

Зоологический журнал, 2019, T. 98, № 5, стр. 485-503
Население дождевых червей (Oligochaeta, Lumbricidae) бассейна среднего течения реки Большая Лаба (Северо-Западный Кавказ, буферная зона Кавказского заповедника)
И. Б. Рапопорт 1, *, Н. Л. Цепкова 1, **
1 Институт экологии горных территорий им. А.К. Темботова РАН
360000 Нальчик, Россия
* E-mail: rap-ira777@rambler.ru
** E-mail: cenelli@yandex.ru
Поступила в редакцию 16.03.2018
После доработки 06.10.2018
Принята к публикации 09.11.2018
Аннотация
Впервые изучены видовой состав и структура населения дождевых червей лесных и луговых формаций среднего течения реки Большая Лаба. Среди морфо-экологических форм не отмечены норники, особенно требовательные к хорошему почвенному дренажу. Нетипично редко встречаются крымско-кавказские виды. Эрозионный характер рельефа в сочетании с большим количеством осадков приводят к возрастанию гетерогенности среды обитания и формированию специфичных таксоценов в зависимости от гидроморфного или автоморфного режима почв.
Бассейн среднего течения р. Большая Лаба охватывает горную часть Западного Кавказа. По классификации Соколова, Темботова (1989) рассматриваемая территория расположена в пределах кубанского варианта западно-северокавказского типа поясности. Высокими горными хребтами местность защищена от проникновения в нее холодных северных и северо-восточных ветров. С юго-западной стороны открыт доступ влажным воздушным массам, проникающим с Черного моря. Климат мягкий и влажный. В среднем за год здесь выпадает более 1000 мм осадков. Осень продолжительная, теплая, с обилием солнечных дней. Из-за сложностей орографических условий, пояса широколиственных и темнохвойных лесов четко не выражены и местами один пояс глубоко вклинивается в другой пояс (Олейникова и др., 2008). На склонах троговых долин темнохвойные леса разделяются эрозионными ложбинами с кленом и буком. Бук, вероятно, претерпевший массовые рубки в недавнем прошлом, в виде чистых древостоев встречается редко. На днищах долин лесные сообщества чередуются с полянами с луговой растительностью и кустарниками. Вдоль русел рек и по теневым склонам распространены ольха серая и ольха черная. Береза Литвинова встречается в виде преобладающей породы на низких надпойменных террасах и в верхней части склонов. Многочисленны неглубокие эрозионные ложбины с водной эрозией с кустарниками и элементами субальпийских лугов. В целом флора рассматриваемого района имеет выраженные мезофильные черты (Шильников, 2009).
Цель работы – изучение видового состава, структуры населения и биотопического распределения дождевых червей семейства Lumbricidae в почвах эталонных лесных и луговых фитоценозов буферной зоны Кавказского заповедника.
МАТЕРИАЛ И МЕТОДИКА
Сбор дождевых червей выполнен в июне 2014 г. на границе поясов темнохвойных и широколиственных лесов. Все лесные фитоценозы, в которых проводились почвенно-зоологические работы, для удобства описаний систематизированы по доминирующему виду. Изучены леса с преобладанием ели восточной (Picea orientalis (L.) Link), пихты Нордманна (Abies nordmanniana (Stev.) Spach), бука восточного (Fagus orientalis Lipsky), березы плакучей (Betula pendula Roth), клена остролистного (Acer platanoides L.), ясеня (Fraxinus excelsior L.), ольхи черной (= ольхи клейкой) (Alnus glutinosa (L.)), ольхи серой (Alnus incana (L.) Moench 1794) (табл. 1) и луговые биогеоценозы. В случае, когда фитоценозы отнесены к одной ассоциации, совпадают по ландшафтному положению, а различия между видовым составом и показателями обилия дождевых червей в них находятся в пределах достоверного сходства, данные по нескольким “точкам” объединены в одну выборку. В итоге получилось 3 кленовника, 3 ясенника, 2 букняка, 1 пихтарник (объединены данные по трем биогеоценозам), 5 ельников, 3 сероольшаника, 1 черноольшаник, 3 луговых сообщества. Фитоценотические характеристики указаны в табл. 1. Все исследованные формации произрастают на северо-западном склоне, с разницей в высоте 100 м. Ландшафтное положение изученных биогеоценозов приведено согласно принятой классификации (Исаченко, 1991).
Таблица 1.
Краткая характеристика мест сбора материала в бассейне р. Карасу (Центральный Кавказ)
| № | Фитоценоз | Древесный ярус | Напочвенный покров | Проективное покрытие (ОПП, %) | Сомкнутость крон | Уклон склона, ° |
|---|---|---|---|---|---|---|
| 1 | Кленовник ясеневый | Клен остролистный и ясень обыкновенный; в подлеске лещина обыкновенная | Разнотравный: вербейник обыкновенный, гравилат речной и др. | 35 | 0.6 | 35 |
| 2 | Кленовник ясеневый (эрозионная лощина) | Клен остролистный, ясень обыкновенный; в подлеске лещина обыкновенная | Разнотравный: вербейник обыкновенный, гравилат речной и др. | 40 | 0.6 | 15 |
| 3 | Кленовник грабово-ясеневый с примесью вяза и ели | Преобладают клен полевой, ясень обыкновенный и граб обыкновенный; единично встречаются ель обыкновенная и вяз гладкий; подлесок отсутствует | Разнотравный: подлесник европейский, ясменник кавказский, пахучка обыкновенная, земляника лесная, чистец лесной и др. | 30 | 0.5 | 35 |
| 4 | Ясенник травянистый с примесью сосны | Ясень обыкновенный, сосна обыкновенная; подлесок отсутствует | Разнотравно-бобовый: клевер седоватый, к. предальпийский, горошек обрубленный, вязель восточный, в. пестрый, земляника лесная, пахучка обыкновенная, тысячелистник обыкновенный, душица обыкновенная и др. | 55 | 0.5 | 30 |
| 5 | Ясенник грабово-кленовый | Ясень обыкновенный, граб обыкновенный, клен остролистный, единично груша лесная; подлесок отсутствует | Вейник тростниковидный, мятлик боровой, валериана липолистная, вероника теневая, первоцвет крупночашечковый, вороний глаз неполный | 45 | 0.6 | 10 |
| 6 | Ясенник кленовый | Клен полевой, ясень обыкновенный, единично ель обыкновенная, груша лесная; в подлеске волчник обыкновенный | Разнотравный: ясменник кавказский, первоцвет крупночашечковый, купена душистая, земляника лесная, цицербита кистевидная, чистец лесной и др. | 30 | 0.5 | 35 |
| 7 | Букняк пихтовый | Бук восточный, пихта Нордманна; подлесок отсутствует | Мертвопокровный | – | 0.9 | 5 |
| 8 | Букняк еловый | Бук восточный и ель обыкновенная; подлесок отсутствует | Редкопокровный: единично встречаются подмаренник душистый, кислица обыкновенная, пыльцеголовник длиннолистный, гнездовка обыкновенная, вероника теневая, ожика Форстера | 10 | 0.9 | 3–6 |
| 9 | Пихтарник буковый | Пихта Нордманна, бук восточный, единично – клен остролистный | Мертвопокровный, местами с лишайниковыми и зеленомошными пятнами | – | 1.0 | 3–5 |
| 10 | Ельник сосново-буковый | Ель обыкновенная, сосна обыкновенная и бук восточный; подлесок отсутствует | Редкопокровный: подлесник быкновенный, земляника лесная, подмаренник душистый, вороний глаз неполный | 8–10 | 0.7 | 35 |
| 11 | Ельник | Ель обыкновенная; подлесок отсутствует | Мертвопокровный | – | 0.9 | 5 |
| 12 | Ельник кленово-грабовый | Ель обыкновенная, граб обыкновенный, клен остролистный; подлесок отсутствует | Борщевик шероховатый | 70 | 0.7 | 5 |
| 13 | Ельник буковый | Ель обыкновенная, бук восточный; подлесок отсутствует | Мертвопокровный | – | 0.9 | 40 |
| 14 | Ельник буковый | Ель обыкновенная, бук восточный; подлесок отсутствует | Мертвопокровный | – | 0.9 | 5–7 |
| 15 | Ольшанник кленовый | Ольха серая, клен остролистный | Белокопытник белый | 40 | 0.7 | 10 |
| 16 | Ольшаник | Ольха серая; в подлеске бузина черная | Разнотравный: бутень, хвощ лесной и др. | 35 | 0.9 | 10 |
| 17 | Ольшанник | Ольха серая; подлесок из ежевики кавказской | Папоротниково-ежевичный: страусник обыкновенный (15%) и щитовник мужской (ОПП 7%) | 22 | 0.7 | – |
| 18 | Черноольшанник | Ольха клейкая, или черная | Страусниковый: страусник обыкновенный, единично встречается недотрога | 75 | 0.6 | – |
| 19 | Березняк | Береза плакучая; подлесок отсутствует | Злаково-разнотравный: вейник тростниковидный, коротконожка лесная, мятлик боровой, тимофеевка луговая, первоцвет крупночашечковый, цицербита кистевидная | 30 | 0.5 | 3–5 |
| 20 | Березняк елово-буковый | Береза плакучая, бук восточный и ель обыкновенная, подлесок отсутствует | Злаково-бобово-разнотравный: коротконожка лесная, мятлик боровой, горошек обрубленный и клевер луговой, подмаренник душистый, первоцвет крупночашечковый, земляника лесная, купена душистая и др. | 25 | 0.6 | – |
| 21 | Шпажниково-осоково- разнотравный | – | Средняя высота травостоя 40 см, участвуют: шпажник обыкновенный (опп 20%), осоки (ОПП 35%), разнотравье (ОПП 45%); из разнотравья преобладают влаголюбивые виды – вербейник обыкновенный, черноголовка обыкновенная, гравилат речной, черноголовка обыкновенная и др. | 100 | – | – |
| 22 | Разнотравно-ситниковый | – | Средняя высота травостоя 45 см; сырой участок луга с доминированием ситника развесистого, в разнотравье участвуют влаголюбивые виды – вербейник обыкновенный, гравилат речной и др. | 90 | – | – |
| 23 | Разнотравно-лютиковый | – | Средняя высота травостоя 30 см; доминант лютик ползучий (ОПП 45%), в разнотравье участвуют вербейник обыкновенный, мать-и-мачеха, манжетка, щавель конский, мята кавказская и др. | 65 | – | – |
На пробных площадках 30 × 30 м2 после проведения подробных геоботанических описаний, координатной и ландшафтной привязки к местности отбирали 5–8 проб. Раскопки проводили на глубину встречаемости беспозвоночных. Почвенные монолиты 25 × 25 см2 разбирали вручную (Гиляров, 1975). Исследование полевой влажности и температуры почв проведено с помощью влагомера “TRdiTuroni&c. Snc.”. Содержание гумуса измерено по методу Тюрина в модификации Никитина (Казеев и др., 2004), pHводной вытяжки из почвы – потенциометрически (Аринушкина, 1970).
Определение люмбрицид и характеристика морфо-экологических форм выполнены согласно работам Перель (Перель, 1979; Всеволодова-Перель, 1997). Т.к. население дождевых червей биогеоценозов, выделенных в пределах формаций с единым эдификатором достоверно отличается по видовому составу и другим синэкологическим характеристикам, в работе приведены средние значения численности и соотношения морфо-экологических и хорологических групп для каждого таксоцена отдельно, а не в среднем для формаций.
Для изучения биотопического распределения видов использована степень относительной биотопической приуроченности, вычисленная по формуле: Fij = (nijN – niNj)/(nijN + niNj – 2nijNj), где nij – число особей i-го вида в j-выборке объемом Nj; ni – число особей этого вида во всех сборах объемом N (Песенко, 1982). Степень относительной биотопической приуроченности видов учитывает долю вида в структуре сообществ разных мест обитания и не требует равного объема исследований в разных типах биотопов. Встречаемость вида рассчитана от общего числа проб, встречаемость доминирования – от числа положительных проб.
Индексы биоразнообразия (Шеннона), выровненности (Пиелу) и доминирования (Симпсона) приведены для удобства сравнения сообществ дождевых червей, характеризующихся равным количеством видов, но разным соотношением их численности (Одум, 1975). Коэффициенты позволяют проследить “перекос” в сторону доминирования одного из видов.
Для обработки полученных данных использовали наиболее распространенную иерархическую классификацию – кластерный анализ, результаты которого изображали графически в виде дендрограммы (Халафян, 2007). В кластерном анализе (Ward’smethod, или метод Уорда) в программе “Statistica-10” сравнивали выборки по численности морфо-экологических и хорологических групп.
Краткое описание исследованных биогеоценозов. Кленовники, букняки, ельники и ясенники сформированы на автоморфных почвах. Для почв луговых, ольховых и березовых биогеоценозов характерно близкое залегание грунтовых вод. Пихтарники произрастают на педиментах северо-западного склона.
Ельники, букняки и пихтарники характеризуются высокой сомкнутостью древесного яруса и редким травяным покровом. В ельниках и пихтарниках отмечена самая низкая полевая влажность почв (табл. 2).
Таблица 2.
Видовой состав и синэкологические характеристики населения дождевых червей в бассейне среднего течения р. Большая Лаба
| Растительные сообщества |
Средняя численность видов | Общие показатели обилия | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D. octaedra | D. tellermanica | D. attemsi | O. lacteum | L. rubellus | Ei. tetraedra | D. schmidti | E. fetida | De. rubidus | A. jassyensis | A. c. trapezoides | A. rosea | D. hortensis | D. veneta | Численность, экз./м2 | Биомасса, г/м2 | ||
| Кленовники | 1 | 23 ± 4 | 8 ± 1 | – | 58 ± 8 | – | – | – | – | – | – | – | – | – | – | 89 ± 13 | 22.9 ± 5.1 |
| 2 | 11 ± 3 | – | – | – | – | 16 ± 3 | 11 ± 3 | – | – | – | – | – | – | 38 ± 7 | 16.6 ± 3.2 | ||
| 3 | – | 11 ± 2 | 5 ± 1 | – | – | – | – | – | – | – | – | – | – | – | 16 ± 2 | 3.3 ± 1.2 | |
| Ясенники | 4 | 4 ± 1 | 8 ± 1 | 4 ± 1 | – | – | – | – | – | – | – | – | – | – | – | 16 ± 3 | 5.5 ± 1.0 |
| 5 | – | 11 ± 2 | 21 ± 5 | – | – | – | – | – | – | – | – | – | – | – | 32 ± 7 | 9.3 ± 2.0 | |
| 6 | 5 ± 1 | 5 ± 1 | – | – | – | – | – | – | – | – | – | – | – | – | 11 ± 2 | 0.3 ± 0.1 | |
| Букняки | 7 | – | – | – | 21 ± 5 | – | – | – | – | – | 5 ± 1 | – | – | – | – | 27 ± 5 | 10.7 ± 3.1 |
| 8 | – | 11 ± 4 | – | 5 ± 1 | – | – | – | – | – | – | – | – | – | 16 ± 3 | 6.1 ± 1.8 | ||
| Пихтарники | 9 | – | – | – | 27 ± 5 | 16 ± 3 | – | 27 ± 5 | – | – | – | – | – | – | – | 70 ± 5 | 19.6 ± 5.8 |
| Ельники | 10 | – | – | – | – | – | – | – | – | 10 ± 2 | – | – | – | – | – | 10 ± 2 | 2.3 ± 0.6 |
| 11 | 8 ± 1 | – | 8 ± 2 | – | – | – | – | – | 4 ± 1 | – | – | – | – | – | 20 ± 2 | 2.6 ± 0.7 | |
| 12 | 36 ± 6 | – | 24 ± 3 | – | – | 12 ± 2 | – | 12 ± 2 | 8 ± 1 | – | – | – | – | – | 92 ± 4 | 4.6 ± 0.5 | |
| 13 | – | 12 ± 2 | – | – | – | – | – | – | – | – | – | – | – | – | 12 ± 2 | 2.7 ± 0.6 | |
| 14 | – | – | – | – | – | – | 5 ± 1 | – | – | – | – | – | 11 ± 3 | – | 16 ± 4 | 2.5 ± 0.4 | |
| Сероольшаники | 15 | 5 ± 1 | 11 ± 3 | 5 ± 2 | – | – | – | 16 ± 2 | – | – | – | – | – | – | (ед.) | 37 ± 5 | 28.4 ± 9.8 |
| 16 | – | – | 128 ± 41 | 150 ± 43 | 11 ± 3 | – | – | – | – | – | 5 ± 2 | – | – | 293 ± 48 | 109.7 ± 28.3 | ||
| 17 | 5 ± 1 | – | 5 ± 1 | 5 ± 2 | 11 ± 3 | – | – | – | – | – | – | – | – | 27 ± 4 | 5.8 ± 1.4 | ||
| Черноольшаник | 18 | 5 ± 1 | – | – | – | – | – | – | 5 ± 2 | – | – | – | – | – | – | 11 ± 3 | 3.7 ± 1.0 |
| Березняки | 19 | 11 ± 3 | 21 ± 4 | – | 59 ± 6 | 21 ± 6 | – | – | – | – | – | – | – | – | – | 112 ± 20 | 37.5 ± 7.9 |
| 20 | 16 ± 3 | – | 6 ± 2 | 23 ± 4 | – | – | 11 ± 2 | – | – | – | – | – | – | – | 55 ± 4 | 24.0 ± 6.1 | |
| Луга | 21 | 80 ± 12 | – | – | 10 ± 2 | – | – | – | – | – | – | – | – | – | – | 90 ± 13 | 11.4 ± 3.2 |
| 22 | – | – | – | – | 64 ± 8 | – | 32 ± 4 | – | – | – | 11 ± 3 | – | – | – | 106 ± 6 | 43.97 ± 12.0 | |
| 23 | – | – | – | – | 64 ± 12 | 16 ± 1 | – | – | – | – | – | – | – | – | 80 ± 13 | 41.2 ± 15.1 | |
Примечания. Биогеоценозы с доминированием: клена остролистного (Ко): 1 – кленовник ясеневый разнотравный-1, 2 – кленовник ясеневый разнотравный-2, 3 – кленовник грабово-ясеневый с примесью вяза и ели разнотравный; ясеня обыкновенного (Яо): 4 – ясенник травянистый с примесью сосны, 5 – ясенник грабово-кленовый разнотравный, 6 – ясенник кленовый разнотравный; бука восточного (Бв): 7 – букняк пихтовый мертвопокровный, 8 – букняк еловый редкопокровный, 9 – пихтарник буковый мертвопокровный; ели восточной (Ев): 10 – ельник сосново-буковый редкопокровный, 11 – ельник мертвопокровный, 12 – ельник кленово-грабовый с борщевиком, балка, 13 – ельник буковый мертвопокровный-1, СЗ, 14 – ельник буковый мертвопокровный-2, С-СЗ; ольхи серой (Ос): 15 – ольшаник кленовый белокопытниковый, 16 – ольшаник разнотравный, 17 – ольшаник папоротниково-ежевичный; ольхи черной (Оч): 18 – черноольшаник страусниковый; березы плакучей (БЛ): 19 – березняк злаково-разнотравный, 20 – березняк елово-буковый злаково-бобово-разнотравный; луга: 21 – шпажниково-осоково-разнотравный, 22 – разнотравно-ситниковый, 23 – разнотравно-лютиковый. Жирным шрифтом выделены доминирующие виды и экстремальные значения общей численности и биомассы.
Ельники произрастают на горно-лесных темно-бурых почвах с признаками оподзоливания. Для этих почв характерны маломощная (до 1–3 см) лесная подстилка, сменяющаяся гумусовым горизонтом (3–5 см), глубже которого залегает резко осветленный подзолистый горизонт с серовато-палевой окраской и непрочной ореховато-комковатой структурой. Почвы изученных ельников имеют слабокислую реакцию и высокое содержание гумуса. В июне они прогреваются меньше, чем под другими породами по причине высокой сомкнутости древесного яруса (табл. 1). Изученные ельники – мертвопокровные, монодомининтые, или редкопокровные сообщества.
Буковые леса сформированы на бурых лесных остаточно-оподзоленных почвах с хорошим дренажем и протяженным гумусовым горизонтом (25 см). По отношению к влаге бук восточный является мезофитом, плохо переносит переувлажнение и заболачивание (Погребняк, 1955). В почвах букняков содержится меньше гумуса, чем в ельниках, но прогреваются они сильнее. Мощность лесной подстилки 3–6 см. Сопутствуют буку пихта Нордманна и ель обыкновенная, напочвенный покров редок, местами не выражен (табл. 1).
Почвы пихтарников – горно-лесные бурые среднемощные, развиты в нижней части склонов и террас, характеризуются преимущественно близкой к нейтральной или слабокислой реакцией почвенного раствора (табл. 2), слой А0 – 2 см. Процентное содержание гумуса ниже, чем в почвах еловых и буковых формаций. В древесном ярусе встречается бук восточный, единично – клен остролистный, все исследованные пихтарники – мертвопокровные (табл. 1).
Ясень обыкновенный и клен остролистный образуют смешанные древостои. Оба вида предпочитают глубокие, рыхлые, плодородные, глинисто-песчаные почвы (Овсяников, 1931; Зонн, 1950; Справочник лесничего, 2003). Содержание гумуса под ясенем немного выше, чем под кленом и является самым высоким в исследованном районе (табл. 2). Реакция почвенного раствора в кленовниках слабощелочная, в ясенниках – слабокислая, подстилка 2–3 см. Клен доминирует на более затеняемых участках. Кленовники сформированы преимущественно в эрозионных ложбинах, в рассматриваемый период в большинстве из них отмечена самая высокая полевая влажность. Фитоценозы с преобладанием ясеня образованы на опушках, или светлых полянах, в них отмечена примесь дикоплодных деревьев. Травяной покров богаче, чем в кленовниках (табл. 1). Сомкнутость древесного яруса таких сообществ меньше, чем в кленовниках, поэтому почва прогревается сильнее (табл. 2).
Исследованные гидроморфные почвы характеризуются меньшей гумусированностью (табл. 2). Березняки произрастают на речных террасах на горно-аллювиальных маломощных почвах со значительным содержанием песчаной фракции, глубина А0 2–3 см. Прогреваются они почти также, как под ясенем и отличаются самыми низкими полевой влажностью и содержанием гумуса. Наряду с березой в древесном пологе бук восточный и ель обыкновенная, подлесок отсутствует. Травяной покров представлен злаками и разнотравьем, проективное покрытие не высокое, как и сомкнутость древесного яруса (табл. 1).
Черноольховые формации сформированы в условиях постоянного избыточного увлажнения на глинистых слабокислых иловато-глеевых почвах, сероольшаники – на среднемощных глинисто-супесчаных гидроморфных почвах (речные террасы) и теневых склонах. Подстилка в сообществах ольхи серой и ольхи черной 3–5 см. Черноольшаник выделяется заболоченностью, самой низкой гумусированностью почв и наиболее выраженной слабощелочной реакцией почвенного раствора. Из-за высокой плотности древесного яруса и насыщенности почвы влагой, его почва прогревается меньше, чем в сообществах ольхи серой. В сероольшаниках с низкой сомкнутостью древесного яруса и высоким проективным покрытием травостоя отмечен максимум текущей температуры и нейтральная реакция водной вытяжки из почвы. В обеих формациях обычны папоротники и разнотравье.
Луга занимают глинисто-супесчаные гидроморфные почвы с почти нейтральной реакцией рН и близкой к верхнему экстремуму температурой верхнего почвенного слоя. Исследованы: шпажниково-осоково-разнотравный, разнотравно-ситниковый и разнотравно-лютиковый биогеоценозы. Травостой 40–45 см, велика представленность влаголюбивых видов (табл. 1).
РЕЗУЛЬТАТЫ
Общая характеристика фауны дождевых червей среднего течения реки Большая Лаба
В пределах рассматриваемой территории отмечены 14 видов дождевых червей: Aporrectodea caliginosa trapezoides (Dugès 1828), A. jassyensis (Michaelsen 1891), A. rosea (Savigny 1826), Dendrobaena attemsi (Michaelsen 1903), D. hortensis (Michaelsen 1890), D. octaedra (Savigny 1826), D. schmidti Michaelsen 1903, D. tellermanica Perel 1966, D. veneta (Rosa 1886), Dendrodrilus rubidus tenuis (Eisen 1874), Lumbricus rubellus (Hoffmeister 1843), Eisenia fetida (Savigny 1826), Eiseniella tetraedra tetraedra (Savigny 1826), Octolasion lacteum (Örley 1885) (табл. 3). Из них D. veneta зарегистрирован лишь однажды – в произвольных сборах на берегу р. Большая Лаба под камнями в почве кленово-ольхово-белокопытникового сообщества. Наибольшее разнообразие – шесть видов – отмечено у рода Dendrobaena Eisen 1973, три вида относятся к роду Aporrectodea Örley 1885, у остальных пяти родов выявлено по одному виду.
Таблица 3.
Степень относительной биотопической приуроченности (СБтП) и встречаемость (%) дождевых червей в бассейне среднего течения р. Большая Лаба
| Растительные сообщества | Вид/показатели | D. octaedra | D. tellermanica | D. attemsi | O. lacteum | L. rubellus | E. tetraedra | D. schmidti | E. fetida | De. rubidus | A. jassyensis | A. c. trapezoides | A. rosea | D. hortensis |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Кленовники | СБтП | 0.28 | 0.33 | –0.15 | 0.17 | –1.00 | 0.54 | –1.00 | 0.65 | –1.00 | –1.00 | –1.00 | –1.00 | –1.00 |
| Встречаемость | 44 | 66 | 33 | 22 | 0 | 22 | 0 | 22 | 0 | 0 | 0 | 0 | 0 | |
| Ясенники | СБтП | 0.04 | 0.75 | 0.87 | –1.00 | –1.00 | –1.00 | –1.00 | –1.00 | –1.00 | –1.00 | –1.00 | –1.00 | –1.00 |
| Встречаемость | 66 | 84 | 66 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Ельники | СБтП | 0.37 | 0.04 | 0.72 | –1.00 | –1.00 | 0.35 | –0.52 | 0.62 | 1.00 | –1.00 | –1.00 | –1.,00 | 1.00 |
| Встречаемость | 40 | 20 | 40 | 0 | 0 | 0,2 | 40 | 20 | 60 | 0 | 0 | 0 | 20 | |
| Сероольшаники | СБтП | –0.74 | –0.48 | –0.30 | 0.14 | 0.39 | 0.27 | –0.45 | –1.00 | –1.00 | –1.00 | –1.00 | 1.00 | –1.00 |
| Встречаемость | 55 | 33 | 60 | 60 | 30 | 66 | 30 | 0 | 0 | 0 | 0 | 30 | 0 | |
| Черноольшаник | СБтП | 0.57 | –1.00 | –1.00 | –1.00 | –1.00 | –1.00 | –1.00 | 0.95 | –1.00 | –1.00 | –1.00 | –1.00 | –1.00 |
| Встречаемость | 80 | 0 | 0 | 0 | 0 | 0 | 0 | 80 | 0 | 0 | 0 | 0 | 0 | |
| Пихтарники | СБтП | –1.00 | –1.00 | –1.00 | 0.15 | –0.03 | –1.00 | 0.76 | –1.00 | –1.00 | –1.00 | –1.00 | –1.00 | –1.00 |
| Встречаемость | 0 | 0 | 0 | 75 | 75 | 0 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Березняки | СБтП | 0.06 | 0.33 | –0.15 | 0.30 | –0.36 | –1.00 | –0.24 | –1.00 | –1.00 | –1.00 | –1.00 | –1.00 | –1.00 |
| Встречаемость | 100 | 50 | 50 | 100 | 50 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Букняки | СБтП | –1.00 | 0.60 | –1.00 | 0.37 | –1.00 | –1.00 | –1.00 | –1.00 | –1.00 | 1.00 | –1.00 | –1.00 | –1.00 |
| Встречаемость | 0 | 40 | 0 | 70 | 0 | 0 | 0 | 0 | 0 | 50 | 0 | 0 | 0 | |
| Луга | СБтП | 0.44 | –1.00 | –1.00 | –0.82 | 0.42 | 0.21 | 0.09 | –1.00 | –1.00 | –1.00 | 1.00 | –1.00 | –1.00 |
| Встречаемость | 25 | 0 | 0 | 30 | 66 | 30 | 30 | 0 | 0 | 0 | 33 | 0 | 0 | |
| Общая встречаемость вида | 48 | 39 | 39 | 35 | 22 | 22 | 22 | 13 | 13 | 4 | 4 | 4 | 4 | |
| Встречаемость доминирования вида в положительных пробах | 54 | 78 | 22 | 62 | 80 | 40 | 40 | 67 | 33 | 0 | 0 | 0 | 1 | |
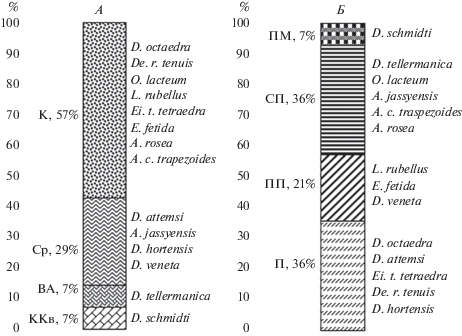
В исследованных сообществах региональных эндемиков не зарегистрировано, D. schmidti – крымско-кавказский субэндемик, D. tellermanica – близкородственный D. schmidti вид с дизъюнктивным восточноевро-азиатским ареалом. Средиземноморским распространением характеризуются четыре вида, 8 видов – космополиты (рис. 1А). Равное число видов относится к морфо-экологическим группам подстилочных и собственно почвенных люмбрицид (рис. 1Б). Полиморфный D. schmidti представлен собственно-почвенной, почвенно-подстилочной и наиболее редкой подстилочной морфо-экологическими формами, отличающимися глубиной стратификации в почвенном профиле, особенностями фенотипа и временем ответной реакции на раздражение. Норники в рассматриваемом районе нами не зарегистрированы.
Рис. 1.
Разнообразие хорологических (А) и морфо-экологических (Б) групп дождевых червей в бассейне р. Большая Лаба: А– хорологические группы: ВА – восточноевро-азиатская, СР – средиземноморская, ККв – крымско-кавказская, К – космополиты; Б – жизненные формы: П – подстилочный, ПП – почвенно-подстилочный, СП – собственно почвенный, Н – норник, ПМ – полиморфный вид.
Население дождевых червей
Биотопическое распределение. Состав доминантов в сообществах дождевых червей среднего течения р. Большая Лаба отличался значительным разнообразием (табл. 3). Наиболее часто преобладали D. octaedra, O. lacteum и D. tellermanica (доминирование этих видов отмечено в 6, 6 и 5 таксоценах соответственно). Экстремумы плотности D. octaedra отмечены в почвах шпажниково-ситниково-разнотравного луга (около 80 экз./м2) и ельника кленово-грабового (в среднем 36 экз./м2). Преобладал по численности этот вид в кленовнике ясеневом разнотравном, ясеннике кленовом разнотравном, ельнике мертвопокровном и черноольшанике страусниковом. В расположенном на крутом участке склона кленовнике ясеневом разнотравном, ясеннике травянистом, ольшанике кленовом белокопытниковом и в ольшанике папоротниково-ежевичном D. octaedra не относился к числу доминантов, однако регистрировался часто.
Доминирование O. lacteum отмечено в лесных сообществах, максимальная численность наблюдалась в ольшанике разнотравном (в среднем 128 экз./м2), березняке злаково-разнотравном и расположенном в эрозионной ложбине кленовнике ясеневом разнотравном (в обоих случаях почти 60 экз./м2). Преобладал по численности этот вид в пихтарнике мохово-лишайниковом, букняке пихтовом мертвопокровном и березняке елово-буковом, со средним обилием 20–30 экз./м2.
Численность D. tellermanica в исследованных таксоценах изменялась меньше, чем выше приведенных видов (5–12 экз./м2). Доминировал D. tellermanica в почвах кленовника грабово-ясеневого разнотравного, ельника букового мертвопокровного и ольшаника кленового белокопытникового. Однако наиболее высокое обилие отмечено в березняке злаково-разнотравном, где этот вид не относился к числу доминантов.
Значительно реже – в двух–трех сообществах – преобладали L. rubellus, D. attemsi, Eiseniella tetraedra tetraedra и E. fetida. L. rubellus был наиболее обилен (64–150 экз./м2) в тех таксоценах (ольшаника разнотравного и обоих лугов), в которых являлся доминантом. Со значительно меньшим обилием отмечен в пихтарнике и березняке злаково-разнотравном. У остальных перечисленных видов столь высоких экстремумов численности не зарегистрировано. D. attemsi преобладал в почве ясенника грабово-кленовогои ельника мертвопокровного (7–28 экз./м2), встречался в кленовнике грабово-ясеневом разнотравном, березняке елово-буковом, ольшаннике кленовом белокопытниковом, ясеннике травянистом с примесью сосны, ольшанике папоротниково-ежевичном и ельнике кленово-грабовом с борщевиком. Eiseniella tetraedra tetraedra доминировал в расположенном в ложбине кленовнике ясеневом разнотравном и ольшанике папоротниково-ежевичном (9–18 экз./м2). Сравнимое обилие вида отмечено в ельнике кленово-грабовом с борщевиком, ольшанике разнотравном и на разнотравно-лютиковом лугу, где он к числу доминантов не относится. E. fetida из перечисленных видов наиболее редок, преобладает по численности в почвах кленовника ясеневого (эрозионная лощина) и черноольшаника страусникового, зарегистрирован в ельнике кленово-ясенево-грабовом с борщевиком.
Dendrodrilus rubidus tenuis собран исключительно в ельниках, в одном из которых (ельник сосново-буковый редкопокровный) отмечено его доминирование. В почве ельника буковового мертвопокровного, расположенного на более пологом склоне, преобладает по численности другой редкий вид – D. hortensis. Остальные четыре вида встречаются локально.
Биотопическое видовое богатство. Максимальное число – 5 видов дождевых червей – отмечено в почве ельника кленово-грабового (табл. 3), травяной покров в котором представлен почти исключительно борщевиком. В целом же еловые таксоцены – сообщества с наиболее изменчивым таксономическим разнообразием: в двух из них (в ельнике буковом мертвопокровном и ельнике сосново-буковом редкопокровном) отмечено по 1 виду люмбрицид. Стабильно высоко – 4 вида – видовое богатство сероольшаников. В остальных сообществах на выделенных площадках зарегистрированы по 2–3 вида дождевых червей.
Встречаемость и степень относительной биотопической приуроченности дождевых червей. Показатель встречаемости вида и степень относительной биотопической приуроченности отражают разные аспекты экологии вида. Степень относительной биотопической приуроченности учитывает обилие видов, тогда как встречаемость характеризует равномерность их распределения в пространстве. Как видно из табл. 4 , наиболее часто в рассмотренных сообществах встречаются D. octaedra, D. tellermanica, D. attemsi и O. lacteum. Однако частота доминирования, рассчитанная по числу положительных проб, совпадает с высоким показателем встречаемости вида только у D. tellermanica и O. lacteum. Несмотря на более низкие значения встречаемости, L. rubellus и E. fetida преобладают по численности в большинстве положительных проб, что, возможно, свидетельствует об их более высокой избирательности к условиям местообитаний.
Степень относительной биотопической приуроченности варьирует от –1 до 1 (табл. 4 ), причем для многих видов ее значения близки к нулю в большинстве биогеоценозов. Высокие значения степени относительной биотопической приуроченности отмечены у часто встречающихся видов – D. tellermanica и D. attemsi. Не всегда степень относительной биотопической приуроченности вида коррелирует с данными по встречаемости. Так, например, в кленовниках, ясенниках, сероольшанниках и березняках D. octaedra отмечен в 44–100% собранных проб, но из-за низкой численности не показывает взаимосвязи с этими сообществами (табл. 3, 4 ). В то же время редкие виды A. caliginosa trapezoides, A. rosea, D. hortensis, D. veneta несмотря на высокую степень относительной биотопической приуроченности не могут считаться специфичными для формаций, в которых они найдены, т.к. зарегистрированы только в одном биогеоценозе, хотя и во всех пробах.
В том случае, если высокая степень относительной биотопической приуроченности совпадает с экстремумами встречаемости вида, можно говорить о том, что вид характерен для формации в целом. В данном случае для ельников были характерны D. tellermanica и Dendrodrilus rubidus tenuis, для ясенников – D. tellermanica и D. attemsi, для пихтарников – D. schmidti и черноольшаника – E. fetida. На уровне тенденции к букнякам тяготели D. tellermanica и A. jassyensis, к – черноольшаникам D. octaedra, к лугам – L. rubellus. Наименьшая связь с эдификатором биогеоценозов в наших сборах отмечена у O. lacteum и L. rubellus.
Наиболее обширным спектром специфичных видов выделяются ельники и ясенники. Минимальная связь с эдификатором фитоценоза отмечена у дождевых червей, обитающих в березовых лесах.
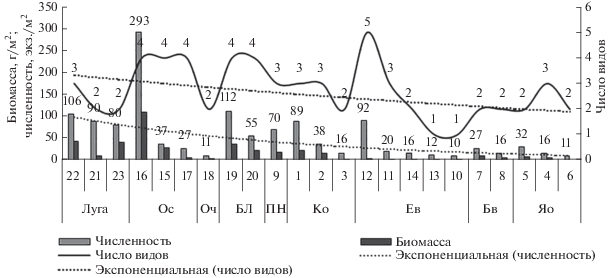
Показатели обилия. Средние численность и биомасса дождевых червей варьировали в широком диапазоне (рис. 2). Максимум показателей обилия отмечен в ольшанике разнотравном, минимум – в ельнике сосново-буковом редкопокровном. Верхние экстремумы численности и биомассы дождевых червей, выявленные в пределах формаций с единым эдификатором, часто совпадают с максимальным биотопическим видовым богатством (табл. 3), как например, в ельнике кленово-ясенево-грабовом с борщевиком. Основной тренд снижения средних показателей обилия дождевых червей направлен от формаций, сформированных на гидроморфных почвах (лугов → ольшаников → березняков) к автоморфным почвам (рис. 2), однако стабильными показателями выделяются только луговые таксоцены, остальные данные высоко вариабельны. У люмбрицид ельников, ясенников и букняков, сформированных на автоморфных почвах, – численность и биомасса невелики.
Рис. 2.
Показатели обилия дождевых червей бассейна среднего течения р. Большая Лаба. Обозначения фитоценозов с 1 по 23 – как и в табл. 1.
ОБСУЖДЕНИЕ
На небольшой территории среднего течения р. Большая Лаба зарегистрированы около 35% всех видов люмбрицид, известных в настоящее время для Северного Кавказа (Перель, 1979; Всеволодова-Перель, 1997; Рапопорт, 2013, 2014; Рапопорт, Комаров, 2017). Население дождевых червей небольшой территории бассейна среднего течения реки Большая Лаба – 14 видов люмбрицид из 7 родов – разнообразно в таксономическом и экологическом аспектах. 7 видов относятся к роду Dendrobaena, который является вторым после рода Eisenia по числу видов, представленных в фауне РФ (Всеволодова-Перель, 1997), и одним из ведущих по числу видов, зарегистрированных на Северном Кавказе (Всеволодова-Перель, 1997). Омодео (Omodeo, 1952) первоначально считал род Dendrobaena кавказским из-за большого видового разнообразия в регионе представителей этого рода, однако более детальное изучение фауны дождевых червей средиземноморских и субсредиземноморских территорий лишило Кавказский перешеек звания центра происхождения одного из наиболее многочисленных родов люмбрицид – только в северо-восточном Средиземноморье отмечено более 60 видов этого рода (Szederjesi, 2017).
Все люмбрициды ранее отмечены на Северо-Западном Кавказе (Малевич, 1976; Перель, 1979; Всеволодова-Перель, 1997; Квавадзе, 1985; Рапопорт, 2014). Основная частьиз них образует лесной комплекс, распространенный на Кавказе от лесостепного пояса до нижней границы субальпики (Квавадзе, 1985; Перель, 1979; Всеволодова-Перель, 1997; Рапопорт, 2013). Это бореальные D. octaedra и Dendrodrilus rubidus tenuis, неморальные L. rubellus, E. fetidaи Eiseniella tetraedra tetraedra, а также D. veneta и D. hortensis. Амфибионт Eiseniella tetraedra tetraedra, найденный вне пойменных почв, вероятно, попал в эрозионные ложбины в кленовниках, ельниках и на лугах с весенне-раннелетними временными водотоками. Неморально-степной A. jassyensis на Северном Кавказе тяготеет к экотонным сообществам и послелесным лугам. Неожиданным результатом исследования оказалась редкая встречаемость полизонального D. schmidti, преобладающего на этой высоте в большинстве сообществ Северного Кавказа (Перель, 1979; Рапопорт, 2013, 2014; Рапопорт и др., 2017). Неморально-степной характер распространения присущ D. tellermanica, O. lacteum, A. caliginosa trapezoides и A. rosea. Последние два вида редки и отмечены только в одном биогеоценозе.
Не менее шести видов люмбрицид относятся к автохтонной кавказской фауне. Помимо крымско-кавказского субэндемика D. schmidti, это D. tellermanica – вид кавказского происхождения, отмеченный не только на Кавказе (в том числе на юго-западном подножье Малого Кавказа в пределах Турции, где также зарегистрирован D. schmidti (Csuzdietal., 2006; Szederjesi, 2017)) и в Крыму, но и на Среднерусской возвышенности, Южном Урале, Алтае и в Казахстане (Всеволодова-Перель, 1997), а также представители наиболее древней средиземноморской фауны Кавказа D. hortensis, D. veneta, D. attemsi и A. jassyensis (Оmodeo, 1952). Основной ареал последних четырех видовлежит в пределах Средиземноморской области Голарктики (Перель, 1979). Из-за использования в вермикультурах D. veneta и D. hortensis в настоящее время распространились более широко, поэтому ряд авторов считают их голарктами или даже космополитами (Stojanović, Karaman, 2005; Сsuzdi, Zicsi, 2003; Csuzdietal., 2006). Как и везде на Северном Кавказе, самой многочисленной оказалась группа всесветно распространенных видов. На территории среднего течения р. Большая Лаба не обнаружены виды с западно-палеарктическим распространением (род Helodrilus), известные для Северного Кавказа по сравнительно небольшому числу находок (Перель, 1979; Квавадзе, 1985; Проконова, 2005; Рапопорт, Комаров, 2017).
Основываясь на представлениях о том, что фитоценотический облик ландшафта является отражением эдафо-климатических условий местообитаний (Раменский, 1938; Сукачев, 1942; Погребняк, 1955 и др.) и о том, что наиболее информативной характеристикой условий произрастания фитоценоза является вид эдификатора, средообразующая деятельность которого также в значительной степени формирует среду обитания беспозвоночных, в графиках представлены биогеоценозы в порядке, объединяемом по доминирующей породе. Таксоны с космолитным типом ареала преобладают не только по видовому богатству, но и по обилию, особенно многочисленны они в луговых, ольховых и березовых сообществах (рис. 3). Однако, как и везде в горах, среда обитания отличается мозаичностью, свойственной как строению древесного и кустарникового ярусов, так и ландшафтному положению (табл. 1, 2). Поэтому общие закономерности наблюдаются в виде тенденции и данные приводятся по каждому биогеоценозу отдельно. Меньше всего космополитных люмбрицид зарегистрировано в ясенниках. Средиземноморские виды и восточноевро-азиатский D. tellermanica преимущественно локализованы в автоморфных почвах ясенников и ельников, элиминируются в пихтарниках, где доминирует крымско-кавказский субэндемик D. schmidti, и на лугах. Ольшаники, березняки и луга в бассейне среднего течения реки Большая Лаба сформированы на гидроморфных почвах. Из-за расположения на педиментах склонов, относительно стабильный режим увлажнения, вероятно, имеют и пихтарники. В рассматриваемом районе D. schmidti показывает преференцию именно к гидроморфным почвам. Небольшая представленность в сборах крымско-кавказского субэндемика D. schmidti может объясняться несколькими причинами. Во-первых, лабильностью вида, позволяющей переживать неблагоприятные периоды у корней деревьев и на возвышенных микростациях (Рапопорт, 2011), благодаря этой лабильности, не смотря на большую биомассу и локализацию в верхних почвенных слоях, D. schmidti не “смывается” водными потоками на относительно ровные формы рельефа, но и не попадается в июньских пробах. Во-вторых, не изучена его способность переносить амплитудный режим увлажнения. В степной зоне D. schmidti замещается близкородственным восточноевропейско-азиатским D. tellermanica (Рапопорт, 2011), возможно, подобное замещение происходит и в данном случае.
Рис. 3.
Представленность хорологических групп в бассейне среднего течения р. Большая Лаба. Обозначения фитоценозов с 1 по 23 – как и в табл. 1.
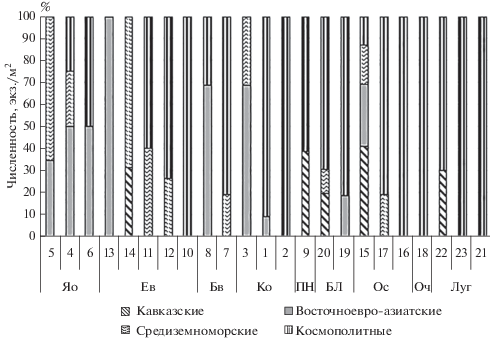
В хорологическом отношении наиболее разнородны таксоцены дождевых червей ельников, собственно кавказский элемент населения в которых представлен восточноевро-азиатскими и средиземноморскими видами. В ряде сообществ, например в ельнике сосново-буковом редкопокровном, обнаружены только космолиты; преобладают так называемые “перегринные”, по выражению Михаэльсена (Michaelsen,1903), виды в ельнике мертвопокровном и ельнике кленово-грабовом с борщевиком.
Представители всех хорологических групп дождевых червей отмечены только в одном биогеоценозе – ольшаннике кленовом белокопытниковом. В этом случае, вероятно, значительную роль играет экотонное – на границе лесных и луговых сообществ – положение биогеоценоза.
В бассейне среднего течения р. Большая Лаба у ряда видов отмечена высокая степень относительной биотопической приуроченности (табл. 3). Биотопическая преференция отмечена у крымско-кавказского эндемика D. schmidti, восточноевро-азиатского D. tellermanica и средиземноморского D. attemsi (табл. 3). Наиболее эвритопными оказались экологически пластичные виды с космополитным типом ареала. Наименьшая связь с эдификатором биогеоценозов в наших сборах отмечена у O. lacteum и L. rubellus. O. lacteum встречается в широком спектре экологических условий, судя по встречаемости, это преимущественно лесной вид. L. rubellus на этой высоте приурочен к лесным местообитаниям с гидроморфными почвами, что, вероятно, объясняется его недостаточной морозостойкостью (Мещерякова, Берман, 2014). Однако и у некоторых космополитных дождевых червей выявлена приуроченность к определенному типу формаций. Так, Dendrodrilus rubidus tenuis отмечен в 60% проб, собранных в ельниках, и не найден за пределами этих растительных сообществ. Не в полной мере можно говорить о предпочитаемости почв черноольховых формаций E. fetida, т.к. исследован только один биогеоценоз. Однако находка теплолюбивого вида в гидроморфных почвах черноольшаника закономерна – обилие незамерзающих проточных водоемов позволяет E. fetida пережить в этих сообществах сезонные колебания температуры и влажности.
Отличительная черта морфо-экологической структуры дождевых червей изученных сообществ – отсутствие в сборах норников. Представители этой морфо-экологической группы требовательны к хорошему почвенному дренажу. При учете средней численности видов, относящихся к разным морфо-экологическим группам, выявляются два варианта сообществ (рис. 4). В автоморфных почвах основную долю в общей численности дождевых червей наиболее часто составляют представители одной морфо-экологической группы: в букняках и кленовниках это собственно почвенные виды, в ельниках – подстилочные.
Рис. 4.
Представленность хорологических групп в бассейне среднего течения р. Большая Лаба. Обозначения фитоценозов с 1 по 23 – как и в табл. 1.
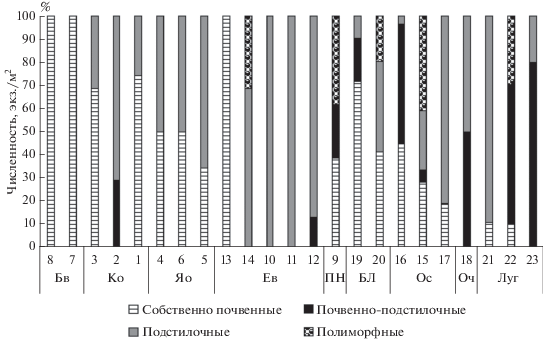
Исследованные кленовники расположены наиболее часто в проточных водосборных понижениях и эрозионных ложбинах на крутых склонах, поэтому под влиянием обильных весенне-раннелетних поверхностных водотоков более благоприятные условияв почвах этих биогеоценозов складываются для собственно почвенных люмбрицид. Многочисленны виды собственно почвенной морфо-экологической группы и в буковых биогеоценозах, почвы которых отличаются хорошим дренажем. Небольшая представленность люмбрицид, питающихся на поверхности почвы, обусловлена так же мощным слоем буковой подстилки, разлагающейся медленнее опада других пород (Вальтер, 1974). Несмотря на присутствие в кишечнике дождевых червей дрилодефензина, играющего важную роль в разложении полифенолов (Liebeke et al., 2015), по предпочитаемости дождевыми червями буковый опад занимает одно из последних мест, уступая только опаду хвойных деревьев (Стриганова, 1980).
В почвах ельников выраженный оподзоленный горизонт (Зонн, 1950) с низкой влажностью и комковатой структурой приводит к концентрации жизни в подстилке и верхнем почвенном слое, обусловливая большое разнообразие подстилочных люмбрицид. В ясенниках преобладание видов, относящихся к этой морфо-экологической группе, вероятно связано с повышенной инсоляцией сообществ (не благоприятной для почвенно-подстилочных видов), и вхождением в состав древесного яруса дикоплодных деревьев. Обилие подстилочных видов, долговременная экспозиция более высоких текущих температур определяют в свою очередь лучшую, по сравнению с кленовниками, гумусированность почв (табл. 2).
Отличительная черта населения гидроморфных почв – более сложная морфо-экологическая структура, а также более частая представленность в сборах полиморфного D. schmidti и почвенно-подстилочных люмбрицид (рис. 4). Вероятно, этот факт связан с общей мозаичностью микроусловий, возникающих в гидроморфных почвах и в целом более благоприятным их характером для такой влаголюбивой группы, как дождевые черви.
Наиболее сбалансирована морфо-экологическая структура пихтовых и березовых таксоценов. Структура населения ольховых и луговых сообществ люмбрицид изменчива и, наряду с другими факторами, зависит от глубины залегания грунтовых вод и типа предшествующих растительных формаций. Косвенным свидетельством вторичности шпажниково-ситниково-разнотравного луга является высокая численность в сборах типично лесного вида D. octaedra. Ольховые фитоценозы, как и луга, могут являться одной из сукцессионных стадий, следующих за сведением бука (Нечаев, 1960; Шхагапсоев, Волкович, 2002).
Достаточно часто морфо-экологическая и хорологическая структура таксоценов выделяется из общего тренда особенностей, характерных для населения почв формаций с единым эдификатором. Так, например, в почве буково-елового мертвопокровного леса найден только один вид – D. tellermanica, что, вероятно, связано, с крутосклонным положением местообитания. В целом же большинство растительных сообществ являются сложносоставными как по структуре древесного и кустарникового яруса, так и характеру микрорельефа (табл. 1, 2). Для дождевых червей – представителей крупной почвенной мезофауны, но мелкоразмерных в общем-то организмов, определяющим может быть наличие небольшой зеленомошной синузии, куртины злаков, валежника на определенной стадии разложения и других локальных особенностей биогеоценоза для формирования условий обитания вида. Вероятно, совокупность множества неучтенных факторов и оказывает влияние на население дождевых червей в среднегорных сообществах бассейна реки Большая Лаба.
Индексы Шеннона, Симпсона и Пиелу. Основные тренды увеличения индексов биоразнообразия и выровненности направлены от таксоценов дождевых червей, обитающих в автоморфных почвах (букняков → ельников → кленовников → ясенников), к населению гидроморфных почв ольшаников и березняков, среди которых также оказываются таксоцены пихтарников (рис. 5).
Рис. 5.
Индексы биоразнообразия (Шеннона), доминирования (Симпсона) и выровненности (Пиелу) дождевых червей в бассейне р. Большая Лаба. Обозначения фитоценозов с 1 по 23 – как и в табл. 1.
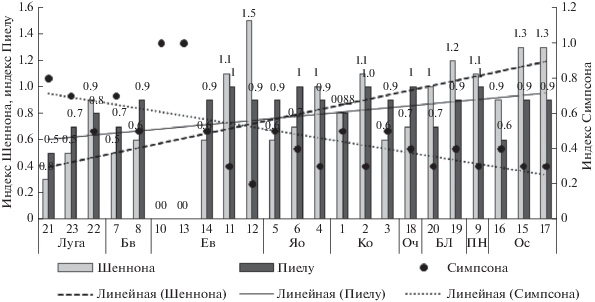
Несмотря на то, что луга также занимают гидроморфные почвы, луговые таксоцены отличаются низкими индексами биоразнообразия, что связано со значительным превышением обилия отдельных видов.
Наиболее изменчивы индексы Шеннона у дождевых червей ельников. Именно в ельниках отмечены как максимум (ельник кленово-грабовый с борщевиком), так и минимум (ельник сосново-буковый редкопокровный и ельник букововый мертвопокровный) показателя. Верхний экстремум индекса выровненности зарегистрирован в пихтарнике буковом, в котором мертвопокровные участки чередуются с мохово-лишайниковыми пятнами, а минимальные совпадают с локализацией нижнего экстремума индекса Шеннона. Индекс доминирования имеет обратный индексу биоразнообразия характер и также сильнее варьирует у дождевых червей ельников по сравнению с таксоценами других формаций.
Как было показано выше, таксоцены ельников выделяются не только изменчивостью синэкологических показателей (индексов Шеннона, Симпсона и Пиелу, общих численности, биомассы, видового богатства дождевых червей), но и наибольшим числом специализированных видов (табл. 3). Как и бук восточный, пихта Нордмана и ель восточная являются реликтами третичного периода. По экологическому статусу (мезофильности и мезотрофности, несколько большей теплолюбивости), бук и пихта ближе друг к другу, что проявляется в широком распространении смешанных древостоев (Бебия, 2002). Леса с господством ели развивается на сухих каменистых склонах, или на почвах с более легким механическим составом. Преобладание ели в древесном ярусе обусловливает формирование оподзоленного горизонта с комковатой структурой и низкой влажностью. Т.к. ель достаточно редко образует чистые формации, значительная примесь других пород приводит к изменению почвенных свойств и гетерогенности трофических ресурсов смешанного леса, обусловливая в свою очередь увеличение разнообразия и неоднородности сообществ дождевых червей.
Кластерный анализ. Изучение соотношения морфо-экологических и хорологических групп дождевых червей с помощью кластерного анализа позволило выделить два основных кластера (рис. 6), объединяющихся за пределами допустимого сходства.
Рис. 6.
Сходство хорологической и экологической структуры населения дождевых червей в лесных и луговых формациях среднего течения р. Большая Лаба (метод Уорда, Евклидовы дистанции, достоверность на уровне 300). Обозначения фитоценозов с 1 по 23 – как и в табл. 1.
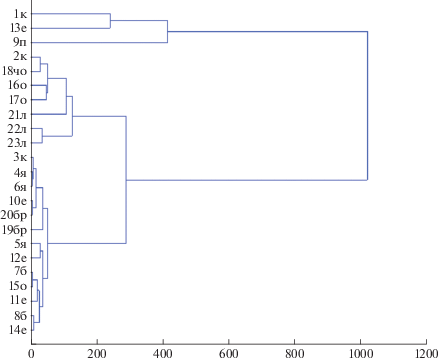
Наиболее многочисленный нижний кластер объединяет два схожих между собой субкластера, нижний из которых делится на два блока. К первому блоку относятся таксоцены дождевых червей ясенников, одного кленовника, одного ельника и березняков. В ясениках и кленовнике встречаются только специализированные собственно-почвенные и подстилочные виды, относящиеся к роду Dendrobaena (табл. 3). Это восточноевро-азиатский вид D. tellermanica, известный способностью к диапаузе, космополитный D. octaedra и средиземноморский D. attemsi. В монодоминантном сообществе ельника отмечен только один всесветно распространенный подстилочный вид дождевых червей – Dendrodrilus rubidus tenuis. Этот вид часто встречается в зеленомошных синузиях, под корой валежника на люмбрицидной стадии разложения (Перель, 1979; Мамаев, 1960), что позволяет не только переживать временную засуху, но и обильные сезонные водотоки. В березовых биогеоценозах также многочисленны собственно почвенные виды, близко подступающие грунтовые воды обусловливают высокую плотность O. lacteum, обильны здесь и подстилочные люмбрициды (рис. 4). Структура населения сложнее – встречаются почвенно-подстилочный L. rubellus и полиморфный D. schmidti. Таким образом, маркером первого блока можно считать высокую численность подстилочных видов – бореального космополита D. octaedra, неморального средиземноморского вида D. attemsi, замещаемых в ельнике экологически викарирующим бореальным космополитом Dendrodrilus rubidus tenuis, и частую встречаемость собственно почвенного восточно-евроазиатского неморально-степного D. tellermanica, дополняемого в березняках другим неморально-степным собственно почвенным видом O. lacteum с более широким – всесветным распространением. Растительные сообщества, в пределах которых выделены перечисленные биогеоценозы, характеризуются меньшей по сравнению с другими формациями сомкнутостью древесного яруса, более низкой влажностью и высокой текущей температурой почв (табл. 1, 2). Ясенники, кленовник и ельник произрастают на трансэлювиальных ландшафтных местоположениях, уклон которых превышает 30% (табл. 2). Мы считаем, что расход влаги на этих позициях рельефа происходит преимущественно на поверхностный сток и испарение. Березняки сформированы на низких надпойменных террасах на маломощных аллювиальных почвах со значительным включением песчаной фракции, слой А1 которых в летний период вероятно также характеризуется переменным гидротермическим режимом из-за низкой сомкнутости древесного яруса. Однако близкое залегание грунтовых вод обусловливает мозаичность микрорельефа и большее разнообразие экологической и хорологической структуры населения люмбрицид.
Ко второму блоку первого субкластера относится население люмбрицид букняков, основной части ельников, одного ясенника и одного сероольшаника. В хорологическом отношении эта группа сообществ дождевых червей характеризуется обилием средиземноморских видов (рис. 3). Исключение – население дождевых червей букняка елового редкопокровного, где зарегистрированы восточноевро-азиатский D. tellermanica и космополитный O. lacteum. В морфо-экологической структуре населения этого блока преобладают либо собственно почвенные, либо подстилочные виды (рис. 4), причем в таксономическом отношении обе группы представлены более разнообразно (табл. 3). Исключение – тот же таксоцен леса с доминированием бука, объединяемый с другими сообществами высокой численностью собственно почвенных видов. Именно в сообществах этого блока зарегистрированы большинство редких видов люмбрицид. Перечисленные биогеоценозы сформированы преимущественно на автоморфных почвах с мощным профилем, занимающих трансакумулятивные позиции либо проточные водосборные понижения и эрозионные лощины на пологих склонах. Бук и клен – мезофиты и эвтрофы, плохо переносящие избыточное увлажнение и заболачивание (Погребняк, 1955; Справочник лесничего, 2003). Сероольшаник произрастает на склоне, уклон которого составляет 10°.
Ко второму субкластеру относятся таксоцены большинства ольшаников, лугов и одного ясенево-кленового разнотравного участка леса, черви в котором собраны в эрозионной лощине на месте временного водотока, о чем свидетельствует высокая численность амфибионта Eiseniella tetraedra tetraedra и влаголюбивого E. fetida. Оба вида часто встречаются и в других биогеоценозах, относящихся к этому кластеру. В хорологическом отношении население дождевых червей характеризуется наибольшим обилием космополитных видов. Здесь зарегистрированы разные морфо-экологические группы, в том числе почвенно-подстилочные виды, редкие в других таксоценах. Все биогеоценозы сформированы на аккумулятивных и трансакумулятивных ландшафтных положениях. Занимаемые ими почвы преимущественно гидроморфного ряда, за исключением тех, в пределах которых произрастает ясенево-кленовый разнотравный участок леса, но и он вероятно подвержен сезонному проточному заболачиванию. Вцелом первый субкластер характеризуется преобладанием в структуре населения специализированных мезофильных видов, второй – влаголюбивых широко распространенных форм.
Три таксоцена – население кленовника ясеневого разнотравного, ельника букового мертвопокровного и пихтарника букового с чередованием мертвопокровных и мохово-лишайниковых участков выделилось в отдельный кластер, объединяемый за пределами достоверного сходства (рис. 6). Ясенево-кленовый таксоцен отличается особенно высоким обилием космополитных видов дождевых червей, буково-еловый мертвопокровный – монодоминантным составом населения, в котором представлен исключительно восточноевро-азиатский D. tellermanica (табл. 3). В морфо-экологической структуре этих сообществ более заметно преобладают собственно почвенные виды (рис. 4). По эдафическим и ландшафтным условиям оба последних биогеоценоза мало отличаются от основной массы ельников и кленовников, объединенных в первом субкластере. Наиболее оригинален пихтовый таксоцен, характеризующийся содоминированием космополитного и крымско-кавказского видов и значительным отличием морфо-экологической структуры – только здесь отмечены собственно почвенный, почвенно-подстилочный и полиморфный виды.
Наблюдаемая картина, на наш взгляд, может определяться выпадением большого количества осадков – более 1000 мм в год (Бебия, 2002). Значительная часть осадков приходится на весенне-раннелетний период, второй максимум отмечается в конце летнего сезона. Прерываются эти максимумы относительно сухим среднелетним периодом (Зонн, 1950). Из-за эрозионного характера рельефа на трансэлювиальных позициях ландшафта формирование таксоценов дождевых червей происходит под воздействием периодических обильных водотоков и некоторого недостатка влаги в среднелетний период. Таким образом сезонное избыточное увлажнение является одним из основных факторов, определяющих структуру населения беспозвоночных. Несмотря на в целом комфортные текущие значения температуры и влажности в ясенниках, ельниках и березовых биогеоценозах в период исследований, мы считаем, что долговременная экспозиция более высоких текущих температур (табл. 2) и различия в механическом составе и мощности почв приводят к более существенным различиям в среднесезонных температуре и влажности их органогенного слоя по сравнению, например, с ольховыми биогеоценозами.
ЗАКЛЮЧЕНИЕ
В исследованных сообществах отмечены 14 видов дождевых червей. Неожиданностью стало нахождение вне пойменных почв амфибионта Eiseniella tetraedra tetraedra, вероятно, попавшего в эрозионные ложбины с временными водотоками. Наряду с влаголюбивыми L. rubellus и E. fetida, зарегистрированы виды, чаще встречающиеся в степных и луговых ландшафтах, однако в бассейне среднего течения р. Большая Лаба, за исключением D. tellermanica, эти люмбрициды не многочисленны.
В хорологическом отношении фауна дождевых червей бассейна р. Большая Лаба отличается от других районов Северного Кавказа. Нетипично редко встречаются собственно кавказские виды. Крымско-кавказский субэндемик D. schmidti, доминирующий на северном макросклоне Большого Кавказа, в большинстве сообществ замещается восточноевро-азиатским D. tellermanica. D. tellermanica – неморально-степной партеногенетический вид, хорошо приспособленный к неблагоприятным гидротермическим условиям. В степи D. tellermanica встречается в тех же биогеоценозах, что и O. lacteum и A. rosea – люмбрициды, хорошо переносящие как переувлажнение, так и пересыхание почв.
У ряда таксонов выявлена высокая степень относительной биотопической приуроченности. Биотопическая преференция отмечена у крымско-кавказского эндемика D. schmidti, восточноевро-азиатского D. tellermanica и средиземноморского D. attemsi. Наиболее эвритопными, за небольшим исключением, оказались виды с космополитным типом ареала.
Экологическая структура населения дождевых червей так же имеет свои особенности. В среднем течении р. Большая Лаба не зарегистрированы норники, не переносящие заболачивания почв. Реже, чем в других районах, встречаются почвенно-подстилочные виды. Зависимость структуры населения дождевых червей от водного режима почв хорошо иллюстрируется соотношением численности морфо-экологических форм.
Распределение дождевых червей – в целом очень влаголюбивой группы – определяется множеством факторов, ведущим из которых, безусловно, является гидроэдафический. Влияние ландшафта проявляется опосредованно – через автоморфный или гидроморфный характер почв и локальные особенности биогеоценозов.
Наибольшей изменчивостью синэкологических показателей населения дождевых червей и числом специализированных видов выделяются таксоцены ельников. Подобный факт ранее отмечался в буковых лесах Центрального Кавказа (Рапопорт и др., 2017). Вероятно, именно в зональных формациях у различных видов реализуется наибольшее число экологических ниш, что подчеркивает биогеоценотическую значимость этих реликтовых фитоценозов как важных рефугиумов флоры и фауны Кавказа.
БЛАГОДАРНОСТИ
Искренне благодарим Ф.А. Темботову и ${\text{Т }}{\text{.С }}{\text{.}}\,{\text{В с е в о л о д о в у - П е р е л ь }}$ за всестороннюю поддержку и интерес к нашей работе; М.Х. Кармокова, В.И. Ланцова и Н.Б. Винокурова – за помощь в отборе почвенных проб; сотрудников лаборатории почвенно-экологических исследований за выполнение физико-химического анализа почв.
Работа выполнена при частичной финансовой поддержке программы фундаментальных исследований Президиума РАН “Живая природа: современное состояние и проблемы развития”, 2012–2014 гг. и РФФИ (18-04-00961).
Список литературы
Аринушкина Е.В., 1970. Руководство по химическому анализу почв. М.: Изд-во МГУ. 487 с.
Бебия С.М., 2002. Пихтовые леса Кавказа. М.: МГУЛ. 270 с.
Вальтер Г., 1974. Лиственные леса умеренной зоны Северного полушария – неморальная зона // Растительность земного шара. Эколого-филогенетическая характеристика. М.: Прогресс. 422 с.
Всеволодова-Перель Т.С., 1997. Дождевые черви фауны России. М.: Наука. 102 с.
Гиляров М.С., 1975. Учет крупных беспозвоночных (мезофауны) // Методы почвенно-зоологических исследований. М.: Наука. С. 12–29.
Зонн С.В., 1950. Горно-лесные почвы Северо-Западного Кавказа. М.-Л.: Изд-во Академии наук СССР. 233 с.
Исаченко И.Г., 1991. Ландшафтоведение и физико-географическое районирование. М.: Высшая школа. 366 с.
Казеев К.Ш., Колесников С.И., Вальков В.Ф., 2004. Биология почв Юга России. Ростов-на Дону: Изд-во ЦВВР. 350 с.
Квавадзе Э.Ш., 1985. Дождевые черви (Lumbricidae) Кавказа. Тбилиси: Мецниереба. 283 с.
Малевич И.И., 1976. Распространение сем. Lumbricidae в европейской части СССР (по картографическим материалам) // Фауна и экология беспозвоночных животных. Ч. 1. М.: Наука. С. 3–11.
Мамаев Б.М., 1960. Зоологическая оценка стадий естественного разрушения древесины. Изв. АНСССР. Сер. Биологическая. № 4. С. 610–617.
Мещерякова Е.Н., Берман Д.И., 2014. Устойчивость к отрицательным температурам и географическое распространение дождевых червей (Oligochaeta, Lumbricidae, Moniligastridae) // Зоологический журнал. Т. 93. № 1. С. 53–64.
Нечаев Ю.А., 1960. Лесные богатства Кабардино-Балкарии. Нальчик: Эльбрус. 143 с.
Овсянников Г.Ф., 1931. Лиственные породы. Пособие для учащихся и лесных специалистов. Владивосток: ОГИЗ – Далькрай. С. 69–71.
Одум Ю., 1975. Основы экологии. М.: Мир. С. 186.
Олейникова Д.В., Шальнов В.А., Гончарова Н.Н., 2008. Особенности среднегорных и высокогорных ландшафтов верховий Большой Лабы // Вопросы современной науки и практики. Университет им. Вернадского. №3(13). Т. 2. С. 127–135.
Перель Т.С., 1979. Распространение и закономерности распределения дождевых червей фауны СССР. М.: Наука. 275 с.
Песенко Ю.А., 1982. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука. 288 с.
Погребняк П.С., 1955. Основы лесной типологии. Киев: Изд-во Академии наук Украинской ССР. 456 с.
Проконова Т.В., 2005. Видовой состав и классификация группировок дождевых червей (Lumbricidae, Oligochaeta) Центрального Предкавказья // Известия ВУЗов. Северо-Кавказский регион. Естественные науки. № 3. С. 70–74.
Раменский Л.Г., 1938. Введение в комплексное почвенно-геоботаническое исследование земель. М.: Сельхозгиз. 620 с.
Рапопорт И.Б., 2011. Сезонная активность дождевых червей (Oligochaeta, Lumbricidae) субальпийского пояса Центрального Кавказа // Известия Самарского НЦ РАН. Т. 13. № 5. С. 148–151.
Рапопорт И.Б., 2013. Высотное распределение дождевых червей (Oligochaeta, Lumbricidae) в центральной части Северного Кавказа // Зоологический журнал. № 1. С. 3–10.
Рапопорт И.Б., 2014. Фауна, структура сообществ и высотно-поясное распределение дождевых червей (Oligochaeta, Lumbricidae) центральной части кубанского варианта поясности (Северо-Западный Кавказ, Республика Адыгея) // Вестник АГУ. № 4. С. 78–85.
Рапопорт И.Б., Зенкова И.В., Цепкова Н.Л., 2017. Население дождевых червей (Oligochaeta, Lumbricidae) лесных и луговых сообществ бассейна реки Карасу (Центральный Кавказ) // Зоологический журнал. Т. 96. № 2. С. 172–183. [Rapoport I.B., Zenkova I.V., Tsepkova N.L., 2017. Earthworm (Oligochaeta, Lumbricidae) Populationsof the Karasu River Basin (Central Caucasus) // Biology Bulletin. V. 44. № 8. Р. 941–951].
Рапопорт И.Б., Комаров Ю.Е., 2017. Дождевые черви (Oligochaeta: Lumbricidae) Северной Осетии-Алании (Центральный Кавказ) // Известия Самарского научного центра. № 5(1). С.86–93.
Соколов В.Е., Темботов А.К. 1989. Млекопитающие. Насекомоядные. М.: Наука. С. 3–27.
Справочник лесничего, 2003. М.: ВНИИЛМ. 640 с.
Стриганова Б.Р., 1980. Питание почвенных сапрофагов. М.: Наука. 244 с.
Сукачёв В.Н., 1942. Идея развития в фитоценологии // Сов. Ботаника. № 1–3. С. 5–17.
Халафян А.А., 2007. Statistica 6. Статистический анализ данных. М.: Бином-Пресс. 512 с.
Шильников Д.С., 2009. Флора бассейна реки Большая Лаба и её анализ. Автореф. … канд. биол. наук. Санкт-Петербург. 21 с.
Шхагапсоев С.Х., Волкович В.Б., 2002. Растительный покров Кабардино-Балкарии и его охрана. Нальчик: Эльбрус. 95 с.
Csuzdi Cs., Zicsi A., 2003. Earthworms of Hungary (Annelida: Oligochaeta; Lumbricidae). Budapest: Hungarian Natural History Museum. 271 p.
Csuzdi Cs., Zicsi A., Misirlioğlu M., 2006. An annotated checklist of the earthworm fauna of Turkey (Oligochaeta: Lumbricidae) // Zootaxa. 1175. P. 1–29.
Liebeke M., Strittmatter N., Fearn S., Morgan A.J., Kille P. et al., 2015. Unique metabolites protect earthworms against plant polyphenols // Nat. Commun. № 6. P. 1–6.
Michaelsen W., 1903. Die geographische Verbreitung der Oligochäten. Berlin: R. Friedländer und Sohn, 186 p.
Omodeo P., 1952. Particolaritádella Zoogeografia dei Lombrichi // Bollettino di Zoologia. № 19. P. 349–359.
Stojanović M., Karaman S., 2005. Further contribution to knowledge of the earthworm of Šumadija (Serbia) // Archives of Biological Sciences. Belgrade. V. 57. № 2. P.127–132.
Szederjesi T., 2017. The first combined checklist of earthworms of the Northeastern Mediterranean region (Clitellata: Megadrili) // Opuscula Zoologica. Budapest. V. 48. № 2. P. 77–116.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал