

Зоологический журнал, 2019, T. 98, № 5, стр. 535-542
Мониторинговые учеты белобрюшки (Aethia psittacula, Charadriiformes, Alcidae) – суточная и сезонная динамика численности в колониях Северной Пацифики
Н. Б. Конюхов *
Институт проблем экологии и эволюции РАН
119071 Москва, Россия
* E-mail: konyukhov@gmail.com
Поступила в редакцию 23.04.2018
После доработки 23.06.2018
Принята к публикации 12.07.2018
Аннотация
Мониторинг морских птиц, гнездящихся в задернованных осыпях, где птиц невозможно посчитать на поверхности колонии, задача непростая. Исследования, проведенные в колониях белобрюшек на островах Булдырь и Святого Павла, позволяют проанализировать характер изменения их численности в колониях и прилегающей к ним акватории, как в течение суток, так и в течение сезона размножения. Для учетов белобрюшек на обоих островах были заложены учетные площадки. Птиц учитывали визуально с помощью бинокля: на о-ве Булдырь в прибрежной акватории под колонией, на о-ве Святого Павла, как на поверхности колонии, так и в прибрежной акватории. Помимо серий учетов, охватывающих весь световой день, проводились и учеты только во время пиков активности птиц в колонии и прилегающей к ней акватории. Учеты проводились каждые 15 мин. На о-ве Булдырь в прибрежной акватории у белобрюшек выражена двухпиковая активность с большим утренним пиком: во время выкармливания птенцов он начинается позже, чем во время насиживания. На о-ве Святого Павла пик один: в начале сезона размножения он приходится на утро, а во время выкармливания птенцов – на вечер. Сильный ветер подавляет активность птиц и на поверхности колонии и в прилегающей акватории. На основании результатов наших исследований даны рекомендации по проведению мониторинговых учетов.
Морские птицы – хороший объект в мониторинговых исследованиях морских экосистем, поскольку они, являясь конечными звеньями пищевых цепей, могут быть индикаторами их состояния.
Белобрюшка – типичная морская птица весом около 300 г, связанная с сушей только во время сезона размножения. Остальное время она проводит в открытом море. Белобрюшка – один из пяти видов конюг, гнездящихся на побережье и островах северной части Тихого океана. В отличие от других видов, как правило, гнездится разреженными колониями численностью от нескольких до сотен пар. Колонии в тысячи особей крайне редки. Гнезда устраивает в разного рода укрытиях: щелях и пустотах скальной стенки, в пустотах голых и задерненых осыпей и крайне редко в норах, вырытых другими видами. Питается планктоном, который добывает в зонах стратификации водных масс, ныряя за ним на глубину на десятки метров (Jones et al., 2001).
В то время как численность популяций большинства морских птиц, гнездящихся на скалах открыто, задокументирована, об абсолютной численности и трендах численности популяций закрыто гнездящихся белобрюшек известно немного. Считается, что в дальневосточных водах России обитает примерно 40% мировой популяции этого вида (Sowls et al., 1978; Артюхин, 1999), однако мониторинг данного вида в этом регионе, исключая о-в Талан (Андреев и др., 2010; Голубова, 2015) не ведется. Без мониторинга начальное сокращение численности скрытно гнездящихся видов морских птиц может произойти незаметно и будет обнаружено только тогда, когда произойдут катастрофические изменения. Мониторинг состояния популяций, который основан на учетах гнездовых камер и который проводится в популяциях, например, тупиков Fratercula arctica и топорков Lunda cirrhata (Harris, Murray, 1981), не может быть применен в случае белобрюшек, поскольку птицы не гнездятся в норах, предпочитая трещины в скальных обрывах или пустоты между камнями как на берегу, так и в осыпях. Кроме того, фактором, затрудняющим мониторинг популяций белобрюшек, является то, что большинство мест, где возможно их гнездование, имеет несколько выходов или существуют общие входные отверстия для нескольких гнездовых камер. Зачастую гнездовые камеры находятся глубоко в нагромождениях обломочного материала или в узких извилистых каналах застывшей лавовой массы, что почти никогда не позволяет увидеть само гнездо (неопубликованные данные). Учет птиц на поверхности осыпи порой невозможен по той причине, что эти осыпи покрыты трасянистой растительностью, высота которой может достигать полутора метров.
Для видов, у которых прямой учет гнезд невозможен, единственной возможностью его провести является учет птиц, находящихся на поверхности колонии или на поверхности моря вблизи колонии. Для получения достоверных сведений первоначально необходимо изучить изменения числа птиц в колонии в течение суток и на протяжении всего сезона размножения. Это позволит выбрать оптимальный промежуток времени учетов для корректного определения численности вида в колонии (Byrd et al., 1983).
Данные настоящего исследования представляют собой методическую разработку исключительно мониторинговых учетов, которая поможет избежать ошибок при учетах как белобрюшек, так и подобных им видов птиц, гнездящихся в биотопах, где прямые учеты невозможны.
МАТЕРИАЛ И МЕТОДИКА
Суточные и сезонные изменения числа белобрюшек в колонии и ее прибрежной акватории изучались в 2001 и 2002 гг. на о-ве Булдыре, а также в 2004 г. на о-ве Святого Павла, где их численность соответственно составляет 10–12 и 34 тыс. особей (Byrd, Day, 1986; Sowls et al., 1978). Надо отметить, что на о-ве Булдыре нет и никогда не было наземных хищников, тогда как на о-ве Святого Павла обитает эндемичный прибыловский голубой песец (Vulpes lagopus pribilofensis).
На о-ве Булдыре белобрюшки гнездятся, главным образом, в осыпях, заросших травянистой растительностью. Поскольку буйная растительность на склоне Северо-западной гряды (52°22′24″ с.ш., 175°52′44″ в.д.), где находится большая колония белобрюшек, не позволяла учитывать птиц на ее поверхности, то для сбора данных по динамике численности птиц в 2001 г. на воде была заложена учетная площадка. На этой площадке отмечались социальные взаимоотношения птиц, гнездящихся на склоне. Ее границами являлись выступающие из воды огромные камни, расстояние между которыми составляло 380 м. Птиц учитывали в полосе около 150 м от уреза воды. Точка, с которой проводился учет, находилась на крутом склоне на высоте около 30 м над ур. м.
В 2001 г. в течение сезона размножения учеты проводились тремя пятидневными сериями: 28 мая– 1 июня (период насиживания), 21–25 июня (конец насиживания – самое начало вылупления птенцов) и 14–18 июля (период выкармливания птенцов). Все птицы, присутствовавшие на площадке, учитывались в течение всего периода их активности: утром с предрассветных сумерек до тех пор, пока все птицы не покидали площадку, т.е. до 13–15 часов, и вечером с 20:00 и до наступления темноты, когда силуэты птиц было сложно различать на фоне поверхности воды. Дополнительно к этим трем сериям учетов белобрюшки учитывались еще в течение 18 дней, из которых 7 учетов пришлись на инкубационный период, а 11 – на период выкармливания птенцов. Эти учеты проводились только во время утреннего (7:00–15:00) и вечернего (22:00–24:00) пиков активности птиц в колонии.
В 2002 г. площадка осталась в тех же границах, что и в 2001 г. В этом году в течение всей утренней активности (6:00–15:00) учеты белобрюшек проводились 14 и 24 июня (инкубационный период) и 14 и 27 июля (период выкармливания птенцов). Кроме того, только во время утреннего пика активности птиц (8:00–12:00) проведено еще 9 учетов, 5 из которых пришлись на период насиживания, а 4 – на период выкармливания птенцов.
На о-ве Св. Павла полевые исследования проводились 19 мая–13 августа 2004 г. Белобрюшек учитывали с мыса Турист Пойнт (57°09′28″ с.ш., 170°23′21″ з.д.). Здесь с обеих сторон от мыса была заложена площадка общей протяженностью 450 м. Птиц учитывали как на обрывах колонии, так и в прибрежной акватории на расстоянии 150 м от уреза воды. Точка, с которой проводились учеты птиц, находилась на высоте 7–8 м над ур. м. Утренний учет начинался в 7:00 и продолжался до 15:00 ч; вечерний начинался в 14:30 и продолжался до 22:00 ч. В прединкубационный период было проведено три утренних и два вечерних учета, в период насиживания – шесть утренних и шесть вечерних учетов, а в период выкармливания птенцов – шесть утренних и девять вечерних учетов. Утренний и вечерний учетные дни чередовались.
Прогноз погоды для судов, работающих в данной акватории, мы получали по радиосвязи или смотрели в интернете. Для оценки силы ветра во время учетов использовали шкалу Бофорта. Наблюдения на островах Булдырь в 2002 г. и Святого Павла проводили в дни, когда сила ветра была меньше 12.5 м/с (25 узлов).
Белобрюшек на всех площадках учитывались визуально с помощью 8× бинокля каждые 15 мин.
Общая продолжительность учетов в 2001 г. составила 263 ч 45 мин, в 2002 г. – 68 ч 30 мин, а в 2004 г. – 240 ч 45 мин.
Хронологию размножения белобрюшек на о-ве Булдыре я определял, осматривая гнездовые камеры на репродуктивных площадках, заложенных для мониторинга успеха размножения этого вида. В течение всего сезона размножения гнездовые камеры осматривались 1 раз в пять дней. Зачастую была известна только дата вылупления птенцов. В этом случае я вычислял медиану дат вылупления и от нее рассчитывал дату откладки яиц, приняв продолжительность насиживания, равной 35 дней (Sealy, Bedard, 1973; Jones et al., 2001). На о-ве Святого Павла белобрюшки гнездятся по обрывам; потоки лавы, Формирующие эти обрыва, испещренны труднодоступными пустотами. Длина их “нор” была значительной, что не позволило проводить прямые наблюдения за хронологией периода размножения. Поэтому для определения этапов сезона размножения я использовал косвенные данные: птицы, кормящие птенцов, носят корм в горловом мешке, который хорошо заметен при учетах. Даты, когда такие птицы стали отмечаться во время учетов, я принимал за начало периода вылупления птенцов. Следовательно, раньше этого времени был инкубационный период, а после – период выкармливания птенцов. Как и на о-ве Булдыре рассчитывалось и время начала периода насиживания.
Цифровые данные в тексте представлены следующим образом: X ± SD; в скобках приведено количество данных. Для сравнения выборок использовался Z–тест Манна–Уитни.
В статье дано административное летнее время: для о-ва Булдырь – Гавайско-алеутское стандартное время (UTC –10), для о-ва Святого Павла – Аляскинское стандартное время (UTC –9). Следует отметить, что на о-ве Булдыре астрономическое (солнечное) время отличается от административного примерно на 3 ч, а на о-ве Святого Павла – на 2 ч, т.е. от административного времени, принятого в данном районе, надо отнять соответствующего число часов. Например, восход солнца по астрономическому времени на о-ве Булдыре 25 июня приходился на 3:55, что соответствует 6:55 утра административного времени.
РЕЗУЛЬТАТЫ
Согласно данным, полученным на продуктивных площадках, белобрюшки начали откладывать яйца в конце мая–начале июня, а вылупление происходило в конце июня–начале июля. В 2001 г. период насиживания продолжался с 25 мая (начала полевых работ) до 1 июля–медианы периода вылупления птенцов. Период выкармливания птенцов, соответственно, охватывал период со 2 июля до слета птенцов. В 2002 г. медиана периода откладки яйца пришлась на 2 июня, а вылупления на 4 июля.
На о-ве Святого Павла первые птенцы белобрюшек вылупились примерно 10 июля, следовательно, инкубационный период продолжался с 4 июня по 10 июля, а после этой даты был период выкармливания птенцов.
На о-ве Булдыре у белобрюшек в течение суток, как в колонии, так и в прибрежной акватории отмечены два пика активности (рис. 1). Один из родителей, насиживающий яйцо или обогревающий маленького птенца, проводит ночь в гнезде, а другой в море. В предрассветных сумерках птицы стайками по 2–10 особей начинают прилетать в акваторию колонии. Сначала они собираются далеко от берега, а затем вплавь приближаются к колонии. Во время насиживания число птиц в прибрежных водах увеличивается примерно до 7:00. Затем птицы с воды начинают перемещаться в колонию, чтобы сменить партнера в гнезде. Число птиц на воде перед колонией стабилизируется в 8:00–9:30. Позже, в начале второй половины дня, число птиц постепенно сокращается до полного их разлета из прибрежной акватории. Вечером первые птицы на воде у колонии появляются в 19:00–20:00 ч. В это время, в отличие от первой половины дня, белобрюшки никогда не подходят близко к берегу. Их число достигает пика незадолго до наступления темноты, что происходит примерно в 23:00, а затем число птиц незначительно уменьшается. Позже, когда птиц уже невозможно видеть, какое–то число птиц все еще присутствует на учетной площадке, о чем можно было судить только по их голосам, доносившимся оттуда.
Рис. 1.
Среднее число белобрюшек на учетной площадке на о-ве Булдыре в течение насиживыания и выкармливания птенцов в 2001 г. (На графике отражены данные всех учетов; период насиживания продолжался c 28 мая до 1 июля, период выкармливания – с 2 до 29 июля).
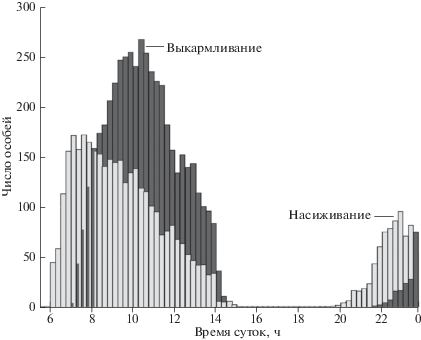
Во время выкармливания птенцов птицы прибывали на площадку позже, чем во время период насиживания (рис. 1), а пик численности приходится на 9:00–11:30, тогда как во время инкубации – на 7:00–10:00. Число птиц, посещавших учетную площадку во время вечернего пика активности, по мере развития сезона размножения постепенно сокращалось.
Неполовозрелые птицы, которых определяли по форме клюва и изношенности оперения, стали посещать колонию в самом конце инкубационного периода. В это время число птиц на учетной площадке увеличилось и достигло своего пика в первой половине периода выкармливания птенцов. Среднее число птиц, присутствовавших на площадке во время пика численности в 2001 г., в период выкармливания птенцов (271.3 ± 94.3 (SD)) было значительно больше, чем во время насиживания (169.5 ± 141.2 (SD); тест Манн–Уитни Z = 6.89 n = 373, P < 0.01). Во второй половине периода выкармливания птенцов число птиц, посещающих площадку, резко сократилось. В колонии остались только птицы, кормившие своих птенцов, а неразмножающиеся птицы и птицы, потерпевшие неудачу в размножении, перестали прилетать.
Как суточная, так и сезонная динамика численности белобрюшек в 2002 г. на о-ве Булдыре была сходна с таковой, что наблюдалась в 2001 г. Птицы в прибрежной акватории присутствовали как утром, так и вечером с той же тенденцией сдвига пика численности на более позднее время в период выкармливания птенцов.
На о-ве Святого Павла до откладки яиц число белобрюшек, посещающих колонию, достигало своего пика в первой половине дня. Пик численности птиц на воде близ колонии предшествовал их пику на скалах (рис. 2).
Рис. 2.
Среднее число белобрюшек на учетной площадке в колонии Турист Пойнт на о-ве Святого Павла в 2004 г. (Представлены данные: Прилет – 3 утренних и 2 вечерних учета, Насиживание – 6 утренних и 6 вечерних учетов, Выкармливание – 6 утренних и 9 вечерних учетов).
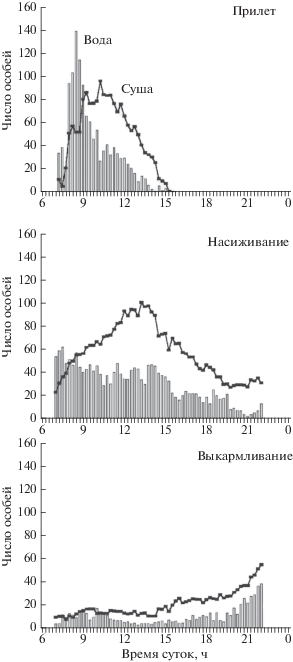
По мере развития сезона размножения, когда птицы стали насиживать яйца, и в колонии, и в прибрежной акватории белобрюшек можно было видеть в течение большего времени, чем в прединкубационный период, а пик их численности, который отмечался только на суше, сдвигался ближе ко второй половине дня. На воде число птиц изменялось в широких пределах с тенденцией уменьшаться в конце дня. В конце инкубационного периода был отмечен только небольшой вечерний пик.
Во время выкармливания птенцов, птицы присутствовали на колонии в течение всего дня с двумя пиками численности: небольшим утром и большим вечером.
Наблюдения показали, что погодные условия сильно влияют на число птиц, присутствующих на учетной площадке. Сильный ветер ведет к сокращению числа белобрюшек в прибрежной акватории. В ветреные дни птицы покидали площадку раньше обычного (рис. 3). В такие дни, как утром, так и вечером, птицы, избегая полосы прибоя, находились от берега дальше, чем обычно. При ветре в порывах 25–30 м/с и более птиц на площадке почти не было, исключая единичных особей, которые покидали прибрежную акваторию, порой даже не садясь на воду. Даже если день, следующий за штормовым, был тихим, птиц на площадке было меньше, чем обычно.
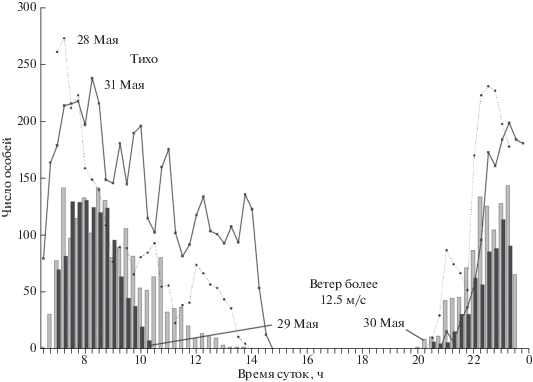
ОБСУЖДЕНИЕ
Среди конюг, исключая малую конюгу (Aethia pygmaea) (Byrd, Williams, 1993; Gaston, Jones, 1998; Зубакин, Конюхов, 1999; Pshenichnikova et al., 2017), у белобрюшки в разных частях ее гнездового ареала наблюдается наибольшее разнообразие суточной динамики численности в колониях. На юге ареала белобрюбшки, на Командорских и Алеутских островах, расположенных в южной части Берингова моря, выражены и утренний, и вечерний пики (Михтарьянц, 1978; Konyukhov, 2002; Konyukhov, Juk, 2001). На о-ве Булдыре двухпиковый паттерн активности птиц в колонии и прилегающей акватории сохраняется в течение всего периода размножения, от прилета птиц весной в колонию до слета птенцов осенью. В северной части Берингова моря (северный предел распространиния белобрюшки) и в заливе Аляска (восточная граница распространения вида), птицы собираются на поверхности колонии и в прилегающей акватории только утром, с рассвета до полудня (Sealy, Bedard, 1973; Конюхов, 1993; Hatch, 2002). На о-ве Талане, в Охотском море, у белобрюшек также выражен утренний пик, причем птицы проводят в колонии меньше времени, чем в прибрежной акватории (Harding et al., 2002). На островах Прибылова в центральной части Берингова моря ситуация с динамикой численности птиц в колонии наиболее интересная. В зависимости от стадии репродуктивного периода паттерн активности птиц в колонии занимает промежуточное положение между одно- и двухпиковой ежедневной активностью. Как уже отмечалось выше, в начале сезона размножения, перед откладкой яиц, птицы присутствовали в колонии и прилегающей акватории только в первой половине дня, т.е. так же, как и на севере и востоке ареала вида. В течение периода насиживания пик численности отмечен только на суше, но просматривается общая тенденция уменьшения числа птиц, как на суше, так и на воде, во второй половине дня. Отсутствие пика численности на воде вызвано, скорее всего, фактором беспокойства. Северные морские котики, которые группами и поодиночке перемещались между кормовыми районами и холостяцким лежбищем, расположенным неподалеку, периодически вспугивали белобрюшек и те улетали за пределы учетной площадки. Это лежбище заполняется котиками в середине июня, как раз во время высиживания и выкармливания белобрюшками птенцов. Формирование небольшого пика вечером в конце инкубационного периода, вероятно, обусловлено тем, что в некоторых гнездах птенцы уже вылупились и родители приносили им корм.
Во время периода выкармливания птенцов небольшой утренний пик формировали, в основном, птицы, приносившие птенцам корм. Эти птицы, как правило, садились прямо перед входом в гнездо и исчезали в нем, оставаясь на поверхности суши минимальное время. Покормив птенца, они сразу же слетали в море. Птицы не были помечены индивидуально, но поскольку родители остаются в гнезде с птенцом только первые 3–5 дней его жизни (неопубликованные данные), то, вероятно, в течение всего сезона выкармливания птенцов, исключая самое его начало, птица, прилетевшая с кормом и покинувшая гнездо несколько минут спустя, была, как я считаю, той же самой, а не ее партнером, который продолжал оставаться в гнезде. Вечерний пик в этот период, вероятно, формировали как птицы, кормящие птенцов, так и птицы, не участвующие в размножении: неполовозрелые и потерпевшие неудачу в размножении взрослые. Когда во второй половине периода выкармливания птенцов птицы, не кормящие птенцов, покидают колонию, то этот пик исчезает. Пока не найдено объяснения такому разнообразию циркадных ритмов у белобрюшки в разных частях ее ареала. Возможно, дальнейшие исследования прольют свет на его причины.
Как было отмечено выше, сильный ветер значительно уменьшал число птиц, посещавших учетную площадку. Сходным образом сильный ветер угнетал и активность ипаток в колонии (Конюхов, 2010). Влияние ветра могло выражаться двояко. Ветер сильнее 25 м/с просто сдувает птиц с поверхности колонии. Но главная причина, вероятно, заключается в том, что он, вызывая сильное волнение, перемешивает поверхностный слой моря и разрушает зоны стратификации водных масс, где кормятся белобрюшки. Птицам для добывания рассеянного корма требуется больше времени, чтобы насытиться, а возможно в это время они голодают. В пользу этого предположения говорит тот факт, что на следующий после окончания шторма день птиц на учетной площадке было учтено меньше, чем обычно. Подобное влияние ветреной погоды отмечено как для большой конюги, так и для тихоокеанского чистика на Чукотке, где рост птенцов из-за недостатка корма приостанавливался (Конюхов, 1997).
Все описанное выше особенности биологии вида нужно принимать во внимание при проведении мониторинговых учетов.
Если в данном районе учеты проводятся впервые, то сначала необходимо определить время суток, когда число птиц в колониях данного региона максимально, и именно в этот период проводить учет.
Учетная площадка должна иметь выраженные границы, что позволит из года в год получать сравнимые данные.
Поскольку ежедневные изменения числа белобрюшек на учтенных площадках были минимальны в период инкубации, то и учеты птиц желательно проводить именно в этот период.
Как было сказано выше, число белобрюшек в колониях уменьшается при увеличении скорости ветра; при скорости ветра в 12.5 м/с (25 узлов) и выше активность птиц на поверхности колонии подавляется почти полностью. Следовательно, учеты не следует проводить в дни с такой силой ветра, а также на следующий день после шторма, когда число птиц в колонии и прилегающей акватории меньше, чем в нормальные дни.
БЛАГОДАРНОСТИ
Я искренне благодарен сотрудникам Аляскинского морского национального природного заказника (Alaska Maritime National Wildlife Refuge) за предоставленную возможность работы на островах Булдырь и Святого Павла, за финансовую и техническую поддержку исследований, а Е.К. Литвину, С.П. Харитонову и В.А. Зубакину – за ценные замечания при подготовке статьи к публикации. Ценные замечания рецензента помогли улучшить качество данной публикации. Данные исследования проводились в рамках Российско-американского соглашения в области охраны окружающей среды.
Список литературы
Андреев А.В., Голубова Е.Ю., Зубакин В.А., Харитонов С.П., 2010. Численность морских птиц на колониях о. Талан: двадцатилетний тренд // Вестник Северо-Восточного научного центра ДВО РАН. № 2. С. 30–42.
Артюхин Ю.Б., 1999. Кадастр колоний морских птиц Командорских островов // Биология и охрана птиц Камчатки. Вып. 1. С. 25–35.
Голубова Е.Ю., 2015. Биология размножения белобрюшки (Cyclorrhynchus psittacula) на острове Талан (Северная часть Охотского моря) // Зоологический журнал. Т. 94. Вып. 7. С. 832–847.
Зубакин В.А., Конюхов Н.Б., 1999. Биология размножения малой конюги (Aethia pygmaea): характер гнездования, активность в колонии и социальное поведение // Известия РАН. Сер. биол. Вып. 5. С. 562–571.
Конюхов Н.Б., 1993. Сравнительная биология некоторых видов закрытогнездящихся видов чистиковых. Автореф. дис. … канд. биол. наук. М.: ИЭМЭЖ РАН СССР. 24 с.
Конюхов Н.Б., 1997. Влияние погодных факторов на биологию размножения чистиковых // Известия РАН. Сер. биол. Вып. 2. С. 228–234.
Конюхов Н.Б., 2010. Суточные и сезонные изменения количества ипаток (Fratercula corniculata) по учетам при помощи фотографирования // Зоологический журнал. Т. 89. Вып. 9. С. 1118–1124.
Михтарьянц Э.А., 1978. Ритмы активности белобрюшки (Cyclorrhynchus psittacula) и топорка (Lunda cirrhata) и связанные с ними регулярные флюктуации их численности на гнездовых колониях // Экология и зоогеография некоторых позвоночных суши Дальнего Востока. Владивосток: ДВНЦ АН СССР. С. 31–39.
Byrd G.V., Day R.H., 1986. The avifauna of Buldir Island, Aleutian Islands, Alaska // Arctic. V. 39. P. 109–118.
Byrd G.V., Day R.H., Knudtson E.P., 1983. Patterns of colony attendance and censusing of Auklets at Buldir island, Alaska // Condor. V. 85. № 3. P. 274–280.
Byrd G.V., Williams J.C., 1993. Whiskered Auklet (Aethia pygmaea) // The Birds of North America. Washington, D.C., Philadelphia: The academy of Natural Sciences. № 76. 12 p.
Gaston A.J., Jones I.L., 1998. The Auks (Bird Families of the World), Oxford: Oxford Univ. 349 p.
Harding A.M.A., Piatt J.F., Byrd G.V., Hatch S.A., Konyukhov N.B., Golubova E.U., 2002. Attendance patterns and population monitoring of crevice–nesting Horned Puffins and Parakeet Auklets in the North Pacific. Homer, Alaska: U.S. Fish and Wildlife Service. 50 p.
Harris M.P., Murray S., 1981. Monitoring of Puffin numbers at Scottish colonies // Bird Study. V. 28. № 1. P. 5–20.
Hatch S.A., 2002. Activity patterns and monitoring numbers of Horned Puffins and Parakeet Auklets // Waterbirds. V. 25. № 3. P. 348–357.
Jones I. L., Konyukhov N.B., Williams J.C., Byrd G.V., 2001. Parakeet Auklet (Aethia psittacula). The Birds of North America. № 594 (Poole A., Gill F., eds). Philadelphia, PA: The Birds of North America, Inc. 20 p.
Konyukhov N.B. 2002. Attendance patterns of parakeet auklets and horned puffins at Buldir Island, Alaska in 2002 // U.S. Fish and Wildl. Serv. Rep. AMNWR 02/05 Homer, Alaska. 45 p.
Konyukhov N.B., Juk K., 2001. Attendance patterns of parakeet auklets and horned puffins at Buldir Island, Alaska in 2001 // U.S. Fish and Wildl. Serv. Rep. AMNWR 01/10 Homer, Alaska. 78 p.
Pshenichnikova O.S., Klenova A.V., Sorokin P.A., Konyukhov N.B., Andreev A.V., et al., 2017. Population differentiation in whiskered auklets Aethia pygmaea: do diurnal and nocturnal colonies differ in genetics, morphometry and acoustics? // Journal of Avian Biology. V. 48. P. 1047–1061.
Sealy S.G., Bedard J., 1973. Breeding biology of the Parakeet Auklet (Cyclorrhynchus psittacula) on St. Lawrence Island, Alaska // Astarte. V. 6. P. 59–68.
Sowls A.L., Hatch S.A., Lensink C.J., 1978. Catalog of Alaskan seabird colonies. Washington, B.C.: U.S. Fish and Wildl. Serv. 252 p.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал