

Зоологический журнал, 2019, T. 98, № 5, стр. 513-524
Замечания по внутривидовой структуре шашечницы Melitaea tangigharuensis de Freina 1980 (Lepidoptera, Nymphalidae) с описанием нового подвида из Восточного Ирана
К. А. Колесниченко *
Биологический факультет Московского государственного университета
им. М.В. Ломоносова
119991 Москва, Россия
* E-mail: kkolesnichenko@gmail.com
Поступила в редакцию 09.04.2018
После доработки 08.06.2018
Принята к публикации 24.06.2018
Аннотация
Приведено описание нового подвида Melitaea tangigharuensis nambi ssp. n. из Восточного Ирана, дана сравнительно-морфологическая характеристика внешних признаков и генитальных структур самцов и самок нового и номинативного подвидов. Описание внешних признаков и гениталий самки номинативного подвида приведено впервые. Обсуждается таксономическое положение вида в составе рода Melitaea.
Очень редкий и локальный вид Melitaea tangigharuensis de Freina 1980 распространен в восточной части Иранского нагорья. На основании морфологических признаков он совместно с M. sarvistana Wiltshire 1941 формирует изолированную группу в составе рода (Колесниченко, 2011; van Oorschot, Coutsis, 2014). Генитальный аппарат самцов характеризуется седловидно-вогнутым дорсальным краем вальвы, раздвоенным на конце каудальным отростком и серповидной гарпой.
M. tangigharuensis описан из окрестностей г. Кабул (Афганистан) по трем самцам и до недавнего времени был известен только по типовой серии и единичным находкам из той же местности (Sakai, 1981). Для Ирана вид приведен Эквайлером (Eckweiler, 2008) и Колесниченко (2011). Сравнение экземпляров M. tangigharuensis из Ирана с фотографиями голотипа и паратипа выявило своеобразие иранских бабочек (Колесниченко, 2011), однако, отсутствие в нашем распоряжении материала из Афганистана не позволило произвести полноценное сравнение обеих популяций. В обзорной работе по роду Melitaea (van Oorschot, Coutsis, 2014) авторы рассматривали иранские популяции M. tangigharuensis в качестве номинативного подвида. Необходимо отметить, что ими была изучена морфология гениталий только одного самца (паратипа) M. tangigharuensis из Афганистана и трех самцов этого вида из Ирана.
С 2012 г. украинскими энтомологами, посетившими Афганистан, собрана и передана нам серия экземпляров этого редкого вида из окрестностей города Пагман, расположенного в непосредственной близости от типового местообитания. Изучение афганских экземпляров и сравнение их внешней морфологии и строения гениталий самцов и самок с экземплярами этого вида из Ирана подтвердило своеобразие иранских популяций. Ниже приводится переописание самцов и описание самки номинативного подвида и описание нового подвида из Восточного Ирана.
Нами изучен материал, хранящийся в коллекции кафедры энтомологии Московского государственного университета им. М.В. Ломоносова и включающий собственные сборы с территории Ирана и сборы других коллекционеров с территории Афганистана. Всего изучено 13♂♂ и 1♀ номинативного подвида M. tangigharuensis из Афганистана, включая фотографии типовых экземпляров из коллекции Дж. Де Фрейны, и 19 самцов и 3 самки M. tangigharuensis с территории Восточного Ирана.
Для большинства экземпляров самцов и самок были изготовлены препараты гениталий. Копулятивные органы препарировали по стандартной методике: у каждого исследуемого экземпляра отделяли часть брюшка, несущую генитальный аппарат, и помещали для мацерирования на трое суток в 10% раствор КОН. Затем препарат извлекали из щелочи, очищали от мышечных и жировых тканей, промывали последовательно в воде и в 75%-ном этиловом спирте, помещали в блистер, наполненный глицерином, и подкалывали под соответствующий коллекционный экземпляр. Препараты гениталий фотографировали при помощи камеры Canon EOS 6D, оснащенной объективом для макросъемки Canon MP-E 65. Съемку каждого препарата проводили в виде серии последовательных кадров с фокусировкой на разные слои объекта с дальнейшим объединением слоев в одном изображении на персональном компьютере при помощи программы-стейкера Combine ZM. Бабочек фотографировали при помощи фотокамеры Canon 5D. Редактирование рисунков и фотографий проводили в программе Adobe Photoshop. Терминология генитальных структур дана по Хиггинсу (Higgins, 1941), терминология крылового рисунка – по: Некрутенко (1985) и Колесниченко (Kolesnichenko, 1999).
В тексте приняты следующие сокращения: ЗММУ – Зоологический музей Московского государственного университета им. М.В. Ломоносова, КЭМГУ – кафедра энтомологии Московского государственного университета им. М.В. Ломоносова.
Melitaea tangigharuensis tangigharuensis de Freina 1980
Melitaea tangigharuensis de Freina, 1980: 17.
Типовая местность: “Afghanistan, Kabul-Fluẞ, Tang-i-Gharu-Schlucht, 1600 m, 22–23.V. 1977”.
Материал. Голотип ♂ (рис. 1, 6; рис. 2, 6), Afghanistan, Kabul-Fluẞ, Tang-i-Gharu-Schlucht, 1600 m, 22–23.V. 1977 (коллекция Дж. Де Фрейны); паратип ♂ (рис. 1, 7; 2, 7), те же данные (коллекция Дж. Де Фрейны); 4 ♂♂, Afghanistan, Kabul Prov., 5 km NW Kabul t., Bandi-Karga gorge, 2000 m., 6.06.2012, O. Pak; 2 ♂♂, Afghanistan, Kabul Prov., Pagman Distr., Qargha vill., 2000 m., 1.06.2010, I. Pljustsch Leg.; 2 ♂♂, 1 ♀, те же данные и сборщик, 7.06.2012; 3 ♂♂, C Afghanistan, Kabul Prov., Paghman Distr., near Qargha vill., 2000 m., 8.06.2016, N 34°32′, E 69°02′, Yu. Skrylnik leg. (все – КЭМГУ).
Рис. 1.
Подвиды Melitaea tangigharuensis de Freina 1980, общий вид бабочек сверху. 1 – M. tangigharuensis tangigharuensis самец: Afghanistan, Kabul Prov., 5 km NW Kabul, Bandi-Karga gorge; 2 – то же, самец: Afghanistan, Kabul Prov., 5 km NW Kabul, Bandi-Karga gorge; 3 – то же, самец: Afghanistan, Kabul Prov., Paghman Distr., near Qargha vill.; 4 – то же, самец: Afghanistan, Kabul Prov., Paghman Distr., near Qargha vill.; 5 – то же, самец: Afghanistan, Kabul Prov., Pagman Distr., Qargha vill.; 6 – то же, самец (голотип): Afghanistan, Kabul river, Tang-i-Gharu gorge, 1600 m, 22–23.05.77 (photo by J.J. de Freina); 7 – то же, самец (паратип), те же данные, что и у голотипа (рhoto by J.J. de Freina); 8 – то же, самка: Afghanistan, Kabul Prov., Pagman Distr., Qargha vill.; 9 – M. tangigharuensis nambi ssp. n., самец (голотип): Iran, Sistan and Baluchestan province, 35 km NE Zahedan town; 10 – то же, самец (паратип): те же данные, что и у голотипа; 11 – то же, самец (паратип): Iran, Khorasan province, Qaen t. area, 15 km SW Khezri Dasht Beyaz t.; 12 – то же, самец (паратип): Iran, Khorasan province, Qaen t. area, 15 km SW Khezri Dasht Beyaz t.; 13 – то же, самец (паратип): Iran, Khorasan province, Qaen t. area, 15 km SW Khezri Dasht Beyaz t.; 14 – то же, самец (паратип): Iran, Sistan and Baluchestan province, 35 km NE Zahedan town; 15 – то же, самец (паратип): Iran, Khorasan province, 65 km SW Khusf town, Dasht-e Lut desert, Ab-e Garm locality; 16 – то же, самка (паратип): Iran, Khorasan province, Qaen t. area, 15 km SW Khezri Dasht Beyaz t.

Рис. 2.
Подвиды Melitaea tangigharuensis de Freina 1980, общий вид бабочек снизу. 1 – M. tangigharuensis tangigharuensis, самец: Afghanistan, Kabul Prov., 5 km NW Kabul, Bandi-Karga gorge; 2 – то же, самец: Afghanistan, Kabul Prov., 5 km NW Kabul, Bandi-Karga gorge; 3 – то же, самец: Afghanistan, Kabul Prov., Paghman Distr., near Qargha vill.; 4 – то же, самец: Afghanistan, Kabul Prov., Paghman Distr., near Qargha vill.; 5 – то же, самец: Afghanistan, Kabul Prov., Pagman Distr., Qargha vill.; 6 – то же, самец (голотип): Afghanistan, Kabul river, Tang-i-Gharu gorge, 1600 m, 22-23.05.77 (рhoto by J.J. de Freina); 7 – то же, самец (паратип), те же данные, что и у голотипа (рhoto by J.J. de Freina); 8 – то же, самка: Afghanistan, Kabul Prov., Pagman Distr., Qargha vill.; 9 – M. tangigharuensis nambi ssp. n., самец (голотип): Iran, Sistan and Baluchestan province, 35 km NE Zahedan town; 10 – то же, самец (паратип), те же данные, что и у голотипа; 11 – то же, самец (паратип): Iran, Khorasan province, Qaen t. area, 15 km SW Khezri Dasht Beyaz t.; 12 – то же, самец (паратип): Iran, Khorasan province, Qaen t. area, 15 km SW Khezri Dasht Beyaz t.; 13 – то же, самец (паратип): Iran, Khorasan province, Qaen t. area, 15 km SW Khezri Dasht Beyaz t.; 14 – то же, самец (паратип): Iran, Sistan and Baluchestan province, 35 km NE Zahedan town; 15 – то же, самец (паратип): Iran, Khorasan province, 65 km SW Khusf town, Dasht-e Lut desert, Ab-e Garm locality; 16 – то же, самка (паратип): Iran, Khorasan province, Qaen t. area, 15 km SW Khezri Dasht Beyaz t.

Описание. Самец (рис. 1, 1–7; 2, 1–7). Размах крыльев 37–42 мм. Окраска верха крыльев красно-оранжевая. Постдискальное просветление верхней стороны передних крыльев, расположенное кнаружи от дискального ряда, хорошо выражено по всей длине, светлое пятно имеется в дистальной части дискоидальной ячейки. Черный субмаргинальный ряд верха крыльев всегда хорошо выражен и состоит из заостренных черных лунок, черный маргинальный край тонкий с хорошо выраженными маргинальными лунками или треугольными пятнами вдоль внешнего края крыльев. Дискальный ряд верха передних крыльев представлен небольшими черными пятнами, часто отсутствующими между жилками Cu2 и M3 и слитыми друг с другом ближе к костальному краю. На задних крыльях дискальный ряд и базальное затемнение, как правило, отсутствуют. Нижняя сторона передних крыльев с выраженным просветлением в постдискальной и в апикальной областях, а также в дистальной части дискоидальной ячейки. Основной цвет испода задних крыльев белый, маргинальный край – светло-оранжевый, субмаргинальная перевязь желтоватого цвета и состоит из пятен, центрированных ярко-оранжевыми точками и окаймленных черными чешуйками. Ближе к костальному краю ярко-оранжевые точки отсутствуют. Оранжевая дискальная перевязь на исподе задних крыльев состоит из отдельных пятен, не соединенных друг с другом. Ширина булавы усиков не менее чем в 5 раз короче длины (рис. 3., 1–3; 7–9). Основной цвет булавы – светло-коричневый. На верхней стороне в центральной части булавы имеется слабо выраженное темно-коричневое затемнение, вдоль края иногда расположены белые чешуйки (рис. 3, 2). Апикальная часть испода булавы усиков окрашена в светло-коричневые цвета, базальная часть покрыта напылением чешуек серо-белого цвета.
Рис. 3.
Форма и окраска булавы усиков подвидов M. tangigharuensis de Freina 1980. 1 – M. tangigharuensis tangigharuensis, верхняя сторона усиков: Afghanistan, Kabul Prov., 5 km NW Kabul, Bandi-Karga gorge. 2 и 3 – то же, те же данные; 4 –M. tangigharuensis nambi ssp. n., верхняя сторона усиков: Iran, Sistan and Baluchestan Province, 35 km NE Zahedan; 5 – то же: Iran, Khorasan province, Qaen t. area, 15 km SW Khezri Dasht Beyaz t.; 6 – то же: Iran, Khorasan province, 75 km N Birjand town, Sedeh village vicinity; 7 –M. tangigharuensis tangigharuensis, нижняя сторона усиков: Afghanistan, Kabul Prov., 5 km NW Kabul, Bandi-Karga gorge; 8 и 9 – то же, те же данные; 10 –M. tangigharuensis nambi ssp. n., нижняя сторона усиков: Iran, Sistan and Baluchestan Province, 35 km NE Zahedan; 11 – то же: Iran, Khorasan province, Qaen t. area, 15 km SW Khezri Dasht Beyaz t.; 12 – то же: Iran, Khorasan province, 75 km N Birjand town, Sedeh village vicinity.

Гениталии (рис. 4, 1–3; 5, 1–6; 6, 1–4). Ветви саккуса относительно тонкие. Овальной формы вальва резко сужена в дистальной части. Каудальный отросток тонкий, S-образно изогнут и дистально раздвоен в виде вилки. Длина лопастей вилки примерно одинакова. Данный признак совпадает с рисунком вальвы паратипа M. tangigharuensis, приведенным в обзорной работе по роду Melitaea (van Oorschot, Coutsis, 2014): на рисунке довольно хорошо видно, что у самца из Афганистана вентральный шип приблизительно такого же размера, что и дорсальный. Тонкая гарпа имеет форму косы, без дополнительных зубцов на внутренней стороне. Эдеагус прямой, превышает длину вальвы примерно в 1.5 раза, расширен в центральной части и резко сужен дистально.
Рис. 4.
Гениталии самцов и самок M. tangigharuensis de Freina 1980, общий вид. 1 – M. tangigharuensis tangigharuensis, самец: Afghanistan, Kabul Prov., 5 km NW Kabul, Bandi-Karga gorge; 2 – то же, самец: Afghanistan, Kabul Prov., Pagman Distr., Qargha vill.; 3 – то же, самец: Afghanistan, Kabul Prov., Pagman Distr., Qargha vill.; 4 – M. tangigharuensis nambi ssp. n., самец (паратип): Iran, Khorasan province, 75 km N Birjand town, Sedeh village vicinity; 5 – то же, самец (паратип): Iran, Sistan and Baluchestan province, 35 km NE Zahedan town; 6 – то же, самец (паратип): Iran, Khorasan province, 65 km SW Khusf town, Dasht-e Lut desert, Ab-e Garm locality; 7 – M. tangigharuensis tangigharuensis, самка: Afghanistan, Kabul Prov., Pagman Distr., Qargha vill.; 8 – M. tangigharuensis nambi ssp. n., самка (паратип): Iran, Khorasan province, Qaen t. area, 15 km SW Khezri Dasht Beyaz t.; 9 – то же, самка (паратип): Iran, South Khorasan Province, 65 km SW Husf t., Dasht-e Lut, Obe-Garme locality.
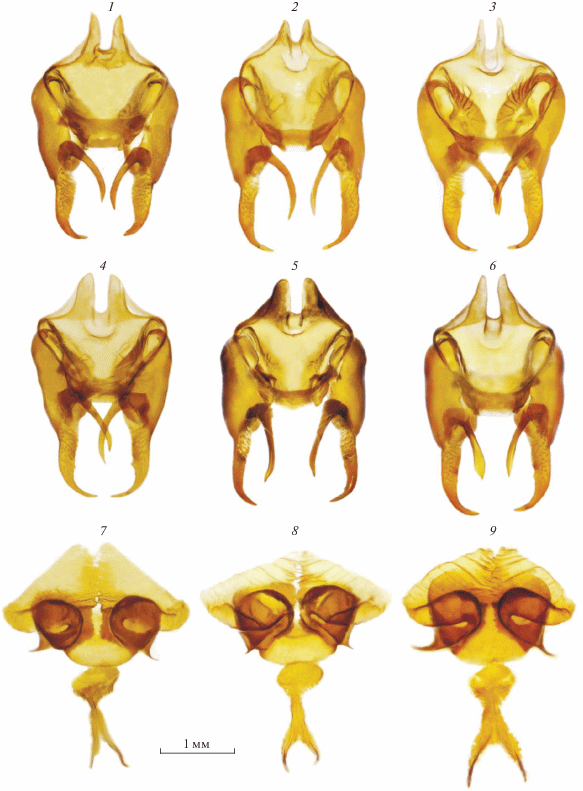
Самка (рис. 1, 8; 2, 8). У единственной самки, находящейся в нашем распоряжении, размах крыльев составляет 40 мм. Окраска верхней стороны крыльев красно-оранжевая. Постдискальное просветление верхней стороны передних крыльев, расположенное кнаружи от дискального ряда, а также светлое пятно в дистальной части дискоидальной ячейки хорошо выражены. Черный рисунок верха крыльев развит: дискальный ряд передних крыльев представлен черными точками, слитыми друг с другом, постдискальный ряд представлен относительно крупными слитыми друг с другом точками у костального края, крупные субмаргинальные лунки оттенены черными чешуйками. На верхней стороне задних крыльев имеются черные пятна и штрихи в дискальной области, а базальное затемнение практически отсутствует. Рисунок испода крыльев сходен с таковым самцов. Две самки номинативного подвида, изображенные книге “Butterflies of Afghanistan” (Tshikolovets et al., 2018) на таблице LIII (fig. 11, 12), по внешнему облику еще более контрастные, чем изученная нами самка: точки дискального и постдискального рядов оттенены черным и сильно контрастируют с постдискальным просветлением.
Гениталии (рис. 4, 7). Антевагинальная пластинка с широко расставленными лопастями и заметно сужена к основанию. Поствагинальная пластинка треугольная (ее основание в 3 раза превышает длину), а вершинная часть значительно выступает за пределы антевагинальной пластинки. Округлой формы аурикулы широко расставлены: расстояние между ними составляет приблизительно 1.5 длины базальной части антевагинальной пластинки.
Рис. 5.
Гениталии самцов подвидов M. tangigharuensis de Freina 1980. A – вальва, B – эдеагус. 1 – M. tangigharuensis tangigharuensis: Afghanistan, Kabul Prov., 5 km NW Kabul, Bandi-Karga gorge; 2 – то же: Afghanistan, Kabul Prov., 5 km NW Kabul, Bandi-Karga gorge; 3 – то же: Afghanistan, Kabul Prov., Pagman Distr., Qargha vill.; 4 – то же: Afghanistan, Kabul Prov., Pagman Distr., Qargha vill.; 5 – то же: C Afghanistan, Kabul Prov., Paghman Distr., near Qargha vill.; 6 – то же: Afghanistan, Kabul Prov., 5 km NW Kabul, Bandi-Karga gorge; 7 – M. tangigharuensis nambi ssp. n., паратип: Iran, Sistan and Baluchestan Province, 35 km NE Zahedan; 8 – то же, паратип: Iran, Sistan and Baluchestan Province, 35 km NE Zahedan; 9 – то же, паратип: Iran, Khorasan province, 75 km N Birjand town, Sedeh village vicinity; 10 – то же, паратип, те же данные; 11 – то же: Iran, Khorasan province, Qaen t. area, 15 km SW Khezri Dasht Beyaz t.; 12 – то же, паратип: Iran, South Khorasan Province, 65 km SW Husf t., Dasht-e Lut, Obe-Garme locality.
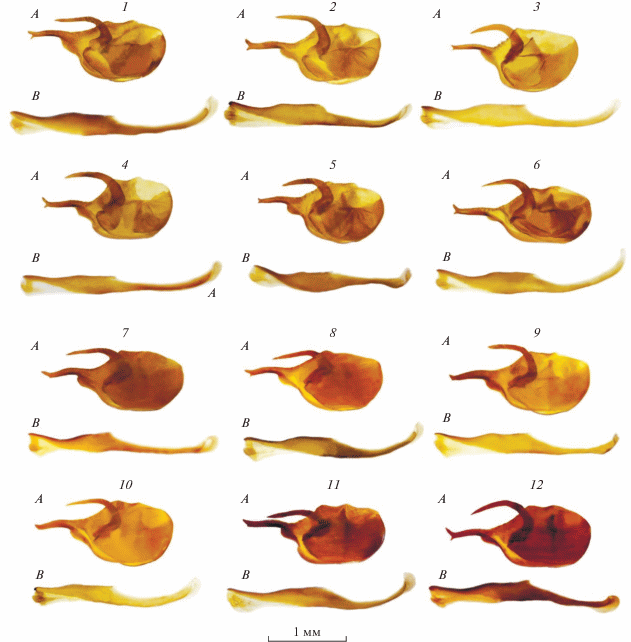
Рис. 6.
Гениталии самцов подвидов M. tangigharuensis de Freina 1980. Форма дистальной части каудального отростка вальвы. 1 – M. tangigharuensis tangigharuensis: Afghanistan, Kabul Prov., 5 km NW Kabul, Bandi-Karga gorge; 2 – то же: Afghanistan, Kabul Prov., 5 km NW Kabul, Bandi-Karga gorge; 3 – то же: Afghanistan, Kabul Prov., Pagman Distr., Qargha vill.; 4 – то же: Afghanistan, Kabul Prov., 5 km NW Kabul, Bandi-Karga gorge; 5 – M. tangigharuensis nambi ssp. n.: Iran, Sistan and Baluchestan Province, 35 km NE Zahedan; 6 – то же: Iran, South Khorasan Province, Qaen t. area, 15 km SW Khezri Dasht Beyaz t.; 7 – то же: Iran, South Khorasan Province, 75 km N Birjand, Sedeh v. vicinity; 8 – то же: Iran, South Khorasan Province, 30 km S Qaen t., Hunik pass.

Биология. Лёт имаго приходится на конец мая–начало июня в зависимости от высоты. Бабочки обитают по аридным склонам с вегетирующей эфемерной растительностью на высоте от 1500 до 2000 м (рис. 7, 1).
Рис. 7.
Биотопы подвидов M. tangigharuensis de Freina 1980. 1 – Место обитания M. tangigharuensis tangigharuensis de Freina 1980 (Afghanistan, Kabul Prov., 5 km NW Kabul, Bandi-Karga gorge): щебнистый склон с вегетирующей эфемерной растительностью (фото О.В. Пака); 2 – место обитания M. tangigharuensis nambi ssp. n. (Iran, Khorasan province, Qaen t. area, 15 km SW Khezri Dasht Beyaz t.): щебнистый склон с деревьями безлистного миндаля Amygdalus scoparia Spach; 3 – место обитания M. tangigharuensis nambi ssp. n. (Iran, South Khorasan Province, 65 km SW Husf t., Dasht-e Lut, Obe-Garme locality): сухие щебнистые склоны обращенные в сторону пустыни Деште Лут.

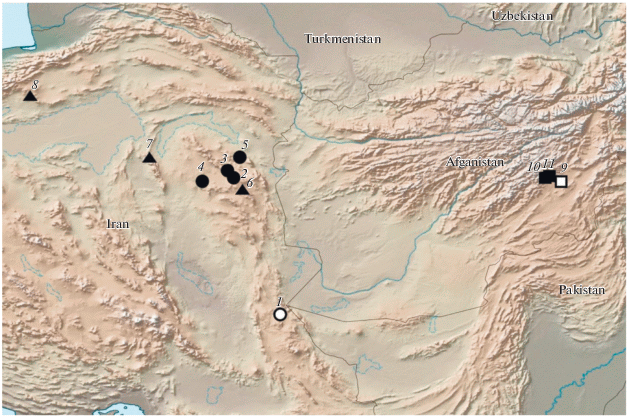
Распространение (рис. 8). До сих пор номинативный подвид известен только из окрестностей г. Кабул (de Freina, 1980; Sakai, 1981; Tschikolovets et al., 2018)
Рис. 8.
Распространение M. tangigharuensis по: де Фрейна (de Freina, 1980), Сакаи (Sakai, 1981), Эквайлеру (Eckweiler, 2008) и нашим данным. ⚪ – Голотип M. tangigharuensis nambi ssp. n.; ⚫ – паратипы M. tangigharuensis nambi ssp. n.; ▲ – литературные данные о распространении M. tangigharuensis в Иране. ◻ – голотип M. tangigharuensis tangigharuensis; ◼ – распространении M. tangigharuensis tangigharuensis. 1 – Sistan and Baluchestan Province, 35 km NE Zahedan; 2 – South Khorasan Province, 75 km N Birjand, Sedeh v. vicinity; 3 – South Khorasan Province, 30 km S Qaen t., Hunik pass.; 4 – Khorasan province, 65 km SW Khusf town, Dasht-e Lut desert, Ab-e Garm locality; 5 – Khorasan province, Qaen t. area, 15 km SW Khezri Dasht Beyaz t.; 6 – Iran, Khorasan Province, Kahi; 7 – Iran, 25 km E Tabaz t., Kharveola [Kharv-e-Bola]; 8 – Semnan Province, 40 km E Semnan; 9 – Kabul, Tang-i-Gharu valley; 10 – Aliabad, Kabul; 11 – Kabul Province, Paghman district, Qargha L.
Melitaea tangigharuensis nambi Kolesnichenko ssp. n.
Материал. Голотип ♂: Iran, Sistan and Baluchestan Province, 35 km NE Zahedan, 10.04.2004, K. Kolesnichenko leg. Паратипы: 4 ♂♂, те же данные, что и голотип; 3 ♂♂, Iran, South Khorasan Province, Qaen t. area, Khezri Dasht Beyaz t. vicinity, 1800–2300 m., 25.05.2008, K. Kolesnichenko leg.; 1 ♂, 1 ♀, те же данные, A.Klimenko leg.; 1 ♂, те же данные, A. Devyatkin leg.; 3 ♂♂, Iran, South Khorasan Province, Qaen t. area, 15 km SW Khezri Dasht Beyaz t., 1800–2000 m., N 33°56′, E 58°43′, 30.05.2009, K. Kolesnichenko leg.; 1 ♀, Iran, South Khorasan Province, 30 km S Qaen t., Hunik pass, 2000 m., 25.05.2009, A. Devyatkin leg.; 2 ♂♂, Iran, South Khorasan Province, 75 km N Birjand, Sedeh v. vicinity, 1500 m., 27–28.04.2004, K. Kolesnichenko leg.; 4 ♂♂, 1 ♀, Iran, South Khorasan Province, 65 km SW Husf t., Dasht-e Lut, Obe-Garme, 29.04.2006, K. Kolesnichenko leg. Голотип будет передан в ЗММУ. Паратипы хранятся в коллекции КЭМГУ.
Описание. Самец (рис. 1, 9–15; 2, 9–15). Размах крыльев 37–40 мм, у голотипа – 39 мм. Основной цвет верха крыльев ярко-красный или красный. На верхней стороне передних крыльев субмаргинальный ряд представлен черными лунками, редуцированными между жилками Cu1 и M3, желтое просветление дискальной области отсутствует либо выражено незначительно (рис. 1, 11–13), черные точки дискального ряда редуцированы между жилками Cu2 и M3. Область центральной ячейки однотонная, окрашена в цвета верха крыльев. Дискальный ряд верхней стороны задних крыльев отсутствует. На нижней стороне передних крыльев белое просветление выражено только ближе к апикальному краю между маргинальным краем и субмаргинальным рядом и между субмаргинальным и постдискльным рядами. Рисунок испода задних крыльев сходен с таковым номинативного подвида. Ширина булавы усиков не менее чем в 3 раза короче длины (рис 3, 4–6; 10–12). Основной цвет булавы – светло-коричневый. На верхней стороне в центральной части булавы имеется хорошо выраженное темно-коричневое затемнение. Вдоль края булавы, начиная с базальной части и до половины, расположена заметная полоса из белых чешуек. Рисунок испода булавы усиков состоит из светло-коричневой вершинной части и серого напыления чешуек в центральной и базальной частях.
Гениталии (рис. 4, 4–6; 5, 7–12; 6, 5–8). Ветви саккуса утолщенные, их ширина в основании в 2–3 раза превышает ширину основания саккуса номинативного подвида. Дистальная часть каудального отростка вилкообразно раздвоена, причем вентральная лопасть заметно короче дорсальной. Исключение составляет единственный экземпляр самца из восточной окраины пустыни Деште Лут (Obe-Garme), у которого лопасти дистальной части каудального отростка приблизительно одинаковой длины (рис. 4, 12). У экземпляров с северной части провинции Систан и Белуджистан каудальный отросток (при просмотре вальвы сбоку) повернут таким образом, что вентральная лопасть вилки расположена на внутренней поверхности каудального отростка. Эдеагус нового подвида по форме и размерам как у номинативного.
Самка (рис. 1, 16; 2, 16). Размах крыльев 42–44 мм. Окраска верхней стороны крыльев оранжевая или желто-оранжевая, с относительно выраженным просветлением в дискальной области, в постдискальной области у костального края и в центральной ячейке. Черный рисунок верха передних крыльев развит: дискальный ряд хорошо выражен и представлен черными точками, слитыми друг с другом у костального края; постдискальный ряд хорошо выражен также у костального края и представлен слившимися точками, расположенными между костальным краем и жилкой M2, и штрихами, расположенными уже ближе к анальному краю; постдискальный ряд выражен вдоль переднего и на заднем крыле у костального края, крупные субмаргинальные лунки оттенены черными чешуйками. На верхней стороне задних крыльев имеются черные пятна в дискальной области, а базальное затемнение практически отсутствует. Рисунок испода крыльев сходен с таковым самцов.
Гениталии (рис. 4, 8, 9). У трех исследованных нами самок массивная антевагинальная пластинка разделена на лопасти и имеет форму квадрата за счет отсутствия заметного сужения к основанию. Поствагинальная пластинка округло-треугольной формы, ее основание почти в 2.5–3 раза превышает длину, а вершинная часть незначительно выступает за пределы антевагинальной пластинки. Крупные аурикулы хорошо выражены, а их внутренние края расположены в непосредственной близости друг к другу: расстояние между ними в 5–6 раз меньше длины основания гениталий (базальной части антевагинальной пластинки).
Диагноз. Самцы M. tangigharuensis nambi ssp. n. хорошо отличаются от самцов номинативного подвида редукцией постдискального просветления и отсутствием светлого пятна в дистальной части дискоидальной ячейки, а также ярко-красным или красным основным цветом верхней стороны передних крыльев. Вальвы самцов нового подвида имеют характерные и существенные отличия: вилка дистальной части каудального отростка имеет разную длину лопастей, причем вентральная лопасть заметно короче дорсальной (рис. 6, 5–8). У номинативного подвида лопасти вилки примерно одинаковы по длине и размеру (рис. 6, 1–4). Различия в размере и форме данной структуры довольно существенны, но часто при первом взгляде не заметны из-за сложной трехмерной структуры вальвы: каудальный отросток иногда повернут таким образом, что вентральная лопасть вилки направлена вовнутрь (перпендикулярно плоскости вальвы) и просто не видна при рассматривании вальвы сбоку. Булава усиков у самцов нового подвида широкая: ее длина не более чем в три раза превышает ширину, тогда как у номинативного подвида булава усиков заметно вытянута, и ее длина не менее чем в 5 раз превышает ширину. Кроме того, у нового подвида на верхней стороне булавы по краю выражена полоса из белых чешуек.
Самки нового подвида характеризуются менее контрастным рисунком, в первую очередь, за счет более блеклого оранжево-желтого основного цвета верха крыльев. Сходная по внешним признакам самка из провинции Семнан (40 km E Semnan) изображена в книге “The butterflies of Iran and Iraq” (Tshikolovets et al., 2014) на таблице LX (fig. 18). Особенностью строения генитального аппарата самок нового подвида являются форма и пропорция антевагинальной и поствагинальной пластинок. Антевагинальная пластинка массивная, но более компактная, чем у номинативного подвида. Для нее характерна квадратная форма за счет широкого основания, которое по длине почти равно расстоянию между крайними внешними точками лопастей (рис. 4, 8, 9). У номинативного подвида форма данной пластинки обратно-треугольная за счет ее сужения к основанию (рис. 4, 7). Поствагинальная пластинка у нового подвида округло-треугольной формы, лишь слегка выступает за край антевагинальной пластинки своей вершинной частью, тогда как у номинативного подвида она значительно выступает за края антевагинальной пластинки. Кроме того, особенностью строения гениталий самок нового подвида следует считать небольшое расстояние между внутренними сторонами аурикул: у номинативного подвида это расстояние в 3–4 раза больше.
Биология. Лёт имаго M. tangigharuensis nambi ssp. n. наблюдается в крайне аридных горных биотопах и приурочен, как и у номинативного подвида, к вегетации эфемерной растительности. Время появления бабочек зависит от высоты и широты места распространения. На южных границах ареала (провинция Систан и Белуджистан) очень свежие самцы нового подвида пойманы 10 апреля в низкогорной (не выше 1200 м) части отрогов хребта Пеленган. Бабочки летали совместно с Hypermnestra helios (Nickerl 1846) и Pontia glauconome (Klug 1829). В центральной части Восточного Ирана в низкогорном массиве (около 1500 м) в районе г. Бирджент (п. Седех) лёт имаго приходится на конец апреля. В низкогорных массивах, обращенных в сторону пустыни Деште-Лут (рис. 6, 3), довольно облётанные имаго также пойманы в невысоких горах (1500 м) в конце апреля. Здесь бабочки летали совместно с Pontia glauconome (Klug 1829) и M. robertsi Butler 1880. Сдвиг фенологической фазы имаго M. tangigharuensis nambi ssp. n. с ранней весны практически на начало лета происходит при проникновении его высоко в горы: в высокой части горной системы Кайен (рис. 7, 2) на высотах около 2000 м свежие самцы и самки летали в конце мая. Самцы активно патрулируют территорию на горных склонах вдоль сухих русел временных водотоков (рис. 7, 2, 3), а самки обычно летают у входа в ущелья. Бабочки в окрестностях г. Захедан (провинция Систан и Белуджистан) летали по дну сухих “вади” и кормились на одиночных цветущих растениях.
Распространение (рис. 8). Основная масса находок M. tangigharuensis nambi ssp. n. сосредоточена в Восточно-Иранской горной области, от высокой части горной системы Кайен на севере до южных отрогов хребта Пеленган в северной части Иранского Белуджистана (Eckweiler, 2008; Колесниченко, 2011). Бабочки, сходные по внешней морфологии с новым подвидом, указывались для провинции Семнан (40 км к востоку от г. Семнан) (Eckweiler, 2008; Tshikolovets et al., 2014).
Этимология. “Nambi” – название сезонного южного ветра, которому подвержена юго-восточная территория Ирана, включая провинцию Систан и Белуджистан.
Ареал M. tangigharuensis охватывает несколько крупных и крайне аридных областей Иранского нагорья – от поднятий Восточно-Иранской горной области до провинции Кабул в Афганистане. Самое западное достоверно известное местообитание M. tangigharuensis расположено в невысоких горах провинции Семнан (Eckweiler, 2008), которые входят уже в горную систему Эльбурса (Петров, 1955). Таксономический статус этих популяций пока не ясен, но по внешней морфологии имаго их можно отнести к M. tangigharuensis nambi ssp. n.
Наличие M. tangigharuensis в восточной части Ирана в относительном удалении от известных мест распространения номинативного подвида может вызывать удивление: между обеими популяциями расположены высокие горы и довольно безжизненные окраинные пустыни, например, Систанская котловина. Однако примеров видов с подобным дизъюнктивным ареалом довольно много. Локальные и редкие виды, распространенные в горных областях Афганистана, часто имеют продолжение своего ареала в Иране. Примером тому может служить вид M. timandra Coutsis et Oorschot 2014, ареал которого сосредоточен в западных пределах хребта Гиндукуш в области озер Банди-Амир и северных склонов хребта Кох-и-Баба (провинция Бамиан, Афганистан), а также в юго-восточных Каракумах. В Иране этот вид известен с высокой части изолированного хребта Кухе-Беналуд, входящего в состав Туркмено-Хорасанских гор, а западным достоверно известным пределом распространения этого вида являются горы Шах-Кух в провинции Семнан (van Oorshot, Coutsis, 2014; Tschikolovets et al., 2014, наши данные). Другим примером подобного дизъюнктивного ареала может служить Colias wiskotti Staudinger 1882. Его крайний западный подвид hofmannorum Eckweiler 2000 обитает в высокой части Восточно-Иранской горной области, а ближайший подвид sweadneri Clench et Shoumatoff 1956 известен уже для высокогорий хребта Кох-и-Баба (Grieshuber, 2014) в Афганистане. Сходные дизъюнктивные ареалы имеют Hyponephele capella (Christoph 1877), H. davendra (Moore 1865) (Eckweiler, Bozano, 2011) и другие бабочки. Разрыв в распространении можно объяснить приуроченностью видов к определенным ландшафтам и локалитетам, в которых произрастают их кормовые растения. С подобной географической изоляцией связаны различия не только в характере рисунка крыльев, но и в деталях морфологии генитального аппарата самцов и самок. В нашем случае для каждого из подвидов характерна своя форма вилки дистальной части каудального отростка вальвы самцов, а для самок – форма и пропорции поствагинальной пластинки и различия в расстоянии между внутренними сторонами аурикул.
Систематическое положение M. tangigharuensis в составе рода до сих пор не определено. Считается, что родственным видом является M. sarvistana (Eckweiler, 2008; Колесниченко, 2007, 2011). Однако необходимо отметить, что, несмотря на внешнее сходство бабочек, разница в морфологических признаках генитальных структур самцов обоих видов весьма существенна. Данный факт дал основание Хиггинсу (Higgins, 1981) произвести, казалось бы, немотивированную перегруппировку в его системе подсемейства Melitaeinae: оба вида были отнесены к разным родам (или видовым группам в нашем понимании). При этом M. tangigharuensis рассматривался этим автором в составе рода Melitaea совместно с представителями группы cinxia, а M. sarvistana отнесен к роду Cinclidia совместно с представителями группы phoebe. Наличие тонкого изогнутого каудального отростка вальвы, действительно сближает M. tangigharuensis с представителями группы cinxia, а точнее – с M. arduinna (Esper [1783]). Позже было предложение рассматривать M. tangigharuensis, но уже совместно с M. sarvistana, в качестве изолированного комплекса в составе группы cinxia (Колесниченко, 2011). С другой стороны, Эквайлер (Eckweiler, 2008), основываясь на формальных признаках, рассматривал M. tangigharuensis в качестве представителя группы phoebe. Наконец, в обзорной работе по роду Melitaea (van Oorschot, Coutsis, 2014) M. tangigharuensis вместе с M. sarvistana был выделен в отдельную группу sarvistana. С последним заключением можно согласиться, учитывая крайнюю специфику морфологических признаков этих видов. Своеобразие M. tangigharuensis (как и M. sarvistana), на наш взгляд, во многом обусловлено адаптацией к ксерофитным условиям. Необходимо отметить, что южный предел распространения M. tangigharuensis совпадает с южным пределом распространения рода Melitaea и приурочен к аридным горным ландшафтам, окруженным областью равнин с крайне жарким климатом, часто пустынно-тропического типа.
БЛАГОДАРНОСТИ
За предоставленную возможность работы с материалом при подготовке данной статьи автор выражают глубокую и искреннюю признательность И.Г. Плющу (г. Киев, Украина), О.В. Паку (г. Донецк, Украина). Особую благодарность автор выражает г-ну Дж. де Фрейне (Германия) за предоставление фотографий типа и паратипа номинативного подвида M. tangigharuensis.
Работа выполнена при поддержке РНФ (14-50-00029: сбор и обработка материала, изготовление иллюстраций) и государственной темы № AAAA-A16-116021660095-7 (подготовка рукописи).
Список литературы
Колесниченко К.А., 2007. Эколого-таксономический обзор шашечниц рода Melitaea (Lepidoptera, Nymphalidae) // Бюллетень Московского общества испытателей природы. Отдел биологический. Т. 112. Вып. 3. С. 27–37.
Колесниченко К.А., 2011. Шашечница Melitaea tangigharuensis (Lepidoptera, Nymphalidae) в Иране // Бюллетень Московского общества испытателей природы. Отдел биологический. Т. 116. Вып. 2. С. 65–68.
Некрутенко Ю.П., 1985. Булавоусые чешуекрылые Крыма: Определитель. Киев: Наукова думка. 152 с.
Петров М.П., 1955. Иран (Физико-географический очерк). М.: Государственное издательство географической литературы. 184 с.
de Freina J.J., 1980. Melitaea tangigharuensis n. sp., eine neue Nymphalidenart für Afghanistan nebst vergleichender Betrachtung zur M. sarvistana Wiltshire, 1941 (Lepidoptera, Nymphalidae) // Atalanta. Zeitschrift der Deutschen Forschungszentrale für Schmetterlingswanderungen. Bd. XI. Heft 1. P. 11–18.
Eckweiler W., 2008. Melitaea tangigharuensis de Freina, 1980 in Iran (Lepidoptera, Nymphalidae) // Nachrichten Entomologischen Vereins Apollo. Bd. 29. Heft 1/2. P. 93–96.
Eckweiler W., Bozano G.C., 2011. Tribe Satyrini. Subtribe Maniolinae. Maniola, Pyronia, Aphantopus, Hyponephele. Bozano G.C. (ed.). Guide to the butterflies of the Palearctic region. Satyrinae part IV. Milano: Omnes Artes. 102 p.
Grieshuber J., 2014. Subfamily Coliadinae. Tribe Coliadini. Bozano G.C. (ed.). Guide to the butterflies of the Palearctic region. Pieridae part II. Milano: Omnes Artes. 86 p.
Higgins L.G., 1941. An illustrated catalogue of the Palearctic Melitaea (Lep. Rhopalocera) // Transactions of the Royal Entomological Society of London. V. 91. № 7. 86 p.
Higgins L.G., 1981. A revision of Phyciodes Hübner and related genera, with a review of the classification of the Melitaeinae (Lepidoptera: Nymphalidae) // Bulletin of the British Museum (Natural History). Entomology. Vol. 43. № 3. P. 77–243.
Kolesnichenko K.A., 1999. A review of the East-Palaearctic taxa of the Melitaea didyma (Esper, 1779)-group. I. The M. ala Staudinger, 1895 – M. chitralensis Moore, 1901 complex (Lepidoptera, Nymphalidae) // Atalanta. Zeitschrift der Deutschen Forschungszentrale für Schmetterlingswanderungen. Bd. 30. Heft 1/4. P. 87–117. Col. Pls. VI, VII.
Sakai S., 1981. Butterflies of Afghanistan. Tokyo: Koudansha. 272 p. [in Japanese].
Tshikolovets V., Naderi A., Eckweiler W., 2014. The butterflies of Iran and Iraq. Pardubice: Tshikolovets Publications. 366 p. Col. Pls. I–LXV.
Tshikolovets V., Pliushch I., Pak O., Skrylnik Y., 2018. The butterflies of Afghanistan. Kiev: Tshikolovets Publications. 263 p. Col. Pls. I–LXIV.
van Oorschot H., Coutsis J.G., 2014. The genus Melitaea Fabricius, 1807 (Lepidoptera, Nymphalidae, Nymphalinae). Taxonomy and systematics with special reference to the male genitalia. Pardubice: Tshikolovets publications. 360 p.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал