdus ruficollis Pallas) и ее таксономическая оценка // Филогения и систематика птиц. Труды ЗИН АН СССР, Т. 102. С. 72–109.
Потокина Е.К., Лебедева МВ., Ульянич П.С., Левкоев Э.А., Волков В.А., Жигунов А.В., 2017. Гетероплазмия и ядерные копии митохондриальных генов (NUMTs) в зоне интрогрессивной гибридизации ели европейской и ели сибирской // Генетика популяций: прогресс и перспективы. М. С. 223–225.
Пфандер П.В., 2018. Трагедия околовидовой систематики // Русский орнитологический журнал. Т. 27. № 1558. С. 301–335.
Редькин Я.А., 2001. Новые представления о таксономической структуре группы “желтых трясогузок” // Достижения и проблемы орнитологии Северной Евразии на рубеже веков. Казань. С. 150–165.
Редькин Я.А., 2001а. Таксономические отношения форм в эволюционно молодых комплексах птиц на примере рода Motacilla L., 1785 (Таксономическая ревизия подрода Budytes). Автореф. дис. … канд. биол. наук. М. 18 с.
Редькин Я.А., 2012. Семейство Трясогузковые. Полный определитель птиц европейской части России. Ч. 3. М.: Фитон ХХI. С. 38–75.
Редькин Я.А., 2015. Для чего нужно продолжать целенаправленный сбор коллекционных тушек птиц на территории России и сопредельных регионов в XXI веке // Тезисы IX Междунар. конф. хранителей орнитол. колл. “Орнитологические коллекции: из прошлого в будущее” (Москва, 12–16 октября 2015 г.). С. 80–81.
Редькин Я.А., Архипов В.Ю., Волков С.В., Мосалов А.А., Коблик Е.А., 2015. Вид или не вид? Спорные таксономические трактовки птиц Северной Евразии // XIV Международная орнитологическая конференция Северной Евразии (Алматы, 18–24 августа 2015 г.) Т. 2. Доклады. Алматы. С. 104–138.
Редькин Я.А., Коновалова М.В., 2003. К вопросу о репродуктивных отношениях двух форм зарнички Phylloscopus inornatus (Blyth, 1842) на основе анализа внешних морфологических признаков и деталей распространения // Русский орнитологический журнал. Т. 12. № 247. С. 1407–1430.
Рубцов А.С., 2015. Репродуктивная изоляция и понятие вида у птиц // Зоологический журнал. Т. 94. № 7. С. 816–831.
Рябицев В.К., 2014. Птицы Сибири: справочник определитель. Москва, Екатеринбург. Т. 2. 452 с.
Сотников В.Н., 2006. Птицы Кировской области и сопредельных территорий. Т. 2 (1) Воробьинообразные. Киров. 448 с.
Спиридонова Л.Н., Вальчук О.П., 2017. Новый случай рекомбинации между ядерным и митохондриальным геномами в роде Calliope: гипотеза происхождения Calliope pectoralis // Генетика популяций: прогресс и перспективы. М. С. 274–275.
Спиридонова Л.Н., Вальчук О.П., Редькин Я.А., 2019. Новый случай рекомбинации между ядерным и митохондриальным геномами в роде Calliope Gould, 1836 (Muscicapidae, Aves): гипотеза происхождения Calliope pectoralis Gould, 1837 // Генетика. Т. 55. № 1. С. 1–12.
Спиридонова Л.Н., Вальчук О.П., Редькин Я.А., Сайто Т., Крюков А.П., 2017. Филогеография и демографическая история соловья-красношейки Luscinia calliope // Генетика. Т. 49. № 6. С. 735–742.
Спиридонова Л.Н., Редькин Я.А., Вальчук О.П., Крюков А.П., 2016. Ядерные копии митохондриальных генов – источник новых вариантов гаплотипов гена цитохрома b Luscinia calliope (Muscicapidae, Aves) // Генетика. Т. 52. № 9. С. 1069–1080.
Степанян Л.С., 1978. Состав и распределение птиц фауны СССР. Воробьинообразные Passeriformes. М.: Наука. 392 с.
Степанян Л.С., 1983. Надвиды и виды-двойники в авифауне СССР. М.: Наука. 294 с.
Степанян Л.С., 2003. Конспект орнитологической фауны России и сопредельных территорий (в границах СССР как исторической области). М.: Наука. 808 с.
Тайкова С.Ю., Редькин Я.А., 2014. О формах группы серых сорокопутов в Европейской России и Украине в свете современных взглядов на систематику этого комплекса // Редкие виды птиц Нечерноземного центра России. Материалы V совещания “Распространение и экология редких видов птиц Нечерноземного центра России” (Москва, 6–7 декабря 2014 г.). М. С. 223–240.
Arnaiz-Villena A., Guillen J., Ruiz-del-Valle V., Lowy E., Zamora J., Varela P., Stefani D., Allende L.M., 2001. Phylogeography of crossbills, bullfinches, grosbeaks, and rosefinches // Cellular and Molecular Life Sciences. V. 58. P. 1–8.
Barani-Beiranvand H., Aliabadian M., Irestedt M., Qu Y., Darvish J., Szekely T., van Dijk R.E., Ericson P.G.P., 2017. Phylogeny of penduline tits inferred from mitochondrial and microsatellite genotyping // J. Avian Biology. V. 48. P. 932–940.
Barrera-Guzman A.O., Aleixo A., Shawkey M.D., Weir J.T., 2017. Hybrid speciation leads to novel male secondary sexual ornamentation of an Amazonian bird // Proc. Natl. Acad. Sci. USA. V. 115. P. e218-e225. https://doi.org/10.1073/pnas.1717319115
Becker J., 2007. Nachtigallen Luscinia megarhynchos, Sprosser L. luscinia und ihre Hybriden im Raum Frankfurt (Oder) – weitere Ergebnisse einer langjahrigen Beringungsstudie // Vogelwarte. V. 45. № 1. P. 15–26.
Benkman C.W., Parchman T.L., Mezquida E.T., 2010. Patterns of coevolution in the adaptive radiation of crossbills // Annals of the New York Academy of Sciences. V. 1206. P. 1–16.
Brelsford A., 2011. Hybrid speciation in birds: allopatry more important than ecology?// Molecular Ecolology. V. 20. P. 3705–3707. https://doi.org/10.1111/j.1365-294X.2011.05230.x
Brelsford A., Milá B., Irwin D.E., 2011. Hybrid origin of Audubon’s warbler // Molecular Ecology. V. 20. № 11. P. 2380–2389.
Clegg S.M., Degnan S.M., Moritz C., Estoup A., Kikkawa J., Owens I.P., 2002. Microevolution in island forms: the roles of drift and directional selection in morphological divergence of a passerine bird. // Evolution. V. 56. № 10. P. 2090–2099.
Clements J.F., Schulenberg T.S., Iliff M. J., Roberson D., Fredericks T.A., Sullivan B.L., Wood C.L., 2017. Clements checklist of birds of the world. http:// www.birds.cornell.edu/clementschecklist.
Coyne J.A., Orr H.A., 2004. Speciation. Sinauer Associates, Inc. 545 p.
del Hoyo J., Collar N.J., (Eds.), 2014. HBW and BirdLife International Illustrated Checklist of the Birds of the World. V. 1. Non-passerines. Lynx. 903 p.
del Hoyo J., Collar N.J., (Eds.), 2016. HBW and BirdLife International Illustrated Checklist of the Birds of the World. V. 2. Passerines. Lynx. 1013 p.
Dickinson E.C., Christidis L. (Eds.), 2014. The Howard & Moore Complete Checklist of the Birds of the World. 4th. Edition, V. 2. Eastbourne. 752 p.
Dickinson E.C., Remsen J.V. (Eds.), 2013. The Howard & Moore Complete Checklist of the Birds of the World. 4th. Edition, V. 1. Eastbourne. 461 p.
Edelaar P., Van Eerde K., Terpstra K., 2008. Is the nominate subspecies of the common crossbill Loxia c. curvirostra polytypic? II. Differentiation among vocal types in functional traits // J. Avian Biology. V. 39. P. 108–115.
Frankham R., 2008. Genetic adaptation to captivity in species conservation programs // Molecular Ecolology. V. 17. № 1. P. 325–333.
Garnett S.T., Christidis L., 2017. Taxonomy anarchy hampers conservation. The classification of complex organisms is in chaos // Nature. № 546. P. 25–27.
Haffer J., 1992. The history of species concepts and species limits in ornithology // In: Monk J.F. (ed.). Avian Systematics and Taxonomy. Bull. British Ornithologists Club. Centenary Supplement 112A. P. 107–158.
Haffer J., 1997. Species Concepts and Species Limits in Ornithology // In: del Hoyo J., Elliot A., Sargatal J. (Eds.) Handbook of the Birds of the World. V. 4. Sandgrouse to Cuckoos. Lynx. P. 11–24.
Hailer F., Kutschera V.E., Hallström B.M., Klassert D., Fain S.R., Leonard J.A., Arnason U., Janke A., 2012. Nuclear genomic sequences reveal that polar bears are an old and distinct bear lineage // Science. V. 336. № 6079. P. 344–357. https://doi.org/10.1126/science.1216424
Haring E., Gamauf A., Kryukov A., 2007. Phylogeographic patterns in widespread corvid birds // Molecular Phylogenetics and Evolution. V. 45. № 3. P. 840–862.
Harr B., Price T., 2012. Speciation: clash of the genomes // Current Biology. V. 22. № 24. R1044-R1046. https://doi.org/10.1016/j.cub.2012.11.005
Helbig A.J., Knox A.G., Parkin D.T., Sangster G., Collinson J.M., 2002. Guidelines for assigning species rank // Ibis. V. 144. № 3. P. 518–525.
Hubbard J.P., 1969. The relationships and evolution of Dendroica coronata complex // Auk. V. 86. P. 393–432.
Irwin D.E., Rubtsov A.S., Panov E.N., 2009. Mitochondrial introgression and replacement between yellowhammers (Emberiza citrinella) and pine buntings (Emberiza leucocephalos) (Aves: Passeriformes) // Biological Journal of the Linnean Society. V. 98. P. 422–438.
Johnson N.K., Remsen J.V., Cicero C., 1999. Resolution of the debate over species concepts in ornithology: a new comprehensive biologic species concept // In: Adams N.J., Slotow R.H. (Eds.) Proceedings of the 22nd International Ornithological Congress, Durban, South Africa, August 1998. Johannesburg. P. 1470–1482.
Kováts D., Végvári Z., Varga, Z., 2013. Morphological patterns of a nightingale population in a contact zone of Luscinia megarhynchos and L. luscinia // Acta Zoologica Academiae Scientiarum Hungaricae. V. 59. № 2. P. 157–170.
Kryukov A.P., Spiridonova L.N., Mori S., Arkhipov V.Yu., Red’kin Ya.A., Goroshko O.A., Lobkov E.G., Haring E., 2017. Deep phylogeographic breaks in Magpie Pica pica across the Holarctic: concordance with bioacoustics and phenotypes // Zoological Science. V. 34. № 3. P. 185–200. https://doi.org/10.2108/zs160119
Lamichhaney S., Han F., Webster M.T., Andersson L., Grant R.B., Grant P.R., 2017. Rapid hybrid speciation in Darwin’s finches // Science. https://doi.org/10.1126/science.aao4593
Le Gros A., Clergeau Ph., Zuccon D., Cornette R., Mathys B., Samadi S., 2016. Invasion history and demographic processes associated with rapid morphological changes in the Red-whiskered bulbul established on tropical islands // Molecular Ecology. V. 25. №21. P. 5359-5376.
Lewontin R.C., Birch L.C., 1966. Hibridization as a source of variation for adaptation to new environments // Evolution. V. 20. № 3. P. 315–336.
Liebers D., De Knijffa P., Helbig A.J., 2004. The herring gull complex is not a ring species // Proceedings of the Royal Society B: Biological Sciences. V. 271. P. 893–901.
Mallet J., 2006. Hybridization, ecological races and the nature of species: empirical evidence for the ease of speciation // Philosophical Transactions of The Royal Society B. Biological Sciences. V. 363. P. 2971–2986.
Mayr E., 1942. Systematics and the origin of species from the viewpoint of a zoologist. Columbia University Press. 334 p.
Mayr G., 2008. Avian higher-level phylogeny: well-supported clades and what we can learn from a phylogenetic analysis of 2954 morphological characters // Journal of Zoological Systematics and Evolutionary Research. V. 46. № 1. P. 63–72.
McCarthy E.M., 2006. Handbook of avian hybrids of the world. Oxford University Press. 583 p.
Milá B., Irwin D.E., 2011. A cryptic contact zone between divergent mitochondrial DNA lineages in southwestern North America supports past introgressive hybridization in the yellow-rumped warbler complex (Aves: Dendroica coronata) // Biological Journal of the Linnean Society. V.103. № 3. P. 696–706.
Miller W., Schuster S.C., Welch A.J., Ratan A., Bedoya-Reina O.C., Zhao F., Kim H.L., et al., 2012. Polar and brown bear genomes reveal ancient admixture and demographic footprints of past climate change // Proceedings of the National Academy of Sciences. V. 109. № 36. P. e2382–e2390.
Millien V., 2006. Morphological evolution is accelerated among island mammals // PLoS Biology. V. 4. № 10. e321. https://doi.org/10.1371/journal.pbio.0040321
Opaev A.S., Red’kin Ya.A., Kalinin E.D., Golovina M.V., 2018. Species limits in Northern Eurasian taxa of the common stonechats, Saxicola torquatus complex (Aves: Passeriformes, Muscicapidae) // Vertebrate Zoology. V. 68. № 3. P. 199–211.
Pagel M., Venditti Ch., Meade A., 2006. Large punctuational contribution of speciation to evolutionary divergence at the molecular level // Science. V. 314. P. 119–121.
Parchman T.L., Benkman C.W., Britch S.C., 2006. Patterns of genetic variation in the adaptive radiation of New World crossbills (Aves: Loxia) // Molecular Ecology. V. 15. № 7. P. 1873–1887.
Piertney S.W., Summers R.W., Marquiss M., 2001. Microsatellite and mitochondrial DNA homogeneity among phenotypically diverse crossbill taxa in the UK // Proceedings of the Royal Society B: Biological Sciences. V. 268. P. 1511–1517.
Price T. D., 2008. Speciation in birds. Greenwood Village, CO. 470 p.
Rasmussen P.C., Anderton J.C., 2005. Birds of South Asia. The Ripley Guide. V. 1–2. Washington D.C. & Barcelona: Smitsonian Institution & Lynx Edicions. 1067 p.
Red’kin Ya.A., Konovalova M.V., 2006. The eastern Asiatic races of Sitta europaea Linnaeus, 1758 // Systematic notes on Asian birds. 63. Zoologische Mededelingen Leiden. V. 80. № 15. P. 241–261.
Redkin Ya.A., Arkhipov V.Yu., Volkov S.V., Mosalov A.A., Koblik E.A., 2016. Art oder keine Art? Strittige taxonomishe Ansichten zu den Vögeln Nord-Eurasiens // Ornithologische Mittelungen. V. 68. № 11/12. P. 327–354.
Red’kin Ya.A., Konovalova M.V., 2004. On the taxonomy of Yellow-browed Warbler Phylloscopus inornatus sensu lato, based on the analysis of collection specimens (Aves: Sylviidae) // Zoosystematica Rossica. V. 13. № 1. P. 137–150.
Sangster G., 2009. Increasing numbers of bird species result from taxonomic progress, not тaxonomic inflation // Proceedings of the Royal Society B: Biological Sciences. V. 276. P. 3185–3191.
Sangster G., 2014. The application of species criteria in avian taxonomy and its implications for the debate over species concepts // Biological Reviews. V. 89. № 1. P. 199–214.
Sangster G., 2018. Integrative taxonomy of birds: the nature and delimitation of species // In: Bird Species. How They Arise, Modify and Vanish. Switzerland AG: Springer Nature. P. 9–37.
Song H., Buhay J.E., Whiting M.F., Crandall K.A., 2008. Many species in one: DNA barcoding overestimates the number of species when nuclear mitochondrial pseudogenes are coamplified // Proceedings of the National Academy of Sciences. V. 105. P. 13486–13491.
Soubrier J., Gower G., Chen K., et al., 2016. Early cave art and ancient DNA record the origin of European bison // Nature Communications. V.7. P. 1–6. https://doi.org/10.1038/ncomms13158
Summers R.W., Dawson R.J., Phillips R.E., 2007. Assortative mating and patterns of in-heritance indicate that the three crossbill taxa in Scotland are species // J. Avian Biology. V. 38. P. 153–162.
Summers R.W., Jardine D.C., Marquiss M., Rae R., 2002. The distribution and habitats of crossbills Loxia spp. in Britain, with special reference to the Scottish crossbill Loxia scotica // Ibis. V. 144. P. 393–410.
Thomson S.A., Pyle R.L., Ahyong S.T., Alonso-Zarazaga M., Ammirati J., Araya J.F., et al., 2018. Taxonomy based on science is necessary for global conservation. PLoS Biol. V. 16. № 3. e2005075. https://doi.org/10.1371/journal.pbio.2005075
Tobias J.A., Seldon N., Spottiswoode C.N., Pilgrim J.D., Fishpool L.D.C., Collar N.J., 2010. Quantitative criteria for species delimitation // Ibis. V. 152. № 4. P. 724–746.
Töpfer T., Haring E., Birkhead T.R., Lopes R.J., Severinghaus L.L., Martens J., Päckert M., 2011. A molecular phylogeny of bullfinches Pyrrhula Brisson, 1760 (Aves: Fringillidae) // Molecular Phylogenetics and Evolution. V. 58. P. 271–282.
Vaurie Ch., 1959. The birds of the Palearctic fauna. Order Passeriformes. L. 762 p.
Vijay N., Bossu Ch.M., Poelstra J.W., Weissensteiner M.H., Suh A., Kryukov A.P., Wolf J. B.W., 2016. Evolution of heterogeneous genome differentiation across multiple contact zones in a crow species complex // Nature Communications. V.7. P. 1–10. https://doi.org/10.1038/ncomms13195
Wiley E.O., Liberman B.S., 2011. Phylogenetics: the theory and practice of phylogenetic systematics. Hoboken. 406 p.
Wiley E.O., Mayden R.L., 1985. Species and speciation in phylogenetic systematics, with examples from the North American fish fauna // Annals of the Missouri Botanical Garden. V. 72. P. 596–635.
Zhang Z-Q., 2013. Animal biodiversity: An update of classification and diversity in 2013 // Zootaxa V. 3703. № 1. P. 5–11. https://doi.org/10.11646/zootaxa.3703.1.3
Zink R.M., 2004. The role of subspecies in obscuring avian biological diversity and misleading conservation policy // Proceedings of the Royal Society B: Biological Sciences. V. 271. P. 561–564.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал