

Зоологический журнал, 2019, T. 98, № 2, стр. 163-174
Морфо-экологическая характеристика преимагинальных стадий ксилобионтных видов рода Phaonia Robineau-Desvoidy 1830 (Muscidae, Diptera) – обитателей дупел деревьев и натеков сока
Н. П. Кривошеина *
Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Россия
* E-mail: dipteranina@rambler.ru
Поступила в редакцию 19.01.2018
После доработки 23.05.2018
Принята к публикации 23.05.2018
Аннотация
Изучена биология преимагинальных стадий двукрылых, сопутствующих обитающим в натеках сока и древесных дуплах ксилофагам. Впервые приведены сведения по биологии для 4 видов и дополнены для 2 видов рода Phaonia Robineau-Desvoidy 1830 (Muscidae). Установлены их основные диагностические признаки. Виды различаются по образу жизни личинок, строению их передних и задних дыхалец, ползательных валиков, ротоглоточного аппарата и анальной пластинки последнего сегмента тела. Разработана определительная таблица преимагинальных стадий ксилобионтных видов рода Phaonia.
Натеки сока на стволах деревьев – специфическая сапробиотическая среда обитания – сложное сочетание бродящего древесного сока, различных групп микроорганизмов и продуктов их жизнедеятельности. Такая среда богата органическими веществами и вследствие постоянного поступления новых порций сока сохраняет повышенную влажность в течение длительного времени. Последнее имеет особое значение для многих групп ксилобионтов, особенно двукрылых, в условиях аридного климата.
Наиболее обычны натеки на стволах лиственных пород: березы, тополя, вяза, ильма и дуба, в Южном Приморье характерны для ильма долинного (Ulmus japonica Rehder). В пустынных регионах натеки наблюдаются на стволах туранги (Populus diversifolia Schrenk) и на карагаче (Ulmus minor Mill.), особенно при повреждении стволов гусеницами древоточца Cossus cossus (L. 1758) (Кривошеина, Ягдыев, 1985), а также на участках со следами механического повреждения коры, вдоль трещин, сухобочин, или в местах выпадения сучков.
Комплекс обитателей натеков – лацидиколов – достаточно изучен в Европе и в России и включает в основном преимагинальные стадии насекомых, преимущественно двукрылых (Carter, 1945; Cole et al., 1970; Chandler, 1973; Кривошеина, 1975; Perry, Stubbs, 1978; Barták, Roháček, 2011; и др.). Характерными представителями таких биотопов оказываются личинки – сапро- и некрофаги семейств Anisopodidae, Mycetobiidae, Ceratopogonidae, Culicidae, Syrphidae и сопутствующие им личинки-хищники семейств Dolichopodidae, Odiniidae, Muscidae и др. (Кривошеина, 1973, 1974, 1975; Кривошеина Н., Кривошеина М., 2015).
В качестве биотопов для личинок двукрылых большое значение имеют также дупла, формирующиеся обычно в нижней части древесных стволов после выпадения или спила нижних ветвей, растрескиваний коры и т.п. В них накапливаются остатки гнилой древесины, насекомых и их экскрементов. Такие дупла в пустынных регионах характерны для тополей, туранги, грецкого ореха (Juglans regia L.) и платана пальчатолистного (Platanus digitata Gordon). Дупла, в зависимости от состояния и степени увлажненности, заселяются преимущественно жесткокрылыми и двукрылыми насекомыми (Кривошеина, 1975; Компанцева, 1984, 1993) и играют существенную роль в распространении нетипичных для территории видов. Так, изучение обитателей дупел на территории Приморского края позволило впервые обнаружить на территории России представителя рода Tipulodina Enderlein 1912, тяготеющего к тропикам и распространенного преимущественно в Ориентальной области (Кривошеина, 1972; Young, 1999). Личинки обитали в заполненном водой дупле чозении (Chosenia arbutifolia (Pall.)), на высоте 1.5 м (Кривошеина, 1972). В Туркменистане, в долинах Копетдага во влажной трухе дупел грецкого ореха, впервые для территории бывшего СССР, были обнаружены личинки львинок Eupachygaster tarsalis Zetterstedt (Stratiomyidae) (Кривошеина, 1975а).
Рассматриваемые среды обитания имеют наибольшее значение для двукрылых – обитателей субаридной и аридной зон. Сохранение высокой влажности субстрата в течение длительного времени, особенно в летний период, определяет его заселение личинками различных видов-ксилофагов и сопутствующих им хищников.
Сбор материала проводили из дупел на стволах грецкого ореха в предгорьях Копетдага (Туркменистан) и из натеков сока на стволах туранги на территории заповедника “Тигровая балка”(Таджикистан) в течение 1970−1987 гг.
Основное внимание уделялось личинкам-зоофагам рода Phaonia Robineau-Desvoidy 1830 (Muscidae) – постоянным спутникам различных комплексов ксилобионтов.
В процессе предварительного определения имагинального материала по роду Phaonia, был выявлен ряд новых видов, близких к Ph. boletina (Rondani 1866) Ph. rufipalpis (Macquart 1835) и Ph. trimaculata (Bouché 1834). По этим материалам Сорокиной (Sorokina, 2015) были описаны новые для науки виды: Ph. babarabica Sorokina 2015, Ph. juglans Sorokina 2015, Ph. modesta Sorokina 2015 и Ph. ninae Sorokina 2015.
В представленной работе приведены описания их преимагинальных стадий и обсуждаются отличия от близких видов.
МОРФО-ЭКОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА ЛИЧИНОК-ЗООФАГОВ РОДА PHAONIA ROBINEAU-DESVOIDY 1830
Для личинок Phaonia характерен широкий диапазон экологических связей с различными древесными породами. Прослеживаются три основные экологические группы: обитатели зоны коры, толщи древесины и субстратов, пропитанных древесным соком (Кривошеина, 2012а). Такой тип питания как хищничество, в значительной степени объясняет эвритопность большинства видов. Личинки зарегистрированы в натеках сока на коре как хвойных, так и лиственных пород. При этом ряд видов предпочитает стволы, пораженные жуками-сверлилами (Lymexylonidae). Личинки обычно встречаются в заполненных соком ходах жуков и в натеках сока в ходах гусениц древоточца Cossus cossus L.
Phaonia babarabica Sorokina 2015 (рис. 1, 1−7; 2, 1−8).
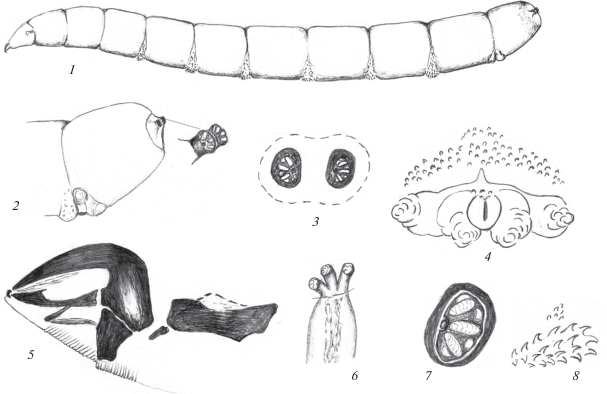
Рис. 1.
Личинка Phaonia babarabica Sorokina 2015: 1 – общий вид, 2 – конец тела сбоку, 3 – анальное поле, 4 – переднее дыхальце, 5 – кутикулярные структуры ползательных валиков, 6 – задние дыхальца, 7 – ротоглоточный аппарат: a – околоанальный бугорок, b – экстраанальный бугорок, das – дорсальный акцессорный склерит, dp – дорсальный отросток фарингеального склерита, ds – зубной склерит, mds – мандибулярные крючья, ors – ротовой акцессорный склерит, phs – фарингеальный склерит, vas – вентральный акцессорный склерит, vp – вентральный отросток фарингеального склерита.
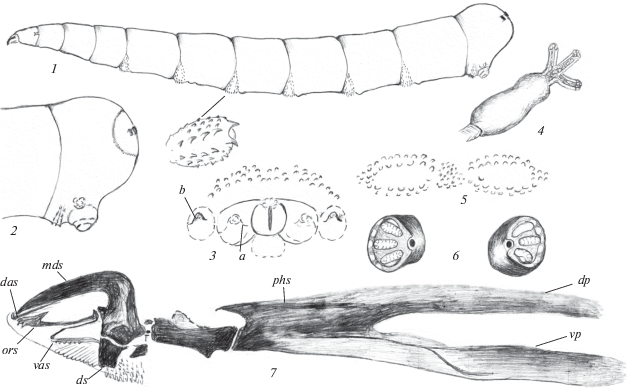
Материал. 1 личинка, 2 пупария, 1♂, 1♀, Туркменистан, Бабараб, 29.04.1982, № 112, в стволе карагача, в ходах Cossus cossus; там же, 27.04.1982, № 102, под корой ивы (Н. Кривошеина).
Описание. Личинка. Тело светло-желтое, последний сегмент закруглен, гладкий (рис. 1, 1, 2).
Ротоглоточный аппарат узкий, удлиненный. Парные дорсальные акцессорные склериты узкие, стержневидные. К ним спереди примыкает небольшой дуговидный склерит. Ротовые акцессорные склериты на вершине с дорсальным заостренным выступом и четкими конусовидными зубцами с вентральной стороны (рис. 1, 7). Вентральные акцессорные склериты очень узкие, соединены светлыми тяжами с зубными склеритами. Фарингеальный склерит удлиненный, с параллельными, почти равной длины и ширины дорсальными и вентральными отростками. Длина его вентральных отростков в 2 раза превышает длину базального отдела фарингеального склерита (7 : 15). Вентральные отростки с четкой продольной темной складкой. Зубные склериты в нижнем отделе почти прямоугольные. Гипостомальные склериты с закругленным вершинным выступом сзади и четким конусовидным выступом с вентральной стороны. Длина мандибулярных крючьев относится к длине гипостомальных и фарингеального склеритов как 3 : 2 : 12, т.е. мандибулярные крючья совместно с гипостомальными склеритами почти в 2.5 раза короче фарингеального склерита.
Длина грудных сегментов равна их ширине или превышает ее. Первый брюшной сегмент короткий, его ширина меньше длины. Длина II–VI брюшных сегментов равна их ширине или несколько меньше ее, VII брюшной сегмент удлиненный, VIII – короткий, закругленный.
Ползательные валики с дуговидно изогнутыми поперечными рядами конусовидных светлых шипиков (рис. 1, 5). Наиболее крупные образуют 2 сближенных передних ряда и на некотором расстоянии от них – 2 задних ряда. В центре между ними расположена группа мелких шипиков.
Анальная пластинка небольшая, овальная. Анальное поле без четких границ, с 2 парами бугорков: 2 околоанальными и 2 экстраанальными (рис. 1, 3). Перед анальной пластинкой расположены 2–3 ряда светлых шипиков, ряды которых доходят до экстраанальных бугорков.
Передние дыхальца с 3 удлиненными выростами, несущими дыхальцевые камеры (рис. 1, 4). Атриум удлиненный, массивный. Задние дыхальца расположены на коротких, слегка выступающих цилиндрических бугорках (рис. 1, 6). Дыхальцевая пластинка овальная, с 3 светлыми радиально расходящимися изолированными дыхальцевыми камерами. Перитрема широкая. Дыхальцевые бугорки расположены на расстоянии, в 1.5 раза превышающем их диаметр.
Длина тела 11 мм.
Пупарий. Окраска пупария светло-коричневая. Задние дыхальца относительно широко расставлены.
Ротоглоточный аппарат удлиненный (рис. 2, 1, 2). Парные ротовые акцессорные склериты узкие, с небольшим дорсальным выступом в передней трети, со слабо выраженными зубцами, парные вентральные акцессорные склериты короткие, расположены под углом к ротовым склеритам. Фарингеальный склерит удлиненный, с параллельными почти равной ширины и длины узкими дорсальными и вентральными отростками и узкой прозрачной фарингеальной мембраной. Его вентральные отростки со слабо заметным дорсальным выступом. Длина отростков в 2 раза превышает длину базального отдела фарингеального склерита. Совместная длина мандибулярных крючьев и гипостомальных склеритов почти в 2.5 раза меньше длины фагингеального склерита.
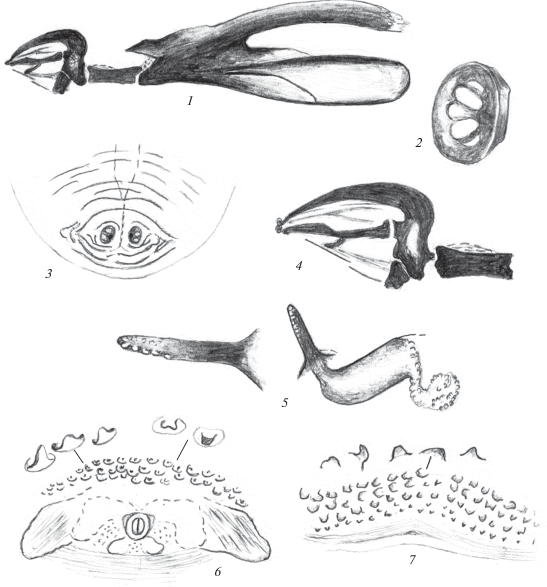
Рис. 2.
Пупарий Phaonia babarabica Sorokina 2015: 1, 2 – ротоглоточный аппарат; 3 – переднее дыхальце; 4 – анальное поле; 5 – кутикулярные структуры ползательных валиков; 6 – последний сегмент с дорсальной стороны; 7, 8 – задние дыхальца.
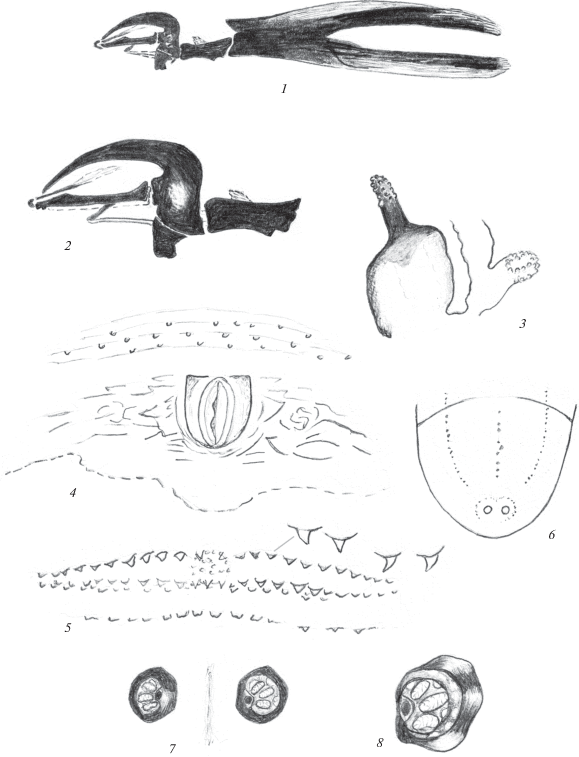
Ползательные валики с четко выраженными 2 рядами прозрачных конусовидных выступов и относительно более слабо выраженным третьим рядом кутикулярных структур (рис. 2, 5). В среднем отделе валика расположено небольшое морщинистое пятно, окруженное мелкими бугорками.
Последний сегмент с дорсальной стороны с 3 продольными бороздками, несущими единичные темные ямки (рис. 2, 6). Анальная пластинка небольшая, овальная или с небольшими боковыми выступами спереди, окружена короткими продольными и поперечными складками. Анальное поле более светлое по сравнению с окружающими покровами (рис. 2, 4). Характерное для личинок Phaonia поле шипиков, расположенное перед анальной пластинкой, слабо развито. Имеется лишь несколько поперечных складок, несущих единичные тупые бугорки.
Передние дыхальца удлиненные, с прозрачными округлыми дыхальцевыми камерами в их вершинной части (рис. 2, 3). Задние дыхальца расположены на расширенных в основании темных бугорках (рис. 2, 7, 8). Расстояние между бугорками в 1.5 раза превосходит их диаметр. Дыхальцевая пластинка задних дыхалец овальная, черная, расположена под углом к поверхности тела. Перитрема дыхалец широкая, черная, Дыхальцевые щели длинные, светлые, слегка расходятся на вершине. Между ними расположены светлые овальные площадки с периспиракулярными железами.
Длина пупария 9 мм.
Биология. Личинки обитали вместе с личинками Mycetobia turkmenica Mamaev 1987 (Mycetobiidae) в натеках сока на ивах, произрастающих вблизи арыков, и во влажной трухе в ходах Cossus cossus на карагаче, где в массе развивались личинки Scatopsidae и Drosophilidae.
Phaonia juglans Sorokina 2015 (рис. 3, 1−6).
Рис. 3.
Пупарий Phaonia juglans Sorokina 2015: 1, 2 – ротоглоточный аппарат; 3 – переднее дыхальце; 4 – стигмальное поле; 5 – анальное поле; 6 – кутикулярные структуры на ползательных валиках VII брюшного сегмента.
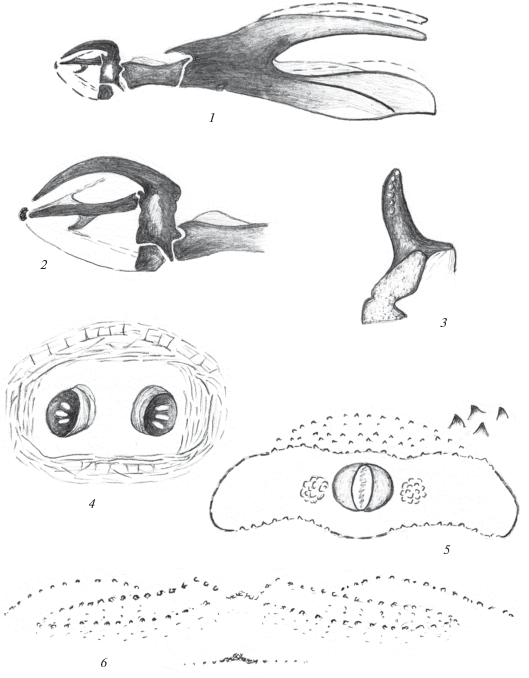
Материал. 1 пупарий, 1♂, Туркменистан, Копетдаг, Ипай-Кала, 6.06.1971, № 237, дупло грецкого ореха. Вылет имаго 18.06.1971 (Н. Кривошеина).
Описание. Пупарий. Парные дорсальные акцессорные склериты в виде тонких тяжей. Ротовые акцессорные склериты закруглены на вершине, без зубцов. Перед ними – небольшой дуговидный поперечный склерит (рис. 3, 1, 2). Парные вентральные акцессорные склериты короткие, соединены тяжами с зубными склеритами. Длина мандибулярных крючьев равна длине гипостомального склерита. Их совместная длина в 2 раза меньше длины фарингеального склерита. Дорсальные отростки фарингеального склерита заметно ýже вентральных. Последние массивные, с удлиненным выступом по дорсальному краю. Базальная часть фарингеального склерита почти в 2 раза короче вентральных отростков.
Ползательные валики с тремя рядами симметрично расположенных бугорков, почти сходящихся в центре (рис. 3, 6).
Передние дыхальца расположены на первом брюшном сегменте, вблизи границы с грудными сегментами. Дыхальца удлиненные, слегка дуговидно загнуты, темные, на боковой стороне с округлыми светлыми камерами (рис. 3, 3). Задние дыхальца расположены на небольшом, поперечном стигмальном поле, ограниченном узкими поперечными и продольными складками (рис. 3, 4). Дыхальца заметно выступают над поверхностью сегмента, расположены на небольших темных склеротизованных выступах, расстояние между которыми не превышает их диаметр. Стигмальная пластинка с тремя удлиненными, слегка расходящимися дыхальцевыми щелями.
Поверхность последнего сегмента с дорсальной стороны гладкая, без заметных продольных борозд. Анальная пластинка округлая, анальное поле поперечное, по границе с редкими тупыми бугорками (рис. 3, 5). Перед анальным полем разрозненные бугорки образуют 4–5 поперечных рядов.
Размеры пупария 7 мм.
Примечание. Пупарий, в отличие от пупария Ph. cincta, описанного из того же биотопа, имеет следующие признаки: анальная пластинка округлая, не трапециевидная, последний сегмент без продольных дорсальных бороздок, задние дыхальца расположены друг от друга лишь на расстоянии, практически не превышающем их диаметр. У Ph. cincta расстояние между дыхальцами почти в 1.5 раза превышает их диаметр.
Phaonia modesta Sorokina 2015 (рис. 4, 1−6).
Рис. 4.
Пупарий Phaonia modesta Sorokina 2015: 1 – последний сегмент с вентральной стороны; 2 – конец тела с дорсальной стороны; 3 – кутикулярные структуры на ползательных валиках VII брюшного сегмента; 4 – стигмальное поле сзади; 5, 6 – передние дыхальца.
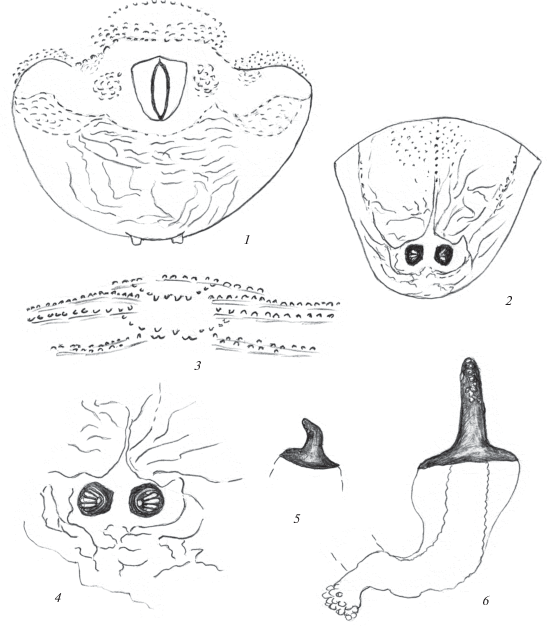
Материал. 1 пупарий, 1 ♂, Туркменистан, Геок-Тепе, Копетдаг, ущелье Чули, 13.06.1984. Пупарий в плодовом теле гриба, вылет имаго 10.06.1984 (Н. Кривошеина).
Описание. Пупарий. Покровы светло-коричневые, с более светлыми участками в области анального поля и вокруг дыхалец. Сегменты тела гладкие, с затемненными поперечными полосками, несущими микробугорки в области ползательных валиков (рис. 4, 3). Анальное поле поперечное, с удлиненными боковыми выступами. Анальная пластинка с продольной анальной щелью, расширена и обрублена спереди, но сужена и закруглена сзади. Сбоку от анальной пластинки – небольшие бугорки с несколькими тупыми шипиками. Перед анальной пластинкой 3–4 поперечных ряда шипиков. За анальной пластинкой покровы на всем протяжении до задних дыхалец морщинистые, с поперечными и продольными складками. Расстояние между бугорками, несущими задние дыхальца, меньше их диаметра (рис. 4, 4). Расстояние между задними дыхальцами (вид снизу) в 2.5 раза превышает диаметр дыхалец (рис. 4, 1). Последний сегмент с дорсальной стороны с двумя продольными сближенными срединными и двумя боковыми бороздками (рис. 4, 2).
Ползательные валики с симметрично расположенными тремя–четырьмя поперечными рядами мелких, слабо развитых бугорков, соединенных короткими рядами таких же структур в центре (рис. 4, 3).
Передние дыхальца расположены на боковой стороне первого брюшного сегмента, в виде удлиненных, конусовидных бугорков. Их верхняя часть с многочисленными округлыми светлыми камерами (рис. 4, 5, 6). Стигмальные пластинки задних дыхалец расположены под углом к поверхности сегмента. Дыхальца небольшие, значительно меньше несущих их бугорков. Стигмальное поле, участок покровов вокруг задних дыхалец, гладкое, в виде поперечной, почти прямоугольной пластинки, окруженной узкими бороздками (рис. 4, 4).
Размеры пупария 9 мм.
Биология. Личинки Ph. modesta зарегистрированы в плодовых телах ксилотрофных грибов. Близкий Ph. modesta вид Ph. trimaculata (Bouché 1834) развивается преимущественно в подстилке и даже в овощах, отмечен в древесине (Keilin, 1917; Ciampolini, 1960; Кривошеина, 2013).
Phaonia ninae Sorokina 2015 (рис. 5, 1−8; 6, 1−7).
Рис. 5.
Личинка Phaonia ninae Sorokina 2015: 1 – общий вид сбоку; 2 – конец тела сбоку; 3, 7 – задние дыхальца; 4 – анальное поле; 5 – ротовой аппарат и гипостомальный склерит; 6 – переднее дыхальце; 8 – кутикулярные структуры на ползательных валиках VII брюшного сегмента.
Материал. 1 личинка, 2 пупария, 1♂, 7♀♀, Таджикистан, заповедник “Тигровая балка”, Северный кордон (окрестности Джиликуля), 27.03.1986, 11.05.1987 и 15.06.1987, № 13, 17, 20, в натеках сока на туранге, Populus diversifolia Schrenk. Вылет имаго 16.04.1986 и 1.06.1987 (Н. Кривошеина).
Описание. Личинка. Тело светло-желтое, с заостренным головным и закругленным последним сегментом. Задние дыхальца расположены на небольших бугорках (рис. 5, 1, 2).
Ротоглоточный аппарат с хорошо развитыми удлиненными ротовыми акцессорными склеритами, несущими дорсальный выступ в передней трети. Дорсальные акцессорные склериты в виде слабо выраженных перепончатых тяжей. Перед ними расположен короткий дуговидный склерит. Короткие, направленные вперед вентральные акцессорные склериты соединены слабо выраженными перепончатыми тяжами с зубными склеритами (рис. 5, 5). Отношение частей ротоглоточного аппарата (мандибулярных крючьев, гипостомального и фарингеального склеритов) 3 : 2 : : 8.0–8.4. Отношение длины базальной части фарингеального склерита к длине его отростков 3 : 5. Зубные склериты неправильной формы (рис. 5, 5).
Длина II–III грудных и I брюшного сегментов несколько уступает ширине, длина остальных почти равна ширине. Наиболее длинный VII брюшной сегмент.
Ползательные валики в виде парных поперечно-овальных бугорков, разобщенных вдоль средней линии сегментов. На них расположены светлые крючковидные шипики (рис. 5, 8).
Анальная пластинка овальная, светлая, сбоку от нее расположены 2 пары бугорков: 2 субанальных и 2 экстраанальных. Перед ней – 4–5 поперечных рядов тупых светлых шипиков, расположенных в шахматном порядке. Ряды шипиков доходят до уровня экстраанальных бугорков (рис. 5, 4).
Передние дыхальца светлые, с тремя дыхальцевыми камерами, атриум относительно массивный (рис. 5, 6). Задние дыхальца расположены на коротких, светлых в основании и затемненных на вершине бугорках (рис. 5, 2, 3, 7). Длина затемненного участка бугорка в 2 раза меньше его ширины. Стигмальная пластинка задних дыхалец почковидная, с тремя удлиненными, радиально расходящимися светлыми дыхальцевыми камерами. Между ними хорошо заметны светлые участки с периспиракулярными железами. Дыхальцевые бугорки сближены, расстояние между ними не больше их диаметра.
Длина тела 9 мм.
Пупарий. Покровы светло-желтые, ползательные валики с рядами светлых бугорков. Задние дыхальца расположены на коротких темных выступах. На уровне передних дыхалец пупарий слегка сужен.
Ротоглоточный аппарат массивный, по строению близок к таковому Ph. cincta. Мандибулярные крючья с внутренним выступом, к которому примыкают базальные отделы ротовых акцессорных склеритов (рис. 6, 1, 4). Ротовые акцессорные склериты расширены в переднем отделе, без заметных зубцов и выступов. Парные дорсальные акцессорные склериты слабо склеротизованы, тонкие и почти примыкают к ротовым акцессорным склеритам. Парные вентральные акцессорные склериты темные и короткие. Фарингеальный склерит массивный, его дорсальные отростки тоньше вентральных, дуговидно изогнуты. Вентральные отростки с плоским дорсальным выступом в основании, неравномерно склеротизованы. Длина базальной части фарингеального склерита относится к длине его отростков как 2 : 3. Длина мандибулярных крючьев превосходит длину гипостомального склерита (7 : 6). Совместная длина этих структур в 2 раза меньше длины фарингеального склерита. Зубной склерит неправильной формы (рис. 6, 4).
Рис. 6.
Пупарий Phaonia ninae Sorokina 2015: 1, 4 – ротоглоточный аппарат; 2 – задние дыхальца; 3 – конец тела с задними дыхальцами; 5 – передние дыхальца; 6 – анальное поле; 7 – кутикулярные структуры ползательных валиков.
Ползательные валики, расположенные в переднем отделе брюшных сегментов, с поперечными рядами слабо склеротизованных неправильной формы бугорков (рис. 6, 7). Среди них обычно более заметен 1 ряд, образованный более крупными и слегка затемненными бугорками. В целом можно говорить о 4–5 рядах. За их рядами покровы пупария с тонкими поперечными параллельными складками.
Последний сегмент тела с дорсальной стороны не морщинистый, перед дыхальцами с несколькими поперечными складками (рис. 6, 3).
Анальная пластинка небольшая, светлая, почти овальная или со слабо выраженными закругленными угловидными выступами в переднем отделе. Анальное поле со слабо намеченными границами (рис. 6, 6). Небольшие морщинистые участки расположены на уровне околоанальных бугорков. Далее вблизи боковых краев анального поля расположены ряды косых параллельных складок. Перед анальной пластинкой расположены ряды редких тупых светлых бугорков, ближе к боковой стороне преимущественно плоских и неправильной формы. Бугорки образуют по 3–4 симметричных поперечных ряда, которые не доходят до боковых краев анального поля.
Передние дыхальца конусовидные, удлиненные, слегка изогнуты в вершинной трети, где расположены овальные, светлые дыхальцевые камеры (рис. 6, 5). Длина дыхалец в 4.5–5 раз превышает ширину в основании. Атриум с характерным округлым придатком. Задние дыхальца овальные, с широкой темной перитремой и тремя светлыми удлиненными дыхальцевыми камерами (рис. 6, 2). Стигмальная пластинка небольшая, овальная. Бугорки, несущие дыхальца, расположены на расстоянии, практически равном их диаметру, спереди и сзади к ним примыкают несколько тонких поперечных складок (рис. 6, 3).
Длина пупария 7–8 мм.
Биология. Личинки и пупарии были собраны из натеков на стволах туранги. Натеки формировались вдоль трещин коры, а также в результате повреждения стволов сартским дровосеком Aeolesthes sarta (Solsky 1871) (Cerambycidae). Личинки встречались в скоплениях темной трухи совместно с личинками мокрецов Dasyhelea sp., Culicoides sp. и сирфид. Вылет имаго наблюдался в середине апреля 1986 г. и в первой половине июня 1987 г.
Примечание. Вид внешне сходен с Ph. rufipalpis (Macquart 1835).
В литературе (Skidmore, 1985) приводятся рисунки пупария самки Ph. rufipalpis из Великобритании. В строении пупария наблюдаются четкие отличия от Ph. ninae. У Ph. rufipalpis фарингеальные отростки узкие, при этом вентральные без выступа по дорсальному краю. Задние дыхальца расположены на расстоянии, в 3 раза превышающем их диаметр, перед анальной пластинкой ряды бугорков выражены слабо. У Ph. ninae вентральные отростки заметно шире дорсальных, с четким выступом по дорсальному краю. Задние дыхальца расположены на расстоянии, не более чем в 1.5 раза превышающем их диаметр, перед анальной пластинкой несколько поперечных рядов бугорков. Соотношение размеров частей ротоглоточного аппарата также иное.
В рассматриваемый комплекс входят также личинки Phaonia exoleta (Meigen 1826) и Ph. cincta (Zetterstedt 1876), описанные ранее в литературе (Keilin, 1917; Skidmore, 1985; Кривошеина, 2012).
Phaonia exoleta (Meigen 1826). Вид широко распространен в Северной и Центральной Европе, известен из Таджикистана и Туркменистана. Личинки Ph. exoleta развиваются в дуплах деревьев, отчасти заполненных водой. На территории Европы личинки обычны в дуплах клена белого (Acer pseudoplatanus L.), где регистрируются, как правило, совместно с личинками Brachyopa sp. (Syrphidae) (Keilin, 1917; Skidmore, 1985).
В Туркменистане, в долинах Копетдага, в заполненных водой дуплах грецкого ореха личинки обитают совместно с личинками мокрецов Culicoides arboreus Gutsevich 1952 (Ceratopogonidae), кровососущих комаров Aedes caspius (Pallas 1771) и A. pulchritarsis asiaticus (Edwards 1926) (Culicidae).
Cведения о личинке Ph. exoleta опубликованы несколькими авторами (Keilin, 1917; Tate, 1935; Skidmore, 1985; Кривошеина, 2012). По литературным данным, только личинка этого вида обладает хорошо развитыми парными ложными ножками на брюшных сегментах. Между тем, наблюдаются некоторые различия в строении преимагинальных стадий из разных регионов Палеарктики. Так, анальная пластинка пупария поперечно-овальная, правильной формы (Ирландия и Англии – Skidmore, 1985; горы Копетдага, Туркменистан – Кривошеина, 2012 ) или боковые стороны пластинки сужены и неправильной формы (Англия – Keilin, 1917; Таджикистан – Кривошеина, 2012). Ложные ножки личинок с удлиненными выступами, несущими на вершине по несколько шипов (Кривошеина, 2012; Тате, 1935), или выступы ложных ножек с одним простым шипом (Keilin, 1917). Эти данные позволяют предположить, что в действительности существует не один вид со столь своеобразными личинками.
Phaonia cincta (Zetterstedt 1876). Вид широко распространен в Европе от Швеции до Италии, известен из Турции и Сирии. Личинки зарегистрированы в Европе как обитатели натеков сока на стволах деревьев широколиственных пород, преимущественно Ulmus L. и каштана конского Aesculus L. (Skidmore, 1985). На территории Туркменистана личинки собраны из дупел грецкого ореха (Кривошеина, 2012).
Личинки – неспециализированные облигатные хищники, питаются личинками двукрылых: мокрецов (Ceratopogonidae), мицетобиид (Mycetobiidae), сирфид (Syrphidae), лонхеид (Lonchaeidae) и др. (Skidmore, 1985; Smith, 1989), а также личинками и имаго жесткокрылых: ризофагид (Rhizophagidae), стафилинид (Staphilinidae), различных видов блестянок рода Epuraea Erichson, 1843 (Nitidulidae) (Keilin, 1917; Skidmore, 1985).
Описанная нами личинка Ph. cincta из Туркменистана (Кривошеина, 2012) по большинству признаков сходна с личинкой Ph. cincta, описанной по материалам из Европы, но наблюдаются некоторые отклонения. Так, у личинок из дупел ореха (Копетдаг – Кривошеина, 2012) и натеков сока (Keilin, 1917) ротовые акцессорные склериты с зубцами у вершины. У личинок из конского каштана (Skidmore, 1985) ротовые склериты на вершине заужены и без зубцов. Анальная пластинка пупария из дупла грецкого ореха (Кривошеина, 2012) трапециевидная, а из конского каштана (Skidmore, 1985) поперечная, овальная. Расстояние между задними дыхальцами, по разным данным, варьирует: равно диаметру дыхальца (Keilin, 1917), в 1.5 раза (Кривошеина, 2012) и более чем в 2 раза (Skidmore, 1985) превосходит диаметр дыхалец. Наблюдаемые различия говорят или о возможном существовании не одного, а двух близких видов, или о варьировании морфологических структур личинок в пределах P. cincta.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПРЕИМАГИНАЛЬНЫХ СТАДИЙ ИССЛЕДОВАННЫХ ВИДОВ PHAONIA
1. Тело с конусовидно суженным задним концом. Брюшные сегменты с хорошо развитыми парными ложными ножками, на вершине которых расположены дуговидные черные шипики. Фарингеальный склерит ротоглоточного аппарата с расширяющимися к вершине отростками. Личинки обитают в трухе заполненных водой дупел лиственных древесных пород …………Ph. exoleta (Meig.)
– Тело с закругленным задним концом. В переднем отделе брюшных сегментов расположены слегка выступающие ползательные валики, несущие многочисленные конусовидные светлые шипики. Отростки фарингеального склерита равной или разной ширины, но параллельносторонние, не расширены на вершине………………………………2
2. Переднегрудные дыхальца личинки светлые, с двумя слабо заметными дыхальцевыми камерами. Парные поля шипиков на вентральной стороне брюшных сегментов сближены, соединены узким участком из более мелких шипиков. Отростки фарингеального склерита разные по ширине, дорсальные более узкие. Личинки обитают во влажной древесной трухе в дуплах, а также в натеках сока на стволах …………….Ph. cincta (Zett.)
– Переднегрудные дыхальца с тремя–четырьмя дыхальцевыми камерами.……..........................3
3. Фарингеальные склериты с узкими дорсальными и вентральными отростками ………………….4
– Фарингеальные склериты с отростками разной ширины, вентральные более массивные и широкие. Расстояние между бугорками, несущими задние дыхальца, равно их диаметру…………..6
4. Задние дыхальца расположены друг от друга на расстоянии, почти равном их диаметру ………5
– Задние дыхальца расположены на расстоянии, в 2 раза превышающем их диаметр, дыхальцевые бугорки − на расстоянии, в 1.5 раза превышающем их диаметр. Личинки под корой, в натеках сока на стволах ………….Ph. babarabica Sorokina
5. Дорсальная поверхность последнего сегмента гладкая, с четкими тремя продольными бороздками. Поля шипиков не сужены в среднем отделе, где шипики не мельче остальных. Анальное поле практически гладкое, лишь с темными бугорками вблизи боковых сторон анальной пластинки. Личинки под корой лиственных пород деревьев ………………..........Ph. trimaculata (Bouché)
– Дорсальная поверхность последнего сегмента морщинистая, в многочисленных складках (рис. 4, 2). Продольные борозды заметны лишь у границы последнего сегмента. Пупарий обнаружен в трутовых грибах...………Ph. modesta Sorokina
6. Стигмальное поле пупария гладкое, поперечное, с четко обозначенными границами. Анальная пластинка округлая (рис. 3, 5). Направленный назад вырост ротовых акцессорных склеритов хорошо выражен, длинный. Длина передних дыхалец пупария в 3 раза превосходит их ширину. Личинки в дупле грецкого ореха.…….Ph. juglans Sorokina
– Стигмальное поле пупария с многочисленными складками, без четких границ. Анальная пластинка пупария расширена в переднем отделе (рис. 6, 6). Направленный назад вырост ротовых акцессорных склеритов очень короткий или отсутствует (у пупария). Длина передних дыхалец пупария в 4–5 раз превосходит их ширину (рис. 6, 5). Личинки в натеках сока на туранге..……Ph. ninae Sorokina
Список литературы
Компанцева Т.В., 1984. Особенности формирования комплексов жесткокрылых в дуплах деревьев в южной подзоне тайги. Соколов В. Е. (ред.). Животный мир южной тайги. М.: Наука. С. 211–219.
Компанцева Т.В., 1993. Сообщества насекомых – обитателей дупел деревьев в лесах долины р. Вахш. Энтомокомплексы долинных лесов пустынной зоны. М.: Наука. С. 71–80.
Кривошеина Н.П., 1972. К биологии видов рода Tipulodina Alex. (Diptera, Tipulidae), нового для фауны СССР // Доклады Академии наук СССР. Т. 202. № 1. С. 240–242.
Кривошеина Н.П., 1973. Личинки рода Systenus (Diptera, Dolichopodidae) // Зоологический журнал. Т. 52. № 7. С. 1095–1097.
Кривошеина Н. П., 1974. Морфология личинок мух-зеленушек рода Medetera Fisch. (Diptera, Dolichopodidae) // Энтомологическое обозрение. Т. 53. Вып. 2. С. 309–323.
Кривошеина Н.П., 1975. Формирование комплексов двукрылых насекомых в сапробиотических средах – натеках сока на стволах деревьев // Энтомологические исследования на Дальнем Востоке. Вып. 3. Владивосток. С. 70–75.
Кривошеина Н.П., 1975а. Новые данные о ксилофильных двукрылых насекомых (Stratiomyidae, Pachygasterinae), развивающихся в тугайной растительности Средней Азии // Научные доклады высшей школы. Биологические науки. № 10. С. 11–17.
Кривошеина Н.П., 2012. Морфоэкологическая характеристика ксилобионтных личинок рода Phaonia Robineau-Desvoidy, 1830 (Diptera, Muscidae) с описанием личинки Ph. canescens Stein, 1919 // Энтомологическое обозрение. Т. 91. Вып. 3. С. 658–668.
Кривошеина Н.П., 2012а. К экологии личинок рода Phaonia (Diptera, Muscidae) // Зоологический журнал. Т. 91. № 12. С. 1489–1497.
Кривошеина Н. П., 2013. Новые данные по экологии и морфологии преимагинальных стадий Phaonia trimaculata (Bouché, 1834) (Diptera: Muscidae) // Кавказский энтомологический бюллетень. Т. 9 (1). С. 197–200.
Кривошеина Н.П., Ягдыев А. 1985. Ивовый древоточец (Cossus cossus L.) – вредитель плодовых в Туркменистане // Известия Академии наук Туркменской ССР, серия биологических наук. № 6. С. 73–74.
Кривошеина Н. П., Кривошеина М. Г., 2015. Определитель двукрылых насекомых подотряда Brachycera-Orthorrhapha по личинкам. Москва: Товарищество научных изданий КМК. 384 с.
Barták M., Roháček J., 2011. Records of interesting flies (Diptera) attracted to meat baited pyramidal trap on sapping stump of Europen walnut (Juglans regia) in Central Bohemia (Czech Republic) // Časopis Slezského Musea v Opava (A). V. 60. P. 223–233.
Chandler P.J., 1973. Some Diptera and other insects associated with decaying elm (Ulmus procera Salisbury) am Bromley, Kent, with some additional observations on these and related species // Entomologist’s Gasette. V. 24. P. 329–346.
Carter J.C., 1945. Westwood of elm // Bulletin of the Illinois State Natural History. Survey division. V. 23 (4). P. 401–448.
Ciampolini M., 1960. La Phaonia trimaculata Bouché (Diptera, Anthomyiidae) parassita delle larve di Temnorrhinus mendicus Gyll. // Redia. V. 45. P. 245–253.
Cole E., James Jr., Streams Fr. 1970. Insects emerging from brown slime fluxes in southern New England // Canadian Entomology. V. 102 (3). P. 321–333.
Keilin D., 1917. Recherches sur les Anthomyides a larves carnivores // Parasitology. V. 9 (3). P. 325–450.
Perry I., Stubbs A.E., 1978. Some micro-habitats. Dead wood and sap runs. Stubbs A., Chandler P. (eds) A Dipterist’s Handbook. Amateur Entomologist. V. 15. P. 65–73.
Skidmore P., 1985. The biology of the Muscidae of the World. Dordrecht / Boston / Lancaster: Dr. W. Junk Publishers. 550 p.
Smith K.G.V., 1989. An introduction to the immature stages of British flies. Diptera larvae, with notes on eggs, puparia and pupae. Handbooks for the Identification of British Insects. V. 10. Pt. 14. 280 p
Sorokina V. S., 2015. New species of the genus Phaonia R.-D., 1830 (Diptera, Muscidae) from Central Asia // Zootaxa. V. 4013 (4). P. 571–587.
Tate P., 1935. The larva of Phaonia mirabilis Ringdahl, predatory on mosquito larvae (Dipt., Anthomyiidae) // Parasitology. V. 27 (4). P. 556–560.
Young Ch.W., 1999. New species and immature instars of crane flies of subgenus Tipulodina Enderlein from Sulawesi (Insecta: Diptera: Tipulidae: Tipula) // Annals of Carnegie Museum. V. 68. № 2. P. 81–90.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал