

Зоологический журнал, 2019, T. 98, № 4, стр. 420-436
Экология урбанизированной популяции тихоокеанской чайки (Larus schistisagus) в сравнении с естественными колониями. 1. Особенности размещения гнезд и продуктивность
Л. А. Зеленская *
Институт биологических проблем Севера ДВО РАН
685000 Магадан, Россия
* E-mail: larusrissa@gmail.com
Поступила в редакцию 20.03.2018
После доработки 20.05.2018
Принята к публикации 12.07.2018
Аннотация
В первой (из серии статей) работе проанализированы результаты четырнадцатилетнего (2004–2017 гг.) мониторинга тихоокеанской чайки, гнездящейся на крышах г. Магадана. Выявлен ряд особенностей, характерных для гнездовий одной из первых урбанизированных популяций чаек на северо-востоке Азии. У городских чаек, в отличие от чаек природных гнездовий, преобладают поселения одиночных пар, но основная масса гнезд сосредоточена в нескольких крупных колониях. При выборе города как места для размножения чайки с годами все дальше уходят от морского побережья. Расселение пар по городу, как правило, происходит в пределах видимости от старой колонии. Темпы роста колоний составляют в среднем 13% в год, аналогично городским поселениям чаек в Европе. Продуктивность тихоокеанских чаек, гнездящихся на городских крышах, значительно выше, чем в природных колониях. В урбанизированной популяции не отмечены гибель яиц от внутривидового хищничества и гибель птенцов от чаек-соседей. Большинство гнезд урбанизированных чаек строится на одном и том же месте многие годы. Предназначение здания определяет степень посещаемости его кровли людьми, что имеет решающее значение при выборе чайками крыши для строительства гнезд. Как правило, гнезда строятся на крыше около любой преграды (стены, трубы, надстройки и проч.) либо на совершенно открытом месте. Второе более характерно для одиночных пар. Безопасность крыш от хищников, вероятно, является решающим фактором, способствующим урбанизации чаек.
История “освоения” городов разными видами чаек с начала прошлого века приводится во многих сводках (Cramp, 1971; Vermeer et al., 1988; Raven, Coulson, 1997; Rock, 2005; Kubetzki, Garthe, 2007; Soldatini et al., 2008). Размножение чаек в городах вызывает много проблем: шум, мусор и экскременты на крышах, припаркованных автомобилях и улицах, агрессивное поведение по отношению к людям, угроза безопасности самолетов в районах аэропортов и т.д. Рок (Rock, 2005), который более 25 лет проводит мониторинг городской популяции чаек Бристоля и изучает проблемы, связанные с городскими гнездовьями чаек, отмечал, что для лучшего понимания реакции птиц на то или иное воздействие людей и для выбора методов управления их поведением очень важно знать особенности экологии именно урбанизированных популяций чаек. В то же время, основные данные по экологии чаек получены в естественных колониях, потому исследования городских популяций остаются крайне актуальными.
Тихоокеанская чайка (Larus schistisagus), гнездовой ареал которой охватывает все побережье Охотского моря, Камчатки и Приморья, Курильские острова и побережье о-ва Хоккайдо с прилежащими островками, до недавнего времени избегала поселений людей. Например, отмечалось, что чайки не гнездятся в г. Владивостоке, хотя вне города, на о-ве Карамзина гнездятся ежегодно в природных колониях с 1967 г. (Назаров, 2004). В г. Куширо (Хоккайдо) тихоокеанская чайка начала гнездиться на крышах во второй половине 1980-х годов (Артюхин, 2002). Примерно в этот же период чайки освоили крыши в г. Саппоро (Хоккайдо), но информация об этом появлялась только в местных газетах (Hiroyoshi Higuchi, личное сообщение). В 1998 г. отмечены первые гнезда тихоокеанской чайки на крышах г. Магадана (Зеленская, 2004). С 2013 г. тихоокеанская чайка гнездится на крышах пос. Южно-Курильск (о-в Кунашир, Южные Курилы) (Зеленская, 2016). На Камчатке тихоокеанская чайка пока не гнездится в городах, но начинает осваивать брошенные поселки на морском побережье (Артюхин, Вяткин, 2012).
В работе представлены результаты четырнадцатилетнего (2004–2017 гг.) мониторинга популяции тихоокеанской чайки, гнездящейся на крышах г. Магадана. Целью данной работы было выявление особенностей экологии городской популяции тихоокеанской чайки, способствующих успешному освоению новой среды гнездования.
МАТЕРИАЛ И МЕТОДЫ
Мониторинг гнездовой популяции начался в 2004 г. и проводился ежегодно за исключением 2007, 2011 и 2012 гг. В 2015 г. были осмотрены только колонии. В период окончания строительства гнезд и начала инкубации (25 мая–10 июня) мы обследовали все крыши города на наличие гнезд чаек, проверяя квартал за кварталом. Второй осмотр зданий проводили в конце лета (25 июля–5 августа) для учета количества слетков и выяснения успеха размножения чаек. Мы оценивали продуктивность – отношение числа слетков к количеству пар птиц, приступивших к строительству гнезд. Отдельно проводили расчеты для одиночных пар, маленьких и крупных колоний. Второй учет не был проведен в 2008, 2013–2014 гг. Осмотр проводили с земли, если крыша была скатной или если она хорошо просматривалась из-за неровности ландшафта (город расположен на склонах сопок). Также крыши осматривали в 8-кратный бинокль с соседних, более высоких зданий. Некоторые колонии осматривали, непосредственно поднимаясь к ним на крышу.
Поселение, т.е. население крыши одного здания, может состоять как из одной пары чаек, так и из группы пар. Если на одном здании гнездились 1–2 пары чаек, мы считали такое поселение одиночным, т.к. гнезда располагались на максимально большом расстоянии друг от друга. Маленькой колонией мы считали поселение на одном здании 3–9 пар. Большая колония содержала 10 и более гнезд. Формальное разделение размеров колоний мы считаем правомерным, основываясь на утверждении Харитонова (2011), что понятие “колония” принципиально не точно. Колонией можно назвать любую пространственно-обособленную группировку гнезд, при этом внутри данной группировки могут присутствовать и группировки более низкого ранга. Из принципиальной неточности определения колонии следует очень важный методологический момент: мы можем рассматривать любую концентрацию гнезд как колонию, не оглядываясь на наличие группировок в пределах этой концентрации, а сама названная “колонией” группировка может являться частью группировки более высокого ранга (Харитонов, 2006).
Обнаружив гнездо/колонию на крыше, фиксировали следующие параметры здания: высоту (количество этажей), тип крыши (плоская, скатная), материал кровли, предназначение строения. Здания по их предназначению условно разделяли на следующие категории: 1 – жилое, 2 – детское (детские сады, школы, интернаты) или учебное (ПТУ, техникум, университет и их общежития), 3 – лечебное (больницы, поликлиники, диспансеры), 4 – административное (офисы, библиотеки, музей, магазины, церкви и проч.), 5 – промышленное (склады, цеха, корпуса и башни заводов). Учитывали точное местоположение гнезда на крыше. Для колонии составляли план крыши, с указанием расположения всех труб, надстроек и проч., иногда с помощью космоснимков Google, позволяющих повысить точность изображения. Далее пользовались готовым бланком, отмечая на нем гнезда текущего сезона.
Чтобы выяснить, есть ли предпочтение у тихоокеанской чайки заселять здания с определенным типом крыши и определенной высоты, на семи участках города были учтены все здания. Участки были выбраны формально: они были ограничены трех–четырехполосными автомагистралями и не были равными ни по площади, ни по числу зданий в них. Эти участки охватывали почти все районы города, в которых отмечены гнезда чаек, и могли представлять выборку для города в целом. Для каждого здания в каждом участке учитывали: есть ли на крыше гнезда чаек (независимо: колония или одиночное гнездо), характер крыши (плоская, скатная), количество этажей. Анализ проведен стандартными методами (использован критерий χ 2).
РЕЗУЛЬТАТЫ
История заселения города
Город Магадан занимает возвышенный перешеек п-ова Старицкого между бухтами Нагаева и Гертнера (рис. 1). Обе бухты – часть Тауйской губы в северной части Охотского моря. В бухту Гертнера впадает р. Магаданка, долина которой около моря заболочена. Река ограничивает городскую застройку.
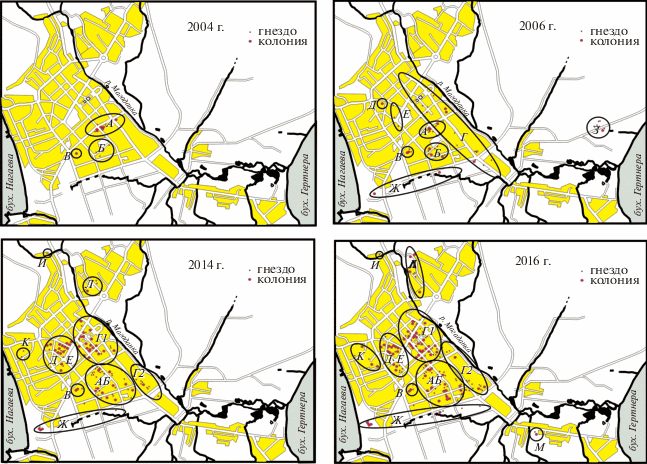
Заселение города чайками началось с центральной части. В 2004 г. в городе было три удаленных друг от друга района гнездования чаек (рис. 1, 2004 г.): А – вдоль ул. Якутская, где она поднимается от р. Магаданки. Это были 5 крупных колоний на административных (библиотека, музей, церковь) и учебных зданиях (школа и общежитие политехникума) и отдельные гнезда на соседних жилых домах. Все гнездовья в этом районе находились в зоне видимости друг друга. Б – удаленная примерно на 1 км в сторону бухты Гертнера и закрытая парком группа одиночных гнезд была построена на крышах зданий поликлиники, политехнического института и на жилом 9-этажном доме. В – колония на здании школы-интерната, изолированная и удаленная от второго участка на 1 км, почти на вершине склона.
На следующий год на участке А насчитывалось 11 колоний. К одиночным гнездам присоединились новые пары и новые одиночные пары появились на других крышах в зоне видимости друг друга. Колония на участке Б росла в геометрической прогрессии, на соседних жилых домах появились одиночные гнезда. Та же картина была и на участке В, где сформировались две маленькие колонии. Кроме того, маленькие колонии и одиночные гнезда появились в “спальном” районе вдоль реки (Г) (рис. 1, 2006г.). Одиночные гнезда были найдены еще в двух, визуально не связанных с существующими гнездовьями местах на вершине сопки –Д и Е, но через перевал к бухте Нагаева чайки не перешли.
Шумные колонии на участке А почти все подверглись разрушению гнезд летом 2005 и весной 2006 гг., и, вероятно, птицы для гнездования переселились на участки Г и Б. Численность поселений и число гнезд в них на этих участках быстро росли. В 2006 г. появились первые небольшие колонии в промзоне вдоль края городской застройки (Ж) (рис. 1, 2006 г.). Крайняя колония была расположена практически над бухтой Нагаева на гигантском законсервированном цехе. Доступ людей на крышу цеха (300 × 300 м, высота около 15 м, 500 м от моря) был невозможен (все лестницы срезаны почти на уровне крыши). В бухте Гертнера в 2006 г. на руинах зверофермы (полуразобранные шеды, бетонные основания разрушенных построек, высота их от 2 до 5 м, 1 км от моря) появились колония и одиночные гнезда вокруг нее (З, рис. 1, 2006 г.). Разбор руин положил конец всем поселениям чаек на участке З через 3 года.
В городе в период наблюдений происходил снос старых бараков, капитальный ремонт кровли школ. С 2009 г. активизировалась реконструкция крыш жилого фонда (замена шифера на металл). Гнезда чаек при этом уничтожались, но рост численности гнездящихся чаек продолжался. Происходило перераспределение гнездовий внутри уже обжитых птицами районов. В 2008 г. стал единым район, где чайки гнездились на крышах застройки 70-х годов в центре (А–Б), а к 2014 г. – объединились районы исторического центра (“сталинки”) и верхушка сопки с “больничным городком” (Д–Е). А район гнездования вдоль реки к 2010 г. разделился по типу его заселения чайками на две области – административный центр, плотно заселенный в основном колониями (Г1) и спальные районы, заселенные в основном одиночными парами (Г2). В промзоне (Ж) одиночные гнезда и колонии то появлялись, то разорялись, была стабильной только колония над бухтой Нагаева (рис. 1, 2014 г.).
Периодически появлялись и затем либо развивались, либо разорялись поселения чаек в других районах города. В 2009 г. появились колония и одиночные гнезда вокруг на крышах здания администрации ТЭЦ и окружающих станцию реконструируемых зданиях (7 км от моря, 4–5 этажей). Через несколько лет после смены кровли и окончания ремонта осталось 2 гнезда на одной крыше (И). В микрорайоне “Нагаево” в 2013 г. появилось первое гнездо на школе (0.5 км от моря) (К). В 2014 г. на другой стороне р. Магаданка, в ее широкой долине была отмечена колония чаек на складе таможни и одиночные гнезда вокруг нее (Л) (рис. 1, 2014 г.).
В настоящее время центр города заселен чайками наиболее плотно, и численность их гнезд продолжает расти. Ранее относительно изолированные поселения чаек в разных районах города в настоящий момент объединяются. Вероятно, через несколько лет в центре будет единая зона гнездования чаек (А–Б–Г1–Г2–Д–Е) (рис. 1, 2016 г.). Если ориентироваться по рельефу местности, почти все гнезда чаек сосредоточены на склоне сопки восточной экспозиции, открывающейся в долину р. Магаданки. Гнезда на крышах домов, расположенных на противоположном склоне сопки, открытой в сторону бухты Нагаева, известны давно. Но именно этот склон наиболее неохотно используется чайками (исключение – колония в промзоне). В последние годы началось освоение чайками другого берега реки, где кроме крупной колонии на складе таможни сформировались несколько колоний на зданиях жилого фонда. В пределах видимости вокруг них гнездятся одиночные пары (Л). В 2016 г. зарегистрирована крупная колония на брошенном цехе-бараке в колонии строгого режима (2 этажа, 4 км от моря) (М) (рис. 1, 2016 г.). На охраняемой территории без беспокойства со стороны людей рост этой колонии, вероятно, будет стремительным. Так, в 2017 г. эта колония уже увеличилась на треть. За весь период наблюдений ни разу не отмечены гнезда на строениях в районах рыбного и торгового портов на берегах бухты Нагаева. Численность урбанизированной популяции на крышах г. Магадана в 2017 г. составляла: 1302 гнезда в колониях и 139 одиночных гнезд.
Можно отметить постепенное удаление гнездовий чаек от морского побережья. В 2004 г. гнезда чаек строились на расстоянии около 5 км от бухты Гертнера, куда впадает р. Магаданка. В 2009 г. их гнезда впервые появились в районе ТЭЦ (6.8 км от бухты Гертнера). В 2017 г. гнезда чаек впервые найдены в микрорайоне “Пионерный” (7.9 км от бухты Гертнера).
Особенности городских поселений
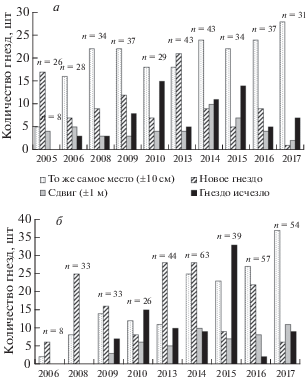
За весь период наблюдений количество одиночных поселений (т.е. одиночных гнезд) всегда было максимальным, а количество крупных колоний – всегда невелико и рост их численности был низким (рис. 2а). При этом именно большие колонии содержали основную массу гнезд чаек (рис. 2б). Одиночные поселения самые нестабильные. В течение 14 лет наблюдений ежегодно на новых местах появлялось в среднем 28 одиночных гнезд (от 16 до 44). Из них в среднем 9 гнезд (от 0 до 22) уже на следующий год исчезают навсегда. При этом одиночные гнезда могут существовать многие годы (до 11 лет). Иногда одиночные пары пропускали какой-то сезон, но на следующий год снова строили гнездо.
Рис. 2.
Количество поселений (занятых зданий) одиночных пар и колоний разной величины (а) и численность гнезд в них (б) в период наблюдений.
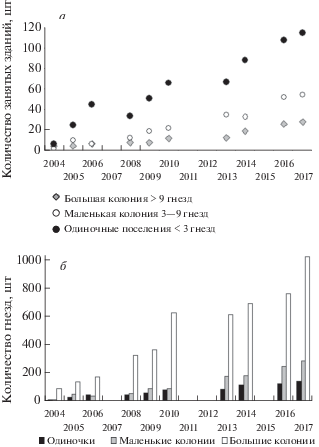
Статус поселения может быстро меняться. Через год-два одиночное поселение становилось маленькой колонией. Затем либо маленькая колония стремительно превращается в большую, либо ее рост приостанавливается на несколько лет. Не редок и обратный процесс, когда на месте крупной колонии оставались одна-две пары. Обычно это связано с ремонтом и сменой кровли крыш, а также с планомерным уничтожением колонии.
Темпы роста быстрорастущих колоний, не затронутых действиями людей, укладываются в следующую грубую схему. В первый год темпы роста максимальны – в среднем 72.3% в год (57.7–85.7%, n = 10) – это период превращения одиночного поселения в маленькую колонию. Затем темпы роста снижаются, в среднем до 40.5% в год (29–47.7%, n = 10) – это период превращения маленькой колонии в большую и первые годы роста численности в большой колонии. Большая колония растет относительно стабильно, пока хватает места на крыше. Дальнейший рост идет за счет оккупации смежных крыш. Темпы роста остаются в среднем 13% в год (7.4–23.8%, n = 11).
Однако формирование и рост большинства колоний, не затронутых действиями людей, протекают значительно медленнее (n = 13). Происходит это за счет того, что стадия одиночного гнездования продолжается 2–3 года, затем 3–8 лет (в среднем 5 лет) существует маленькая колония, которая, наконец, увеличивается до большой колонии. Существуют исключения. Так, колония, расположенная на огромной плоской крыше промышленного объекта, увеличилась за 2 года с 20 до 130 гнезд, еще за 2 года до 340 гнезд, а далее оставалась на этом уровне несколько лет, хотя гнезда занимали не более 2/3 площади крыши. Полностью крыша была занята только через 11 лет.
По нашим наблюдениям, в городских условиях (как в г. Магадан, так и в пос. Южно-Курильск) тихоокеанские чайки строят гнездо из сухой травы, собранной на газонах. В выстилку также могут попадать куски мха с почвой, найденные на этой же крыше, а также валяющийся рядом на крыше мусор. Объем мусора в выстилке гнезд был крайне индивидуален. Подавляющее большинство гнезд его не содержало. Аналогичное поведение отмечено нами у этого вида и при строительстве гнезд в природных колониях.
Выбор здания для гнездования
Предназначение здания определяет степень посещаемости его кровли людьми, что имеет важнейшее значение для чаек. Как правило, полностью исключают выход людей на крышу во избежание несчастных случаев в детских, учебных и лечебных заведениях. Крыши промышленных объектов, которые находятся под постоянной охраной, люди посещают так же редко. Редко люди выходят и на крыши административных зданий. Крыши жилых многоквартирных домов посещаются людьми беспорядочно.
Большинство зданий с гнездами чаек – жилые многоквартирные дома, они составляют основную массу городской застройки. Однако общая численность гнезд, построенных на жилых домах, изначально была не высока. Только в последние несколько лет в связи с продолжающейся экспансией чаек в “спальные” районы города, жилые здания сделались лидерами по численности гнезд.
Второй категорией зданий по частоте использования чайками являются детские и учебные заведения. Они же были “постоянными лидерами” по числу гнезд, построенных на их крышах до 2009 г, а позже входили в “первую тройку”. Административные здания, начиная с 2014 г., являются следующими по частоте заселения их чайками. Но численность гнезд на них всегда была невысока. Число лечебных заведений и промышленных объектов, используемых чайками, невелико. Но именно промышленные объекты поддерживают наибольшую численность гнезд в последние годы. Самая крупная колония, содержащая более 340 гнезд, находится на крыше брошенного цеха.
Большинство плоских крыш в городе покрыты рубероидом, а большинство скатных крыш – шифером. Начиная с 2009 г. в историческом центре, а позже и в других районах шиферные крыши массово перекрывают металлом. При этом здания жилого сектора (за исключением исторического центра) и промышленные объекты, как правило, имеют плоские крыши. Исторический центр застроен зданиями со скатными крышами с множеством “архитектурных излишеств”.
В 2016 г. анализ застройки в центральной части города показал, что она крайне неоднородна. В исследуемом районе было зафиксировано 357 зданий с плоской крышей и 181 здание со скатной. Соотношение домов с плоскими и скатными крышами занятых чайками в разных участках изменялось от 23 до 100%. Различия были статистически достоверны (χ2 = 17.09, df = 6 на скатных крышах; χ2 = 14.49, df = 6 на плоских крышах). Таким образом, подтвердилось предположение, что при выборе здания у чаек нет определенных предпочтений при выборе типа крыши и кровельного покрытия. Распределение гнезд по крышам с разным типом кровли отражает лишь особенности застройки (плоские или скатные) и реконструкцию городских строений (замена шифера металлом). Более того, чайки, гнездившиеся на шиферных крышах, после замены кровли на металлочерепицу, продолжали строить гнезда на новой поверхности, если их гнезда больше не тревожили.
Без учета характера крыши суммарно на всех 7 участках были отмечены следующие факты. Ни на одном из одноэтажных домов (46 зданий) не было обнаружено гнезд чаек. Из 48 двухэтажных зданий было заселено всего 2 или 1%. Из 53 трехэтажных зданий заселено 41.6%; из 89 четырехэтажных – 52.9%; из 273 пятиэтажных – 18.7%; из 31 зданий в шесть этажей и выше – 42%. Чайки избегали гнездиться на одноэтажных зданиях. Охотно селились на трех–четырехэтажных и на высоких зданиях (в 6, 7, 9 этажей). Менее охотно гнездились на двух– и пятиэтажных зданиях. При этом почти все спальные районы г. Магадана застроены пятиэтажными домами с плоской крышей, а внутри кварталов старой застройки преобладают двух–трехэтажные дома со скатными крышами.
Выбор места для гнездового участка и расположения гнезда на крыше
Чаще всего гнездо было расположено рядом с преградой (за трубой, около стены, около любого оставленного на крыше крупного предмета). Гнездо на открытом месте (более 50 см от любого объекта или постройки на крыше) чайки строят не часто. Но при этом они охотно занимают совершенно открытую самую высокую точку на крыше (на широком коньке) или любую надстройку на плоской крышке (крыша балкона, труба, будка, слуховые окна и проч.). Чайки редко строят гнезда под навесом или между ящиками и прочими преградами. Расположение гнезд в желобах, откуда есть хороший обзор, встречается часто в колониях на крышах со скатом. Мы приводим суммарную картину для колониальных и одиночных пар за весь период наблюдений, т.к. мы не нашли значительных изменений по годам (рис. 3).
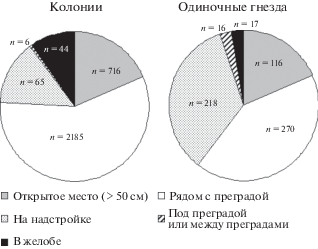
Отмеченные нами различия в предпочтениях чаек, гнездящихся одиночно и колониально (рис. 3), скорее всего, отражают то, что одиночные пары более осторожны и помимо защитной стены предпочитают места с хорошим обзором, т.к. не имеют возможности, как колониальные птицы, воспользоваться предупреждением об опасности от чаек-соседей.
Подробное картирование гнезд в колониях позволило выявить факт, что большая часть гнезд на крышах, как правило, из года в год размещается на одном и том же месте (±10 см относительно прошлого сезона). Как правило, к началу нового сезона размножения от прошлогодних гнезд на кровле нет и следов, остатки гнездового материала зимние ветра сбрасывают на землю. Только модернизация крыши заставляет чаек переместить место гнезда. Так, после 9 лет ежегодного успешного размножения одиночное гнездо было перенесено на 4 м после установки на старом месте антенн мобильной связи. В колониях отдельные гнезда фиксировались нами на том же самом месте на протяжении 12 лет. Одно из гнезд (отмечалось ежегодно 9 лет) перестали строить на том же самом месте только после уничтожения надстройки, на которой птицы его воздвигали.
Как модельные площадки были выбраны две колонии, подвергавшиеся минимальному воздействию людей. Обе колонии прослеживались одновременно (2004–2017 гг.) и размещались на зданиях общежития политехникума (рис. 4) и школы-интерната (рис. 5). Каждое здание состояло из двух корпусов. Сначала обе колонии занимали один корпус (“центр”), потом, разрастаясь, заняли и крышу соседнего корпуса (“периферия”). Колонии расположены в разных районах города, на крышах разного типа (общежитие имеет плоскую крышу, покрытую рубероидом, интернат – скатную крышу, покрытую шифером).
Рис. 4.
Число и статус гнезд (объяснение в тексте) в “центре” (а) и на “периферии” (б) колонии на плоских крышах общежития в течение периода наблюдений. n – общее количество гнезд на данном участке колонии.
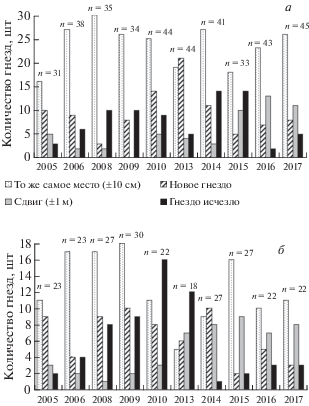
Рис. 5.
Число и статус гнезд (объяснение в тексте) в “центре” (а) и на “периферии” (б) колонии на скатных крышах интерната в течение периода наблюдений. n – общее количество гнезд на данном участке колонии.
Большинство гнезд в этих колониях “стабильные”, они строятся на одном и том же месте из года в год. Небольшая часть гнезд в следующем сезоне строится со сдвигом на расстояние примерно до 1 м относительно местоположения прошлого сезона. Обычно этот “сдвиг” вызывается появлением гнезда новой пары, из-за которого “приходится отодвигаться”, а затем место гнезда становится постоянным. Либо в течение 2–3 сезонов гнездо “поднимается” по склону крыши, пока не займет “идеальное положение”, после чего место гнезда становится постоянным. Часть гнезд в конкретном сезоне – “новые”, т.е. построены впервые на данном месте. “Новым” гнездо может быть только единственный сезон. На следующий сезон гнездо либо строится на том же самом месте, либо передвигается немного в сторону/выше, либо это место чайками больше не используется. В последнем случае мы считаем гнездо “исчезнувшим”. Отражая расположения гнезд на рис. 4 и 5, мы добавили информацию о гнездах, которые “исчезли”, т.е. их нет в данном сезоне.
В обеих колониях в “центрах” соотношение гнезд разного статуса (“стабильных”, “новых”, “сдвинутых”) сходно (рис. 4а, 5а). Возможно, в благополучных колониях, в давно заселенном “центре”, состав гнездящихся пар более постоянен. Если происходят изменения, они отражают общегородскую ситуацию с распределением гнезд чаек. Резкое возрастание иммигрантов в 2013 г. отмечено в обеих колониях. Возможно, это связано с массовой плановой заменой кровельного покрытия на зданиях жилого фонда города в тот период. Согнанные с ремонтируемых крыш пары чаек переселялись в другие колонии. По распределению гнезд периферии колоний отличались от “центров” и рзличались между собой (рис. 4б, 5б). Вероятно, здесь отражаются процессы, происходящие именно в конкретной колонии и ее окрестности.
Продуктивность
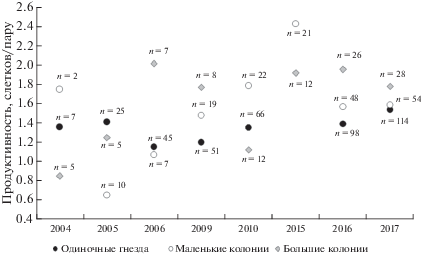
Продуктивность гнездящихся на крышах г. Магадана тихоокеанских чаек только в первые годы наблюдений была менее 1 птенца на пару (рис. 6). В г. Магадане мы ни разу не видели ограбления/расклевывания кладок, оставленных слетевшими при нашем приближении чайками. За все годы мониторинга в городе была найдена единственная погадка (n = 494) со скорлупой конспецифичного яйца. Наблюдения показали, что и гибель птенцов в городе очень редка. Мы не отмечали на головах или шеях птенцов на городских крышах ран от клювов чаек-соседей.
Рис. 6.
Продуктивность гнездящихся на крышах тихоокеанских чаек в разных поселениях в период мониторинга. n – количество занятых гнездами крыш.
На крыши г. Магадана не проникают хищные млекопитающие, ни дикие, ни домашние. В городе ни разу не отмечали таких пернатых хищников, как белоплечий (Haliaeetus pelagicus) и белохвостый (H. albicilla) орланы, которые гнездятся в окрестностях города. По нашим многолетним наблюдениям в природных колониях тихоокеанской чайки, как в Северном Охотоморье, так и на Камчатке, только при появлении орланов наблюдается массовая коллективная защита гнезд. Охотящийся на городских голубей (Columba livia) сапсан (Falco peregrinus) не вызывает защитных реакций у гнездящихся на крышах чаек. Слабую агрессию вызывают только вóроны (Corvus corax), редко появляющиеся над гнездовыми участками чаек в городе.
Основная причина разорения гнезд и гибели чаек в городе – действия людей. В начале формирования городской популяции чаек (первые годы наблюдений) больше всего страдали от человека именно колонии, как крупные, так и маленькие (рис. 6). Рост колоний и стабильность размножения чаек в них определялись желанием людей уничтожить конкретную колонию и их настойчивостью или полным равнодушием к ее существованию. Методом “проб и ошибок” чайки определяли безопасные крыши для колонии.
Разорение одиночных гнезд обычно происходит случайно, попутно с ремонтными работами и/или установкой антенн. Наиболее обычным вариантом разорения колоний является плановый ремонт крыши. При этом уничтожаются все гнезда с кладками. Но уже на следующий сезон чайки начинают пробовать вновь заселять реконструированную крышу. Второй вариант – уничтожение колонии по указанию владельцев крыши из-за проблем, связанных с жизнедеятельностью чаек. Для этих целей опробовались различные методы. Единственный раз применялась специальная техника – пожарная машина, для уничтожения колонии на здании областной библиотеки в 2004 г. Одна из самых старых и крупных колоний в городе в течение весны 2010 г. подвергалась обстрелу из воздушного ружья по собственной инициативе одного из жителей соседнего более высокого дома. Были отстреляны, ранены и впоследствии погибли около двух десятков взрослых птиц. Отстрелы были прекращены силами других жителей окрестных домов, которым нравились чайки. Колония на данной крыше продолжает существовать и поныне.
В период, когда птенцы становятся на крыло и начинают подлетывать, ежегодно отмечается падение отдельных птенцов на землю, когда они пытаются убежать/улететь от людей, поднявшихся на крышу. На улицах/на земле родители не могут защитить птенцов, они гибнут под колесами автотранспорта, от нападения собак и кошек. Но доля погибших слетков-неудачников очень мала.
Самыми стабильными по продуктивности все годы наблюдений в Магадане были поселения отдельных пар (рис. 6). Одиночные пары после сбора яиц людьми обычно бросают гнездовой участок, но разорение одиночных пар не частое явление. Если гнезда одиночных пар не разоряются, они, как правило, поднимают на крыло всех птенцов. В последние три года наблюдений средняя продуктивность городских чаек была выше 1.4 птенца на пару во всех типах поселений (рис. 6).
ОБСУЖДЕНИЕ
Выбор города как места гнездования
Решающими могут быть два фактора – питание и безопасность. Питание – тема для отдельного следующего сообщения. Первостепенность безопасности для размножения подчеркивает тот факт, что крупные природные колонии тихоокеанских чаек существуют на недоступных для наземных хищников островах и скальных участках побережья. В г. Магадан гнездящийся на земле вид сразу же стал строить гнезда на крышах. Не было ни одной попытки строительства гнезд на земле. Это вполне соответствует выводам специальных исследований, показавшим, что и в городе гнездование на земле не безопасно. При этом хищничество по отношению к гнездам на земле в центре и в жилой застройке города происходит по-разному, и оно выше, чем в окрестности города (Jokimaki et al., 2005). Монаган (1979) предположила, что для чаек здания в городах являются искусственными “островами в море бетона”, и крыши имеют многие из признаков, предоставляемых естественными гнездовьями.
Безопасность крыш от наземных хищников как решающий фактор, расширяющий урбанизацию чаек, отмечали и другие исследователи (Vermeer et all., 1988; Raven, Coulson, 1997; Виксне, 2006). Это подчеркнуто в специальном исследовании сизой чайки (L. canus), численность которой в естественных местообитаниях на Балтийском побережье в Германии с 70-х годов прошлого столетия существенно уменьшилась. Увеличение численности чаек в городах, по мнению авторов, есть ответ на хищничество куницы, норки и лисы, численность которых увеличилась. Этому соответствует тот факт, что вдоль побережья Северного моря, где пресс хищников еще низок, нет гнездования сизой чайки на крышах, и продолжается рост численности птиц в естественных популяциях (Kubetzki, Garthe, 2007).
Давно отмечено, что выбрав город для гнездования, чайки уходят от морского побережья во внутренние области. Этот процесс растянут во времени и дистанция от берега варьирует для каждого вида чаек. Так, серокрылая чайка (L. glaucescens) в Сиэтле (США) в 1960 гнездилась на расстоянии до 320 м от моря, в 1981 – на расстоянии в 1 км (Hooper, 1988). Гнездовые колонии серебристой чайки (L. argentatus) и клуши (L. fuscus) на крышах в Англии в 1969 г. встречались на расстоянии не далее 8 км от побережья, а в 1976 г. – в 25 и 56 км для серебристой чайки и клуши, соответственно (Monaghan, Coulson, 1977). Постепенное увеличение со временем дистанции от морского побережья до гнезд мы наблюдали для тихоокеанской чайки в г. Магадане. Для естественных популяций тихоокеанской чайки гнездование даже в нескольких километрах от моря является отклонением от поведенческих стереотипов. Каждый такой случай привлекает внимание исследователей. Только две природные популяции тихоокеанской чайки гнездятся в 40 км от моря на крупнейших пресноводных озерах Камчатки (Лобков, 1986; Зеленская, 2014, 2017). Как исключительно редкое явление отмечено гнездование одиночной пары в 3 км от моря на о-ве Хоккайдо (Япония) (Mazda, Higuchi, 1976) и четырех уединенных пар тихоокеанской чайки на Камчатке на искусственных отстойных озерах в 90–100 км от моря (Лобков, Карпухин, 2010).
Первые гнезда в г. Магадане найдены в центре города. Возможно, здесь отражается тенденция, отмеченная сначала в Англии, что чайки располагают гнезда первоначально на относительно высоких и безопасных зданиях центра города (Monaghan, 1979; Raven, Coulson, 1997; Coulson, Coulson, 2009).
Особенности городских поселений
Изучая экологию тихоокеанской чайки в природных условиях около 30 лет, мы исключительно редко наблюдали поселение этого вида отдельными парами. Чаще всего тихоокеанская чайка гнездится небольшими колониями, насчитывающими, как правило, несколько десятков пар. Крупные колонии (250 пар и более) встречаются не часто. Это характерно как для Камчатки (Фирсова и др., 1982), так и для северного Охотоморья (Голубова, Плещенко, 1997). В то же время в городе у тихоокеанской чайки преобладают поселения именно одиночных пар и маленькие колонии. Крайне разреженное гнездование в природных условиях у тихоокеанской чайки отмечено только на краю болот в кочковатой тундре на крайнем юге Корякского нагорья, где минимальное расстояние между гнездами было 100 м. На юго-восточном побережье Камчатки гнезда на болотах были найдены в низовьях и устьях только трех рек (Лобков, 1981). Одиночные гнезда отмечены недавно на выброшенных морем и полузатопленных судах в Олюторском заливе на Камчатке. При этом в заливе обнаружены и мелкие естественные колонии (Артюхин, Вяткин, 2012).
На крышах городов плотность размещения гнезд чаек, независимо от вида чаек, ниже, чем в природных колониях (Monaghan, 1979; Vermeer et al., 1988; Hooper, 1988; Belant, 1993; Soldatini et al., 2009). Для видов, чьи гнезда являются объектом нападения хищников, сравнимых по силе с гнездящимися особями, выгодна либо очень малая плотность гнездования, когда хищник редко находит гнезда (Burger, 1984; Харитонов, 2006), либо сразу высокая плотность, когда хищник не может разорить больше определенного предела (Харитонов, 2006), либо чайки в крупных колониях нападают на хищника совместно (Burger, 1974). Если хищником является человек, успех размножения выше именно в редко распределенных гнездах – как отмечено в природных колониях, подвергающихся разорению людьми у желтоногой чайки (L. livens) (Spear, Anderson, 1989), так и в городе у средиземноморской чайки (L. michahellis) (Soldatini et al., 2008).
Как другую гипотезу, объясняющую массовое появление одиночного гнездования в городе, можно рассматривать городскую среду обитания как нестабильную, в которой человек может разорять гнезда также непредсказуемо, как происходят колебания уровня воды в естественных местах гнездования чаек в болотах и низинах. В таком случае можно видеть прямую аналогию с выбором мест гнездования у городских чаек Магадана и серебристой чайки (L. mongolicus) в дельте р. Селенга, описанную Пыжьяновым (1985). В дельте Селенги основная часть колоний (численность 11–30 пар) занимает выбранный участок 3–5 лет подряд, и только отдельные крупные колонии (от 100 пар и более) сохраняются на одном месте до 10 лет и более (Мельников, 2014). Из-за колебаний уровня воды, что является основной причиной гибели кладок у чаек, птицы выбирают места для гнездования методом “проб и ошибок”. При этом частая гибель гнезд и способность птиц повторять кладки приводят к тому, что в одних случаях чайки подселяются к “благополучным” колониям, в других – образуют небольшие поселения в новых местах. Стремясь использовать особенности микрорельефа, чайки нередко расселяются на очень большой территории. Расстояния между гнездами составляют десятки и даже сотни метров, в то время как на самых разреженных скальных колониях они не превышают 10–15 м (Пыжьянов, 1985). Возможно, деятельность человека по отношению к гнездовьям чаек в городе воздействуют на популяцию птиц аналогично неуправляемой стихии и рассеянное распределение гнезд на крышах города можно предполагать адаптацией к гнездованию в условиях нестабильной среды и воздействия человека как основного хищника.
Рост и развитие колоний чаек в г. Магадан (ряд: одиночное гнездование–маленькая колония–большая колония) происходит так же, как в Европе у других видов крупных чаек. В Англии поселение из 1–2 пар может легко исчезнуть, но при повышении их численности колония уже не исчезнет без влияния человека (Raven, Coulson, 1997).
Рост численности чаек в колониях на крышах в Англии – 17% в год. (Monaghan, 1979). Факт иммиграции подтверждается окольцованными в природной колонии птицами, размножающимися затем на крышах в 180–200 км от их родной колонии (Monaghan, Coulson, 1977). Аналогичные данные получены в Италии для средиземноморской чайки. Рост численности городской популяции в первое десятилетие в г. Триесте составил 46.6% в год, в следующие 5 лет – 31.3%. Такой высокий уровень роста явно включает иммиграцию, наиболее вероятный источник которой – ближайшие естественные колонии (Soldatini et al., 2008). Предполагается, что расширение городских колоний есть результат насыщенности или почти насыщенности природных колоний, откуда идет иммиграция (Monaghan, Coulson, 1977). Вероятно, в Магадане увеличение численности городской популяции также происходит за счет иммиграции из природных колоний в окрестностях города.
Появление новых гнездовий в Магадане, как правило, происходило в пределах видимости от старой колонии. Эта черта характерна для естественных колоний разных видов крупных чаек на островах без хищников-млекопитающих. Так, у чернохвостой чайки (L. crassirostris) новая колония начиналась с одиночного гнезда на участке, откуда хорошо видна основная колония, и новые участники добавлялись постепенно (Литвиненко, 1980). Ту же картину мы наблюдали у тихоокеанской чайки в Тауйской губе: на о-вах Шеликан, Умара, островных барах Ольской лагуны. Тенденцию гнездиться рядом с конспецифичными соседями отмечали у урбанизированных серокрылых чаек (Vermeer et al., 1988), клуш и серебристых чаек, гнездящихся в Англии (Monaghan, 1979) и Шотландии (Coulson, Coulson, 2009), где одиночные гнезда располагались в пределах 100 м друг от друга. По мнению исследователей, такое размещение гнезд в городе демонстрирует наличие социальных взаимоотношений между гнездовыми парами и колониальную природу гнездования. Новички не селятся наугад на подходящих местах, а в пределах или рядом с существующей группой (Coulson, Coulson, 2009).
Состав гнездового материала у городских тихоокеанских чаек не отличался, как правило, от гнезд в природных колониях, где в состав выстилки наравне с природным материалом мог включаться и мусор, если он соседствовал с гнездом (Зеленская, 1992). У только начавшей гнездиться на крышах в г. Венеции средиземноморской чайки гнезда были построены только из естественных материалов (Soldatini et al., 2009). У серебристой чайки в Огайо (США) гнездовой материал был естественным только в 3% гнезд на крышах в городе и в 77% гнезд в природной колонии на острове (Belant, 1993). Таким образом, состав гнездового материала городских чаек отражает их огромную пластичность в способности “подстроиться” к возможностям среды обитания. Как крайний вариант подобного поведения можно рассматривать отношение к мусору серебристых чаек, гнездящиеся на засоренных берегах острова в Нью-Йоркской гавани. Чайки строили больше гнезд на участках, которые изобиловали обломками мусора и имели более крутой склон. Мусор служил чайкам не только гнездовым материалом, но и укрытием от ветров, убежищем для птенцов и, обладая плавучестью, спасал гнезда во время паводка (Maccarone et al., 1993).
Выбор здания для гнездования
Какому материалу крыши чайки отдают предпочтение – пока не ясно. В Магадане, как и в Англии (Rock, 2005), замена шифера на металл не отвратила чаек в Магадане от гнездования. По данным некоторых авторов определенные предпочтения имеются, например, если у клуш был выбор, то они предпочитали крыши, где был мох или трава (Sellers, Shackleton, 2011). У серебристых чаек расстояния между гнездами на засыпанных гравием крышах, были меньше таких расстояний на металлических или смоляных (последние не различались по плотности гнезд) (Belant, 1993).
Также нет единого мнения о том, предпочитают чайки скошенные или, наоборот, плоские крыши. Возможно, эти предпочтения видоспецифичны. Утверждается, что средиземноморские чайки предпочитают черепичные (скатные) крыши и поселяются на них, даже когда плоские крыши доступны (Soldatini et al., 2009). Серебристые чайки также предпочитают скошенные крыши (Monaghan, 1979; Raven, 1997). В Англии большинство клуш гнездилось на плоских или мягко скошенных крышах (Raven, 1997; Sellers, Shackleton, 2011). Но, возможно, предпочтение серебристыми чайками скатных крыш в Англии определено архитектурой застройки центров старых городов. В Германии сизые чайки гнездились на плоских крышах, и только единичные гнезда располагались на обычных скошенных крышах (Kubetzki, Garthe, 2007). Не найдено предпочтения определенного типа крыши для серокрылых чаек (Hooper, 1988; Vermeer et al., 1988).
Важнейшее значение, на наш взгляд, имеет предназначение зданий. Давно отмечено в Британской Колумбии для серокрылых чаек (Hooper, 1988), а также в Шотландии для клуш и серебристых чаек, что птицы с высокой плотностью гнездились на плоских крышах индустриальных и коммерческих зданий, а низкая плотность размещения пар была отмечена на жилых домах (Coulson, Coulson, 2009). При этом первоначально чайки в Англии гнездились на жилых и административных зданиях и очень редко – на индустриальных. Сейчас гнездовий на индустриальных объектах в Англии значительно больше (Raven, Coulson, 1997; Rock, 2005). Аналогичную картину мы наблюдали при заселении тихоокеанскими чайками г. Магадан. Переход к массовому гнездованию на промышленных объектах происходит не только из-за перераспределения колоний из центра города в промышленную зону, но из-за того, что крыши индустриальных зданий, имея большую площадь, позволяют существовать намного более плотным и многочисленным колониям (Vermeer et al., 1988; Raven, Coulson, 1997; Rock, 2005). Кроме того, подобные крыши гарантируют безопасность от хищников, отличный обзор и простор для взлета (Hooper, 1988).
Выбор гнездового участка и места для построения гнезда
Как отмечал Харитонов (1993), в понятие “гнездовой консерватизм” может вкладываться как расширенная трактовка – способность птиц возвращаться на прежнее место размножения, так и узкая – значительная для птиц, возвращающихся на прежнее место размножения. При этом “место гнездования” также может иметь различный смысл: от конкретного места прошлогоднего гнезда до того же самого географического района. В данной работе гнездовой консерватизм понимается в узком смысле и “прежнее место гнездования”, это как минимум – прошлогодний гнездовой участок.
Мы не маркировали взрослых тихоокеанских чаек в городе ни кольцами, ни краской, и не можем утверждать, что гнездо, возводимое ежегодно на одном и том же месте на крыше, принадлежит той же самой паре, что и в прошлые сезоны. Однако обзор литературы позволяет с большой вероятностью это предполагать. Согласно результатам наблюдений за индивидуально помеченными серебристыми чайками Тинбергеном (1974), в некоторых случаях гнездо сооружалось на одном месте два года подряд, а однажды – три года. Он подчеркивал, что после зимних ветров от прошлогоднего гнезда не оставалось даже следа. По данным кольцевания и индивидуального мечения чаек, гнездовой консерватизм – скорее правило для таких видов больших белоголовых чаек, как: серебристая чайка (Тинберген, 1974; Chabrzyk, Coulson, 1976; Пыжьянов, 1985), серокрылая чайка (Vermeer, 1963; Blokpoel, Courtney, 1980), западная чайка (L. occidentalis) (Annett, Pierotti, 1999; Pyle et al., 1991), сизая чайка (Онно, 1970), кольцеклювая чайка (L. delawarensis) (Southern, Southern, 1979, 1982), средиземноморская чайка (Soldatini et al., 2008, 2009), клуша (Rock, 2005), хохотунья (L. cachinnans) (Панов и др., 1990), большая морская чайка (L. marinus) (Краснов, 1998).
Как известно, плотность гнездования – индикатор размеров территории (Burger, 1984). Однако плотность может отражать микроландшафт (очень плотная растительность, невозможность взлететь с гнезда), а не фактический размер территории. Выбор гнездового участка отражает предпочтение той или иной микросреды обитания (Kharitonov, Siegel-Causey, 1988). Другая характеристика среды обитания – минимальное расстояние от гнезда или гнездовой территории, ближе которого не позволено находится незнакомцу (Харитонов, 1982). “Зрительная изоляция” способствует сокращению расстояния между гнездами и устраняет конфликты между близкими соседями (Burger, 1984). “Зрительную изоляцию” обеспечивают в естественной среде обитания растительный покров, камни и бревна, а в условиях города – любые преграды в виде труб, вентилей, стен, а также резкий уклон кровли, желоба, козырьки и прочие “изломы” крыш. Склонность чаек устраивать гнездо около преграды свойственна многим видам: серебристой чайке (Haycock, Threllfall, 1975; Monaghan, 1979; Belant, 1993; Raven, 1997; Sellers, Shackleton, 2011), клуше (Monaghan, 1979; Raven, 1997), серокрылой чайке (Hooper, 1988; Vermeer et al., 1988), чернохвостой чайке (Lee et al., 2006), западной чайке (Pierotti, Annett, 2001), сизой чайке (Kubetzki, Garthe, 2007). В полной мере эта особенность отмечена нами у тихоокеанской чайки как в природных популяциях (Зеленская, 1992), так и в урбанизированных гнездовьях. Кроме того, наличие преград может служить укрытием для птенцов от ветров и атак взрослых конспецифичных чаек.
“Сдвиги” места гнезда на гнездовом участке относительно их положения в прошлом сезоне, отмеченные нами в колониях тихоокеанских чаек на крышах г. Магадана, явление широко распространенное в природных колониях у озерной чайки (L. ridibundus) (Харитонов, 2011), у западной чайки (Pyle et al., 1991). У серебристой чайки в естественных колониях где их не беспокоили, 18% пар год от года уменьшали или увеличивали расстояния между гнездами больше чем на 5 м (Coulson, Coulson, 2009). Пары, которым не удалось успешно завершить инкубацию или которые потеряли птенцов в первые 5 дней после их вылупления, вероятнее всего изменят местоположение гнезда на своем участке в последующем сезоне (Graves et al., 1986). На крышах Шотландии межгодовые изменения расстояний между гнезд у чаек были связаны с беспокойством людьми, даже если конкретным парам не удаляли их гнезда (Coulson, Coulson, 2009).
Продуктивность
По нашим данным, в успешных природных колониях Тауйской губы продуктивность тихоокеанской чайки была следующей: о-в Шеликан – 0.60 ± 0.14 птенца на пару; о-в Умара – 0.35 ± 0.07; о-в Талан (неопубликованные данные Л.Ф. Кондратьевой) – 0.60±0.31, что значительно ниже, чем у городских чаек (рис. 6). В Англии именно на городских скошенных крышах у серебристых чаек отмечали наибольшую продуктивность (1.9–2.0) (Monaghan, 1979). В Бельгии отмечена более высокая продуктивность у городских серебристых чаек (1–1.5 птенцов/на пару), по сравнению с естественными колониями (0.5–1) (Francois, 2002). В Мурманской области успешность размножения серебристых чаек в естественных колониях 1–1.5 птенцов/пару, в городе Мурманск: 2006 г. – 2.5, в 2008 г. – 2.4, в 2009 г. – 2.3 (Горяев и др., 2011).
Известно, что в колониях на участках с высокой плотностью гнезд оперялось меньше птенцов чаек, чем на участках с низкой плотностью (Hunt, Hunt 1976). Авторы предполагали, что при отсутствии межвидового хищничества, большие территории могут давать репродуктивное преимущество у чаек из-за уменьшения конспецифичного хищничества (Hunt, Hunt 1976). Позже это предположение было подтверждено (Butler, Trivelpiece, 1981).
Но не все однозначно. Успех размножения двух видов чаек (серебристая и кольцеклювая) на крыше промобъекта неподалеку от оз. Эри, Огайо, был ниже, чем для этих видов указывалось другими исследователями для естественных колоний на островах и волнорезах (Belant et al., 1998). Гнездящиеся на крыше чайки не были более молодыми и менее опытными, чем чайки на острове. По мнению исследователей, крыши – подходящий ресурс для обитания, который только начал использоваться (Belant, 1993). Вероятно, нужно некоторое время для приспособления к новой среде обитания и для выработки и закрепления определенных ответов на воздействие человека.
В настоящее время тихоокеанская чайка продолжает гнездиться в естественных колониях на морском побережье и островах в Тауйской губы, в том числе и рядом с городом. Численность птиц в природных колониях быстро растет независимо от городской популяции (Зеленская, Хорева, 2006; Зеленская, 2008, 2012, 2012а, 2013). В настоящий момент численность городской популяции составляет около 4% от численности тихоокеанской чайки, гнездящейся в природных колониях островов и побережья Тауйской губы. При этом, как показано выше, продуктивность в городе значительно выше, чем в природных колониях.
Эти два параметра – доля городских чаек от природных популяций и соотношение успеха размножения у “диких” и урбанизированных чаек – различаются при сравнении более старых и совсем недавно сформированных городских популяций у крупных чаек. История гнездования серебристых чаек на крышах в Англии насчитывает более 100 лет. За последние 45 лет численность их в природных колониях Великобритании уменьшилась, но в городах рост популяции продолжается. По крайней мере, 15% всех гнездящихся в Англии серебристых чаек в 2000 г. жило в городах (Coulson, 2015). При этом успех их размножения в городах в разные годы был выше, чем в природных колониях (Monaghan, 1979; Raven, Coulson, 1997; Rock, 2005).
Гнездование средиземноморских чаек на крышах г. Венеции продолжалось менее десятилетия в период изучения. Городская популяция составляла 1.03% от растущей популяции чаек, размножающихся в Венецианской лагуне (Soldatini et al., 2009). Успех размножения в г. Венеция (0.41) был лишь немного выше, чем в окружающих естественных колониях (0.35). И единственным преимуществом в городе, по мнению авторов, было отсутствие наводнения и хищников. Однако было отмечено отсутствие различий в успехе размножения в городе за 3 года, в то же время в естественных колониях доля гнезд-неудачников достоверно различалась (Soldatini et al., 2008). При сравнении успеха размножения в городах Венеция и Триест отмечено, что в Триесте он выше, вероятно, из-за более длинной истории колонизации, т.е. птицы имели больший опыт урбанизации (Soldatini, et al., 2008).
Возможно, в разных городах Великобритании успех размножения популяций определен именно опытом урбанизированности. Так, на крышах в северной Шотландии не было никаких существенных различий в продуктивности клуш и серебристых чаек в разных колониях (1.5–2.5 птенца/пару) (Sellers, Shackleton, 2011). На северо-востоке Англии в смешанной колонии успех размножения для серебристой чайки составил 1.45 птенцов/гнездо, а для клуши этот показатель был достоверно ниже – 0.62 птенца/гнездо (Raven, 1997).
Кроме опыта урбанизации популяций, для повышения успеха размножения, вероятно, важна степень склонности к хищничеству у конкретного вида чаек. Так, у серокрылой чайки успех размножения в колониях в городе либо не отличался от аналогичного показателя для природных колоний (Hooper, 1988), либо в городе был ниже (0.35 слетка/пару), чем в рассеянном и уединенном гнездовании (1.1 слетка/пару) (Vermeer et al., 1988). Причина – только внутривидовое хищничество. На естественных колониях высокая птенцовая смертность из-за конспецифичной агрессии была зарегистрирована у нескольких видов крупных чаек (Parsons, 1971; Hunt, Hunt, 1976). Как известно, даже временное отсутствие взрослых птиц на гнездовом участке в природных колониях, связанное с его защитой, может значительно уменьшить репродуктивный успех для большинства видов чаек (Hunt, 1972; Gillet et al., 1975; Robert, Ralph, 1975; Hand, 1980; Burger, 1981; Fetterolf, 1983; Brown, Morris, 1994; Carney, Sydeman, 1999).
У тихоокеанских чаек, по нашим наблюдениям на природных колониях, гибель яиц от внутривидового хищничества иногда превышала 50%, а социальные конфликты являются одной из самых распространенных и массовых причин травматизма и смертности птенцов. Особенно это актуально для колоний, расположенных на участках со слабо развитой растительностью или вообще лишенных растительного покрова – укрытия для спасающихся бегством птенцов (Зеленская, 2008). Потому особенно поразительно отсутствие внутривидового хищничества яиц и избиения птенцов чайками-соседями на крышах г. Магадана, где история урбанизации насчитывает около 20 лет.
Теория синурбизации (урбанизации) (Luniak, 2004) обосновывает несколько существенных экологических и поведенческих различий между популяциями одних и тех же видов, живущих в городах и в природе. Мы сопоставили полученные нами данные с данными о типичных чертах синурбических популяций, приведенные Луняком (2004) (табл. 1). Из 9 позиций положение одной не ясно, совпадают четыре и столько же – нет. Возможны два варианта интерпретации: либо магаданская городская популяция еще слишком “молода”, и ее формирование еще продолжается, либо теория синурбизации не совершенна и требует доработки.
Таблица 1.
Соответствие типичных черт популяции тихоокеанской чайки г. Магадана современной теории о синурбизации природных популяций
| Типичные черты синурбической популяции (по Luniak, 2004) | Совпаде-ние | Наши данные |
|---|---|---|
| Намного более высокая плотность популяции в связи с сокращением размеров территории пар | Нет | Плотность размещения гнезд чаек в городе ниже, чем в природных колониях |
| Сокращенное миграционное поведение в связи с лучшими условиями для зимовки в городе | Нет | Миграционное поведение городских и природных популяций полностью совпадает |
| Удлинение сезона размножения из-за благоприятных микроклиматических условий | Нет | Все фенологические даты совпадают |
| Увеличение продолжительности жизни за счет лучшего переживания зим и снижения пресса хищников, в сочетании со снижением индивидуального отбора (выживание инвалидов, птиц с высокой инвазией паразитов, альбинизмом и проч.) | ? | Наличие различий в физиологических показателях не проверялось, но ни случаев альбинизма (даже частичного), ни явных инвалидов у чаек на крышах мы не отмечали |
| Длительная циркадная деятельность | Да | Искусственное освещение явно усиливает ночную активность чаек в городе. |
| Изменения в гнездовых привычках включая использование антропогенных объектов в качестве убежищ, гнездовых мест, материала для гнезд | Да | Есть изменения в гнездовых привычках включая использование антропогенных объектов в качестве убежищ, гнездовых мест, материала для гнезд |
| Изменения в кормовом поведении | Да | Есть изменения в кормовом поведении |
| Терпимость к человеку | Да | Отмечена терпимость к человеку |
| Увеличение внутривидовой агрессии | Нет | Наблюдаются обратные тенденции |
ЗАКЛЮЧЕНИЕ
Первые единичные гнезда в г. Магадане отмечены в 1998 г. Численность урбанизированной популяции в 2017 г. составила более 1.4 тыс. гнезд. Безопасность крыш от хищников, вероятно, является решающим фактором, способствующим урбанизации чаек. В настоящее время центр города заселен чайками наиболее плотно, и численность их гнезд продолжает расти. В городе Магадан, в отличие от природных гнездовий, преобладают поселения одиночных пар и маленькие колонии (до 10 пар). Однако основная масса гнезд сосредоточена в относительно небольшом числе крупных колоний.
Расселение пар по городу, как правило, было в пределах видимости от старой колонии. В то же время, отдельные гнезда, не имеющие соседей в пределах видимости, успешно существуют много лет. Освоение новых районов города также начиналось с отдельных пар, не имевших соседей в зоне видимости. При успешном размножении вселившейся пары, новые пары-иммигранты могут начать гнездиться рядом. Если нет противодействия людей, то развитие новой колонии происходит стремительно. Темпы роста составляют в среднем 13% в год, аналогично городским поселениям чаек в Европе.
При выборе города как места для размножения, чайки уходят от морского побережья. С годами дистанция от новых гнезд до моря увеличивается. Отмечено избегание гнездования на крышах одноэтажных строений. Мы не нашли предпочтения определенного типа крыши и кровельного покрытия у тихоокеанских чаек при выборе здания для гнездования. Предназначение здания определяет степень посещаемости его кровли людьми. Это имеет решающее значение при выборе чайками крыши для строительства гнезда. Переход к массовому гнездованию на крышах промышленных объектов происходит не только из-за перераспределения колоний из центра города в промзону. Главное – недоступные для людей крыши индустриальных зданий позволяют существовать намного более плотным и многочисленным колониям. Та же картина наблюдается и во многих странах Европы и Америки. Тихоокеанские чайки строят гнездо в городе из сухой травы, собранной на газонах. Объем мусора в выстилке гнезд крайне индивидуален. Большинство гнезд его не содержит.
В поселениях урбанизированных чаек отмечен сильный гнездовой консерватизм. Большинство гнезд строится на одном и том же месте многие годы. Небольшая часть гнезд в следующем сезоне строится “со сдвигом” на расстояние примерно до 1 м относительно прошлого сезона. Обычно этот “сдвиг” вызывается появлением нового гнезда-соседа, из-за которого “приходится отодвигаться”. Либо в течение 2–3 сезонов гнездо немного “перемещается”, пока не займет “идеальное положение”, затем место гнезда становится постоянным. Как правило, гнезда строятся около любой преграды на крыше (стены, трубы, надстройки и проч.) либо на абсолютно открытом месте. Второе более характерно для одиночных пар, которые более осторожны и помимо защитной стены предпочитают места с хорошим обзором. Преграды способствуют “зрительной изоляции” от чаек-соседей и укрытию для птенцов от непогоды и атак взрослых конспецифичных чаек.
Продуктивность тихоокеанских чаек, гнездящихся на городских крышах, значительно выше, чем в природных колониях. Причина этого явления – отсутствие или значительное ослабление межвидового и внутривидового хищничества в городе по сравнению с хищничеством в природных колониях. Основная причина неудачи гнездования у городских чаек – прямое воздействие людей.
Вероятно, городской популяции нужно некоторое время для приспособления к новой среде обитания. В городских популяциях, существующих в течение длительного периода, прослеживается увеличение продуктивности колоний. Вероятно, этому способствуют два параллельно протекающих процесса. Первый – перераспределение гнездящихся чаек внутри города, т.е. переселение на крыши, где нет беспокойства. Второй – массовое поселение отдельными парами или маленькими колониями, что ведет к снижению/исчезновению внутривидового хищничества.
Список литературы
Артюхин Ю.Б., 2002. Необычное гнездование уссурийского баклана Phalacrocorax filamentosus и тихоокеанской чайки Larus schistisagus на юге Дальнего Востока // Биология и охрана птиц Камчатки. Вып. 5. М. 117.
Артюхин Ю.Б., Вяткин П.С., 2012. Гнездование тихоокеанских чаек Larus schistisagus на искусственных сооружениях в Камчатском Крае // Сохранение биоразнообразия Камчатки и прилегающих морей: материалы XIII международной научной конференции. Петропавловск-Камчатский. Камчатпресс. С. 60–62.
Виксне Я., 2006. Гнездование чайковых птиц и куликов на крышах в Латвии // Орнитологические исследования в Северной Евразии: Тезисы XII Международной орнитологической конференции Северной Евразии. Ставрополь: Изд-во СГУ. С. 112–113.
Голубова Е.Ю., Плещенко С.В., 1997. Колонии морских птиц северной части Охотского моря // Видовое разнообразие и состояние популяций околоводных птиц Северо-востока Азии. Магадан. СВНЦ ДВО РАН. С. 141–162.
Горяев Ю.И., Горяева А.А., Татаринкова И.П., 2011. Крупные чайки в антропогенных ландшафтах Западного Мурмана (Кольский полуостров) // Беркут. Т. 20 (1-2). С. 90–110.
Зеленская Л.А., 1992. Зависимость плотности гнездования, устройства гнезда и успеха инкубации от биотопического размещения гнезд у тихоокеанской чайки // Зоологический журнал. Т. 71. Вып. 7. С. 93–107.
Зеленская Л.А., 2004. Гнездование тихоокеанской чайки Larus schistisagus на крышах зданий Магадана // Биология и охрана птиц Камчатки. Вып. 6. М. С. 85–90.
Зеленская Л.А., 2008. Тихоокеанская чайка (Larus schistisagus Stejneger). Магадан: СВНЦ ДВО РАН. 156 с.
Зеленская Л.А., 2012. Состояние колоний морских птиц окрестностей г. Магадана // Вестник СВНЦ ДВО РАН. № 2. С. 45–49.
Зеленская Л.А., 2012а. Условия гнездования и численность тихоокеанской чайки Larus schistisagus (Laridae) в Ольской лагуне (Тауйская губа, Охотское море) // Вестник СВНЦ ДВО РАН. № 1. С. 37–42.
Зеленская Л.А., 2013. Колонии морских птиц Тауйской губы и полуострова Кони // Вестник СВНЦ ДВО РАН. № 3. 87–100.
Зеленская Л.А., 2014. Экология питания тихоокеанской чайки (Larus schistisagus) озера Курильского (южная Камчатка) // Зоологический журнал. Т. 93. № 4. С. 570–584.
Зеленская Л.А., 2016. Новые урбанизированные гнездовья тихоокеанской чайки Larus schistisagus // Русский орнитологический журнал. Т. 25. Экспресс-выпуск 1277. С. 1455–1465.
Зеленская Л.А., 2017. Экология тихоокеанской чайки (Larus schistisagus), гнездящейся на озере Кроноцком (центральная Камчатка) // Зоологический журнал. Т. 96. № 1. С. 67–82.
Зеленская Л.А., Хорева М.Г., 2006. Увеличение численности гнездовой колонии тихоокеанской чайки (Larus schistisagus) и деградация растительного покрова на о. Шеликан (Тауйская губа, Охотское море) // Экология. № 2. С. 140–148.
Краснов Ю.В., 1998. Экология и морфология морских и серебристых чаек Баренцева моря // Биология и океанография Карского и Баренцева морей (по трассе Севморпути). Апатиты. С. 260–325.
Литвиненко Н.М., 1980.Чернохвостая чайка. М.: Наука. 144 с.
Лобков Е.Г., 1981. О гнездовании тихоокеанской чайки на болотах на севере Камчатского Края // Размещение и состояние гнездовий околоводных птиц на территории СССР. М.: Наука. С. 41–42.
Лобков Е.Г., 1986. Гнездящиеся птицы Камчатки. Владивосток: ДВНЦ АН СССР. 304 с.
Лобков Е.Г., Карпухин Н.С., 2010. Орнитологический комплекс южной части Корякского нагорья и его трансформация в результате освоения Сейнав-Гальмоэнанского платинового узла // Биология и охрана птиц Камчатки. Вып. 9. М. С. 47–61.
Мельников Ю.И., 2014. Динамика пространственной структуры чайковых птиц в 11-летнем климатическом цикле (дельта р. Селенги, южный Байкал) // Экология. № 1. С. 53–61.
Назаров Ю.Н., 2004. Птицы города Владивостока и его окрестностей. Владивосток: Изд-во Дальневост. ун-та. 276 с.
Онно С., 1970. Об этологических механизмах, определяющих гнездовой консерватизм у сизой чайки // Материалы 7-й Прибалтийской орнитологической конференции. Рига. № 1. С. 65–68.
Панов Е.Н., Зыкова Л.Ю., Грабовский В.И., 1990. Биология гнездования, поведение и таксономия хохотуньи Larus cachinnans. 1. Пространственная структура и поведенческие аспекты гнездовой биологии // Зоологический журнал. Т. 69. Вып. 12. С. 92–105.
Пыжьянов С.В., 1985. Пространственная структура поселений колониальных птиц в зависимости от условий среды обитания (на примере серебристой чайки) // Теоретические аспекты колониальности у птиц. Материалы совещания по теоретическим аспектам колониальности у птиц. 16-18 окт., 1984. М. С. 121–128.
Тинберген Н., 1974. Мир серебристой чайки. М.: Мир. 272с.
Фирсова Л.В., Лобков Е.Г., Вяткин П.С., 1982. Тихоокеанская чайка (Larus schistisagus Stejneger) в Камчатской области // Бюллетень Московского общества испытателей природы. Отдел биол. Т. 87. № 1. С. 30–35.
Харитонов С.П., 1982. Регуляция плотности гнездования в колонии озерной чайки // Биологические науки. № 10. С. 28–34.
Харитонов С.П., 1993. Гнездовой консерватизм (гнездовая филопатрия) и переселения озерных чаек Larus ridibundus в пределах колонии // Русский орнитологический журнал. Вып. 2. № 3. С. 361–382.
Харитонов С.П., 2006. Пространственно-этологическая структура колоний околоводных птиц. Автореф. дис. … докт. биол. наук. М.
Харитонов С.П., 2011. Пространственно-этологическая структура колоний околоводных птиц // Зоологический журнал. Т. 90. № 7. С. 845–860.
Annett C.A., Pierotti R., 1999. Long-term reproductive output in Western Gulls: consequences of alternate tactics in diet choice // Ecology. V.80. № 1. P. 228–297.
Belant J.L., 1993. Nest-site selection and reproductive biology of roof- and island-nesting Herring Gulls // Trans. 58th N.A. Wilfl. & Natur. Resour. Conf. P. 78–86.
Belant J.L., Ickes S.K., Seamans T.W., 1998. Importance of landfills to urban-nesting herring and ring-billed gulls // Landscape and Urban Planning. V. 43. P. 11–19.
Blokpoel H., Courtney P., 1980. Site tenacity in a new Ring-billed Gull colony // Journal of Field Ornithology. V. 51. № 1. 1–5p.
Brown K.M., Morris R.D., 1994. The influence of investigator disturbance on the breeding success of Ring-billed Gulls (Larus delawarensis) // Colonial Waterbirds. V. 17. № 1. P. 7–17.
Burger J., 1974. Breeding biology and ecology of the Brown-hooded Gull in Argentina // The Auk. V. 91. P. 601–613.
Burger J., 1981. Effects of human disturbance on colonial species, particularly gulls // Colonial Waterbirds. V. 4. P. 28–36.
Burger J., 1984. Pattern, mechanism, and adaptive significance of territoriality in Herring gulls (Larus argentatus) // Ornithilogical Monographs. № 34. P. 1–91
Butler R.G., Trivelpiece W., 1981. Nest spacing, reproductive success, and behavior of the Great Black-backed Gull (Larus marinus) // The Auk. V. 98. № 1. P. 99–107.
Carney K.M., Sydeman W.J., 1999. A review of human disturbance effects on nesting colonial waterbirds // Waterbirds. V. 22. № 1. P. 68–79.
Chabrzuk G., Coulson J.C. 1976. Survival and recruitment in the Herring Gull Larus argentatus // J. Animal Ecology. V. 45. № 1. P. 187–203.
Coulson J.C., 2015. Re-evalution of the role landfills and culling in the historic changes in the Herring Gull (Larus argentatus) population in Great Britain // Waterbirds. V. 38. № 4. P. 339–354.
Coulson J.C., Coulson B.A., 2009. Ecology and colonial structure of large gulls in an urban colony: investigations and management at Damfries, SW Scotland // Waterbirds. V. 32. № 1. P. 1–15.
Cramp S., 1971. Gulls nesting on building in Britain and Ireland // Brithish Birds. V. 64. № 11. P. 476–484.
Fetterolf P.M., 1983. Effects of investigator activity on Ring-billed Gull behavior and reproductive performance // Wilson Bulletin. V. 95. № 1. P. 23–41.
Francois R., 2002. Numbers and behaviour of roof-nesting Herring Gulls Larus argentatus and Lesser Black-backed Gulls Larus fuscus in Belgium // Natuur oriolus. V. 68. № 3. P. 123–126.
Gillet W.H., Hayward J.L., Stout J.F., 1975. Effects of human activity on egg and chick mortality in a Glaucous-winged Gulls colony // Condor. V. 77. P. 492–495.
Graves J., Whiten A., Henzi P., 1986.Reproductive success and the location of the nest site in the territory of the Herring Gull (Larus argentatus) // Bird Study. V. 33. № 1. P. 46–48.
Hand J.L., 1980. Human disturbance in Western Gull Larus occidentalis livens colonies and possible amplification by intraspecific predation // Biol. Conserv. V. 18. № 1. P. 59–63.
Haycock K.A., Threalfall W., 1975. The breeding biology of the Herring Gull in Newfoundland // Auk. V. 92. P. 678–697.
Hooper T.D., 1988. Habitat, reproductive parameters, and nest-site tenacity of urban-nesting Glaucous-winged gulls at Vctoria, British Columbia // The murrelet. V. 69. № 1. P. 10–14.
Hunt G.L., 1972. Influence of food distribution and human disturbance on the reproductive success of Herring Gulls // Ecology. V. 53. № 6. P. 1051–1061.
Hunt G.L., Hunt M.W., 1976. Gull chick survival: the significance of growth rates, timing of breeding and territory size // Ecology. V. 57. P. 62–75.
Jokimaki J., Kaisanlahti- Jokimaki M.-L., Sorace A., Fernandez-Juricic E., Rodriguez-Prieto I., Jimenez M.D., 2005. Evalution of the “safe nesting zone” hypothesis across an urban gradient: multi-scale study // Ecography. V. 28. P. 59–70.
Kharitonov S.P., Siegel-Causey D., 1988. Colony formation in seabirds // Current Ornithology. V. 5. P. 223–272.
Kubetzki U., Garthe S., 2007. Nests with a view: Distribution, nest habitats and diets of roof-breeding common gulls (Larus canus) in Northern Germany // Waterbirds. V. 30. № 4. P. 602–608.
Lee W.-S., Kwon Y.-S., Yoo J.-C., Song M.-Y., Chon T.-S., 2006. Multivariate analysis and self-organizing mapping applied to analysis of nest-site selection in Black-tailed Gulls // Ecological Modelling. V. 193. P. 602–614.
Luniak M., 2004. Synurbization – adaptation of animal wildlife to urban development // Proceeding 4th International urban wildlife symposium. Swan et al., eds. P. 50–55.
Maccarone A.D., Brzorad J.H., Parsons K.C., 1993. Nest site selection by Herring Gulls in an urban Estuary // Colonial Waterbirds. V. 16. № 2. P. 216–220.
Mazda M., Higuchi Y., 1976. Редкий случай гнездования тихоокеанской чайки на Хоккайдо // Tori Ornithological Society of Japan V. 25. № 100. Р. 81–82.
Monaghan P., 1979. Aspects of the breeding biology of herring gulls Larus argentatus in urban colonies // Ibis. V. 121. P. 475–481.
Monaghan P., Coulson J.C., 1977. Status of large gulls, nesting on buildings // Bird Study. V. 24. P. 89–104.
Parsons J., 1971. Cannibalism in Herring Gull Larus argentatus // Ibis. V. 118. P. 537–546.
Pierotti R., Annett C.A., 2001. The ecology of Western Gulls in habitats varying in degree of urban influence. In: Marzluff J.M., Bowman R., Donelly R. (Eds), Avian Conservation in an Urbanizing World. Klumer Academic Publishers. Boston. P. 307-329.
Pyle P., Spear L.B., Sydeman W.J., Ainley D.G., 1991. The effects of experience and age on the breeding performance of Western Gulls // The Auk. V. 108. P. 25–31.
Raven S. J., 1997. Aspects of the ecology of gulls in the urban environment // Durham theses, Durham University. Available E-Tesis Online: http//etheses.dur.ac.uk/5069/
Raven S., Coulson J.C., 1997. The distribution and abundance of Larus gulls nesting on buildings in Britain and Ireland // Bird Study. V. 44. № 1. P. 13–34.
Robert H.C., Ralph C.J., 1975. Effects of human disturbance on the breeding success of gulls // Condor. V. 77. P. 495–499.
Rock P., 2005. Urban gulls: problems and solutions // British Birds. V. 98. P. 338–355.
Sellers R.M., Shackleton D., 2011. Numbers, distribution and population trends of large gulls breeding in Cumbria, northwest England // Seabirds. V. 24. P. 90–102.
Soldatini C., Albores-Barajas Y.V., Mainardi D., Monaghan P., 2008. Roof nesting by gulls for better or worse? // Italian Journal of Zoology. V. 75. № 3. P. 295–303.
Soldatini C., Albores-Barajas Y.V., Mainardi D., Tgorricelli P., 2009. A widespread gull population in a complex wetland: habitat specific methods to census breeding pairs // Avocetta. V. 33. P. 205–210.
Southern L.K., Southern W.E., 1979. Philopatry in Ring-billed Gulls // Proc. 1979 Conf. Colonial Waterbird Group. V. 3. P. 27–32.
Southern L.K., Southern W.E., 1982. Effect of habitat decimation on Ring-billed Gull colony- and nest-site tenacity // The Auk. V. 99. P. 328–331.
Spear L.B., Anderson D.W., 1989. Nest-site selection by Yellow-footed Gulls // The Condor. V. 91. P. 91–99.
Vermeer K., 1963. The breeding ecology of the Glaucous-winged Gull (Larus glaucescens) on Mandarte Island B.C. Occ. Pap. B.C. Prov.Mus. V. 13. P. 1–101.
Vermeer K., Power D., Smith G.E.J., 1988. Habitat selection and nesting biology of roof-nesting Glaucous-winged Gulls // Colonial Waterbirds. V. 11. № 2. P. 189–201.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал