

Зоологический журнал, 2019, T. 98, № 3, стр. 343-352
Распределение и территориальная неоднородность населения мелких млекопитающих южной тайги Западной Сибири
А. А. Кислый 1, Ю. С. Равкин 1, 2, *, И. Н. Богомолова 1, А. А. Одинцева 1, С. М. Цыбулин 1, О. А. Одинцев 3
1 Институт систематики и экологии животных СО РАН
630091 Новосибирск, Россия
2 Томский государственный университет
634050 Томск, Россия
3 Омский государственный педагогический университет
644099 Омск, Россия
* E-mail: zm.nsc@yandex.ru
Поступила в редакцию 15.12.2017
После доработки 18.10.2018
Принята к публикации 08.11.2018
Аннотация
Южная тайга Западной Сибири разделена по меридиану 75° в.д. на две полосы, которые мы условно считаем провинциями. Результаты учета мелких млекопитающих канавками, заборчиками и давилками во второй половине лета 1962, 1967–1971, 1975, 1977–1994, 2005, 2006, 2012 и 2013 гг. усреднены по группам выделов карты “Растительность Западно-Сибирской равнины” (Ильина и др., 1976) за все годы работ в пределах провинций. На основании усредненных вариантов населения по результатам кластерного анализа составлена классификация 29 видов зверьков по сходству их распределения. Выявлено 6 типов преференции: эвритопный, лесной, полевой, болотный, пойменный и синантропный. При классификации сообществ мелких млекопитающих выделено четыре типа населения: 1 – лесов и полей-перелесков, 2 – низинных и переходных болот, 3 – верховых болот и 4 – пойм крупных рек. Первые три типа разделены по сходству сообществ на два подтипа каждый, что совпадает с различиями по провинциальности. Наиболее высокие показатели плотности населения, общего количества встреченных видов и фонового богатства свойственны населению лесного типа. Во всех типах населения по обилию лидирует обыкновенная бурозубка (Sorex araneus (Linnaeus 1758)). По составленной классификации населения и структурному графу прослежена связь изменений сообществ с заболоченностью, облесенностью, продуктивностью фитоценозов (в поселках – с кормностью), провинциальностью, застроенностью, рельефом (пойменный – не пойменный) и распашкой. Сила и общность связи факторов среды и неоднородности сообществ максимальна по заболоченности. Плотность и видовое богатство населения мелких млекопитающих увеличиваются с возрастанием облесенности и трофности болот и уменьшаются при увеличении заболоченности, степени распаханности и заливания в половодье. Кроме того, суммарное обилие мелких млекопитающих повышается с запада на восток. Показано преимущество использования усредненных данных для выявления закономерностей неоднородности населения. Сопоставлены настоящие и более ранние результаты анализа пространственной неоднородности населения мелких млекопитающих исследованной подзоны.
Южная тайга – одна из подзон лесной зоны Западной Сибири. Подзона расположена между 57° и 60° с.ш. и приурочена в основном к Средне-Обской впадине, большая часть которой представляет собой плоские слабодренированные участки (Орлов, 1963).
Межгодовая динамика численности мелких млекопитающих существенна. Поэтому, при исследовании их распределения и пространственной неоднородности населения, сравнимость данных можно увеличить, усредняя материалы в пределах групп выделов геоботанической или ландшафтной карты в пределах зон, подзон и провинций. Поскольку учеты были проведены на различных территориях в разные годы, средние значения по группам выделов в некоторой степени отражают пространственные различия в целом за годы работ.
Ранее проанализировано население мелких млекопитающих южной тайги Западной Сибири как в отдельности, так и в составе сообществ позвоночных в целом (Равкин, Лукьянова, 1976). Распределение зверьков рассмотрено по всей Западно-Сибирской равнине с разделением на зоны и подзоны (Равкин и др., 1996). Различия сообществ по годам описаны на примере Приобья (Цыбулин, Богомолова, 2012) и Прииртышья (Одинцева, 2013; Кислый и др., 2014). Основная цель настоящей статьи – выявление пространственной неоднородности населения мелких млекопитающих южной тайги Западной Сибири и оценка связи этой изменчивости с факторами среды, определяющими пространственно-типологические различия сообществ.
МАТЕРИАЛЫ И МЕТОДЫ
Для проведения анализа использован ряд сведений из банка данных лаборатории зоологического мониторинга Института систематики и экологии животных СО РАН, опубликованных ранее (Никифоров и др., 1968; Равкин, Лукьянова, 1976; Сапогов, 1983; Пучковский, 1984; Стариков, 1985; Равкин и др., 1996; Цыбулин, Богомолова, 2012; Одинцева, 2013; Кислый и др., 2014). Информация о банке данных и правила для вкладчиков доступны на сайте лаборатории (Банк данных: информация, правила для вкладчиков, 2012). Учеты мелких млекопитающих проведены в 23 географических пунктах в период с 16 июля по 31 августа в течение 29 полевых сезонов: 1962, 1967–1971, 1975, 1977–1994, 2005, 2012, и 2013 гг. (рис. 1, табл. 1). Всего проанализированы результаты отловов зверьков в 306 биотопах, считая места повторного сбора данных в аналогичных местообитаниях и в разные годы.
Рис. 1.
Точки проведения учетов (1–23) мелких млекопитающих в южной тайге Западной Сибири. Подробная информация о местах сборов материалов приведена в табл. 1.
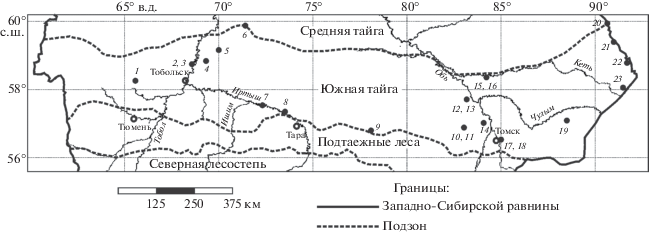
Таблица 1.
Места проведения учетов мелких млекопитающих в южной тайге Западной Сибири
| № | Места работ* | Координаты, град | Годы проведения учетов | Участники работ | Публикация материалов | |
|---|---|---|---|---|---|---|
| в.д. | с.ш. | |||||
| 1 | Свердловская обл., Тавдинский р-н, д. Ленино | 58.13 | 65.34 | 1992 | Юдкин В.А., Торопов К.В. | Равкин и др., 1996 |
| 2 | Тюменская обл., Уватский р-н, р. Миссия | 58.71 | 66.63 | 1968 | Равкин Ю.С., | Равкин, Лукьянова, 1976 |
| 3 | с. Горнослинкино | 58.76 | 68.8 | |||
| Лукьянова И.В. | ||||||
| 1971 | Равкин Ю.С. | |||||
| 1991 | Шор Е.Л., Торопов К.В. | Равкин и др., 1996 | ||||
| 2012 | Одинцева А.А. | Одинцева, 2013 | ||||
| 2013 | Одинцева А.А., Макаров А.В. | Кислый и др., 2014 | ||||
| 4 | п. Чебунтан | 58.94 | 69.14 | 1979–1981 | Стариков В.П. | Стариков, 1985 |
| 5 | п. Демьянка | 59.54 | 70 | |||
| 6 | Нефтеюганский р-н, п. Салым | 60.01 | 71.46 | 1975 | Пучковский С.В. | Пучковский, 1984 |
| 7 | Омская обл., Тевризский р-н, с. Тевриз | 57.51 | 72.4 | 1979, 1980 | Стариков В.П. | Стариков, 1985 |
| 8 | Прииртышье в пределах Омской обл. | ~57 | ~73.5 | 1968–1988 | Соловьев С.А. | Равкин и др., 1996 |
| 9 | Новосибирская обл., Северный р-н, с. Останинка и с. Украинка |
56.72 | 78.26 | 1993 | Шор Е.Л., Бобков Ю.В., Торопов К.В., Вартапетов Л.Г., Богомолова И.Н. |
|
| 10 | Томская обл., Бакчарский р-н, с. Карагай | 56.93 | 83 | 1967–1969 | Лукьянова И.В. | Равкин, Лукьянова, 1976 |
| 1970 | Равкин Ю.С. | |||||
| 1990 | Юдкин В.А., Шор Е.Л., Богомолова И.Н. | Равкин и др., 1996 | ||||
| 2005 | Цыбулин С.М., Климова Н.В. | Цыбулин, Богомолова, 2012 | ||||
| 11 | п. Плотниково | 56.86 | 83.08 | 1990 | Юдкин В.А., Шор Е.Л., Богомолова И.Н. | Равкин и др., 1996 |
| 2005 | Цыбулин С.М., Климова Н.В. | Цыбулин, Богомолова, 2012 | ||||
| 12 | Чаинский р-н, с. Коломино | 57.83 | 83.31 | 1970 | Равкин Ю.С. | Равкин, Лукьянова, 1976 |
| 1990 | Юдкин В.А., Шор Е.Л., Богомолова И.Н. | Равкин и др., 1996 | ||||
| 2006 | Цыбулин С.М. | Цыбулин, Богомолова, 2012 | ||||
| 13 | п. Кузурово | 57.87 | 83.34 | 1970 | Равкин Ю.С. | Равкин, Лукьянова, 1976 |
| 1990 | Торопов К.В., Морковина Т.А., Богомолова И.Н. | Равкин и др., 1996 | ||||
| 2006 | Цыбулин С.М. | Цыбулин, Богомолова, 2012 | ||||
| 14 | Кривошеинский р-н, пойма Оби | ~57 | ~84 | 1978, 1981–1992 | Москвитина Н.С. | Равкин и др., 1996 |
| 15 | оз. Монатка | 57.36 | 84.15 | 1978, 1981–1983 | ||
| 16 | Верхнекетский р-н, д. Тайное | 58.39 | 84.11 | 1983 | ||
| 17 | г. Томск, Советский р-н, п. Заварзино | 56.47 | 85.1 | 1981–1988 | ||
| 18 | Томская обл., Томский р-н, д. Протопопово | 56.43 | 85.16 | 1978–1988 | ||
| 19 | Тегульдетский р-н, п. Четь-Конторка | 57.06 | 88.12 | 1994 | Бобков Ю.В., Торопов К.В. | |
| 20 | Красноярский край, Енисейский р-н, д. Фомка | 60.04 | 90.57 | 1977 | Бурский О.В., Вахрушев А.А. | Сапогов, 1983 |
| 21 | д. Колмогорово | 59.26 | 91.32 | |||
| 22 | с. Погодаево | 58.7 | 92.05 | |||
| 23 | Западная часть Красноярского края (левый берег Енисея) |
58 | 92 | 1962 | Наумов Р.Л., Никифоров Л.П. | Никифоров и др., 1968 |
Материалы собраны при отловах в 50-метровые канавки и заборчики из полиэтиленовой пленки с пятью цилиндрами или конусами, на четверть залитыми 4% раствором формалина, а также с помощью давилок. Методика учетов численности зверьков и обработки данных подробно описана ранее (Равкин, Ливанов, 2008). В целом, при учетах мелких млекопитающих в южной тайге Западной Сибири отработано около 45 тыс. цилиндро-суток и 20 тыс. давилко-суток. Материалы в банке данных хранятся в виде количества особей каждого вида в пересчете на 100 цилиндро- или давилко-суток в период проведения учета. Информации о числе отловленных зверьков нет. Все показатели обилия приведены к значениям в пересчете на 100 цилиндро-суток (ц.с.). Для этого число особей грызунов на 100 давилко-суток сначала пересчитано к их количеству на 1 км2 (умножением на 400), а потом уменьшено в 145 раз (множитель перевода со 100 ц.с. на 1 км2; Равкин, Ливанов, 2008). Собранные давилками материалы по обилию насекомоядных исключены из расчетов, поскольку эти зверьки очень редко попадают в давилки. Средние показатели рассчитаны без учета соотношения площадей местообитаний.
Для анализа все материалы предварительно усреднены по годам и группам выделов карты “Растительность Западно-Сибирской равнины” (Ильина и др., 1976) в пределах западной и восточной ее частей. Эти части мы условно считаем провинциями, разделенными по меридиану 75° в.д. Это позволяет в некоторой мере нивелировать как межгодовые, так и частные колебания обилия мелких млекопитающих. В указанные в тексте группы выделов входят коренные формации и их производные за исключением мелколиственных лесов, а также полностью или частично распаханных участков, которые рассмотрены в качестве отдельных местообитаний. Поэтому, если в тексте названа лишь коренная формация, сказанное о ней относится и к ее производным. Например, указание “в темнохвойной тайге” включает темнохвойно-мелколиственные и мелколиственно-темнохвойные леса, а также вырубки, гари и шелкопрядники. К многочисленным отнесены виды с обилием 10 и более особей/100 ц.с., к обычным – от 1 до 9, к редким – от 0.1 до 0.9, очень редким – менее 0.1 (Кузякин, 1962). Лидерами считали первые пять по обилию видов. Деление на фаунистические типы дано по Галкиной (по: Равкин, Лукьянова, 1976). Названия видов стандартизированы по сводке Павлинова и Лисовского (2012).
Пространственно-типологическая структура населения выявлена одним из методов кластерного анализа (Трофимов, 1976). В качестве меры сходства использован коэффициент Жаккара (Jaccard, 1902) для количественных признаков (Наумов, 1964). Оценка связи с факторами среды и природно-антропогенными режимами, как совокупностями неразделимых сочетаний факторов (Равкин, Ливанов, 2008), проведена с помощью линейной качественной аппроксимации (Равкин и др., 1978). Статистическая обработка материала выполнена с использованием программного обеспечения банка данных лаборатории зоологического мониторинга ИСиЭЖ СО РАН (Равкин, Ефимов, 2009).
После формализованного разбиения состав выделенных классов идеализирован, т.е. концептуально переработан. Для этого отклоняющиеся от разработанной концепции пробы перенесены в те группы, в которые они должны входить в соответствии с принятым объяснением. При этом, с одной стороны, оценка информативности классификации (доля учтенной ею дисперсии) снижается. С другой стороны, подобная перестановка упрощает понимание классификации и приводит ее в соответствие с нашими реальными представлениями о предмете исследования (Равкин, Ливанов, 2008).
Классификация видов мелких млекопитающих по сходству распределения выполнена экспертно по предпочтению ими обследованных местообитаний. Формализованный подход привел по предметным соображениям к менее четким представлениям.
РЕЗУЛЬТАТЫ
Распределение мелких млекопитающих
При многолетних учетах в южной тайге Западной Сибири отмечено 29 видов мелких млекопитающих. Это кроты: европейский (Talpa europaea (Linnaeus 1758)) и алтайский (T. altaica (Nikolsky 1883)); сибирская белозубка (Crocidura sibirica (Dukelsky 1930)); обыкновенная кутора (Neomys fodiens (Pennant 1771)); бурозубки: обыкновенная (Sorex araneus (Linnaeus 1758)), тундряная (S. tundrensis (Merriam 1900)), крупнозубая (S. daphaenodon (Thomas 1907)), средняя (S. caecutiens (Laxmann 1788)), равнозубая (S. isodon (Turov 1924)), плоскочерепная (S. roboratus (Holister 1913)), крошечная (S. minutissimus (Zimmermann 1780)) и малая (S. minutus (Linnaeus 1766)); лесная мышовка (Sicista betulina (Pallas 1779)); обыкновенный хомяк (Cricetus cricetus (Linnaeus 1758)); лесной лемминг (Myopus schisticolor (Liljeborg 1844)); полевки: красносерая (Craseomys rufocanus (Sundevall 1846)), европейская рыжая (Myodes glareolus (Schreber 1780)), красная (M. rutilus (Pallas 1779)), водяная (Arvicola amphibius (Linnaeus 1758)), узкочерепная (Lasiopodomys gregalis (Pallas 1779)), экономка (Alexandromys oeconomus (Pallas 1776)), обыкновенная (Microtus arvalis (Pallas 1778)) и темная (M. agrestis (Linnaeus 1761)); мыши: малютка (Micromys minutus (Pallas 1771)), малая лесная (Sylvaemus uralensis (Pallas 1811)), восточноазиатская (Apodemus peninsulae (Thomas 1907)), полевая (A. agrarius (Pallas 1776)), домовая (Mus musculus (Linnaeus 1758)), а также серая крыса (Rattus norvegicus (Berkenhout 1769)). Из них в среднем по подзоне (без учета соотношения площадей) многочисленна лишь обыкновенная бурозубка. Обычны 11 видов: средняя, равнозубая и малая бурозубки, полевки – европейская рыжая, красная и экономка, а также (в нижней части пределов обилия по баллу) обыкновенная кутора, красносерая, обыкновенная и темная полевки и полевая мышь. Редки тоже 11 видов и очень редки – шесть. По сходству распределения все виды разделены на шесть типов преференции (1–6) и девять подтипов (2.1–4.3).
1. Широко распространенный вид (темная полевка).
Виды, предпочитающие в основном11
2 – леса (европейский и алтайский кроты, средняя и равнозубая бурозубки, красная полевка и малая лесная мышь),
2.1 – особенно мелколиственные леса, а также низинные и переходные болота (обыкновенная кутора, лесная мышовка и лесной лемминг),
кроме того:
2.2 – переходные и верховые болота (крошечная бурозубка),
2.3 – поймы (европейская рыжая полевка),
2.4 – поля-перелески (обыкновенная и тундряная бурозубки, красносерая полевка и восточноазиатская мышь);
3 – поля-перелески и переходные болота:
3.1 – в основном поля-перелески (обыкновенный хомяк, обыкновенная полевка и полевая мышь),
3.2 – поля-перелески и переходные болота (мышь-малютка);
4 – переходные болота,
кроме того:
4.1 – леса (сибирская белозубка и плоскочерепная бурозубка),
4.2 – низинные болота (малая бурозубка),
4.3 – поймы (крупнозубая бурозубка и узкочерепная полевка);
5 – поймы (полевки – водяная и экономка);
6 – поселки (домовая мышь и серая крыса).
Более половины видов отнесены к лесному типу преференции. Втрое меньше их предпочитают, в основном, поля-перелески и переходные болота, или преимущественно болота. Еще два типа распределения составляют зверьки, предпочитающие поймы и поселки (по 2 вида). Лишь темная полевка распространена повсеместно.
Неоднородность населения мелких млекопитающих и ее экологическая организация
При классификации сообществ мелких млекопитающих выделено четыре типа населения. Первые три из них разделены на два подтипа каждый (табл. 2).
Таблица 2.
Классификация населения мелких млекопитающих южной тайги Западной Сибири
| Типы* (1–4) и подтипы (2.1–3.2) населения | Лидеры по обилию, % от общей плотности населения | Плотность населения, особей/100 ц.с. | Количество видов/ из них фоновых | Преобладающие по обилию типы фаун, %: | ||
|---|---|---|---|---|---|---|
| Европейский | Сибирский | Транспалеаркты | ||||
| 1 – лесов, полей- перелесков и поселков в их пределах | Обыкновенная и средняя бурозубки 30 и 16, красная и европейская рыжая полевка 15 и 7, равнозубая бурозубка 5 | 85 | 29/14 | 52 | 44 | – |
| 1.1 – западной провинции | Обыкновенная бурозубка 36, европейская рыжая полевка 14, средняя бурозубка 13, красная полевка 10, малая бурозубка 5 | 78 | 21/11 | 69 | 27 | – |
| 1.2 – восточной провинции | Обыкновенная и средняя бурозубки 28 и 19, красная полевка 18, равнозубая бурозубка 6, красносерая полевка 4 | 89 | 28/14 | 55 | 43 | – |
| 2 – низинных и переходных болот | Малая и обыкновенная бурозубки 24 и 14, полевая мышь 12, обыкновенная кутора и мышь-малютка по 7 | 42 | 28/10 | 74 | 17 | – |
| 2.1 – западной провинции | Малая и обыкновенная бурозубки 46 и 25, обыкновенная кутора и средняя бурозубка по 6, мышь-малютка 3 | 32 | 18/5 | 82 | – | – |
| 2.2 – восточной провинции | Обыкновенная и малая бурозубки 23 и 20, темная полевка 12, средняя бурозубка 9, обыкновенная полевка 6 | 69 | 26/13 | 70 | 23 | – |
| 3 – верховых болот | Средняя и обыкновенная бурозубки 35 и 17, красная полевка 13, малая бурозубка 9, европейская рыжая полевка 4 | 23 | 21/5 | 43 | 52 | – |
| 3.1 – западной провинции | Обыкновенная, средняя и малая бурозубки 35, 24 и 12, европейская рыжая и красная полевки по 6 | 17 | 17/5 | 59 | 35 | – |
| 3.2 – восточной провинции | Средняя бурозубка 41, красная полевка 15, обыкновенная и малая бурозубки 11, европейская рыжая полевка 4 | 27 | 17/5 | 33 | 63 | – |
| 4 – пойм крупных рек, долин их притоков и поселков в их пределах | Обыкновенная бурозубка 30, полевки: экономка и красная 24 и 8; малая бурозубка и европейская рыжая полевка по 6 | 50 | 28/10 | 56 | 16 | 28 |
Наибольшая плотность населения свойственна лесам и полям-перелескам, меньшая – поймам, низинным, переходным и особенно верховым болотам. Общее количество встреченных видов и фоновое богатство сообществ изменяется почти так же, уменьшаясь от лесов и полей-перелесков к поймам, и далее от низинных и переходных болот к верховым. Таким образом, плотность и видовое богатство населения мелких млекопитающих увеличиваются с возрастанием облесенности и трофности болот и уменьшаются при увеличении заболоченности, степени распаханности и заливания в половодье. Кроме того, суммарное обилие повышается с запада на восток.
Во всех типах населения по обилию лидирует обыкновенная бурозубка, а малая – почти повсеместно, кроме лесов и полей-перелесков. Равнозубая бурозубка как лидер отмечена только в лесах, а средняя – еще и на верховых болотах. Европейская рыжая и красная полевки входят в число лидеров во всех типах населения, кроме низинных и переходных болот. Там лидируют обыкновенная кутора, полевая мышь и мышь-малютка. Полевка-экономка отмечена как лидер только в поймах.
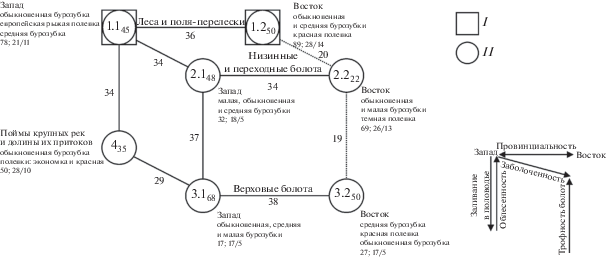
Пространственно-типологическая структура сообществ мелких млекопитающих выявлена при пороге сходства 29 единиц (рис. 2). Тип населения лесов и полей-перелесков за счет подтипов образует основной горизонтальный ряд (подтипы – 1.1 и 1.2). Вертикальную часть графа составляют внепойменные сообщества (1.1) и поймы (4), что отражает влияние заливания в половодье. Два горизонтальных ряда вариантов населения болот (2.1–2.2 и 3.1–3.2) отклонены по третьей оси (2.1–3.1) и коррелируют с заболоченностью территории. Вертикальные связи между ними определяют различия в трофности болот. Все горизонтальные связи соответствуют внутритиповому разделению в связи с провинциальными различиями населения. Максимальное сходство свойственно внутритиповым горизонтальным связям (34–38%). Наибольшее межтиповое сходство прослежено между восточными подтипами 1.2–2.2–3.2 (19 и 20%).
Рис. 2.
Пространственно-типологическая структура населения мелких млекопитающих южной тайги Западной Сибири. Население мелких млекопитающих: I – лесов; II – местообитаний, где чередуются облесенные и открытые участки. Цифрами внутри значков показаны номера типов и подтипов по классификации населения, индексом обозначено внутригрупповое сходство, у связей между значками – межгрупповое. Сплошными линиями показано сверхпороговое (значимое) сходство, пунктиром – дополнительные связи. Рядом со значками приведены названия трех первых по обилию видов, а также показатели населения: плотность (особей/ 100 ц.с.), общее/ фоновое видовое богатство. Стрелками внизу рисунка показаны направления увеличения влияния основных структурообразующих факторов среды. Запад и Восток – соответственно западная и восточная части равнины. Население всех типов, кроме четвертого, занимает территорию междуречий и надпойменных террас крупных рек.
При анализе иерархической классификации и структуры населения мелких млекопитающих южной тайги Западной Сибири прослежено влияние заболоченности, облесенности, кормности местообитаний, провинциальности, а также, в меньшей степени, застроенности, рельефа (пойма – не пойма) и, особенно, степени распаханности (табл. 3). Связь заболоченности и неоднородности сообществ на основании данных, усредненных по годам и группам выделов карты растительности раздельно по провинциям, максимальна по сравнению с другими факторами среды. Далее значимость факторов снижается по облесенности, продуктивности фитоценозов (в поселках – кормности) и провинциальности. Затем следуют менее значимые факторы – застроенность, рельеф и степень распаханности. Локальная значимость этих факторов несомненна, но за счет низкой представленности в целом по подзоне невелика.
Таблица 3.
Сила и общность связи факторов среды и неоднородности населения мелких млекопитающих южной тайги Западной Сибири
| Фактор, режим | Учтенная дисперсия, % | |
|---|---|---|
| индивидуально | нарастающим итогом | |
| Заболоченность | 18 | 18 |
| Облесенность | 13 | 25 |
| Продуктивность, кормность | 8 | 31 |
| Провинциальность | 6 | 37 |
| Застроенность | 3 | 39 |
| Рельеф (поймы и не поймы) | 2 | 41 |
| Степень распаханности | 1 | 42 |
| Все факторы | 42 | 42 |
| Режимы по иерархической классификации | 30 | 57 |
| Режимы по структуре | 40 | 58 |
| Все режимы | 40 | 58 |
| Все режимы и факторы | 58 | 58 |
| Коэффициент множественной корреляции | 0.76 | |
Влияние большей части факторов и режимов не ортогональны, т.е. взаимно скоррелированны. В результате множественная оценка связи меньше простой суммы индивидуальных значений. Степень такой корреляции можно проследить при расчетах нарастающим итогом – последовательным увеличением числа факторов и режимов. Так все факторы дают приращение по отношению к корреляции с заболоченностью в 1–7% дисперсии, а классификационные режимы – 16%. В целом все факторы и режимы объясняют 58% дисперсии (коэффициент множественной корреляции – 0.76).
ОБСУЖДЕНИЕ
В отличие от ранее опубликованной классификации (Равкин, Лукьянова, 1976), выполненной по не усредненным данным, включение дополнительных материалов позволило нивелировать межгодовые и частные различия местообитаний и сформулировать представления о пространственно-типологической неоднородности населения мелких млекопитающих и ее экологической организации. Разделенные до этого по составу лесообразующих пород лесные классы теперь составляют единый тип населения лесов и полей-перелесков. Так же объединены в один тип классы низинных и переходных болот. Отдельный тип включает в себя классы населения верховых болот. Исключение составляют сообщества пойменных лесов и болот, ранее отнесенные в лесной и болотный типы населения. Такие пойменные местообитания объединены по сходству с вариантами сообществ пойм крупных рек и долин их притоков. Центральная часть южной тайги Западной Сибири обследована недостаточно. Это вынудило условно разделить подзону на две полосы (западную и восточную), а не на три, как было сделано ранее для всей Западной Сибири (Равкин и др., 1996). При этом различия облика населения по соответствующим провинциям по t-критерию Стьюдента достоверны (уровень значимости 0.95).
Представления о пространственно-типологической структуре населения тоже отличаются от опубликованных ранее (Равкин, Лукьянова, 1976). Так, добавление новых материалов позволило выявить значимую связь между лесным и пойменным типами населения. Это так же укрепило связь пойм и верховых болот. Кроме того, была обнаружена связь населения лесов с населением низинных и переходных болот (ранее население низинных пойменных и внепойменных болот рассматривали как разные варианты населения).
Использование не усредненных данных по населению мелких млекопитающих при работе с материалами, имеющими широкий разброс по годам и ключевым участкам, затрудняло выявление пространственно-типологической структуры и основных структурообразующих факторов среды по причине значительных межгодовых и частных отличий сообществ. Оцененная по не усредненным (исходным) данным сила и общность связи неоднородности сообществ, обусловленной годовыми колебаниям обилия мелких млекопитающих, составляет 12% учтенной дисперсии, при этом межгодовые различия по t-критерию достоверны (уровень значимости 0.99). Информативность различий ключевых участков составляет 27% дисперсии матрицы сходства исходных проб. Оценка режимов по этим данным объясняет всего 5% дисперсии (40% для усредненных материалов). Таким образом, усредненные данные лучше отражают территориальные тренды изменений, а исходные – реальную информативность наших представлений по географической изменчивости населения мелких млекопитающих.
ЗАКЛЮЧЕНИЕ
При многолетних учетах в южной тайге Западной Сибири отмечено 29 видов млекопитающих. Из них в среднем по подзоне многочисленна лишь обыкновенная бурозубка, кроме нее еще 11 видов обычны и 11 видов редки. Очень редки шесть видов. По сходству распределения зверьки разделены на шесть типов преференции. В общих чертах это: эвритопный, лесной, полевой, болотный, пойменный и синантропный типы.
По результатам кластерного анализа выделено четыре типа населения: 1 – лесов, полей-перелесков и поселков в их пределах; 2 – низинных и переходных болот; 3 – верховых болот; 4 – пойм крупных рек, долин их притоков и поселков в их пределах. Население первых трех типов занимает территорию междуречий и надпойменных террас долин крупных рек, а население четвертого типа – их поймы и целиком долины притоков. Первые три типа разделены по сходству на два подтипа каждый, что совпадает с различиями по провинциальности. Связь заболоченности и неоднородности сообществ на основании данных, усредненных по годам и группам выделов карты растительности раздельно по провинциям, максимальна по сравнению с другими факторами среды. Остальные факторы, с учетом представленности в выборке, менее значимы. Локальное влияние этих факторов несомненно, но за счет низкой представленности в целом по южной тайге невелико.
Классификация населения мелких млекопитающих южной тайги Западной Сибири, разработанная ранее (Равкин, Лукьянова, 1976) по меньшему как в пространстве, так и по годам объему материала и с использованием менее совершенного алгоритма кластерного анализа, показала более дробное, но менее надежное по объяснению разделение сообществ. Это связано с годовыми колебаниями численности животных. После усреднения данных многие таксоны классификации уже не прослеживались, т.е. они были свойственны лишь отдельным годам и после усреднения данных не могли быть выявлены. Списки основных структурообразующих факторов среды по обоим массивам данных одинаковы, но иерархия факторов по их влиянию разная. Последнее связано не только с годовыми колебаниями численности зверьков, но и со спецификой использованных алгоритмов анализа. Кроме того, во время проведения первого этапа работ еще не была разработана программа оценки силы и общности связи факторов среды и изменчивости животного населения. Это также повлекло за собой различия в представлениях об их иерархии.
Список литературы
Банк данных: информация, правила для вкладчиков, 2012. Сайт лаборатории зоологического мониторинга ИСиЭЖ СО РАН [Электронный ресурс]. Режим доступа: http://eco.nsc.ru/zoomonit/zoomonit_ r.htm. Дата обновления: 05.01.2012.
Ильина И.С., Лапшина Е.И., Лавренко Н.Н., Мельцер Л.И., Романова Е.А. и др., 1976. Растительность Западно-Сибирской равнины. Карта масштаба 1 : 1 500 000. М.: ГУГК СССР.
Кислый А.А., Макаров А.В., Одинцев О.А., 2014. Влияние экстремальных погодных условий на обилие мелких млекопитающих // Зоологические чтения – 2014: материалы Всерос. науч.-практ. конф. с междунар. участием. Новосибирск: Изд-во НГПУ. С. 193–199.
Кузякин А.П., 1962. Зоогеография СССР // Ученые записки МОПИ им. Н.К. Крупской. Т. 109. № 1. С. 3–182.
Наумов Р.Л., 1964. Птицы природного очага клещевого энцефалита Красноярского края. Автореф. дис. … канд. биол. наук. М.: Московский областной педагогический институт. 19 с.
Никифоров Л.П., Наумов Р.Л., Бабенко Л.В., 1968. Вопросы эпидемиологии клещевого энцефалита и биологические закономерности в его природном очаге. М.: Медицина. 432 с.
Одинцева А.А., 2013. Межгодовые отличия населения земноводных и мелких млекопитающих южной тайги Западной Сибири // Динамика современных экосистем в голоцене. Казань: Изд-во “Отечество”. С. 264–265.
Орлов В.И., 1963. Зона лесов // Западная Сибирь. Отв. ред. Рихтер Г.Д. М.: Изд-во Академии наук СССР. С. 331–376.
Павлинов И.Я., Лисовский А.А., 2012. Млекопитающие России: систематико-географический справочник. М.: Товарищество научных изданий КМК. 604 с.
Пучковский С.В., 1984. Распространение и численность мелких млекопитающих в тайге Тюменской области // Тезисы докладов VIII Всесоюз. зоогеограф. конф. М. С. 353–354.
Равкин Ю.С., Богомолова И.Н., Ердаков Л.Н., Панов В.В., Буйдалина Ф.Р. и др., 1996. Особенности распределения мелких млекопитающих Западно-Сибирской равнины // Сибирский экологический журнал. Т. 3. № 3‑4. С. 307–317.
Равкин Ю.С., Ефимов В.М., 2009. Банк данных по численности и распределению животных в пределах бывшего СССР // Формирование баз данных по биоразнообразию – опыт, проблемы, решения. Материалы Международной научно-практической конференции. Барнаул: “АRТIКА” (ИП Жерносенко С.С.). С. 205–214.
Равкин Ю.С., Куперштох В.Л., Трофимов В.А., 1978. Пространственная организация населения птиц // Птицы лесной зоны Приобья. Новосибирск: Наука. С. 253–269.
Равкин Ю.С., Ливанов С.Г., 2008. Факторная зоогеография. Новосибирск: Наука. 205 с.
Равкин Ю.С., Лукьянова И.В., 1976. География позвоночных южной тайги Западной Сибири. Новосибирск: Наука. 359 с.
Сапогов А.В., 1983. Зональные особенности населения мышевидных грызунов Енисейской тайги // Животный мир Енисейской тайги и лесотундры и природная зональность. М.: Наука. С. 204–214.
Стариков В.П., 1985. Пространственная структура населения мелких млекопитающих. Лесостепная и лесная зоны Западной Сибири // Пространственно-временная динамика животного населения. Новосибирск: Наука. С. 176–185.
Трофимов В.А., 1976. Модели и методы качественного и факторного анализа матрицы связи // Проблемы анализа дискретной информации. Новосибирск: Наука. Ч. 2. С. 24–36.
Цыбулин С.М., Богомолова И.Н., 2012. Межгодовая изменчивость населения мелких млекопитающих южной тайги Приобья // Актуальные проблемы современной териологии. Новосибирск: ООО “Сибрегион Инфо”. С. 20.
Jaccard P., 1902. Louis de distribution florale dans la zone alpine // Bulletin de la Société vaudoise des sciences naturelles. V. 38. P. 69–130.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал