

Зоологический журнал, 2019, T. 98, № 6, стр. 616-627
Энтомокомплекс трупов крупных животных и особенности их разложения на севере европейской части России
С. Н. Лябзина 1, *, О. С. Лаврукова 1, А. Н. Приходько 2, **, А. И. Азовский 3, В. Л. Попов 4
1 Петрозаводский государственный университет
185910 Петрозаводск, Россия
2 Бюро судебно-медицинской экспертизы Республики Карелия
185035 Петрозаводск, Россия
3 Биологический факультет Московского государственного университета им. М.В. Ломоносова
119991 Москва, Россия
4 Первый Санкт-Петербургский государственный медицинский университет им. И.П. Павлова
197022 Санкт-Петербург, Россия
* E-mail: slyabzina@petrsu.ru
** E-mail: andrey_prihodko@list.ru
Поступила в редакцию 01.04.2018
После доработки 24.06.2018
Принята к публикации 08.07.2018
Аннотация
На примере домашней свиньи (Sus scrofa domesticus L.) рассмотрен энтомокомлекс трупов крупных животных севера европейской части России (Республика Карелия) в двух типах биотопов. Некрофильный энтомокомплекс насчитывает 100 видов членистоногих, относящихся к 25 семействам и пяти отрядам. Между биотопами имеются различия по видовому и количественному составу фауны некрофилов. Анализируются структура комплекса и сукцессия сообществ насекомых в процессе разложения трупа.
В природе трупы или их останки чаще всего подвергаются воздействию некробионтов, которые приводят материал к деструкции и минерализации. Основную роль в этих процессах выполняют двукрылые и жесткокрылые. Состав фауны трупа часто исследуется на небольших приманках или фрагментах органических субстратов (печень, фарш) (Nuorteva, Hasanen, 1972; Kozminykh, Esynin, 1994; Минаев, Пушкин, 2016 и др.). Однако особый интерес представляет изучение процесса разложения туш крупных животных (свыше 50 кг), который приравнивают к процессам гниения человеческих тел. В последнее время было выполнено несколько работ в этом направлении (Caballero, León-Cortés, 2014; Anderson et al., 2015; Matuszewski et al., 2017). Результаты этих экспериментов позволяют выявить не только разнообразие насекомых-некробионтов, но и определить их участие в ходе разложения для возможного применения в экспертных случаях судебной энтомологии.
МАТЕРИАЛ И МЕТОДИКА
Исследования проводили в течение 2015–2017 гг. в Прионежском р-не Карелии (Лососиное, 61.3° с.ш., 34.0° в.д.). Район характеризуется преобладанием старых (до 140 лет) еловых лесов с елью обыкновенной (Picea abies (L.) Karst.) и сибирской (P. obovata Ldb.), в подросте преобладают береза повислая (Betula pendula Roth), ольха серая (Alnus incana L.). Открытые биотопы представлены большей частью болотами и, в меньшем количестве, разнотравными лугами, которые, как правило, граничат с ельниками.
Исследования проводили на четырех трупах домашней свиньи (Sus scrofa domesticus L.) массой 65–100 кг. По две приманки были размещены в двух биотопах (в ельнике черничном и на разнотравном лугу). Закладку туш осуществляли в июне 2015 и 2016 гг. Умерщвление животных было произведено без нарушения анатомической целостности кожных покровов, после чего в течение двух часов их размещали в местах исследований.
Методика постановки опыта описана нами ранее (Лябзина и др., 2016). Приманку помещали в металлическую клетку (120 × 80 × 60 см, размер ячеи сверху и сбоку 10 см, снизу 20 см), которую прочно фиксировали с помощью длинных металлических прутьев (рис. 1). Только такие меры защиты позволяют уберечь еe от позвоночных и провести длительные наблюдения. Изначально, в первых двух опытах, эти условия полностью не были соблюдены и крупные падальщики смогли утащить заложенные туши свиней.

Сбор материала проводили с помощью двух типов ловушек (пирамидальной, почвенной) и ручным способом. Пирамидальную ловушку использовали для отлова летающих насекомых, привлеченных разлагающейся органикой. Ловушка представляла собой капроновую сеть, которую с помощью шеста устанавливали на высоту около 1.5 м. Сеть закрывала клетку с четырех сторон так, чтобы снизу оставался небольшой зазор для проникновения к приманке. Через отверстие в вершинной части ловушки летающие насекомые попадали в ловчий стакан, наполненный водой для фиксации. Такой тип ловушек считается одним из самых эффективных способов отлова с трупов как двукрылых, так и большинства жесткокрылых (Cruise et al., 2018). Для сбора напочвенных насекомых использовали пол-литровые банки, которые заполняли наполовину водой и размещали на расстоянии до 0.5 м от трупа.
Во время исследований вели количественный учет и собирали насекомых, находящихся на трупе, в его ложе и пойманных в ловушки, а для сохранения естественного хода разложения, изымали не более 5–10% особей. Исключением являлись муравьи, которые присутствовали в большом количестве, поэтому учитывали только их наличие. Сбор проводили регулярно три раза в неделю в течение месяца после закладки и в дальнейшем один раз в неделю до полного разложения. Личинок насекомых (двукрылых, жесткокрылых), собранных с трупа, доращивали в лабораторных условиях в термостате на кусочках куриной или говяжьей печени, рыбе, либо на фрагментах гниющей органики, изъятых непосредственно с приманки. Исследования проводили в весенне-осенний период в течение трех лет. Всего было собрано более 25 000 особей имаго и преимагинальных стадий, из них более четверти выведено в лабораторных условиях.
Определения видов отдельных семейств проверены специалистами: Carabidae – С.Д. Узенбаевым (Петрозаводский государственный университет); Staphylinidae – В.Б. Семеновым (Институт медицинской паразитологии и тропической медицины им. Е.И. Марциновского, ПМГМУ им. И.М. Сеченова); Histeridae и Leiodidae – В.К. Зинченко (Институт систематики и экологии животных Сибирского отделения РАН), Sepsidae и Piophilidae – А.Л. Озеровым (Зоологический музей Московского государственного университета).
В качестве характеристик разнообразия некробионтного сообщества использовали число видов на пробу и индекс Шеннона–Уивера (Н ').
Для анализа изменений в составе некробионтов и выделения основных этапов сукцессии применяли анализ соответствий с удаленным трендом (Detrended Correspondence Analysis, DCA). Этот метод непрямой ординации позволяет упорядочивать распределение объектов (видов и описаний) вдоль осей ординации, максимально сохраняя взаимное расположение описаний, оцененное как сходство между ними по видовой структуре. Достоинством метода является использование нелинейной (унимодальной) модели отклика, т.е. распределения обилия видов вдоль осей ординации (Джонгман и др., 1999). Кроме того, процедура удаления тренда (detrending) устраняет “эффект арки”, свойственный простому анализу соответствий. Однако этот метод может сглаживать реально существующие изменения и скрытые градиенты, которые формируют группировки, и границы между ними не всегда могут быть выявлены достоверно (Шитиков и др., 2003).
Для интерпретации осей ординации использовали величины линейной корреляции между положением проб на оси, сроком отбора (в сутках с момента закладки эксперимента) и накопленной суммой эффективных положительных температур. Последний показатель характеризует количество тепла и выражается суммой средних суточных температур воздуха за рассматриваемый период, превышающих 0°С – температурный порог развития личинок и лёта насекомых. Информацию по температуре воздуха района исследования получали с официального сайта по погодным условиям Гисметео (http://gismeteo.ru). Расчеты проведены в программе PAST 3.14 (Hammer et al., 2001).
РЕЗУЛЬТАТЫ
Всего было зарегистрировано 100 видов насекомых, относящихся к 25 семействам и шести отрядам (табл. 1). Большая часть по видовому составу представлена двумя отрядами – жесткокрылые (65) и двукрылые (27). Остальные восемь видов принадлежат к отрядам перепончатокрылых, клопов, тараканов и чешуекрылых. Среди отмеченных насекомых на трупах преобладали жесткокрылые (Necrobia violacea, Sciodrepoides watsoni, Aleochara curtula, Atheta aeneipennis) и двукрылые (Protophormia terraenovae, Hydrotaea dentipes) и два вида из семейства Piophilidae (табл. 1).
Таблица 1.
Видовой состав насекомых, их обилие в исследуемых биоценозах и присутствие на различных этапах в процессе разложения трупа домашней свиньи (Sus scrofa domesticus)
| Таксон | Стадии насекомых, встреченные на трупе | Численность насекомых в исследуемых биоценозах | Этапы разложения | ||||
|---|---|---|---|---|---|---|---|
| ельник черничный | разнотравный луг | раннего | активного | позднего | распад костной ткани | ||
| Blattodea | |||||||
| Ectobiidae | |||||||
| Ectobius lapponicus (Linnaeus 1758) | A, Im | 10 | – | + | + | ||
| Hemiptera | |||||||
| Pyrrhocoridae | |||||||
| Pyrrhocoris sp. | A, Im | 9 | 2 | + | + | ||
| Coleoptera | |||||||
| Carabidae | |||||||
| Carabus glabratus (Paykull 1790) | A | 6 | 4 | + | + | ||
| Pterostichus aethiops (Panzer 1797) | A | 5 | – | + | |||
| P. niger (Schaller 1783) | A | 10 | 14 | + | + | ||
| P. oblongopunctatus (Fabricius 1787) | A | 4 | 2 | + | + | ||
| Cleridae | |||||||
| Necrobia violacea (Linnaeus 1758) | A, Im | 800 | 500 | + | + | + | |
| Elateridae | |||||||
| Prosternon sp. | Im | 4 | 8 | + | |||
| Histeridae | |||||||
| Hister unicolor (Linnaeus 1758) | A | 36 | 55 | + | |||
| Margarinotus brunneus (Fabricius 1775) | A, Im | 130 | 86 | + | + | ||
| M. striola (Sahlberg 1819) | A, Im | 24 | 5 | + | + | ||
| Saprinus semistriatus (Scriba 1790) | A, Im | 123 | 260 | + | + | ||
| Hydrophilidae | |||||||
| Cercyonlateralis (Marsham 1802) | A | 15 | 9 | + | + | ||
| Cryptopleurum crenatum (Panzer 1794) | A | 6 | – | + | |||
| C. minutum (Fabricius 1775) | A | 45 | – | + | + | ||
| Megasternum obscurum (Marsham 1802) | A | 25 | – | + | |||
| Leiodidae | |||||||
| Apocatops nigrita (Erichson 1837) | A | 460 | 40 | + | + | + | |
| Sciodrepoides alpestris (Jeannel 1934) | A | 98 | – | + | + | ||
| S. fumatus (Spence 1815) | A | 140 | – | + | + | ||
| S. watsoni (Spence 1815) | A | 1470 | 450 | + | + | + | |
| Nitidulidae | |||||||
| Nitidula bipunctata (Linnaeus 1758) | A, Im | 230 | 500 | + | + | ||
| Omosita colon (Linnaeus 1758) | A, Im | 60 | 10 | + | |||
| O. depressa (Linnaeus 1758) | A, im | 90 | 60 | + | + | ||
| Scarabaeidae | |||||||
| Aphodius rufipes (Linnaeus 1758) | A | 5 | 60 | + | |||
| Geotrupes stercorarius (Linnaeus 1758) | A | – | 5 | + | |||
| G. stercorosus (Scriba 1791) | A | 500 | 140 | + | + | ||
| Silphidae | |||||||
| Necrodes littoralis (Linnaeus 1758) | A, Im | 170 | 150 | + | |||
| Nicrophorus invetstigator (Zetterstedt 1824) | A | 25 | 12 | + | |||
| N. vespillo (Linnaeus 1758) | A | 2 | 3 | + | |||
| N. vespilloides (Herbst 1793) | A | 40 | 30 | + | |||
| Oiceoptoma thoracica (Linnaeus 1758) | A, Im | 165 | – | + | + | ||
| Thanatophilus dispar (Herbst 1793) | A, Im | – | 130 | + | + | ||
| Th. rugosus (Linnaeus 1758) | A, Im | – | 5 | + | + | ||
| Th. sinuatus (Fabricius 1775) | A, Im | – | 205 | + | + | ||
| Staphylinidae | |||||||
| Aleochara curtula (Goeze 1777) | A, Im | 390 | 340 | + | + | ||
| Anthobium atrocephalum (Gyllenhal 1827) | A | 5 | – | + | |||
| Atheta aeneipennis (Thomson 1856) | A | 530 | 490 | + | + | + | |
| A. corvina (Thomson 1856) | A | 15 | – | + | |||
| A. crassicornis (Fabricius 1792) | A | 420 | 600 | + | + | ||
| A. dadopora (Thomson 1867) | A | 43 | – | + | |||
| A. gagatina (Baudi 1848) | A | 430 | 250 | + | + | ||
| A. nigripes (Thomson 1856) | A | – | 42 | + | |||
| A. paracrassicornis (Brundin 1954) | A | 680 | 450 | + | + | ||
| A. subtilis (Scriba 1866) | A | 260 | 200 | + | |||
| Creophilus maxillosus (Linnaeus 1758) | A, Im | 135 | 80 | + | + | ||
| Lypoglossa lateralis(Mannerheim 1830) | A | 1 | – | + | |||
| Megarthrus depressus (Paykull 1789) | A | 120 | 85 | + | + | ||
| Omalium rivulare (Paykull 1789) | A | 110 | 75 | + | |||
| O. septentrionis (Thomson 1857) | A | 40 | 15 | + | |||
| Ontholestes murinus (Linnaeus 1758) | A | 65 | 70 | + | + | ||
| O. tessulatus (Geoffroy 1785) | A | 95 | 80 | + | + | ||
| Oxypoda alternans (Gravenhorst 1802) | A | 1 | – | + | + | ||
| Pella humeralis (Gravenhorst 1802) | A | 13 | – | + | |||
| Philonthus addendus (Sharp 1867) | A | 76 | 40 | + | |||
| Ph. decorus (Gravenhorst 1802) | A | 12 | – | + | |||
| Ph. fumarius (Gravenhorst 1802) | A | 7 | – | + | |||
| Ph. marginatus (Müller 1764) | A | 10 | 65 | + | |||
| Ph. politus (Linnaeus 1758) | A, Im | 145 | 120 | + | + | ||
| Ph. rectangulus (Sharp 1874) | A | 1 | – | + | |||
| Ph. succicola (Thomson 1860) | A, Im | 137 | 105 | + | + | ||
| Ph. varians (Paykull 1789) | A | 34 | 7 | + | |||
| Proteinus brachypterus (Fabricius 1792) | A | 155 | 70 | + | |||
| Quedius lucidulus (Erichson 1839) | A | 22 | 6 | + | |||
| Tachinus laticollis (Gravenhorst 1802) | A | 225 | 125 | + | |||
| T. marginellus (Fabricius 1781) | A | 90 | 2 | + | |||
| T. pallipes (Gravenhorst 1806) | A | 17 | – | + | |||
| T. proximus (Kraatz 1855) | A | 135 | – | + | |||
| Diptera | |||||||
| Calliphoridae | |||||||
| Calliphora vicina (Robineau-Desvoidy 1830) | A, Im | 230 | 300 | + | |||
| C. vomitoria (Linnaeus 1758) | A, Im | 400 | 430 | + | |||
| Cynomya mortuorum (Linnaeus 1761) | A, Im | 70 | 80 | + | |||
| Protophormia terraenovae (Robineau-Desvoidy 1830) | A, Im | 830 | 750 | + | + | ||
| Lucilia caesar (Linnaeus 1758) | A, Im | 400 | 530 | + | + | ||
| L. silvarum (Meigen 1826) | A, Im | 75 | 420 | + | + | ||
| L. illustris (Meigen 1826) | A, Im | 112 | 580 | + | + | ||
| Dryomyzidae | |||||||
| Dryomyza anilis (Fallén 1820) | A | 2 | 1 | + | |||
| Fanniidae | |||||||
| Fannia canicularis (Linnaeus 1761) | A, Im | 270 | 540 | + | + | ||
| F. coracina (Loew 1873) | A, Im | 10 | 50 | + | |||
| F. scalaris (Fabricius 1794) | A, Im | 20 | 54 | + | |||
| F. serena (Loew 1873) | A, Im | 9 | 7 | + | |||
| Fannia spp. | A, Im | 20 | 200 | + | + | ||
| Helomyzidae | |||||||
| Helomyza serrata (Linnaeus 1758) | A, Im | 270 | 180 | + | + | ||
| Muscidae | |||||||
| Hydrotaea dentipes (Fabricius 1805) | A, Im | 340 | 160 | + | |||
| Hydrotaea spp. | A, Im | 175 | 530 | + | |||
| Muscina levida (Harris 1780) | A, Im | 420 | 560 | + | |||
| Piophilidae | |||||||
| Parapiophila vulgaris (Fallén1820) | A, Im | 450 | 400 | + | + | ||
| Stearibia nigriceps (Meigen 1826) | A, Im | 500 | 550 | + | + | + | |
| Sarcophagidae | |||||||
| Helicophagella melanura (Meigen 1826) | A, Im | 125 | 55 | + | |||
| Sarcophaga albiceps (Meigen 1826) | A | 55 | 152 | + | |||
| S. carnaria (Linnaeus 1758 ) | A | 20 | 60 | + | |||
| S. caerulescens (Zetterstedt 1838) | A, Im | 20 | 106 | + | |||
| Sepsidae | |||||||
| Nemopoda nitidula (Fallén 1823) | A | 105 | 165 | + | |||
| Sepsis punctum (Fabricius 1794) | A | 50 | 145 | + | |||
| Sphaeroсeridae | |||||||
| Sphaerocera sp. | A | 4 | 4 | + | |||
| Tipulidae | |||||||
| Tipula sp. | A, Im | 5 | – | + | |||
| Hymenoptera | |||||||
| Vespidae | |||||||
| Vespula vulgaris (Linnaeus 1758) | A | 6 | 12 | + | |||
| Braconidae | |||||||
| Alysia manducator (Panzer 1799) | A, Im | 56 | 120 | + | |||
| Formicidae | |||||||
| Formica polyctena (Forster 1850) | A | * | – | + | + | + | |
| Lasius niger (Linnaeus 1758) | A | – | * | + | + | + | |
| Myrmica rubra (Linnaeus 1758) | A | – | * | + | + | + | |
| Lepidoptera | |||||||
| Tineidae | |||||||
| sp. | A | 6 | 2 | + | |||
| Количество видов | 93 | 79 | 9 | 74 | 65 | 6 | |
| Количество особей | 14 181 | 13 205 | |||||
Наибольшее число (93 вида) некрофильных представителей выявлено в ельнике черничном (табл. 1). Кроме общих доминирующих видов, здесь преобладали также жуки Atheta paracrassicornis, A. dadopora, Philonthus politus, Tachinus laticollis и двукрылые Sepsis punctum. Для этого биотопа характерен 21 вид, из них восемнадцать жесткокрылых (табл. 1). Например, представители Tachinus pallipes, T. proximus, Oxypoda alternans и Lypoglossa lateralis могут быть связаны не только с трупами, но и с почвенным покровом, разлагающимися растительными остатками или грибами. Также выраженную приуроченность к лесным ландшафтам проявляли жесткокрылые Oiceoptoma thoracica, Apocatops nigrita и Sciodrepoides alpestris. В течение всего длительного процесса разложения на приманках были многочисленны личинки жуков рода Philonthus и лесные муравьи рода Formica.
На лугу отмечено меньшее число (79 видов) некробионтов (табл. 1). Исключительно в открытых ландшафтах встречались поверхностно-падальные мертвоеды рода Thanatophilus, из стафилинид чаще отлавливали Atheta crassicornis и A. nigripes. В таких биоценозах в теплую погоду на приманках иногда попадался Geotrupes stercorarius.
В целом состав некрофильных насекомых в ельнике черничном и на разнотравном лугу весьма схож: 72 вида встречены в обоих исследованных биотопах. Наибольшая доля эврибионтов среди некрофильных двукрылых – 99% (26 видов). Высокая степень сходства связана с активным перемещением насекомых в поисках гниющих субстратов.
Насекомые связаны с трупом на разных стадиях развития: неполовозрелой – яйцо, личинка, куколка (Im – immature) и половозрелой (имаго А – adult) (табл. 1). При этом для одних видов труп может быть источником пищи, для других – местом обитания преимагинальных стадий. Известно, что для полноценного овогенеза самкам насекомых необходимо белковое питание. Вероятно, поэтому имаго некоторых жуков, например водолюбов (Hydrophilidae) и навозников (Scarabaeidae), оказываются привлеченными разлагающимся веществом в качестве питательного субстрата.
Среди насекомых выявлено более 40 видов, часть жизненного цикла которых проходит на трупе, это жуки Silphidae, Nitidulidae и Staphylinidae, а также большая часть двукрылых (табл. 1). Муха P. terraenovae одна из первых обнаруживает падаль и откладывает яйца в большом количестве в виде комочков разных размеров, располагая их по всему телу. В дальнейшем её личинки остаются на поверхности трупа, активно питаются и тут же пупаризируются, образуя массовое скопление. Представители семейства Piophilidae также обильно колонизируют трупы, но перед пупаризацией всегда перемещаются в его ложе. Куколок некоторых жесткокрылых (Necrodes littoralis, Philonthus succicola, Creophilus maxillosus) также можно найти в верхнем почвенном слое вблизи трупа, а другие виды N. violacea, рода Omosita перезимовывают на поверхности останков кожи и костей.
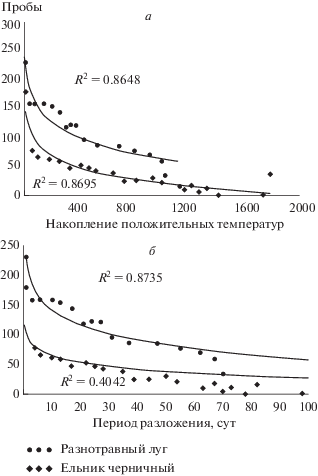
Во время тления меняются состояние и структура всех тканей, а также их химический состав, все это приводит к изменению видового состава в количественном и качественном отношении. Анализ соответствий с удаленным трендом выявил общие и специфичные черты изменения состава насекомых в процессе разложения трупов свиней в двух исследуемых биотопах (рис. 2). Первая ось ординации объясняет бόльшую часть (более 60%) общей изменчивости (собственное число оси равно 0.22) и отражает главным образом ход сукцессии (справа налево). Это подтверждается и тем, что положение последовательных сборов на этой оси сильно коррелирует со сроком сбора (в сутках от начала эксперимента, r = –0.935 и –0.636 для луга и леса, соответственно) (рис. 3). Еще теснее положение сборов на первой оси коррелирует с суммой эффективных (неотрицательных) температур: r = –0.930 и –0.932, соответственно). В целом смена состава некробионтов ярче выражена на лугу. Вторая (вертикальная) ось ординации объясняет существенно меньший процент изменчивости (собственное число – 0.07) и в основном отражает различия между биотопами (лес и луг), особенно заметные на ранних стадиях сукцессии (рис. 2).
Рис. 2.
Пространственно-временная изменчивость населения некрофильных насекомых в исследуемых биотопах и основные этапы сукцессии (анализ соответствий с удалением тренда). Первая (горизонтальная) ось ординации описывает изменение состава некробионтов в процессе разложения и ход сукцессии (справа налево), вторая (вертикальная), ось отражает различия в составе некрофильных сообществ в двух биотопах между биоценозами. Обозначения: F (forest) – лес, M (meadow) – луг, F(или М)1–366 – день разложения, овалами выделены основные стадии сукцессии, стрелки показывают направление хода сукцессии.
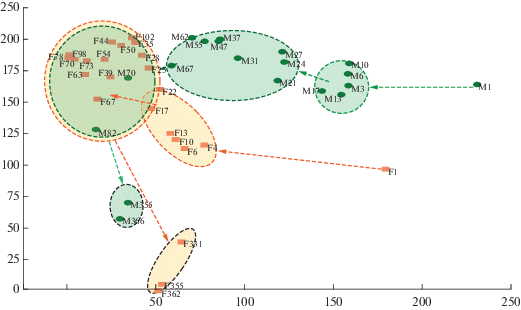
Рис. 3.
Связь положения проб на первой оси ординации с накопленной суммой положительных температур (а) и периода разложения (сут) (б).
В ходе сукцессии прослеживаются несколько стадий, отличающихся друг от друга количественным и видовым составом некробионтов, а также состоянием трупных тканей. В ранний период разложения, который начинается от момента смерти и длится в течение нескольких дней, на трупах отмечали лишь отдельные виды (табл. 1). В первые дни (спустя сутки после закладки эксперимента) на тушах в обоих биотопах зарегистрированы лишь двукрылые Lucilia caesar и L. illustris. Самки активно откладывали яйца в слизистые участки головы, забираясь глубоко внутрь носовой и ротовой полостей, либо около ресниц. Все это позволяло личинкам младших возрастов быстро перейти на питание мягкими тканями. Через два дня в ложе трупа отлавливали падальных жуков из семейства Leiodidae. В этот период процесс гниения не вызывал видимых внешних изменений трупа, и разложение сопровождалось аутолизом внутренних тканей и охлаждением тела.
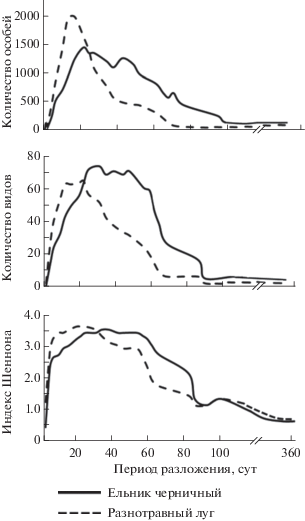
Вторая стадия активного разложения наступает на 3–4-е сутки и длится около месяца в открытых биоценозах и около двух месяцев в лесных. В видовом и количественном соотношении она отличается наибольшим числом некробионтов, а индекс Шеннона-Уивера имеет максимальные значения (рис. 4). Наблюдался стремительный рост обилия посетителей трупа (табл. 1).
Рис. 4.
Динамика общей численности насекомых некробионтов, видов и индекса Шеннона с продолжительностью периода разложения (сут) в двух биотопах трупа домашней свиньи (Sus scrofa domesticus).
В этот период доминировали представители синих мясных мух из родов Calliphora, Cynomya, Lucilia и Protophormia, суммарно составлявшие около четверти всей численности посетителей. Способность этих двукрылых быстро обнаруживать и колонизировать труп позволяет относить их к числу видов, имеющих важное судебно-медицинское значение. Несколько позже попадались Sarcophagidae и Muscidae. Других представителей – Fannia canicularis, Parapiophila vulgaris – также обильно отлавливали на приманках через неделю. В период, когда ткани сильно разжижены, всегда многочисленны Sepsidae и Helomyzidae. Большинство присутствующих двукрылых откладывали яйца или отрождали личинок, постепенно заполняя всю поверхность туловища.
Жесткокрылые появлялись на два–три дня позже двукрылых, и в процессе разложения наблюдалась смена их видового состава. Время появления жуков было связано как с состоянием трупных тканей, так и присутствием других посетителей, для которых эти ткани непосредственно являются пищей. Например, представителей родов Oiceoptoma, Tachinus, Necrodes, Geotrupes привлекали слегка подгнившие мягкие ткани, а виды родов Philonthus и Aleochara обнаруживались, когда биомасса личинок двукрылых была наибольшей. Однако некоторые виды жесткокрылых (например, Necrobia violacea, Apocatops nigrita, Sciodrepoides watsoni, Atheta aeneipennis) и двукрылых (Stearibia nigriceps), а также муравьи находились на приманках в течение большей части времени разложения. На этой стадии широко представлены различные трофические группы насекомых: некро-, сапро-, зоофагов и паразитоидов, и такое разнообразие приводило к утилизации основной массы мягких тканей.
В течение второй стадии резко менялось состояние трупных тканей. К концу первой недели развивалась трупная эмфизема, обусловленная формированием гнилостных газов по всему телу, это приводило к набуханию и значительному увеличению объема туловища. Поверхность кожи становилась осклизлой и грязно-зеленого цвета, а вследствие гниения внутренних органов из естественных отверстий обильно вытекала грязновато-бурая жидкая масса. В дальнейшем за счет пищеварительных ферментов некробионтов, субстрат сильно размягчался, а активное их перемещение образовывало многочисленные отверстия и нарушалась целостность кожных покровов.
При высокой влажности происходил процесс омыления, при этом в образующейся пене скапливались личинки двукрылых, что вероятно, защищало их от пересыхания и других воздействий факторов внешней среды. Несмотря на кажущуюся общую бесформенную личиночную массу, находящуюся на гниющем субстрате, многие виды двукрылых локализовались в определенных местах с учетом своих экологических особенностей. В большом числе около трупа скапливались личинки мух P. terraenovae; видимо, такое совместное обитание облегчает протекание метаболических процессов в организме. Личинки младших возрастов из родов Lucilia и Cynomya группировались вместе, а подросшие особи самостоятельно активно перемещались, углубляясь в почву на несколько сантиметров. Наоборот, личинки Sarcophagidae всегда располагались отдельно от других некробионтов и при малейшей тревоге впадали в танатоз.
Активное воздействие разлагающегося органического вещества и обитателей приводило к образованию хорошо выраженного ложа трупа. Местоположение приманки можно было обнаружить и на следующий год по повреждённому растительному и почвенному покровам.
Следующая стадия – позднего разложения – наступала через один-два месяца и в лесу продолжалась до конца летнего сезона, а на лугу несколько недель. В ходе этого этапа численность и разнообразие некробионтов на лугу постепенно снижалась, тогда как в лесу количественные показатели продолжали оставаться высокими (рис. 4). Процессы деструкции наружных покровов и сухожилий происходили за счет жуков и их личинок сапро- и кератофагов (рода Atheta, Omalium, Tachinus, Geotrupes, Omosita, Necrobia, Nitidula). На этом этапе также встречалось незначительное количество хищных личинок жесткокрылых (Philonthus spp., Creophilus maxillosus) и двукрылых (родов Sarcophaga, Fannia).
В этот период происходит разрушение покровов, и продолжительность этапа зависит от обилия осадков и их интенсивности. Например, частые дожди приводили к быстрому размыванию последних связующих элементов трупа.
Завершающий этап разложения – распад костной ткани – заключается в окончательной деструкции костных останков, волос и других кератинсодержащих компонентов. В лесных биоценозах он начинался на следующий год после закладки, а на лугу – через два месяца. Период отличался резким снижением количественного и видового состава насекомых и низкими показателями индекса разнообразия (рис. 4). На последних этапах в утилизации участвовали лишь небольшая группа некробионтов – это жуки–кератофаги (Omosita, Necrobia, Nitidula) и личинки двукрылых (рода Helomyza, Stearibia) (табл. 1). Кроме того, среди всех этапов разложения этот этап самый продолжительный. Например, в экспериментах на лугу в течение двух лет наблюдений не завершилась полная утилизация останков свиньи, и на ложе оставались клочки волос, кости скелета и черепа (верхняя и нижняя челюсти, ребра).
Проведенные эксперименты в открытых и лесных биотопах позволили отметить общие и специфичные закономерности заселения трупов видами. Обнаружение приманок некробионтами и их колонизация в местах исследования различались – на лугах эти процессы происходили значительно быстрее. Например, наибольшее видовое и количественное разнообразие жесткокрылых и двукрылых на разнотравном лугу регистрировали на 15–20-е сутки (рис. 4). Однако быстрая смена стадий активного и позднего разложения сокращало время присутствия большинства видов. В лесу, наоборот, умеренные колебания температуры воздуха и повышенная влажность заметно затягивали колонизацию трупов свиней, и гниющая органика достаточно длительный период времени привлекала посетителей. Одной из общих тенденцией разложения является то, что одни и те же виды жесткокрылых и двукрылых встречались в ложе трупа на следующий год; в почве в большом количестве находились преимагинальные стадии и имаго Necrobia violacea и Stearibia nigriceps.
ОБСУЖДЕНИЕ
Выявленный видовой состав некробионтов на крупных приманках дополняет список насекомых, ранее отмеченных в южной Карелии на тушках средних размеров (Лябзина, 2011). Общий список некрофауны пополнился за счет Staphylinidae, а длительный период разложения позволил более полно выявить разнообразие двукрылых, связанных в своем развитии с гниющими субстратами. В целом на севере европейской части России состав насекомых некробионтов незначительно уступает разнообразию в центральной Европе. Например, на севере Чешской Республики фауна жесткокрылых-некробионтов насчитывает 145 видов (Kočárek, 2003).
Трофические взаимоотношения членистоногих с трупом и друг с другом очень разнообразны. Среди них выделяют некро-, керато-, сапрофагов, кожеедов, паразитоидов, а также группы со смешанным типом питания – некро-зоофаги и др. (Braack, 1987; Марченко, 1992). Такое многообразие способствует полной и достаточно быстрой утилизации в природе различных органических компонентов. Кроме того, по степени приуроченности к этому типу местообитаний выделяют три группы (Smith, 1986). Облигатные некробионты развиваются исключительно в трупах, факультативные могут обитать на различных эфемерных субстратах (экскременты, навоз, гнезда, растительные остатки, грибы и т.д.), случайные посетители чаще встречаются в иных местобитаниях. Первая группа некробионтов, личинки которых развиваются только в трупных тканях, представлена одиннадцатью видами, из которых большая часть – жесткокрылые, это (представители родов Nicrophorus, Thanatophilus, Necrodes, Oiceoptoma, Nitidula, Omosita) и два вида двукрылых (Cynomya mortuorum и Sarcophaga caerulescens). Их личинки являются облигатными некрофагами, а взрослые особи – полифаги и могут встречаться на разнообразных эфемерных субстратах. Например, O. thoracica и N. vespilloides обитают на грибах, другие виды рода Nicrophorus при конкуренции на трупе нападают на двукрылых (Kozminykh, Esynin, 1994), а самцы и самки C. mortuorum, питаясь только черным хлебом в лабораторных условиях, могут быть активны более десяти дней (Лябзина и др., 2016а). В эту группу включены представители семейства кожеедов (Dermestidae), но их присутствие на субстратах находится в прямой зависимости от влажности (Жантиев, 2009). В исследуемых биотопах виды этого семейства не отмечены, но в северных районах республики в сухих биоценозах, например в сосняке черничном, они попадаются на трупных приманках средней массы (Лябзина, 2011).
Наиболее многочисленна группа факультативных некробионтов. В неё входят жуки, перепончатокрылые и большинство двукрылых. Из встреченных нами, стафилиниды также характеризуются сочетанием различных типов имагинального питания и встречаются на иных гниющих субстратах. Например, А. paracrassicornis обитает на грибах, в навозе и сочетает сапро- и зоофагию, для Creophilus maxillosus и некоторых видов рода Tachinus характерна некро- и зоофагия (Тихомирова, 1973; Kozminykh, Esynin, 1994; Kočárek, 2003; Бельская, Колесникова, 2011). Паразитоиды Alysia manducator и Aleochara curtula развиваются в основном в личинках двукрылых.
Группа случайных посетителей немногочисленна и включает представителей нескольких отрядов, например таракана (Ectobius lapponicus), клопа (Pyrrhocoris sp.), жужелиц (рода Carabus, Pterostichus) и осу (Vespula vulgaris). Их трофические взаимосвязи с трупом определяются как сапро- и зоофагия, но из-за низкой численности воздействовать на скорость утилизации тканей они не могут.
Описанный выше ход некробиотической сукцессии сходен с процессом разложения, описанным другими исследователями. Во всех случаях отмечают несколько стадий разложения, каждой из которых соответствует определённый видовой состав посетителей (Smith, 1986; Anderson, 2015; Matuszewski, 2017; Марченко, 1992). Изменение тканей, происходящее в процессе разложения субстрата, приводит к хорошо выраженному последовательному заселению некробионтами.
Различия между биотопами проявляются не только в видовом составе некробионтов, но и в скорости утилизации тканей. Динамичные абиотические факторы открытых биоценозов (инсоляция, перепады температур и влажности и проч.) ускоряют ход начальных стадий сукцессии и, наоборот, сглаженность этих условий в лесах замедляют протекание этого этапа. Прежде всего, различия выражаются в укорочении второй стадии разложения (Марченко, 1992; Caballero, León-Cortés, 2014).
В течение всего времени разложения труп может функционировать как микробиотоп. Многообразие трофических связей и взаимоотношений посетителей, как с трупом, так и между собой, и позволяет какое-то время ему существовать автономно. Кроме того, на определенном этапе разложения температура внутри субстрата значительно выше, чем снаружи, за счет эндогенного тепла, выделяемого личинками двукрылых при их метаболизме (Anderson, 2015; Марченко, 1992; Лаврукова и др., 2017).
Список литературы
Бельская Е.А., Колесникова А.А., 2011. Видовой состав и экологическая характеристика стафилинид Coleoptera, Staphylinidae) южной тайги среднего Урала // Энтомологическое обозрение. Т. 90. Вып. 1. С. 123–137.
Джонгман Р.Г.Г., Тер-Браак С. Дж.Ф., ван Тонгерен О.Ф.Р., 1999. Анализ данных в экологии сообществ и ландшафтов. М.: Российская академия сельскохозяйственных наук. 306 с.
Жантиев Р.Д., 2009. Экология и классификация жуков-кожеедов (Coleoptera, Dermestidae) фауны Палеарктики // Зоологический журнал. Т. 88. Вып. 2. С. 176–192.
Лаврукова О.С., Попов В.Л., Лябзина С.Н., Сидорова Н.А., Приходько А.Н., 2017. Изменение температуры трупа в процессе его разложения (экспериментальное исследование) // Судебно-медицинская экспертиза. Т. 60. Вып. 3. С.19–22.
Лябзина С.Н., 2011. Видовой состав и структура комплекса членистоногих-некробионтов Южной Карелии // Ученые записки Петрозаводского государственного университета. Т. 117. С. 10–19.
Лябзина С.Н., Приходько А.Н., Лаврукова О.С., 2016. Метод изучения некрофильных насекомых на трупах крупных животных // Принципы экологии. Вып. 2. С. 91–98.
Лябзина С.Н., Приходько А.Н., Лаврукова О.С., Сиккиля Н.С., 2016а. Эколого-биологические особенности мухи мертвых (Сynomya mortuorum) в Карелии и применение этих данных в судебно-медицинской практике // Ученые записки Петрозаводского государственного университета. Т. 159. С. 82–87.
Марченко М.И., 1992. Влияние климатических факторов на продолжительность биологического разложения трупа насекомыми-некробионтами в условиях Северо-Запада Европейской части России // Энтомологическое обозрение. Т. 63. Вып. 4. С. 557–568.
Минаев Д.М., Пушкин С.В., 2016. Фауна некробионтных жесткокрылых Северного Кавказа // Научно-методический электронный журнал Концепт. Т. 11. С. 321–325.
Тихомирова А.Л., 1973. Морфологические особенности и филогенез стафилинид. М.: Наука. 193 с.
Шитиков В.К., Розенберг Г.С., Зинченко Т.Д., 2003. Количественная гидроэкология: методы системной идентификации. Тольятти: ИЭВБ РАН. 463 с.
Anderson G.S., 2015. Human Decomposition and Forensics // Carrion ecology, evolution, and their applications. Benbow M.E., Tomberlin J.K., Tarone A.M. (Ed.). USA CRC Press: Boca Raton, FL. P. 541–560.
Braack, L.E.O., 1987. Community dynamics of carrion–attendant arthropods in tropical African woodland // Oecologia. V. 72. P. 402–429.
Caballero U., León-Cortés J.L., 2014. Beetle succession and diversity between clothed sun–exposed and shaded pig carrion in a tropical dry forest landscape in Southern Mexico // Forensic Science International. V. 245. P. 143–150.
Cruise A., Hatano E., Watson D.W., Shal C., 2018. Comparison of Techniques for Sampling Adult Necrophilous Insects From Pig Carcasses // Journal of medical entomology. P. 1–8. https://doi.org/10.1093/jme/tjx255
Hammer Ø., Harper D.A. T., Ryan P.D., 2001. PAST: Paleontological statistics software package for education and data analysis // Palaeontologia Electronica. V. 4 (1). 9 p.
Kočárek P., 2003. Decomposition and Coleoptera succession on exposed carrion of small mammal in Opava, the Czech Republic // European Journal of Soil Biology. V. 39. P. 31–45.
Kozminykh V.O., Esynin S.L., 1994. Spectra of ecological groups and the structure of Coleopters necrobiont communities // Russian Entomological Journal. V. 3. P. 75–80
Matuszewski S., 2017. A general approach for postmortem interval based on uniformly distributed and interconnected qualitative indicators // International journal of legal medicine. V. 131 (3). P. 877–884.
Nuorteva P., Hasanen E., 1972. Transfer of mercury from fishes to sarcosaprophagous flies // Annales Zoologici Fennici. V. 9. P. 23–26.
Smith K.G.V., 1986. A manual of forensic entomology. London, Trustees of the British Museum. 205 p.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал