

Зоологический журнал, 2019, T. 98, № 5, стр. 567-577
Оценка численности дальневосточного леопарда (Panthera pardus) в России
В. С. Лукаревский 1, *, С. В. Лукаревский 1
1 Информационно-аналитический центр поддержки заповедного дела,
117042 Москва, Россия
* E-mail: vlukarevsky@mail.ru
Поступила в редакцию 18.04.2018
После доработки 08.08.2018
Принята к публикации 09.12.2018
Аннотация
Дальневосточный леопард (Panthera pardus orientalis) – одна из наиболее редких форм крупных кошек мира. Его численность оценивалась в 25–50 особей. Новые методы исследования с использованием автоматических фотокамер и молекулярно-генетической диагностики также не дали исчерпывающей информации. Работу проводили в 2011–2013 гг. на юго-западе Приморского края: от р. Раздольная на северо-востоке, до границы с КНДР на юге и КНР на юго-западе. Нами выбран модельный участок площадью 503 км2, где была установлена сеть из 41 пары фотоловушек Reconyx Rapidfire RC60, HC500, HC600 – высокоскоростных автоматических цифровых камер с инфракрасным датчиком движения и инфракрасным освещением, позволяющих делать 1 кадр в секунду. Снятие информации с фотоловушек проводили один раз в четыре месяца. Для оценки численности леопарда были использованы два метода: прямая экстраполяция числа зарегистрированных леопардов в течение года на модельном участке на весь ареал подвида и модифицированный метод программы Capture с определением плотности популяции, основанный на принципе повторного отлова и учитывающий пространственные связи между регистрациями животных (Space Capture-Recapture, SECR) при использовании программы SPACECAP. Для корректной экстраполяции была определена “эффективная область отлова”, рассчитанная как вариант половины среднего значения расстояния между двумя повторными отловами (1/2MMDM), что позволило получить буферную зону шириной в 5.66 км. Эффективная область отлова составила 1023 км2. Из этой площади вычтены заведомо непригодные местообитания. На 377 фотоснимках, полученных с фотоловушек, нами идентифицировано 27 леопардов, из них 10 самцов, 10 самок, для 7 особей пол не установлен, на 10 фоторегистрациях индивидуальная идентификация была невозможной. Экстраполяция этих данных на всю территорию потенциальных местообитаний леопарда на российском Дальнем Востоке, т.е. на площадь 3502 км2, позволяет заключить, что общая численность леопарда составляет около 96–100 особей, из которых около 60 животных являются резидентными, а 35–40 – молодые, кочующие особи. Однако, учитывая предыдущие учеты численности леопардов, следы которых были обнаружены на площади 2233 км2, общая оценка численности, по-видимому, будет ниже и составит около 60–65 леопардов.
Дальневосточный леопард (P. p. orientali ) – одна из наиболее редких форм крупных кошек мира. Проблема оценки его численности активно обсуждается более 25 лет. Во многих работах приводятся относительно близкие результаты. Так, численность леопарда на Юго-Западе Приморского края оценивалась в 25–50 особей (Абрамов, Пикунов, 1974; Пикунов, Коркишко, 1992; Арамилев, Фоменко, 200011; Пикунов и др., 2009 и др.). Приводимые оценки основаны на конкретных данных, полученных в ходе полевых исследований, но и несмотря на это носят исключительно экспертный и субъективный характер. Это связано с тем, что в этих исследованиях единицей измерения является след, оставленный леопардом на снегу, а решение о его принадлежности конкретной особи каждый исследователь принимал индивидуально. С появлением новых методов исследования – использования автоматических фотокамер (Karanth, 1995; Karanth et al., 2002) и молекулярно-генетической диагностики (Sugimoto et al., 2012; Рожнов и др., 2013) – вопросы индивидуальной идентификации животных были решены. Однако проблемы оценки численности остались.
Мониторинг состояния популяции дальневосточного леопарда, а также оценка численности крайне необходимы. Любые новые данные, такие как структура ареала, структура популяции (генетическая, пространственно-этологическая и др.), вносят неоценимый вклад в его сохранение. Кроме того, оценка состояния популяции редкой формы дальневосточного леопарда, как и любой другой, позволяет понять механизмы существования небольших популяций на протяжении длительного времени. Это дает возможность своевременно разрабатывать и принимать соответствующие решения по его сохранению.
Цель настоящей работы – оценка численности дальневосточного леопарда на основе использования фотоловушек, выявление факторов, определяющих его размещение и плотность популяции. Кроме того, используемые нами методы протестированы на возможность использования их для мониторинга популяции дальневосточного леопарда.
МАТЕРИАЛ И МЕТОДИКА
Работу проводили в 2011–2013 гг. на юго-западе Приморского края: от р. Раздольная на северо-востоке до границы с КНДР на юге и КНР на юго-западе.
Площадь потенциальных местообитаний леопарда в Юго-Западном Приморье по различным оценкам составляет от 3502 (Hebblewhite et al., 2011) до 5217 км2 (Пикунов и др., 2009). Анализ результатов, полученных нами во время маршрутного обследования территории (рис. 1), показал, что распределение леопарда неравномерно, а значительная часть территории вследствие антропогенных или природных условий мало пригодна для обитания леопарда.
Рис. 1.
Схема маршрутов обследования территории и точки встреч следов жизнедеятельности леопарда в 2011–2012 гг.
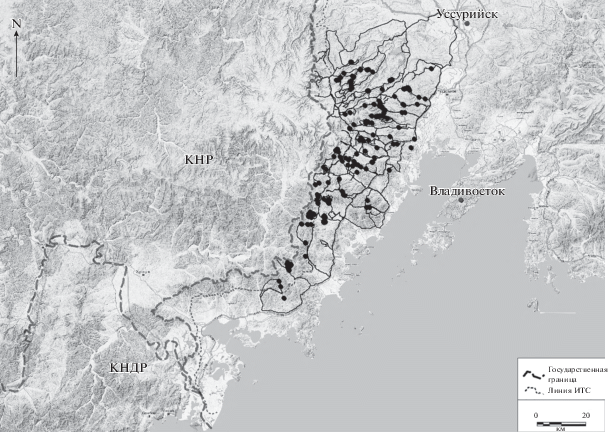
Практически на всей российской части ареала дальневосточного леопарда нами проведены одноразовые маршруты, на которых регистрировали все следы жизнедеятельности леопарда – следы, поскребы, мочевые точки, шерсть, экскременты.
Для изучения выбора леопардом местообитаний, определения структуры и плотности его популяции, факторов, влияющих на распределение, нами был выбран модельный участок площадью 503 км2 или около 19% площади Национального парка “Земля леопарда”, который охватывает бассейны рек Амба, Барабашевка и Нарва (рис. 2).
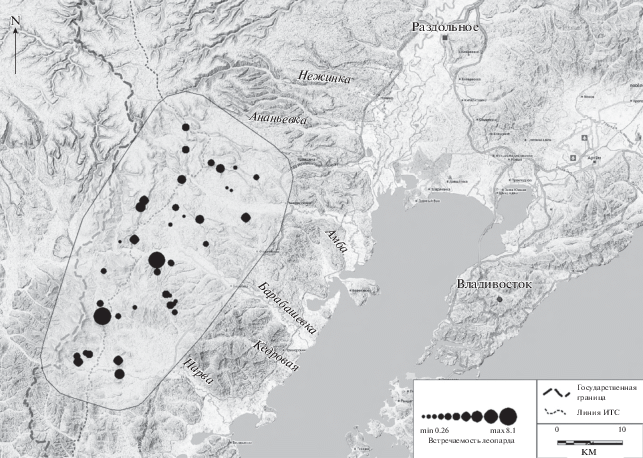
Местообитания леопарда здесь включают различные биотопы: склоны гор с сильно расчлененными ущельями, оврагами, обнажениями материнских пород с растительностью различных формаций – от дубово-широколиственных лесов до пирогенно трансформированных комплексов с господством леспедеции (Lespedeza sp.). Большая часть полей сельскохозяйственного назначения фактически не освоена и заросла высоким разнотравьем, практически ежегодно они полностью выгорают. Значительная часть территории (пойма реки Нарва) занята военным полигоном, где раньше регулярно проводились военные учения. Около трети территорий расположены за линией инженерно-технических сооружений (ИТС), куда местных жителей пропускают только по особым разрешениям. На востоке к модельному участку примыкает заповедник “Кедровая падь”.
На модельном участке нами была установлена сеть из 41 пары фотоловушек Reconyx Rapidfire RC60, HC500, HC600 – высокоскоростных автоматических цифровых камер с инфракрасным датчиком движения и инфракрасным освещением, позволяющих делать 1 кадр в секунду. Снятие информации с фотоловушек проводили один раз в четыре месяца, что позволило исключить фактор “отпугивания” животных. Большинство леопардов привыкли к присутствию фотоловушек и не обращали на них внимания. Самки вели себя аналогично, лишь некоторые – самые осторожные – подходили к фотоловушкам и детально их осматривали. Кроме того, редкое посещение человеком фотоловушек должно было исключить также влияние запаха на присутствие животных.
Фотоловушки были установлены таким образом, чтобы “отснять” всех леопардов, живущих на этой территории. При их установке были учтены особенности поведения леопарда: как и другие крупные кошки, он использует для перемещения особенности рельефа, направляющие его движение (Матюшкин, 1977; Schaller, Crawshaw, 1980; Лукаревский, 2001; Лукаревский, Поярков, 2008).
Участок обитания дальневосточного леопарда имеет площадь от 300–600 км2 у самцов до 250–300 км2 у самок (Коркишко, 1986; Огастин и др., 1996; Чистополова и др., 2012; Рожнов и др., 2015). Фотоловушки устанавливали таким образом, чтобы одна пара была расположена в квадрате 5 × 5 км, поэтому на участке обитания было установлено не менее двух пар фотоловушек. Это позволяет увеличить вероятность съемки всех особей. Фотоловушки располагали друг против друга, чтобы сфотографировать оба бока животного одновременно, так как узор на шкуре леопарда индивидуален и асимметричен. На основе полученных снимков составляли истории съемки каждой особи.
Для оценки численности леопарда были использованы два метода: прямая экстраполяция числа зарегистрированных леопардов в течение года на модельном участке на весь ареал подвида в России и компьютерная модель программы Capture. Метод Capture, основанный на принципе “повторного отлова”, ранее был использован для оценки численности тигра (Panthera tigris) (Karanth, Nichols, 1998; Костыря и др., 2003) и дальневосточного леопарда (Арамилев и др., 2010, 2012).
Нами использован модифицированный метод программы Capture с определением плотности популяции. Метод также основан на принципе повторного отлова, учитывающего пространственные связи между регистрациями животных (Space Capture-Recapture, SECR) при использовании программы SPACECAP (Royle et al., 2009). Этот метод апробирован нами на амурском тигре (Эрнандес-Бланко и др., 2013).
Для верификации (анализа) влияния различных факторов на “уловистость” леопарда в фотоловушках и учитывая, что часть животных, в силу их биологических особенностей или природных условий (в частности, реакции на фотоловушки или наличия котят у самок), избегают фотоловушек, мы провели четыре анализа с учетом возможных сезонных различий: зима – влияние снежного покрова на перемещения леопардов; весна – отсутствие листвы на деревьях и уплотненная листва на земле, что делает перемещения леопарда бесшумными, а возможность хорошего обзора с гребней хребтов делают их привлекательными для охоты и перемещения; лето – развитая листва и ограниченный обзор; осень – отсутствие листвы на деревьях, хороший обзор и опавшая хрустящая листва на земле.
Продолжительность периодов съемки, число локаций и число отснятых леопардов для анализа программой SPACECAP приведены в табл. 1.
Таблица 1.
Регистрации леопарда программой SPACECAP
| Сезон фотоотлова | Период анализа | Число “отловленных” леопардов | Число регистраций |
|---|---|---|---|
| Весна | 13.03.2012–11.05.2012 | 17 | 80 |
| Лето | 19.06.2012–17.08.2012 | 16 | 53 |
| Осень | 30.09.2012–28.11.2012 | 14 | 58 |
| Зима | 25.12.2012–22.02.2013 | 9 | 25 |
Число суток, отработанных каждой фотоловушкой, регистрировали как число дней, прошедших с момента установки фотоловушки/замены батарей до последнего кадра, снятого этой фотоловушкой. При замене батарей к фотоловушкам подходили со стороны камеры, проверяя, в рабочем состоянии она находится или нет.
Годовой цикл работы фотоловушек позволил получить информацию о размещении леопардов и характере их пребывания на модельном участке в течение года. Это позволило нам сравнить и проанализировать полученные при использовании разных подходов результаты.
Поскольку участки обитания части отловленных нами леопардов могут выходить далеко за пределы расположения фотоловушек, для корректной экстраполяции полученных нами результатов съемки (“отлова”) необходимо определить “эффективную область отлова”. В дальнейшем, так как речь идет о фотоловушках, мы позволили себе для удобства заменить слова “съемка” и “фотосъемка” на “отлов”. Для этого используется среднее значение расстояния между двумя повторными отловами (MMDM) или его половины (½MMDM) (Wilson, Anderson, 1985). Нами был использован вариант половины среднего значения расстояния между двумя повторными отловами (½MMDM), что позволило получить буферную зону шириной в 5.66 км. Вариант среднего значения расстояния между двумя повторными отловами (MMDM) неприемлем, ввиду того, что включает участки обитания зверей, которые нашими фотоловушками ни разу не регистрировались, зато регулярно отмечены на прилежащих территориях (Арамилев и др., 2012).
В эффективную область попали территории, заведомо непригодные для обитания леопарда (населенные пункты, сельскохозяйственные поля, стрельбище), а также расположенная вне российской части ареала территория КНР. Поэтому указанные территории были вычтены. Таким образом, общая эффективная область отлова составила 1023 км2.
Зима 2011–2012 гг. была малоснежной: максимальная глубина снежного покрова достигала 15–25 см в северной части ареала на высотах свыше 700 м над ур. м., тогда как в южной части ареала снег глубиной 5–10 см лежал лишь на склонах северной экспозиции. Зима 2012–2013 гг., напротив, была многоснежной: уже в конце ноября установился глубокий снежный покров по всей территории ареала дальневосточного леопарда и сохранился до середины марта. Глубина снежного покрова составляла свыше 40 см, а в верхней части хребтов достигала 1 м и более.
Для выявления влияния погодных условий, в частности снегопадов, на распределение леопарда и сезонных его изменений нами был рассчитан коэффициент встречаемости леопарда для каждого месяца (число локаций леопарда за месяц в пересчете на 100 фотоловушко/суток).
Один из основных факторов, влияющих на распределение леопарда, – состояние популяций основных видов его жертв. Нами была проведена оценка обилия основных объектов питания леопарда – пятнистого оленя (Cervus nippon), сибирской косули (Capreolus pygargus) и кабана (Sus scrofa). Мы исходили из того, что все ловушки были расположены единообразно на всей территории модельного участка, в одинаковых биотопах, а именно на гребнях хребтов, поэтому условия и вероятность регистрации объектов питания леопарда на разных хребтах на исследуемом участке были однообразны в течение всего года. Индекс обилия потенциальных жертв леопарда, так же как в исследованиях по амурскому тигру (Рожнов и др., 2012), характеризовали числом зарегистрированных особей на число фотоловушко/суток. Всего отработано 14 895 фотоловушко/суток, получена и обработана 4671 фоторегистрация потенциальных объектов питания леопарда.
Анализировали число фоторегистраций потенциальных жертв леопарда, к которым относили пятнистого оленя, косулю, кабана, лисицу (Vulpes vulpes), азиатского барсука (Meles leucurus) и зайцев: беляка (Lepus timidus) и маньчжурского (L. mandshuricus). Для каждой фоторегистрации фиксировали дату и время, определяли до вида потенциальные жертвы леопарда. Животных не идентифицировали индивидуально. Несколько фотографий, сделанных фотоловушкой последовательно с интервалом до 3 мин, считали как проход (фоторегистрация) одного животного. Для каждого вида рассчитывали индекс обилия – число фотолокаций животных этого вида на 100 фотоловушко/суток. При расчете обилия жертв леопарда не принимали во внимание фоторегистрации кабарги (Moschus moschiferus) и енотовидной собаки (Nyctereutes procyonoides) из-за небольшого их числа (n = 12 и n = 22, соответственно), а также фоторегистрации белки (Sciurus vulgaris), бурундука (Tamias sibiricus), колонка (Mustela sibirica), харзы (Martes flavigula) и птиц.
РЕЗУЛЬТАТЫ
Численность леопарда на исследуемой территории. На 377 фотоснимках, полученных с фотоловушек, нами идентифицировано 27 леопардов, из них 10 самцов (286 фоторегистраций), 10 самок (68 фоторегистраций), для 7 особей (12 фоторегистраций) пол не установлен, на 10 фоторегистрациях из-за плохого качества изображений индивидуальная идентификация была невозможной.
Анализ результатов регистрации леопардов на фотоловушках в течение года позволил выявить на модельном участке резидентных (регистрируемых на фотоловушках регулярно) и кочующих (регистрируемых лишь на одной или двух фотоловушках в центральной части модельного участка в течение короткого времени) особей. Из 27 леопардов 19 оказались резидентными, а восемь – кочующими. Учитывая, что самки регистрируются на фотоловушках значительно реже самцов (примерно 1 : 4), и тот факт, что молодые самки, в отличие от самцов, не покидают материнские группировки (Коркишко, 1986а), мы отнесли их всех к категории резидентных особей. Нерезидентные леопарды, по-видимому, перемещаются в поисках свободных участков – это либо молодые, либо старые, потерявшие участки обитания самцы. Например, самец Хасан, помеченный нами спутниковым ошейником, возраст которого был оценен в 12–14 лет, после того как на его участке поселился другой самец, покинул территорию и был отмечен нами на самой дальней ловушке, установленной в бассейне р. Амба, после чего его регистрации на фотоловушках прекратились.
Прямая экстраполяция числа зарегистрированных леопардов в течение года на модельном участке на весь ареал подвида (3600 км2) позволяет заключить, что на Российской части ареала общая численность составляет около 95 особей.
Результаты, полученные с использованием программы SPACECAP для одной и той же схемы размещения фотоловушек, но для различных сезонов, весьма различны (табл. 2), что ставит под сомнение возможность ее использования для мониторинга популяции дальневосточного леопарда. Среднее значение численности леопардов, вычисленное с помощью программы SPACECAP, составляет от 36 особей летом до 25 особей зимой. Эффективная площадь отлова, определенная программой SPACECAP, составляет 2615 км2, однако значительная часть этого пространства расположена на территории КНР.
Таблица 2.
Результаты анализа фотоотловов леопарда программой SPACECAP
| Сезон | Число “отловленных” леопардов | Число регистраций | Численность | Плотность средняя | ||
|---|---|---|---|---|---|---|
| средняя | min | max | ||||
| Весна | 17 | 80 | 30.2398 | 19 | 41 | 1.3583 |
| Лето | 16 | 53 | 36.1184 | 22 | 51 | 1.6224 |
| Осень | 14 | 58 | 23.8235 | 15 | 33 | 1.0701 |
| Зима | 9 | 25 | 24.5765 | 10 | 40 | 1.1039 |
Распределение леопардов на исследуемой территории. Обследование территории в декабре 2011 г.–марте 2012 г. показало, что животные встречаются на ней неравномерно. На значительной части территории следы жизнедеятельности леопарда не отмечены (рис. 1). Леопард практически полностью отсутствует на участках пограничной заставы “Таежная” и к северу от нее. Не отмечены следы жизнедеятельности леопарда также в верховьях рек Борисовка, Крауновка и Павлиновка – на площади около 247 км2. Следы жизнедеятельности леопарда практически отсутствуют (за редким исключением) к югу от бассейна р. Рязановка на территории до ИТС.
В пределах установленной матрицы фотоловушек “уловистость”, отражающая распределение леопарда, неравномерна (рис. 2).
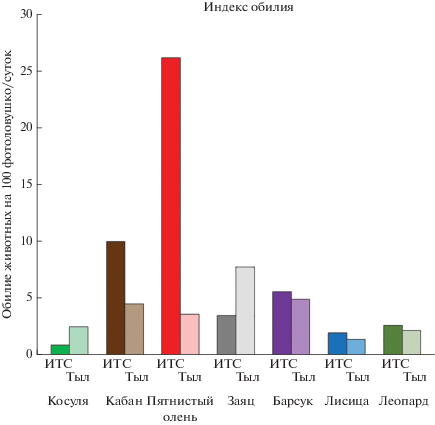
Наиболее плотно леопардом заселены участки, расположенные в непосредственной близости к ИТС и до линии государственной границы: здесь зарегистрировано 18 леопардов. На этих же участках наиболее высокая плотность копытных (рис. 3). Суммарная плотность копытных за линией инженерно-технических сооружений (индекс обилия 37.3) значительно превышает таковую на остальной части модельного участка (в “тылу”), где индекс обилия составляет 10.6 особей на 100 ловушко/суток.
Рис. 3.
Обилие леопарда и его потенциальных объектов питания в зоне инженерно-технических сооружений (ИТС) и на остальной части модельного участка (в “тылу”).
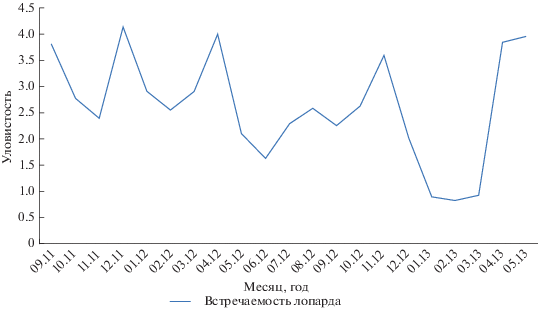
Факторы, влияющие на распределение леопарда. Уловистость леопарда значительно различается в разные сезоны года. Наибольшая “уловистость” леопарда отмечена в осенний (3.7 леопарда на 100 фотоловушко/суток) и весенний (4.0 леопарда на 100 фотоловушко/суток) периоды (рис. 4). Максимальные значения “уловистости” леопарда (4.14 локаций леопарда на 100 фотоловушко/суток), отмеченные в декабре 2011 г., совпали с практически полным отсутствием снега на модельном участке в этот период. Это подтверждает представления о том, что наибольшее значение в распределении леопарда имеет снежный покров.
Снег в значительной степени определяет распределение леопарда и его активность. На модельном участке были пройдены 27 маршрутов, которыми его последовательно обследовали в течение 7–8 дней в декабре 2012 г. и феврале 2013 г. Здесь следы леопарда, которые принадлежали только взрослым самцам, были отмечены только на шести маршрутах из 27. На гребнях отрогов, где в обычных условиях наиболее часто встречались следы жизнедеятельности леопардов, следы были отмечены только на трех маршрутах, на двух маршрутах – на дорогах, и на одном маршруте на склонах, где глубина снега составляла около 30–35 см.
ОБСУЖДЕНИЕ
Вопрос о численности дальневосточного леопарда – один из наиболее запутанных в оценке состояния его популяции.
Для определения численности дальневосточного леопарда долгое время использовался традиционный метод троплений следов на снегу и определение их половой, возрастной и индивидуальной принадлежности (Абрамов, Пикунов, 1974; Пикунов, Коркишко, 1992 и др.), который весьма субъективен. Позже этот метод был несколько модифицирован (Арамилев, Фоменко, 2000), но это еще больше увеличило субъективность оценки численности.
Субъективность получаемых оценок численности леопарда на основании существующих методик и их значительное занижение не удивительны. Согласно ряду наблюдений, дальневосточные леопарды приступают к охоте накануне снегопада, в условиях ограниченной видимости (Коркишко, 1983, 1986). Такое же поведение отмечено и в других северных популяциях леопарда (Лукаревский, 2001). После снегопадов эти хищники длительное время остаются недалеко от добычи или же “сидят” на ней. Именно поэтому оценки численности леопарда по следам после снегопадов зачастую значительно занижают значения численности. Т.е. метод маршрутного учета по следам после снегопада, используемый некоторыми авторами (Пикунов, Коркишко, 1992; Арамилев, Фоменко, 2000; Пикунов и др., 2009 и др.), не позволяет выявить значительную часть животных.
С появлением новых методов исследований (использование автоматических фотокамер) вопрос субъективности оценки численности должен был бы решиться. Однако из-за технических характеристик фотокамер и методических проблем эти вопросы оставались долгое время не решенными.
Так, например, используя компьютерные модели программы Capture-Recapture (Karanth, Nichols, 1998; Костыря и др., 2003), Арамилев с соавторами (Арамилев и др., 2010, 2012) получали на одной и той же площадке в разные годы двукратно различающиеся данные. По данным этих авторов, проводивших исследования на той же территории, что и мы, оценки численности в два раза отличаются от оценок, представленных нами.
Проведенные нами исследования с использованием той же методической основы – программы Capture-Recapture (Karanth, Nichols, 1998; Костыря и др., 2003) и модифицированной Ройял (Royle et al., 2009) – показывают, что на одной и той же территории, на одних и тех же точках регистрации с использованием одной и той же техники, в течение года мы получаем данные о численности и плотности популяции, различающиеся более чем на 30–40%.
Необходимо отметить, что продолжительность периода учета для использования программы Capture-Recapture составляет не более 2 месяцев. Особенности биологии и поведения леопарда таковы, что большая часть взрослых самок при такой продолжительности учета может остаться не зарегистрированной, также могут остаться незарегистрированными нерезидентные особи.
У леопарда отсутствует сезонность в размножении (Eaton, 1977; Коркишко, 1986; Пикунов, Коркишко, 1992; Лукаревский, 2001 и др.), и котята могут родиться в любое время года. В первые месяцы их жизни самка использует небольшую, наиболее продуктивную, часть участка обитания (Лукаревский, 2001; Рожнов и др., 2015), и только когда котята полностью переходят на твердую пищу, она начинает использовать весь участок обитания. Самка с котятами очень осторожна и подозрительно относится к незнакомым предметам, зачастую их избегает, поэтому остается практически “незаметной” для матрицы фотоловушек. Так, самка София, за которой мы вели наблюдения с помощью спутниковой телеметрии, ни разу не сфотографировалась ни на одной из четырех фотоловушек, установленных на ее участке обитания, пока ее котята были маленькие, тогда как резидентный самец (PP018M) регулярно снимался на них. Однако позже, когда котята достигли возраста 15–16 месяцев, София с котятами стала регулярно попадать в объективы на тех же фотоловушках. Данные Арамилева с соавторами (2012) также подтверждают, что значительная часть животных, в первую очередь самки и молодые звери, фотографируется не каждый год.
К сожалению, ни одна из компьютерных моделей – ни программа Capture–Recapture (Karanth, Nichols, 1998), ни программа SPACECAP (Royle et al., 2009) – не учитывает биологические особенности таких видов как леопард. А именно тот факт, что вероятность регистрации для разных категорий особей различна (рис. 5), приводит к искаженным результатам (табл. 2).
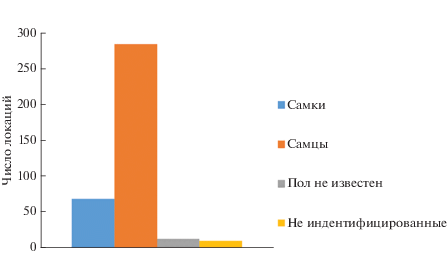
На результаты исследований, по-видимому, могут влиять природные условия, в частности, снегопады. Высокие показатели регистрации леопарда на фотоловушках зимой 2011–2012 гг. (рис. 4) в наших исследованиях обусловлены отсутствием осадков, тогда как зимой 2012–2013 гг. наоборот: глубина снежного покрова составляла в долинах от 40 до 50 см, на склонах до 30–45 см, а на гребнях хребтов образовывались надувы с глубиной снега до 120–150 см.
Высокие показатели встречаемости леопарда на фотоловушках в непосредственной близости от ИТС объясняются отсутствием факторов беспокойства на данных участках, а также высокой плотностью популяций копытных за линией ИТС. Лишь девять леопардов из 27, идентифицированных на модельном участке, не связаны с пограничной полосой. Из них четыре являются резидентами, участки обитания которых включают территорию заповедника “Кедровая падь” (Арамилев и др., 2012) и оленепарк в районе сел Занадворовка и Крауновка (наши неопубликованные данные).
Результаты, полученные с помощью фотоловушек в течение года на модельном участке, позволяют оценить численность дальневосточного леопарда как на территории Национального парка “Земля леопарда”, так и на территории всего ареала леопарда на Дальнем Востоке России.
Экстраполируя полученные нами данные только на территорию национального парка, куда входит наш модельный участок, получаем, что общая численность леопарда составляет 69 особей, из которых 49–50 – взрослые резидентные животные. Маршрутное обследование нами территории национального парка позволяет заключить, что около 15–18% его площади мало пригодна для обитания леопарда, поэтому оценка будет несколько ниже и составит около 57 леопардов.
Экстраполяция этих данных на всю территорию потенциальных местообитаний леопарда на российском Дальнем Востоке, т.е. на площадь 3502 км2 (Hebblewhite et al., 2011), позволяет заключить, что общая численность леопарда составляет около 96–100 особей, из которых около 60 животных являются резидентными, а 35–40 – особи, которые могут кочевать на большие расстояния. Однако, учитывая предыдущие учеты численности леопардов (Арамилев и др., 1999; Пикунов и др., 200022; Арамилев, Фоменко, 2000; Пикунов и др., 2009; Арамилев и др., 2010), следы которых были обнаружены на площади 2233 км2 (Мурзин А.А, Микелл Д., неопубликованные данные33), общая оценка численности, по-видимому, будет ниже и составит около 60–65 леопардов.
Таким образом, по самым осторожным оценкам, современная численность дальневосточного леопарда на российской части ареала составляет не 28–35 особей (Пикунов, Коркишко, 1985; Пикунов и др., 2009), а не менее 60–70 особей.
Учитывая общую площадь и характер местообитаний леопарда, при условии обеспечения эффективной борьбы с пожарами и браконьерством только на российской части ареала численность леопарда может быть увеличена до 150 особей, что позволит вывести вид из-под угрозы вымирания.
БЛАГОДАРНОСТИ
Авторы выражают искреннюю признательность за помощь и техническую поддержку в проведении полевых работ Кузьменко В.Б., Лазуренко А.Т., Подойницыну П.А., Чайникову И.Н., Яценко И.Д., директору Национального парка “Земля леопарда” Бородину А.В. Авторы благодарны Рожнову В.В. за ценные замечания, сделанные при подготовке авторами данной статьи.
Работа выполнена в рамках Постоянно действующей экспедиции РАН по изучению животных Красной книги Российской Федерации и других особо важных животных фауны России в рамках “Программы изучения, сохранения и восстановления дальневосточного леопарда на Российском Дальнем Востоке” при финансовой поддержке Русского географического общества.
Список литературы
Абрамов В.К., Пикунов Д.Г., 1974. Барс на Дальнем востоке СССР и его охрана // Бюллетень московского общества испытателей природы, отделение биологии. Т. 79. Вып. 2. С. 5–15.
Абрамов В.К., Пикунов Д.Г., Коркишко В.Г., 1994. Дальневосточный леопард и проблемы его сохранения в фауне мира // Природоохранные территории и акватории Дальнего Востока и проблемы сохранения биологического разнообразия: Материалы II науч. конф. Уссурийского заповедника. Владивосток. С. 95–98.
Арамилев В.В., Фоменко П.В., Микелл Д., 1999. Учет леопарда в 1998 г. // Зов тайги. № 4. С. 6–11.
Арамилев В.В., Костыря А.В., Соколов С.А., Рыбин А.Н., Макулла Д., Микелл Д.Дж., 2010. Мониторинг популяций дальневосточного леопарда (Panthera pardus orientalis) с помощью фотоловушек // Амурский тигр в Северо-Восточной Азии: проблемы сохранения в XXI веке. Междунар. науч.-практ. конф., 15–18 марта 2010 г., Владивосток. Владивосток: Дальнаука. С. 343–352 .
Арамилев В.В., Костыря А.В., Соколов С.А., Микелл Д.Г., Рыбин А.Н., и др., 2012. Дальневосточный леопард Panthera pardus orientalis в России. Атлас. Владивосток: Дальнаука. 88 с.
Коркишко В.Г., 1983. Зимняя охота леопарда на косулю // Механизмы поведения. Материалы III Всес. конф. по поведению животных. М. Т. 3. С. 60–61.
Коркишко В.Г., 1986. Экологические особенности и поведение дальневосточного леопарда. Автореф. дис… канд. биол. наук. М.: ИЭМЭЖ АН СССР. 26 с.
Коркишко В.Г., 1986а. Социальная организация дальневосточного леопарда // IV съезд Всесоюз. териол. общества. Тез. докл., II том, Москва, 27–31 января 1986 г. М., 1986. С. 136–137.
Костыря А.В., Белозор А.А., Микелл Д., Арамилев В.В., Котляр А.К., 2003. Применение фотоловушек для учетов амурского тигра // Териофауна России и сопредельных территорий (VII съезд Териологического общества). Материалы междунар. совещания 6–7 февраля 2003 г., Москва. М. С. 176–177.
Лукаревский В.С., 2001. Леопард, полосатая гиена и волк в Туркменистане. М.: Сигнар. 128 с.
Лукаревский В.С., Поярков А.Д., 2008. Современное состояние популяции ирбиса (Uncia uncia, Carnivora) в России // Зоологический журнал. Т. 87. № 1. С. 114–121.
Матюшкин Е.Н., 1977. Выбор пути и освоение территории амурским тигром (по данным зимних троплений) // Поведение млекопитающих. М.: Наука. С. 146–178.
Огастин Д., Микелл Д.Дж., Коркишко В.Г., 1996. Леопард выходит в эфир // Зов тайги. № 4. Вып. 27. С. 6–11.
Пикунов Д.Г., Коркишко В.Г., 1985. Современное распределение и численность леопарда (Panthera pardus) на Дальнем Востоке СССР // Зоологический журнал. Т. 64. № 6. С. 897–905.
Пикунов Д.Г., Коркишко В.Г., 1992. Леопард Дальнего Востока. М.: Наука. 192 с.
Пикунов Д.Г., Середкин И.В., Арамилев В.В., Николаев И.Г., Мурзин А.А., 2009. Крупные хищники и копытные юго-запада Приморского края. Владивосток: Дальнаука. 95 с.
Рожнов В.В., Найденко С.В., Эрнандес Бланко Х.А., Лукаревский В.С., Сорокин П.А., и др., 2012. Сезонные изменения кормовой базы амурского тигра: опыт применения матрицы фотоловушек // Зоологический журнал. Т. 91. № 6. С. 746–756.
Рожнов В.В., Сорокин П.А., Лукаревский В.С., Найденко С.В., Эрнандес-Бланко Х.А., Лукаревский С.В., 2013. Индивидуальная идентификация дальневосточных леопардов (Panthera pardus orientalis) молекулярно-генетическими методами и ее использование для оценки численности популяции // Известия РАН. Сер. биол. № 2. С. 138–143.
Рожнов В.В., Чистополова М.Д., Лукаревский В.С., Эрнандес-Бланко Х.А., Найденко С.В., Сорокин П.А., 2015. Участок обитания самки дальневосточного леопарда (Panthera pardus orientalis, Carnivora, Felidae), его структура и использование на протяжении годового цикла // Зоологический журнал. Т. 94. № 5. С. 593–603.
Чистополова М.Д., Эрнандес-Бланко Х.А., Лукаревский В.С., Найденко С.В., Сорокин П.А., Рожнов В.В., 2012. Использование пространства самцом дальневосточного леопарда после гибели соседствующего самца // V Всероссийская конф. по поведению животных. Москва, 20–23 ноября. С. 204.
Эрнандес-Бланко Х.А., Рожнов В.В., Лукаревский В.С., Найденко С.В., Чистополова М.Д., и др., 2013. Метод пространственно-эксплицитного повторного отлова (SECR, SPACECAP): новый подход к определению плотности популяции амурского тигра (Panthera tigris altaica) с помощью автоматических фоторегистраторов // Доклады Академии наук. Т. 453. № 2. С. 230–233.
Eaton R.L., 1977. Reproductive Biology of the Leopard // Zoologischer garten Bd. 47. H. 5. S. 329–351.
Hebblewhite M., Miquelle D.G., Murzin A.A., Aramilev V.V., Pikunov D.G., 2011. Predicting potential habitat and population size for reintroduction of the Far Eastern leopards in the Russian Far East // Biological Conservation 144. P. 2403–2413.
Karanth K.U., 1995.Estimating tiger populations from camera-trap data using capture-recapture models // Biological Conservation. V. 71. P. 333–338.
Karanth K.U., Nichols J.D., 1998. Estimation of tiger densities in India using photographic capture and recaptures // Ecology. V. 79. P. 2852–2862.
Karanth K.U., Kumar N.S., Nichols J.D., 2002. Field surveys: Statistical concepts: estimating absolute densities of tiger using capture-recaptures sampling // In: Monitoring tigers and their prey. Center Wildlife Studies, India. P. 139–150.
Royle J.A., Karanth K.U., Gopalaswamy A.M., Kumar N.S., 2009. Bayesian inference in camera trapping studies for a class of spatial capturerecapture models // Ecology. V. 90. P. 3233–3244.
Schaller G., Crawshaw P.G., 1980. Movement patterns of Jaguar // Biotropica. V. 12. № 3. P. 161–168.
Sugimoto T., Nagata J., Aramilev V.V., McCullough D.R., 2012. Population size estimation of Amur tigers in Russian Far East using noninvasive genetic samples // Journal of Mammalogy. V. 93. № 1. P. 9–101.
Wilson K.R., Anderson D.R., 1985. Evaluation of two density estimators of small mammal population size // Journal of Mammalogy. V. 66. P. 13–21.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал