

Зоологический журнал, 2019, T. 98, № 2, стр. 149-162
Структура видового разнообразия почвенных ногохвосток (Hexapoda, Collembola) сосновых лесов Кавказа и русской равнины: мультимасштабный подход
Н. А. Кузнецова 1, *, А. И. Бокова 1, А. К. Сараева 2, Ю. Б. Швеенкова 3
1 Московский государственный педагогический университет
129164 Москва, Россия
2 Институт леса Карельского научного центра РАН
185910 Петрозаводск, Россия
3 Государственный природный заповедник “Приволжская лесостепь”
127642 Пенза, Россия
* E-mail: mpnk@yandex.ru
Поступила в редакцию 10.05.2018
После доработки 28.06.2018
Принята к публикации 6.06.2018
Аннотация
Сравнивали структуру разнообразия коллембол зеленомошных сосняков, расположенных в горах Кавказа и на Русской равнине. Работа основана на интенсивных количественных учетах с использованием мультимасштабной схемы, позволяющей оценить число видов на пространственных шкалах от нескольких квадратных сантиметров до сотни квадратных метров. Проведение учетов в однотипных местообитаниях разных районов и регионов дало возможность сравнить структуру разнообразия ногохвосток географически удаленных территорий, несмотря на недостаточную изученность их фауны. Результаты показали, что видовое богатство на всех пространственных шкалах в горах примерно вдвое выше, чем на равнине. По данным аддитивного парционирования, различия видового состава на участках разного размера возрастали пропорционально расстоянию между ними. Число видов коллембол в однотипных местообитаниях внутри региона обычно сходно, а различия списков связаны в основном с замещением видов. Соотношение между числом видов региональной и локальной фауны на равнине было ниже, чем в горах. Это указывает на большую степень неполночленности регионального пула видов на Русской равнине. Поскольку в обоих регионах многочисленны одни и те же широкораспространенные виды и на Кавказе видовое богатство выше, можно предположить, что горные леса этого региона были одним из источников заселения коллемболами лесов Русской равнины в голоцене.
Разнообразие биологического сообщества сегодня рассматривают с точки зрения его окружения и истории. Такой подход реализуют в рамках ландшафтной экологии (Forman, Godron, 1986; Wien et al., 1993; Ricklefs, 2005) и/или макроэкологии, если речь идет о крупных пространственных шкалах (Brown, Maurer,1989; Gaston, 2000; Buschke et al., 2014). “Раздвижение горизонтов” пространства и времени позволило рассматривать локальное разнообразие с точки зрения миграции, вымирания и видообразования (MacArthur, Wilson, 1967), а также в аспекте метасообщества, в котором между отдельными местообитаниями происходит обмен видами (Leibold et al., 2004). Был выявлен фрактальный характер пространственного распределения морских донных животных (Azovsky et al., 2007), показано влияние возраста ландшафта на локальное разнообразие (Zaitsev et al., 2013), обнаружены принципиальные различия факторов, связывающих локальное и региональное разнообразие, с одной стороны, и региональное и биогеографическое – с другой (Buschke et al., 2014) и т.д.
Эти подходы лишь недавно начали применять в почвенной зоологии (Bardgett et al., 2005; Decaëns, 2010). Исследования показали, что разнообразие раковинных амеб заповедных лесов Среднего Урала в большей мере зависит от типа леса, чем от разнообразия микросайтов в пределах местообитаний (Tsyganov et al., 2015). Напротив, именно разнообразие микросайтов вносило наибольший вклад в видовое богатство клещей в лесах Ирландии (Bolger et al., 2014). Для коллембол сосновых лесов Русской равнины была дана оценка вклада в разнообразие разных уровней: природных подзон, районов, разных местообитаний и однородных участков внутри местообитания (Kuznetsova, Saraeva, 2018). Было показано, например, что на одном квадратном метре лесной подстилки можно обнаружить в среднем около трети видов коллембол сосновых лесов изученного региона. Наибольший вклад в разнообразие вносили подзоны лесов, существенно различались и входящие в них районы.
Объект наших исследований – коллемболы – повсеместно разнообразная и многочисленная группа мелких почвенных членистоногих. На Русской равнине группа изучена довольно подробно усилиями многих специалистов: Э.Г. Беккера, Е.Ф. Мартыновой, Е.Л. Солнцевой и их последователями сегодня. Тем не менее, число видов в регионе с учетом современных таксономических подходов не известно. По нашей экспертной оценке, в регионе обитает порядка пятисот видов ногохвосток, большинство из них относится к редким. В широколиственных лесах отмечено не менее 120 видов (Чернов и др., 2010), в хвойных – более 160 видов даже по данным 30-летней давности (Кузнецова, 1988).
Региональное разнообразие любых таксонов зависит от климатических и исторических факторов. Низкое видовое богатство в высоких широтах традиционно связывают с недостатком тепла и, как следствие, с низкой продуктивностью экосистем (Чернов, 1975; Одум, 1975; Gaston, 2000). Другая возможная причина – эволюционная молодость сообществ послеледниковых ландшафтов (Qian, Rickfels 2004; Эволюция экосистем Европы…, 2008; Новенко и др., 2016). Факт того, что эти факторы влияют на видовое разнообразие регионов, не вызывает сомнений у большинства специалистов. Однако значение исторических факторов для разнообразия на локальном уровне продолжают обсуждать (Акатов и др., 2007; Ávila-Jiménez, Coulson, 2011; Zaitsev et al., 2013).
На локальном уровне видовая насыщенность зональных сообществ коллембол умеренного пояса оценивается как 30–35 видов при однократных учетах. Интересно, что локальное разнообразие сходно в диапазоне от южных тундр до широколиственных лесов (Ананьева и др., 1987; Чернов, 2008). По данным этих авторов, положительная корреляция числа видов со средней температурой июля проявляется лишь на региональном, но не на локальном уровне. Такая независимость разнообразия ногохвосток на локальном уровне от регионального не характерна для большинства таксонов животных (Gaston, 2000). В этой связи встает вопрос, насколько этот уровень локальной видовой насыщенности сохраняется в регионах с повышенным разнообразием (hot spots), таких, как Кавказ. По имеющимся данным (Добролюбова, 1982, 1984), видовое богатство коллембол в хвойных лесах Тебердинского заповедника на Северном Кавказе находится на том же уровне (порядка 30–35 видов на местообитание в одной учетной серии), что и на равнине. Однако оценка локального разнообразия ногохвосток горных лесов Кавказа нуждается в уточнении с учетом существенных изменений, которые произошли в таксономии группы в последние десятилетия. Так, наши недавние исследования в лесах Кавказа обнаружили немало новых для науки видов (Smolis, Kuznetsova, 2016, 2018 и др.).
В задачи работы входило: 1) сравнить видовое богатство коллембол аналогичных лесов Кавказа и Русской равнины на разных пространственных шкалах, 2) сопоставить соотношение локального и регионального богатства в горах и на равнине, 3) оценить неполночленность фаун гор и равнины.
МАТЕРИАЛЫ И МЕТОДЫ
Подходы к исследованию. Выявление более или менее полного видового состава коллембол на уровне регионов Кавказа и даже Русской равнины – задача вряд ли выполнимая в обозримое время. Фауна коллембол Кавказа, как обсуждалось выше, изучена фрагментарно. Русская равнина изучена лучше, однако и для этого региона видовые списки постоянно пополняются. В такой ситуации сравнение региональных фаун коллембол по накопленным в литературе данным не вполне адекватно и требует иных подходов. Такие подходы разработаны в ландшафтной экологии, где изучают локально-региональные процессы, в том числе сравнивают разнообразие на разных пространственных шкалах, используя стандартную, единообразную процедуру учетов (Colwell, Coddington, 1994). Мы оценивали данные в следующих масштабах: регионов (Кавказ – Русская равнина), районов (Тебердинский и Северо-Осетинский заповедники на Кавказе, Кивач и Приокско-Террасный – на Русской равнине), однотипных местообитаний (по два в каждом районе, кроме Приокско-Террасного заповедника, где было изучено только одно местообитание), однотипных участков площадью 1 м2 внутри местообитаний (рис. 1).
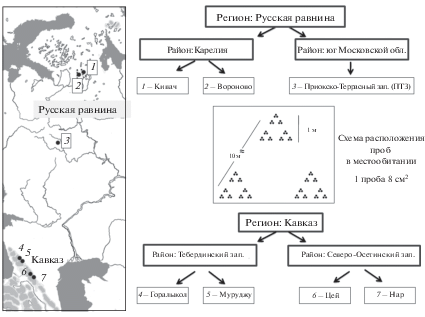
Особенность нашего подхода заключается в интенсивной выборке, предполагающей одновременное изъятие большого числа проб в одном местообитании. Даже в этом случае материал не претендует на полноту выявления видового состава местообитания, но позволяет корректно сравнивать структуру разнообразия в разных масштабах.
Пробные площади. Для сравнения были выбраны зеленомошные сосняки, широкораспространенные как на Русской равнине, так и на Кавказе. Все изученные участки были расположены на территории заповедников. На Русской равнине исследование проводили в заповеднике Кивач (средняя тайга) и Приокско-Террасном заповеднике (подзона хвойно-широколиственных лесов), на Кавказе – на северном макросклоне Главного Кавказского хребта в Тебердинском и Северо-Осетинском заповедниках (рис. 1).
В Карелии и на юге Московской обл. среднегодовая температура воздуха 2.4 и 4.5°С, средняя температура воздуха летом 14.7 и 17.3°С, зимой –9.7 и –9°С, соответственно, при сходном количестве осадков около 600 мм в год. В Тебердинском и Северо-Осетинском заповедниках на высотах от 1.5 до 1.8 км, где были расположены изучаемые леса, среднегодовая температура воздуха 6 и 4°С, средняя температура воздуха летом 14.5 и 12.7, зимой –2.1 и –8.8°С, количество осадков 700 и 800 мм в год, соответственно (Научно-прикладной справочник …2018). Таким образом, горные леса Кавказа в целом существуют в условиях более теплых зим, прохладного лета и большего количества осадков.
Участки равнинных зеленомошных сосняков, где были взяты учетные серии, описаны нами ранее (Сараева и др., 2015; Kuznetsova, Saraeva, 2018). Для данной работы были взяты карельские серии “Кивач”, июнь 2012 г., и “Вороново”, июнь 2011 г., на юге Московской обл. в Приокско-Террасном заповеднике – серия “ПТЗ”, июнь 2010 г.
Серия “Горалыкол” − Тебердинский заповедник, сосняк зеленомошно-вейниковый на высоте 1805 м над ур. м., склон восточной экспозиции. 43.4514 с. ш., 41.8236 в. д. В древесном ярусе − сосна обыкновенная (Pinus sylvestris), сомкнутость крон 0.5–0.6, в подлеске ель восточная (Picea orientalis), береза Литвинова (Betula litwinowii), рябина обыкновенная (Sorbus aucuparia), лещина обыкновенная (Corylus avellanа), жимолость настоящая (Lonicera xylosteum). В напочвенном покрове – Calamagrostis arundinaceae, Oxalis acetosella, преобладает мох Pleurozium schreberi. Средняя толщина подстилки 8 см. Почва – бурозем каменистый.
Серия “Муруджу” − Тебердинский заповедник, сосняк зеленомошно-мертвопокровный на высоте 1588 м над ур. м., склон восточной экспозиции. 43.3829 с. ш., 41.7135 в. д. В древесном ярусе − сосна обыкновенная с примесью пихты (Abies nordmanniana) и ели восточной, сомкнутость крон 0.7, в подлеске − ель восточная, пихта, бук восточный (Fagus orientalis). В напочвенном покрове − C. arundinacea, O. acetosella, преобладает мох Hylocomium splendens. Средняя толщина подстилки 6 см. Почва супесчаная подзолистая. Бурозем с признаками оподзоливания.
Серия “Цей” − Северо-Осетинский заповедник, сосняк зеленомошно-мертвопокровно-рододендроновый на высоте 1676 м над ур. м., склон западной экспозиции. 42.7971 с.ш., 43.9242 в.д. В древесном ярусе − сосна обыкновенная с примесью бука восточного (сомкнутость 0.6), в подлеске − ель восточная, бук восточный, рододендрон желтый (Rhododendron luteum), бересклет бородавчатый (Euonymus verrucosa) и др. В напочвенном покрове – кислица O. acetosella, встречаются Pyrola rotundifolia, Fragaria vesca, C. arundinacea, Lizula pilosa и др. Средняя толщина подстилки 7 см. Почва бурозем.
Серия “Нар”– Северо-Осетинский заповедник, сосняк зеленомошно-мелкотравный на высоте 1786 м над ур. м., склон северной экспозиции. 42.6792 с.ш., 44.0303 в.д. В древесном ярусе − сосна обыкновенная, сомкнутость крон 0.7–0.8, в подлеске − береза Литвинова, рябина обыкновенная, жимолость (Lonicera sp.) и др. В напочвенном покрове – O. acetosella, Poa nemoralis, F. vesca, Veronica sp., Alchemilla sp, преобладает мох H. splendens. Средняя толщина подстилки 11 см. Почва бурозем.
Почва всех участков горных лесов была нейтральной или слабощелочной (рН 6–7).
Изученные зеленомошные сосняки Русской равнины, где были взяты серии “Кивач”, “Вороново” и “ПТЗ”, отличались от горных в целом меньшим участием лиственных пород в древесном ярусе, бóльшим проективным покрытием зеленых мхов, меньшей толщиной подстилки.
Схема учетов в местообитании. В каждом из семи местообитаний было взято для анализа по одной учетной серии из 81 пробы. Образцы отбирали с помощью бура, площадью 8 см2, на всю глубину подстилки с захватом 1–2 см верхних минеральных слоев почвы. Для более корректного сравнения данных из разных местообитаний все участки выбирали в относительно однородном напочвенном покрове межкронового пространства. Серия состояла из трех групп проб, взятых на расстоянии 10 м друг от друга. Каждая группа включала 27 проб в пределах участка, площадью 1 м2. Пробы располагали гнездовым способом для выявления числа видов в разных масштабах: 8 см2 (одна проба), 1 дм2 (три пробы), 6 дм2 (9 проб) и 1 м2 (27 проб) (рис. 1).
Изученные пространственные шкалы. Исследование было основано на иерархическом подходе к изучению структуры разнообразия и охватывало следующие масштабы территорий:
1) регион (их два – Кавказ и Русская равнина);
2) район внутри региона. Всего 4 района, по два в каждом регионе: Северо-Осетинский и Тебердинский заповедник – на Кавказе, Приокско-Террасный заповедник и Карелия – на Русской равнине;
3) пробные площади в местообитаниях (сайт, site), т.е. в конкретных сосновых лесах района. Каждый район был представлен выборками из двух зеленомошных сосняков: Горалыкол и Муруджу в Тебердинском заповеднике, Цей и Нар – в Северо-Осетинском, Кивач и Вороново – в Карелии. Исключение – район Приокско-Террасного заповедника (серия ПТЗ), где был изучен один сосняк;
4) участки, площадью 1 м2, отражающие гетерогенность местообитания, т.е. сайта. Было изучено по три таких участка в каждом местообитании.
Коллембол экстрагировали на воронках Тульгрена. Всего получен 9291 экземпляр. Весь материал заключали в микропрепараты с жидкостью Фора и определяли до вида.
Методы анализа. Для описания пространственной структуры разнообразия использовали метод аддитивного парционирования, при котором количественно оценивают альфа-, бета- и гамма-компоненты (Lande, 1996; Crist et al., 2003; Gering et al., 2003). В нашем случае под альфа-компонентом понимали число видов участка, площадью 1 м2, под бета-компонентом – различия между ними по видовому составу. На следующем уровне альфа-компонент – это число видов в сайте (αsite), а бета-компонент – различия видового состава сайтов (βsite). На верхнем уровне анализа альфа-компонент – это число видов в сосняках района, а бета-компонент – различия между районами по видовому составу, относящимися к одному региону (Кавказ) или разным регионам (Кавказ и Русская равнина). Все эти компоненты рассчитывали, как средние величины для разных масштабов. Результаты аддитивного парционирования показали относительный вклад разнообразия регионов (Кавказ, Русская равнина), сайтов (Горалыкол и др. см. ниже) и участков, площадью 1 м2 внутри сайтов, в общее разнообразие коллембол всех изученных сосняков, γtotal. Принятая нами схема учетов позволяет рассчитать вклад в общее разнообразие: регионов (βreg), сайтов-местообитаний (βsite), мозаичности внутри сайтов (βsam) и участков площадью 1 м2 (αsam). В наиболее полном виде формула для расчетов:
Кроме того, отдельно для Кавказа был рассчитан вклад уровня района (βloc), т.е. Северо-Осетинского и Тебердинского заповедников, в разнообразие коллембол этого региона: γreg = αsam + + βsam + βsite + βloc.
В анализ включали комбинации выборок, по одному местообитанию из каждого района: Горалыкол-Нар-Кивач-ПТЗ, Муруджу-Нар-Вороново-ПТЗ и т.д. Аддитивное парционирование проводили для этих восьми комбинаций учетных серий в интегрированной среде RStudio, пакете программ “vegan” (R Development CoreTeam, 2011). В результате удалось оценить средний вклад в разнообразие коллембол выделов разного масштаба: регионов, районов, местообитаний, их мозаичности и участков площадью 1 м2.
Структура бета-компонента. Сходство видового состава каких-либо выборок традиционно рассчитывают с помощью коэффициентов Жаккара, Серенсена и др. (например, Песенко, 1978). Однако анализ несходства (dissimilarity) выборок дает не менее содержательные результаты (Legendre, 2014). Отличия в видовом составе, которые оценивает индекс несходства D, могут быть проявлением как разницы в числе видов (RichDiff), так и несовпадения списков (“замещения видов”, Repl), т.е. D = Repl + RichDiff (Legendre, 2014). Мы рассчитывали коэффициент несходства по матрице присутствия-отсутствия на основе формулы Жаккара:
Repl = 2 мин (b, c)/(a + b + c), где b – число видов, представленных в первой выборке, но отсутствующих во второй выборке; c – число видов, представленных во второй выборке, но отсутствующих в первой выборке; a – число видов, общих для обеих выборок.
Видовое богатство, т.е. число видов на участках определенной площади, оценивали внутри местообитания для участков 8 см2 (1 проба), 1 дм2 (3 пробы), 6 дм2 (9 проб), 1 м2 (27 проб) и 100 м2 (81 проба). Для анализа связи числа видов с площадью рассчитывали соотношения числа видов между участками разной площади. Полноту выявления видового богатства на участках разной площади оценивали с помощью кумулятивных кривых и индекса Chao2 в программе PAST (Hammer et al., 2001).
Для оценки соотношения локального и регионального разнообразия использовали индекс неполночленности N/S, где N − число видов видового фонда, S – локальное видовое богатство (Акатов и др., 2003; Акатов, Чефранов, 2007). Чем меньше значения N/S, тем выше уровень неполночленности регионального пула. В нашей работе за размер видового пула региона (N) было принято число видов, выявленное во всех сериях либо на Кавказе, либо на равнине. В качестве локального богатства (S) рассматривали число видов изученного сайта-местообитания по данным учетной серии (81 проба).
РЕЗУЛЬТАТЫ
Фаунистические особенности регионов. В материале, собранном на Кавказе, отмечено 79 видов коллембол, в том числе 4 эндемика: Ceratophysella caucasica, Endonura dobrolyubovae, E. kremenitsai, Neanura sp. n. aff. minuta. Эндемиками Кавказа могут оказаться еще 16 видов из родов Friesea, Pseudachorutes, Odontella, Oligaphorura и др. при дальнейшем изучении фауны региона (Приложение 1 ). Общая выборка из равнинных лесов включала всего 36 видов (Приложение 2 ). Все встреченные на равнине виды отмечены и на Кавказе или были выявлены для фауны этого региона ранее. Некоторые рода коллембол в лесном поясе гор отличались большим разнообразием, по сравнению с равниной: Folsomia, Endonura, Friesea, мелкие онихиуриды (Oligaphorura).
Важно отметить, что одни и те же широкораспространенные виды многочисленны в обоих регионах: Parisotoma notabilis, Isotomiella minor, Lepidocyrtus lignorum и др. (Приложение 1, 2 ).
Таксономическая структура фауны коллембол горных и равнинных местообитаний отличалась отношением числа видов к числу родов. На Кавказе эта величина (1.5) выше, чем на равнине (1.1). В сериях из сосновых лесов Кавказа рода Pygmarrhopalites, Ceratophysella, Endonura, Folsomia, Friesea, Mesaphorura, Tomocerus и др., как правило, представлены двумя и больше видами. На равнине виды коллембол одной учетной серии обычно относились к разным родам, несмотря на то, что в региональной фауне другие виды этих родов присутствуют.
Видовое богатство и численность. В пределах одного местообитания при однократном учете обнаруживали от 20 до 23 видов на равнине и почти вдвое больше в горах: 36–46. Общая численность коллембол в сосняках Кавказа была также в среднем вдвое выше, чем на равнине: 26.1 и 13.0 тыс. экз./м2, соответственно.
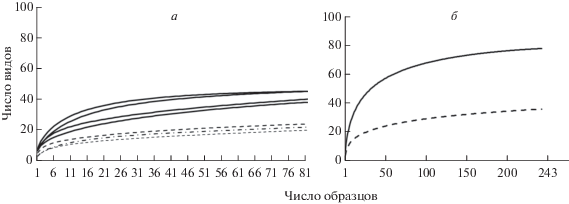
Для выяснения полноты выявления видового состава использовали кумулятивные кривые (рис. 2). Эти кривые, даже по данным однократных учетов, отражают довольно полное выявление видового состава изученных участков на равнине и менее полное на Кавказе, 97% и 72–84% по индексу Chao 2, соответственно.
Рис. 2.
Полнота выявления видового состава в горных и равнинных лесах, а – в отдельных местообитаниях, б – в регионах. Данные серий по: — Кавказу, - - - Русской равнине.
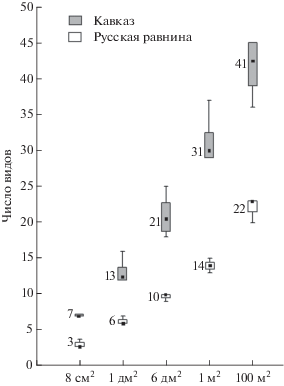
В среднем на участках 1 м2 в местообитаниях равнины отмечено 14–18 видов, в горах – 25–34. Несмотря на эти различия, квадратный метр как равнинных, так и горных сосняков включал 70–75% видов, обнаруженных в местообитании. На всех масштабах рассмотрения число видов ногохвосток в горных лесах было примерно вдвое больше, чем в равнинных (рис. 3).
Рис. 3.
Число видов на участках разной площади внутри зеленомошных сосняков на Кавказе и Русской равнине. Внутри каждой фигуры: точка – медиана, “ящик” – верхний и нижний квартили, “усы” – максимальное и минимальное значения числа видов. Цифра около каждой фигуры – среднее арифметическое значение числа видов.
Соотношение регионального пула и локального видового богатства. Отношение числа видов региональной фауны к локальной (индекс неполночленности, N/S) в горах выше: 2.0 vs 1.6 (табл. 1). Из этого следует, что видовой состав разных сосняков на равнине меньше различается, чем в горах, и, следовательно, в большей мере характеризует региональный пул видов. Таким образом, можно заключить, что неполночленность регионального пула коллембол Русской равнины выше, чем на Кавказе.
Таблица 1.
Соотношение между региональным и локальным числом видов коллембол (значения индекса неполночленности, N/S)
| Сайт | N/S |
|---|---|
| Горалыкол | 1.9 |
| Муруджу | 2.3 |
| Цей | 2.1 |
| Нар | 1.9 |
| Горы в среднем (±SD) | 2.0 ± 0.2 |
| Кивач | 1.5 |
| Вороново | 1.7 |
| ПТЗ | 1.6 |
| Равнина в среднем (±SD) | 1.6 ± 0.1 |
Оценка вклада факторов, связанных с разными пространственными шкалами в суммарное разнообразие коллембол сосновых лесов гор и равнины. Было изучено восемь комбинаций из местообитаний-сайтов так, чтобы в каждую комбинацию входили представители всех четырех изученных районов, по два от каждого региона (табл. 2). Результаты аддитивного парционирования показали, что наибольшие различия связаны с региональной шкалой. Сосняки Кавказа и Русской равнины различались по видовому составу коллембол (βreg) на 40–42%. Гетерогенность внутри местообитания (βsam), т.е. несходство участков площадью 1 м2 – 11–13%, была меньше, чем различия между однотипными местообитаниями-сайтами, βsite – 17–19%. На одном участке площадью 1 м2 (αsam) в среднем обитало 27–30% всех видов, отмеченных в сосняках обоих регионов. Важно отметить, что результаты аддитивного парционирования всех восьми комбинаций пробных площадей, представляющих разнообразие коллембол Кавказа и Русской равнины, оказались весьма сходными.
Таблица 2.
Структура разнообразия коллембол сосняков Кавказа и Русской равнины на разных пространственных шкалах
| Выборка | αsam | βsam | βsite | βreg |
|---|---|---|---|---|
| Горалыкол–Нар–Кивач–ПТЗ | 0.30 | 0.11 | 0.17 | 0.42 |
| Горалыкол–Цей–Вороново–ПТЗ | 0.30 | 0.12 | 0.17 | 0.41 |
| Муруджу–Цей–Вороново–ПТЗ | 0.27 | 0.12 | 0.18 | 0.42 |
| Муруджу–Нар–Вороново–ПТЗ | 0.27 | 0.13 | 0.18 | 0.42 |
| Горалыкол–Цей–Кивач–ПТЗ | 0.30 | 0.11 | 0.19 | 0.40 |
| Горалыкол–Нар–Вороново–ПТЗ | 0.30 | 0.12 | 0.16 | 0.42 |
| Муруджу–Цей–Кивач–ПТЗ | 0.27 | 0.12 | 0.20 | 0.41 |
| Муруджу–Нар–Кивач–ПТЗ | 0.27 | 0.12 | 0.19 | 0.42 |
Отдельно для региона Кавказа по выборке Горалыкол-Муруджу-Цей-Нар была рассчитана оценка вклада в разнообразие уровня района (βloc), т.е. территорий разных заповедников этого региона. Оказалось, что видовой состав ногохвосток в лесах разных заповедников различался на 27%. В этом анализе αsite = 0.53, βsite = 0.20, βloc = 0.27.
Интересные результаты дал анализ структуры разнообразия коллембол сосняков Кавказа и Русской равнины по-отдельности: αsam : βsam : βsite = = 0.37 : 0.16 : 0.47 на Кавказе и 0.44 : 0.17 : 0.39 на Русской равнине, соответственно. Это означает, что видовой состав ногохвосток на участках площадью 1 м2 в разных лесах равнины более сходен, чем в горах, т.к. на участках этого размера можно обнаружить относительно бóльшую долю видов от всего разнообразия региона (αsam = 0.44 vs 0.37). Вклад в разнообразие различий между участками одного местообитания (βsam = 0.16 и 0.17) в горах и на равнине сходен. На уровне местообитаний горные леса больше различаются по видовому составу коллембол, чем равнинные (0.47 vs 0.39). Таким образом, дифференцировка структуры разнообразия коллембол горных и равнинных лесов на уровне сайтов различается, а внутри сайтов – сходна.
Результаты аддитивного парционирования для всех рассмотренных выше выборок отличались от случайных, р < 0.01.
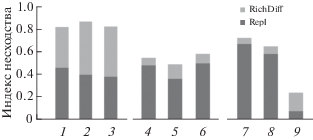
Анализ несходства видового состава. Попарное сравнение данных разных выборок одного объема показало, что различия в видовом составе коллембол разных регионов связаны как с замещением видов, так и с разным видовым богатством (столбики 1–3, рис. 4). Различия однотипных местообитаний внутри регионов были связаны в основном с замещением видов (столбики 4–8). Однако было и исключение: выборки Кивач и Вороново из соседних районов Карелии различались в первую очередь числом видов (столбик 9).
Рис. 4.
Компоненты несходства видовых списков ногохвосток зеленомошных сосняков разных территорий: разница в числе видов (RichDiff) и замещение видов (Repl). 1 – Русская равнина и Кавказ, 2 – Карелия и Северо-Осетинский заповедник, 3 – Карелия и Tебердинский заповедник, 4 – Tебердинский и Северо-Осетинский заповедники, 5 – Горалыкол и Муруджу, 6 – Цей и Нар, 7 – Kивач и ПТЗ, 8 – Вороново и ПТЗ, 9 – Кивач и Вороново.
ОБСУЖДЕНИЕ
Связь числа видов с площадью (species-area relationships, SAR) в разных группах организмов обычно носит степенной, реже – имеет иной характер (Connor, McCoy, 2001). Подобные исследования на почвенно-зоологических объектах немногочисленны (Decaëns, 2010). Обнаруживаемые при этом закономерности, показанные для клещей, ногохвосток и дождевых червей (Judas, 1988; Maraun et al., 2007; Ulrich, Fiera, 2009), как и наши данные, в целом подтверждают тренды, ранее обнаруженные для надземных организмов (Rosenzweig, 1995).
Анализ пространственной структуры разнообразия коллембол показал, что видовое богатство на участках одного размера в пределах местообитания в горах Кавказа выше, чем на равнине, примерно вдвое. Эти различия могут быть обусловлены тремя факторами: климатом, рельефом и историческими причинами. Температурные характеристики макроклимата могут не оказывать существенного влияния на локальное разнообразие ногохвосток больших территорий, если они не выходят за рамки более или менее оптимальных. В случаях перехода в область пессимальных температур локальное разнообразие проявляет явную зависимость от тепла и заметно снижается, например, при переходе от южных тундр к полярным пустыням (Бабенко, 2017). В таких условиях зависимость распределения коллембол от температуры проявляется даже на уровне микрорельефа, т.е. в микромасштабе. В изученных нами районах гор и равнины температурные условия для коллембол вполне подходящие. В горах, однако, больше осадков, что благоприятно для коллембол, в основе склонных к гигрофилии. В целом, климатические различия, связанные с широтным градиентом изученных территорий, вряд ли можно рассматривать в качестве основного фактора локального разнообразия коллембол.
Другой аспект: во время оледенений больше возможностей сохранения видов в рефугиумах, наличие которых обеспечивается сложным рельефом (например, Deharveng et al., 2015). Действительно, в кавказских сериях обнаружено больше видов (36–45), чем на Русской равнине (20–23). Наши данные сопоставимы с оценками разнообразия коллембол хвойных лесов гор Западной Европы в районах, которые не были покрыты плейстоценовыми ледниками: в пихтовом лесу Альп 47 видов (Deharveng et al., 2015), ельниках Богемии 45–50 видов (Čuchta, Shrubovych, 2015), сосняках Пиренеев 46 видов (Benito, Sanchez, 2000). В нашем материале это соотносится с присутствием эндемиков, а также более высокими значениями индекса В/Р, т.е. отношения числа видов к числу родов в лесах Кавказа, по сравнению с Русской равниной. Принято считать, что, чем выше этот показатель, тем больше выражены автохтонные процессы формирования биоты, и наоборот, чем ниже – тем бóльшую роль играет миграция видов, т.е. аллохтонные процессы (Малышев, 1969; Толмачев, 1974). Низкие значения индекса В/Р для коллембол Русской равнины указывают скорее на аллохтонное происхождение фауны ногохвосток этого региона. В населении ногохвосток лесов равнины и гор обычно доминируют одни и те же широкораспространенные виды ногохвосток (Parisotoma notabilis, Isotomiella minor, Приложения 1 и 2 ), но на Кавказе видовое богатство выше. По-видимому, это означает, что Кавказ мог быть одним из источников заселения коллемболами Русской равнины в голоцене. Однако и на Кавказе фауна ногохвосток, по-видимому, сохранила исходное разнообразие не в полной мере, так как эндемичные виды редки даже на заповедных территориях.
Известно, что геологические процессы, такие как плейстоценовые оледенения, существенно повлияли на флору и фауну Русской равнины. Заселение послеледниковых ландшафтов происходило с горных территорий, а также из рефугиумов в поймах крупных рек (Величко, 2002; Эволюция экосистем Европы…, 2008). Динамика ледового покрова Кавказа достаточно подробно изучена (обзор Solomina et al., 2016). Пересеченный рельеф способствовал сохранению рефугиумов в этом регионе, что подтверждают находки реликтовых растений плиоценовой эпохи в Приэльбрусье (Серебрянный, 1980). По-видимому, близость рефугиумов способствовала более быстрому восстановлению популяций коллембол после отступлений ледников в горах.
Возраст ландшафта как фактор разнообразия лишь в последнее время начал привлекать внимание почвенных зоологов. Показано, что этот фактор может иметь весьма большое значение для сообществ микроартропод, что показано для панцирных клещей на территории Голландии (Zaitsev et al., 2013). Равнины, некогда занятые ледниками, сегодня населяют немногие, в основном космополитные, виды дождевых червей, в то время как горные районы отличаются высокой степенью эндемизма и разнообразия (Перель, 1979; Квавадзе, 1985; Pop et al., 2010).
Длительное антропогенное воздействие на лесные экосистемы хорошо документировано для Русской равнины (Эволюция экосистем Европы…, 2008). Однако и на Кавказе уменьшение площади лесов за последние 600–700 лет имеет, вероятно, антропогенную природу (Соломина, 1999). Показателем преобразованности коренных лесов гор может быть и небольшое число эндемичных видов коллембол, отмеченных на Северном Кавказе.
Аддитивное парционирование всей выборки коллембол гор и равнины показало, что наибольшие различия обусловлены особенностями регионов, а в пределах Русской равнины – спецификой лесных подзон (Kuznetsova, Saraeva, 2018). В обоих случаях основной вклад в разнообразие вносили максимально удаленные территории (подзоны или регионы). Как показали компоненты несходства видовых списков сайтов равнины и Кавказа, большая величина βreg (более 40%) обусловлена, прежде всего, разным уровнем видового богатства регионов. Другие авторы также связывают наибольшие фаунистические различия коллембол в бореальных лесах европейской части России именно с экорегионами, соответствующими подзонам тайги, в то время как участки внутри одной подзоны различались в меньшей степени (Saifutdinov et al., 2018). В нашем случае различия в числе видов возрастали с ростом расстояния между участками: различия списков видов регионов больше, чем различия списков районов, и т.д. βreg > βloc > βsite > βsam. Такой на первый взгляд тривиальный результат, по-видимому, обусловлен изучением именно однотипных местообитаний. В случае охвата разных типов сосняков в Карелии, от сухого лишайникового до сырого сфагнового, вклад различий между ними был больше, чем различий на уровне районов: βsite > βloc (Kuznetsova, Saraeva, 2018).
Приложение 1 . Видовой состав коллембол в сосновых лесах Кавказа (число особей на серию проб)
| Семейство, вид | Тебердинский заповедник | Северо-Осетинский заповедник | ||
|---|---|---|---|---|
| Горалыкол | Муруджу | Цей | Нар | |
| Hypogastruridae | ||||
| Ceratophysella caucasica Martynova | 32 | 16 | 3 | |
| Ceratophysella sp. | 38 | 2 | 210 | 10 |
| Xenylla sp. | 3 | |||
| Willemia anophthalma Börner | 11 | 9 | 4 | 2 |
| W. scandinavica Stach | 1 | |||
| W. denisi Mills sensu Hüther | 1 | |||
| Neanuridae | ||||
| Friesea mirabilis (Tullberg) | 10 | |||
| F. truncata Cassagnau | 63 | |||
| Friesea cf. subterranea Cassagnau | 3 | |||
| Friesea sp. 1 | 1 | 12 | ||
| Friesea sp. 2 | 25 | 9 | 2 | |
| Anurida sp. | 2 | 4 | ||
| Micranurida forsslundi Gisin | 3 | 2 | ||
| M. pygmaea Börner | 2 | 9 | 3 | 9 |
| Pseudachorutes cf. dubius Krausbauer | 3 | 2 | 2 | |
| Pseudachorutes sp. 1 | 5 | |||
| Neanura minuta Gisin | 1 | 1 | ||
| Neanura sp. n. aff. minuta Gisin | 2 | |||
| Neanurinae gen. sp. | 1 | |||
| Endonura dobrolyubovae Smolis et Kuznetsova | 12 | 3 | ||
| E. kremenitsai Smolis et Kuznetsova | 7 | 2 | ||
| Endonura sp. | 6 | 2 | ||
| Odontellidae | ||||
| Xenyllodes armatus Axelson | 4 | 2 | ||
| Odontella sp. | 1 | |||
| Onychiuridae | ||||
| Kalaphorura sp. | 1 | |||
| Orthonychiurus sp. | 1 | |||
| Protaphorura sakatoi (Yosii) | 3 | 109 | 16 | |
| P. subarctica Martynova | 54 | 4 | ||
| Protaphorura sp. aff. jernika Kaprus, Weiner et Pasnik | 5 | |||
| Oligaphorura absoloni (Börner) | 9 | 3 | 21 | 9 |
| Oligaphorura cf. caucasica Weiner et Kaprus | 2 | |||
| Oligaphorura cf. difference Bagnall | 5 | |||
| Oligaphorura sp. n. 1 aff. kremenitsai Shvejonkova et Potapov | 3 | |||
| Oligaphorura sp. n. 2 aff. kremenitsai Shvejonkova et Potapov | 10 | 2 | ||
| Oligaphorura сf. pieninensis (Weiner) | 5 | |||
| Tullbergiidae | ||||
| Mesaphorura critica Ellis | 18 | 2 | 14 | 7 |
| M. tenuisensillata Rusek | 4 | 1 | 34 | 3 |
| M. hylophila Rusek | 17 | 72 | 4 | 2 |
| M. italica (Rusek) | 8 | 5 | ||
| Mesaphorura sp. | 1 | |||
| Isotomidae | ||||
| Pseudanurophorus binoculatus Kseneman | 11 | |||
| Folsomia quadrioculata (Tullberg) | 114 | |||
| F. manolachei Bagnall | 107 | 144 | ||
| F. pseudodiplophthalma Stach | 53 | 374 | 277 | 8 |
| F. penicula Bagnall | 1 | |||
| F. litsteri Bagnall | 21 | 3 | 5 | |
| F. fimetarioides (Axelson) | 9 | |||
| F. inoculata Stach | 12 | |||
| Folsomia juv. | 7 | 3 | 1 | |
| Isotomiella minor (Schäffer) | 312 | 168 | 221 | 109 |
| Proisotoma subminuta Denis | 1 | |||
| Parisotoma notabilis (Schäffer) | 232 | 305 | 192 | 198 |
| Parisotoma sp. 1 | 1 | |||
| Desoria hiemalis (Schött) | 11 | 1 | ||
| Desoria sp. | 60 | |||
| Tomoceridae | ||||
| Tomocerina minuta (Tullberg) | 47 | 100 | 1 | 41 |
| Tomocerus sp. 1 | 7 | 1 | 21 | 13 |
| T. vulgaris (Tullberg) | 270 | 7 | 36 | 27 |
| Pogonognathellus flavescens (Tullberg) | 18 | |||
| Tomoceridae juv. | 1 | |||
| Entomobryidae | ||||
| Entomobrya marginata (Tullberg) | 14 | |||
| Entomobrya sp.1 | 3 | 7 | ||
| Lepidocyrtus lignorum (Fabricius) | 237 | 148 | 74 | 224 |
| L. violaceus (Geoffroy) | 30 | 4 | 7 | |
| Pseudosinella alba (Packard) | 7 | 1 | 7 | |
| Pseudosinella cf. zygophora (Schille) | 3 | 4 | ||
| Pseudosinella cf. immaculata (Lie-Pettersen) | 1 | |||
| Orchesella sp. 1 | 92 | 1 | 4 | |
| Orchesella sp. 2 | 25 | |||
| Willowsia buski (Lubbock) | 1 | |||
| Entomobryidae juv. | 15 | 14 | 2 | 37 |
| Sminthurididae | ||||
| Sphaeridia pumilis (Krausbauer) | 32 | |||
| Arrhopalitidae | ||||
| Pygmarrhopalites cochlearifer (Gisin) | 14 | 14 | 34 | 4 |
| P. secundarius Gisin | 20 | 4 | 158 | |
| Katiannidae | ||||
| Sminthurinus elegans (Fitch) | 2 | |||
| Dycirtomidae | ||||
| Dicyrtoma fusca (Lubbock) | 3 | 3 | ||
| D. grinbergsi Stebaeva | 5 | |||
| Dicyrtomina ornata (Nicolet) | 1 | |||
| Dicyrtomina sp. | 5 | |||
| Ptenothrix atra (Linnaeus) | 6 | |||
| Sminthuridae | ||||
| Сaprainea marginata (Schött) | 1 | |||
| Lipothrix lubbocki (Tullberg) | 18 | 39 | 12 | 35 |
| Spatulosminthurus flaviceps Tullberg | 39 | 18 | 9 | 21 |
| Neelidae | ||||
| Megalothorax sp. | 525 | 167 | 56 | |
| Прочие | 2 | 1 | ||
| Число экз. | 2234 | 1647 | 1708 | 1178 |
| Число родов | 29 | 27 | 24 | 29 |
| Число видов | 46 | 36 | 40 | 45 |
| Число видов/родов | 1.6 | 1.3 | 1.7 | 1.6 |
Приложение 2 . Видовой состав коллембол в сосновых лесах Русской равнины (число особей на серию проб)
| Семейство, вид | Kивач | Вороново | ПТЗ |
|---|---|---|---|
| Hypogastruridae | |||
| Ceratophysella denticulata (Bagnall) | 1 | ||
| Schoettella ununguiculata (Tullberg) | 1 | ||
| Willemia anophthalma Börner | 64 | 4 | 16 |
| Xenylla mucronata Axelson | 1 | ||
| Neanuridae | |||
| Neanura muscorum (Templeton) | 2 | 8 | 3 |
| Anurida sp. | 3 | ||
| Micranurida pygmaea Börner | 42 | 14 | 73 |
| Pseudachorutes corticicollus (Schäffer) | 4 | 1 | |
| P. dubius Krausbauer | 3 | ||
| P. subcrassus Tullberg | 5 | ||
| Odontellidae | |||
| Xenyllodes armatus Axelson | 3 | ||
| Onychiuridae | |||
| Protaphorura cancellata (Gisin) | 15 | ||
| Oligaphorura absoloni (Börner) | 31 | 1 | 9 |
| Tullbergiidae | |||
| Mesaphorura yosii (Rusek) | 92 | 18 | |
| M. macrochaeta Rusek | 100 | ||
| Isotomidae | |||
| Anurophorus septentrionalis Palissa | 62 | 34 | |
| Folsomia manolachei Bagnall | 16 | ||
| Proisotoma minima (Absolon) | 1 | 1 | |
| Isotomiella minor (Schäffer) | 338 | 192 | 155 |
| Parisotoma notabilis (Schäffer) | 255 | 46 | 122 |
| Desoria hiemalis (Schött) | 278 | 134 | |
| D. blekeni (Leinaas) | 15 | ||
| D. olivacea (Tullberg) | 1 | ||
| Isotoma viridis Bourlet | 1 | 1 | |
| Tomoceridae | |||
| Pogonognatellus flavescens (Tullberg) | 1 | 2 | |
| Entomobryidae | |||
| Entomobrya nivalis (Linnaeus) | 11 | 1 | |
| Orchesella flavescens (Bourlet) | 53 | 78 | 10 |
| O. bifasciata Nicolet | 5 | ||
| Lepidocyrtus lignorum (Fabricius) | 53 | 45 | 12 |
| Entomobryoidea juv. | 4 | ||
| Sminthurididae | |||
| Sphaeridia pumilis (Krausbauer) | 1 | ||
| Arrhopalitidae | |||
| Pygmarrhopalites cochlearifer (Gisin) | 1 | 6 | 1 |
| P. principalis (Stach) | 6 | 3 | |
| Katiannidae | |||
| Sminthurinus bimaculatus (Axelson) | 1 | ||
| Bourletiellidae | |||
| Heterosminthurus claviger (Gisin) | 1 | ||
| Neelidae | |||
| Megalothorax minimus Willem | 31 | 1 | 30 |
| Число экз. | 1348 | 591 | 585 |
| Число родов | 18 | 19 | 20 |
| Число видов | 23 | 20 | 22 |
| Число видов/родов | 1.3 | 1.1 | 1.1 |
Довольно сложен вопрос, насколько изученные нами сообщества неполночленны. Чем больше сходство локального и регионального наборов видов, тем выше уровень неполночленности регионального пула (Акатов и др., 2003). Подход был предложен и успешно применен для анализа неполночленности различных фитоценозов Кавказа. При этом сравнивали число видов на участках площадью 1 га относительно участков в 300 м2. Неполночленным оказался древесный ярус низко- и верхнегорных лесов Западного Кавказа, в отличие от насыщенных видами среднегорных лесов (Акатов, Чефранов, 2007). Эти результаты, по мнению авторов, указывают на то, что не только региональное, но и локальное разнообразие зависит от исторических процессов. У коллембол неполночленность регионального пула видов на Русской равнине выражена сильнее, чем на Кавказе. Это обстоятельство наряду с другими (обилие на равнине широкораспространенных и партеногенетических видов) позволяет предположить, что процесс формирования разнообразия таксона в лесных экосистемах постгляциального ландшафта далеко не завершен.
БЛАГОДАРНОСТИ
Авторы благодарят А.П. Гераськину и Н.Е. Шевченко за помощь в организации экспедиции и описание пробных площадей, А.Б. Бабенко и В.В. Акатова за ценные замечания при работе над рукописью, М.Б. Потапова за консультации в определении ряда видов.
Работа выполнена при финансовой поддержке РФФИ (16-04-01228).
Список литературы
Акатов В.В., Чефранов С.Г., Акатова Т.В., 2003. Об эволюционной полночленности видовых фондов современных растительных сообществ высокогорной зоны Западного Кавказа // Журнал общей биологии. Т. 64. № 4. С. 308–317.
Акатов В.В., Чефранов С.Г., 2007. Локальное видовое богатство древесного яруса лесов острова Мадагаскар и Западного Кавказа: опыт тестирования исторической гипотезы путем анализа структуры распределения обилия видов // Бюллетень Московского общества испытателей природы. Отд. биол. Т. 112. № 1. С. 65–71.
Ананьева С.И., Бабенко А.Б., Чернов Ю.И., 1987. Ногохвостки (Collembola) в арктических тундрах Таймыра // Зоологический журнал. Т. 66. № 7. С. 1032–1044.
Бабенко А.Б., 2017. Ногохвостки (Collembola) Восточной Чукотки: особенности фауны и населения // Зоологический журнал. Т. 96. № 10. С. 1141–1164.
Величко А.А., 2002. Устойчивость ландшафтной оболочки и ее био- и георазнообразие в свете динамики широтной зональности // Известия АН. Сер. геогр. № 5. С. 7–21.
Добролюбова Т.В., 1982. Пространственное распределение ногохвосток в горных сосновых лесах Тебердинского заповедника // Антропогенное воздействие на фауну почв. М.: МГПИ. С. 25–36.
Добролюбова Т.В., 1984. Фауна коллембол темнохвойных лесов Северо-Западного Кавказа // Фауна и экология ногохвосток. М.: Наука. С. 95–98.
Квавадзе Э.Ш., 1985. Дождевые черви (Lumbricidae) Кавказа. Тбилиси: Мецниереба. 238 с.
Кузнецова H.А., 1988. Типы населения коллембол в хвойных лесах европейской части СССР // Экология микроартропод лесных почв. М.: Наука. С. 24–52.
Малышев Л.И., 1969. Зависимость флористического богатства от внешних условий и исторических фактов // Ботанический журнал. Т. 54. № 8. С. 1137–1147.
Научно-прикладной справочник “Климат России”, 2018 [Электронный ресурс]. Режим доступа: http://meteo.ru/pogoda-i-klimat/197-nauchno-prikladnoj-spravochnik-klimat-rossii. Дата обновления: 15.11.2018.
Новенко Е.Ю., Цыганов А.Н., Волкова Е.М., Бабешко К.В., Лаврентьев Н.В., Мазей Ю.А., 2016. Изменения растительности и климата на северо-западе Среднерусской возвышенности в голоцене // Известия АН. Сер. геогр. № 1. С. 103–114.
Одум Ю., 1975. Экология. Т. 2. Пер. с англ. М.: Мир. 376 с.
Перель Т.С., 1979. Распространение и закономерности распределения дождевых червей фауны СССР. М.: Наука. 272 с.
Песенко Ю.А., 1978. Концепция видового разнообразия и индексы, его измеряющие // Журнал общей биологии. Т. 39. № 3. С. 380–393.
Сараева А.К., Потапов М.Б., Кузнецова Н.А., 2015. Разномасштабное распределение коллембол (Collembola) в однородном напочвенном покрове: устойчивость параметров в пространстве и времени // Зоологический журнал. Т. 94. № 9. С. 1029–1045.
Серебрянный Л.Р., 1980. Древнее оледенение и жизнь. М.: Наука. 128 с.
Соломина О.Н., 1999. Горные ледники Северной Евразии в Голоцене. М.: Научный мир. 264 с.
Толмачев А.И., 1974. Введение в географию растений. Л.: Изд-во ЛГУ. 243 с.
Чернов А.В., Кузнецова Н.А., Потапов М.Б., 2010. Население коллембол восточноевропейских широколиственных лесов // Зоологический журнал. Т. 89. № 5. С. 559–573.
Чернов Ю.И., 1975. Природная зональность и животный мир суши. М.: Мысль. 222 с.
Чернов Ю.И., 2008. Экология и биогеография. Избранные работы. М: Товарищество научных изданий КМК. 500 с.
Эволюция экосистем Европы при переходе от плейстоцена к голоцену (24-8 тыс. л.н.), 2008. М.: Товарищество научных изданий КМК. 556 с.
Ávila-Jiménez M.L., Coulson S.J., 2011. A Holarctic biogeographical analysis of the Collembola (Arthropoda, Hexapoda) unravels recent post-glacial colonization patterns // Insects. V. 2. P. 273–296.
Azovsky A.I., Burkosky I.V., Kolobov M.Yu., Kucheruk N.V., Saburova M.A., et al., 2007. Self-similar properties of the spatial structure of intertidal macro- and microbenthic communities // Журнал общей биологии. Т. 68. № 3. С. 180–194.
Bardgett R.D., Yeates G.W., Anderson J.M., 2005. Patterns and determinants of soil biological diversity // Biological diversity and function in soils (ed. by R.D. Bardgett, M.B. Usher and D.W. Hopkins). Cambridge: Cambridge University Press. P. 100–118.
Benito J.C., Sanchez M.J., 2000. Ecology of soil springtails (Collembola, Insecta) from pine woods and Rhododendron shrublands in the Central and Eastern Pyrenees (North Spain) // Pedobiologia. V. 44. P. 430–441.
Bolger T., Arroyo J., Kenny J., Caplice M., 2014. Hierarchical analysis of mite community structures in Irish forests – a study of the relative contribution of location, forest type and microhabitat // Applied Soil Ecology. V. 83. P. 39–43.
Brown J.H., Maurer B.A., 1989. Macroecology: the division of food and space among species on continents // Science. № 243. P. 1145–1150.
Buschke F.T., Brendonck L., Vanschoenwinkel B., 2014. Differences between regional and biogeographic species pools highlight the need for multi-scale theories in macroecology // Frontiers of Biogeography. V. 6. P. 173–184
Colwell R.K., Coddington J.A., 1994. Estimating Terrestrial Biodiversity through Extrapolation // Philosophical Transactions of the Royal Society of London B: Biological Sciences. V. 345. P. 101–118.
Connor E.F., McCoy E.D., 2001. Species–area relationships // Encyclopedia of Biodiversity. V. 5. NY: Academic Press. P. 397–411.
Crist T.O., Veech J.A., Gering J.C., Summerville K.S., 2003. Partitioning Species Diversity across Landscapes and Regions: A Hierarchical Analysis of α, β and δ Diversity // The American Naturalist. V. 162. № 6. P. 734–743.
Čuchta P., Shrubovych J., 2015. Soil Collembola communities in montane coniferous forests of the Bohemian Forest // Silva Gabreta. V. 21. № 2. P. 149–156.
Decaëns T., 2010. Macroecological patterns in soil communities // Global Ecology and Biogeography. V. 19. P. 287–302.
Deharveng L., Bedos A., Daugeron C., Villemant C., Judson M.L.I., 2015. Organization, usefulness and limitations of an ATBI (All Taxa Biodiversity Inventory): the inventory of terrestrial invertebrates in the Mercantour National Park // Zoosystema. V. 37. № 1. P. 9–30.
Forman R., Godron M., 1986. Landscape Ecology. NY: Wiley. 619 p.
Gaston K.J., 2000. Global patterns in biodiversity // Nature. № 405. P. 220–227.
Gering J.C., Crist T.O., Veech J.A., 2003. Additive partitioning of species diversity across multiple spatial scales: Implications for regional conservation of biodiversity // Biological Conservation. № 17. P. 488–499.
Hammer Ř., Harper D.A.T., Ryan P.D., 2001. PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica. 4,1. http://nhm2.uio. no/norlex/past/download.html.
Judas M., 1988. The species-area relationship of European Lumbricidae (Annelida, Oligochaeta) // Oecologia. V. 76. P. 579–587.
Kuznetsova N.A., Saraeva A.K., 2018. Beta-diversity partitioning approach in soil zoology: A case of Collembola in pine forests // Geoderma. V. 332. P. 142–152.
Lande R., 1996. Statistics and Partitioning of Species Diversity, and Similarity Among Multiple Communities // Oikos. V. 76. № 1. P. 5–13.
Legendre P., 2014. Interpreting the replacement and richness difference components of beta diversity // Global Ecology and Biogeography. № 23. P. 1324–1334.
Leibold M.A., Holyoak M., Mouquet N., Amarasekare P., Chase J.M. et al., 2004. The metacommunity concept: a framework for multi-scale community ecology // Ecology Letters. № 7. P. 601–613.
MacArthur R.H., Wilson E.O., 1967. The Theory of Island Biogeography. Princeton: Princeton Univ. Press. 203 p.
Maraun M., Schatz H., Scheu S., 2007. Awesome or ordinary? Global diversity patterns of oribatid mites // Ecography. V. 30. № 2. P. 209–216.
Pop A. A., Pop V. V., Csuzdi C., 2010. Significance of the Apuseni Mountains (the Carpathians) in the origin and distribution of Central European earthworm fauna: (Oligochaeta: Lumbricidae) // Zoology in the Middle East. T. 51. № 2. P. 89–110.
Qian H., Rickfels R., 2004. Taxon Richness and Climate in Angiosperms: Is There a Globally Consistent Relationship That Precludes Region Effects? // American Naturalist. V. 163. P. 773–779.
R Development CoreTeam, 2011. R: Язык и среда для статистических вычислений. Вена. Австрия: Фонд Р статистических вычислений.
Ricklefs R., 2005. Historical and ecological dimensions of global patterns in plant diversity. Biologiske Skrifter. V. 55. P. 583–603.
Rosenzweig M.L., 1995. Species Diversity in Space and Time. Cambridge: University Press. 436 p.
Saifutdinov R.A., Gongalsky K.B., Zaitsev A.S., 2018. Springtail (Hexapoda: Collembola) fauna in the burnt boreal forests of European Russia // Invertebrate Zoology. V. 15. №1. P. 115–130.
Smolis A., Kuznetsova N., 2016. Remarkable diversity of the genus Endonura Cassagnau, 1979 (Collembola: Neanuridae: Neanurinae) in the Caucasus // Zootaxa. V. 4200. № 1. P. 47–82.
Smolis A., Kuznetsova N., 2018. Paravietnura gen. n., a new intriguing genus of Neanurini from the Caucasus (Collembola, Neanuridae, Neanurinae) // ZooKeys. № 739. P. 41–54.
Solomina O., Bushueva I., Dolgova E., Jomelli V., Alexandrin M. et al., 2016. Glacier variations in the Northern Caucasus compared to climatic reconstructions over the past millennium // Global and Planetary Change. № 140. P. 28–58.
Tsyganov A.N., Komarov A.A., Mitchell E.A.D, Shimano S., Smirnova O.V., Aleynikov A.A., Mazei Yu A., 2015. Additive partitioning of testate amoeba species diversity across habitat hierarchy within the pristine southern taiga landscape (Pechora-Ilych Biosphere Reserve, Russia) // European Journal of Protistology. V. 70. P. 861–864.
Ulrich W., Fiera C., 2009. Environmental correlates of species richness of European springtails (Hexapoda: Collembola) // Oecologica. № 35. P. 45–52.
Wiens J.A., Stenseth N.C., Van Horne B., Ims R.A., 1993. Ecological Mechanisms and Landscape Ecology // Oikos. V. 66. № 3. P. 369–380.
Zaitsev A.S., van Straalen N.M., Berg M.P., 2013. Landscape geological age explains large scale spatial trends in oribatid mite diversity // Landscape Ecology. № 28. P. 285–296.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал