

Зоологический журнал, 2019, T. 98, № 6, стр. 691-705
Влияние “нового” хищника – хохлатого орла (Nisaëtus nipalensis) – на выбор мест отдыха кабаргой (Moschus moschiferus)
В. А. Зайцев *
Институт проблем экологии и эволюции им. А.Н. Северцова РАН
117071 Москва, Россия
* E-mail: zvit09@mail.ru
Поступила в редакцию 12.07.2018
После доработки 28.11.2018
Принята к публикации 29.11.2018
Аннотация
С 1975 г. на стационарных участках в бассейнах двух рек Сихотэ-Алинского заповедника использовали тропления и визуальные наблюдения за привыкшими и непривыкшими к человеку особями кабарги. Определено расположение 1920 лежек кабарги в среде маскирующих и защищающих зверя объектов. Выделены паттерны расположения лежек разных половозрастных групп кабарги. При схожем составе наземных и пернатых хищников и сходных условиях в бассейнах этих рек в бассейне одной из них регулярно с 1967–1975 г. регистрировали хохлатого орла – нового редкого вида для центрального Сихотэ-Алиня. Описаны эпизоды охоты и добычи орлом кабарги. На участке присутствия орла особи кабарги разного пола и возраста выбирали более защищенные от нападения сверху лежки. Способы охоты хохлатого орла, который использует подкарауливание и полет в пологе леса, отличаются от способов охоты других хищников в многовидовой системе “хищник–жертва”. Это обусловило появление в поведении кабарги способов, обеспечивающих ее маскировку и защиту от нападений пернатого хищника. При высокой бдительности кабарги к сигналам птиц, особенно хищных птиц, в поведении этого копытного, согласованного с разными стратегиями поиска хищниками жертвы, сохранились паттерны архетипа поведения, активизирующиеся при вселении “нового” пернатого хищника. Среди сигналов, которые способны указать жертве на появление “нового” хищника, отмечен свистящий клекот орла. Влияние крупных дневных хищных птиц и сов в эволюции, вероятно, было существенным фактором, определившим заселение кабаргой и трагулидами (Tragulidae) густых зарослей.
В центральном Сихотэ-Алине кабарга представляет собой обычную добычу хищных зверей: харзы (Martes flavigula), рыси (Lynx lynx), реже соболя (M. zibellina), тигра (Panthera tigris altaica), бурого (Ursus arctos) и гималайского (U. thibetanus) медведей (Капланов, 1948; Матюшкин, 1974; Бромлей, Кучеренко, 1983; Зайцев, 1991; Олейников. Зайцев, 2014; Середкин и др., 2017). Медведи и соболь обычно используют останки кабарог, добытых другими хищниками. Такие случаи отмечены и для других хищников, в частности, тигра. Арсеньев (1951) неоднократно отмечал преследование кабарги росомахой (Gulo gulo). Этот хищник обычно охотится за кабаргой в Сибири (Устинов, 1978; Туманов, Кожечкин, 2012). Однако в центральном Сихотэ-Алине в настоящее время росомаха очень редка. Случаев, когда кабаргу добывала лисица (Vulpes vulpes), не часто появляющаяся в местах обитания кабарги Сихотэ-Алинского заповедника, и дальневосточный лесной кот (Prionailurus bengalensis euptilurus. syn. P. euptilura), не выявлено. В течение нескольких десятилетий редко (с 1962 по 1981 гг. 8 случаев) отмечали охоту на кабаргу волка (Canis lupus) (Зайцев, 1991).
Кабарга присутствует и в добыче пернатых хищников, в частности беркута (Aquila chrysaëtus) (Зайцев, 1991; Елсуков, 2013). Беркут и орланы (Haliaëtus albicilla, H. pelagicus) зимой обычно занимаются поиском падали. Тем не менее, в дубовом лесу Лазовского заповедника известна добыча беркутом самки пятнистого оленя (Cervus nippon) (Kerley, Slaght, 2013), гораздо более крупной жертвы, чем кабарга. Среди крупных хищных птиц, способных добыть взрослую кабаргу, только хохлатый орел – редкий в заповеднике и в Приморском крае вид (Елсуков, 1977, 2006, 2013; Нечаев, Горчаков, 2001) – весь год встречается в местах обитания кабарги в горных хвойных и смешанных лесах.
С начала исследований в 2004 г. на стационарном участке в бассейне р. Таежная автор обратил внимание на то, что кабарга выбирала более защищенные места для лежки в сравнении с другими обследованными участками. Встречи хохлатого орла недалеко от лежащих кабарог и у останков жертв, нападение этого хищника на кабаргу дали основание для объяснения данных различий влиянием этого хищника. Задача статьи состоит в: характеристике различий в выборе кабаргой мест расположения лежек в связи с влиянием хохлатого орла. Данная задача направлена на решение проблемы соответствия тактики поведения потенциальной жертвы стратегии охоты “нового” хищника.
МАТЕРИАЛ И МЕТОДИКА
Исследования проведены в Сихотэ-Алинском заповеднике и на прилегающих территориях, в основном, в бассейне среднего течения р. Серебрянка на стационарном участке “Зимовейный”, дополнительно на соседних площадях: “Поднебесный”, “Серебряный” (исследования с 1975 г.). Второй основной участок – “Нечетовский”, на котором исследования ведутся, в основном, с 2004 г., как и дополнительные (“Перевальный” и “Водораздельный” – общая территория “Таежный”) расположены в 40–60 км у северо-восточной границы заповедника в бассейне р. Таежная (рис. 1).
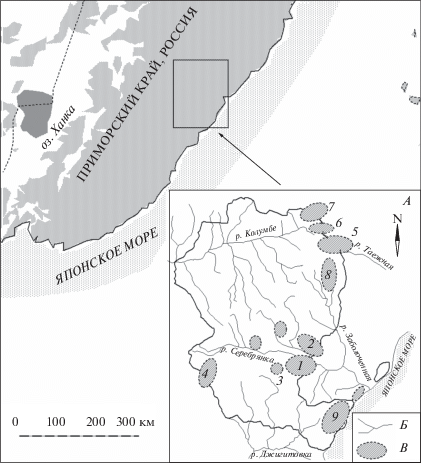
Рис. 1.
Основные стационарные участки в Сихотэ-Алинском заповеднике для исследования экологии кабарги (А), Б – основные реки и ключи, В – участки исследований кабарги, 1 – “Зимовейный”, 2 – “ключ Серебряный и Третий”, 3 – “Поднебесный”, 4 – “Кабаний”, 5 – “Нечетовский”, 6 – “Перевальный”, 7 – “Водораздельный”, 8 – “Солонцовый”, 9 – урочища Благодатное и Голубичное у побережья моря.
Абсолютные высоты на участке Зимовейный составляют 1222–1247 м над ур. м., на участке Нечетовский – 1160.6 м над ур. м. Диапазон высот от вершин водоразделов до русел ключей достигает почти 280–350 м, как и на Нечетовском участке в разных местах до 160–350 м. Зимние температуры воздуха в бассейне р. Серебрянка, обычно, не опускались ниже –18…–20°С, в январе и феврале преобладали температуры до–5…–15°С при обычном муссонном ветре с материка, создающим суровые условия зимовки. На участках в бассейне р. Таежная в январе и в феврале к 7–9 часам утра температура воздуха нередко опускалась до –24…–28°С при обычно маловетренной и безветренной погоде.
Преобладают горные и долинные спелые кедровники (Pinus koraiensis) средней и северной климатической фации (Васильев, Флягина, 2006) сложного породного состава, многоярусной вертикальной и мозаичной горизонтальной структуры, с сомкнутыми кронами древостоев верхних ярусов. Кедровники распространены до высот 600–800 м над ур. м., сменяясь выше (до 1100 м) пихтово-еловыми лесами, с высот 1100 м – каменноберезняками (Betula ermanii), с 1200 м – кедровостланниковыми (Pinus pumila) ассоциациями и с 1400 м – горными тундрами (Шеметова, 1975; Васильев, Флягина, 2006). Сочетание участков высокоствольного леса “паркового” вида – кедровников с дубом (Quercus mongolica) на южных склонах, кедровников с елью (Picea jezoensis, P. koraiensis), пихтой (Abies nephrolepis), лиственницей (Larix cajánderi) на северных склонах и зарослей подроста пихты разного возраста в пологе леса, участков ветровала – создает благоприятные условия для кабарги и хохлатого орла. “Парковые” хвойные, хвойно-широколиственные леса относятся к основным местообитаниям хохлатого орла в Приморском крае (Горчаков, Нечаев, 1994; Нечаев, Харченко, 2012). Пихта, которая в кедровниках обычно выходит во второй древесный ярус и дает обильный подрост, представляет собой наиболее характерный признак местообитаний кабарги (Зайцев, 1991). Подрост пихты на многих участках склонов и в долине заполняет нижние и средние древесные пологи, леса на северных и северо-восточных склонах, нижние и средние части южных склонов и лощины. В обычные годы количество ветровала деревьев верхних ярусов, особенно пихты и елей, достигало от 6.5 до 18 деревьев на 1 га на Зимовейном (Зайцев, 1991) и 4.5–19 на Нечетовском, но местами было больше.
В 1970–1980 гг. и 2004–2008 гг. обычные плотности населения кабарги на участках в бассейне р. Серебрянка, учтенные специальными методами (Зайцев и др., 1988; Зайцев и др., 2013), составляли 11–24 особей на 10 км2. Численность кабарги постепенно снижалась с 1970 гг. В 2012 г. на участке Зимовейный, где вели основные работы, плотность уже не превысила 1–3 особей на 10 км2. На основной площади участка Нечетовский в 2004–2015 гг. она достигала 14–18 особей, но на более обширных территориях с площадями хвойного леса, почти не заселенными кабаргой, по данным учета экскрементов весной, к 2012–2014 гг. составляла всего 3.8–4.6 особей на 10 км2 (Maksimova et al., 2015).
Сбор и анализ данных. Мы применяли многосуточные и фрагментарные тропления кабарог (Зайцев, 1991; Zaitsev et al., 2015). С 1975 г. использованы данные о лежках 22 привыкших к человеку особей. За ними по следам и визуально следили обычно в течение нескольких лет. Учтены данные встреч, троплений 119 непривыкших особей, у которых по внешнему виду и следовым признакам (Зайцев, 1991) были определены пол и возрастная группа. Встречи лежек на маршрутах (более 200) использованы в качестве дополнительных, не включенных в статистическую обработку.
При описании местообитаний использовали показатели проективного покрытия древесных ярусов (в процентах или долях от 1) по 10-балльной шкале (Воронов, 1973). Отмечали особенности устройства лежек кабарги в среде ближних (в радиусе не далее 3–10 м) ориентиров, выделяли варианты их расположения: на буграх или в углублениях, у гребня водораздела, на стволах и комлях упавших деревьев, на скалах или рядом с ними, у ствола дерева, пня, в покопках и на лежках других зверей: изюбрей (Cervus elaphus xanthopygus), косуль (Capreolus pygargus), кабанов (Sus scrofa), очень редко тигра, в зарослях хвойного подроста и тонкомера пихты, под стволами и комлями упавших деревьев (рис. 2). Часть этих мест зарисовывали или фотографировали. Вначале автор на ставил задачу определить эффективность нападения хищных птиц. Однако описания места положения лежки, позволили выполнить данную оценку.
Рис. 2.
Основные варианты, паттерны расположения лежек кабарги (подписи в списке паттернов раздела “Варианты расположения лежек кабарги”).

С использованием выделенных образцов расположения лежки в среде маскирующих и защищающих кабаргу объектов (рис. 2) при троплении с ноября до апреля проанализировано положение 1469 лежек особей определенного пола и возрастной группы на участках в бассейне р. Серебрянка и 451 – в бассейне р. Таежная. 89 лежек встречено на маршрутах в бесснежное время года, когда факт встречи был установлен при расположении лежки на влажной почве у ключа, при встрече кабарги (в зарослях травы, под каменной нишей и др.). Кабарга летом нередко ложится на сухом участке леса без травостоя, что затрудняло поиск таких лежек. Соответственно, статистическое распределение вариантов их расположения было неполным.
Нередко лежки располагались в комплексе выделяемых форм рельефа и объектов растительности: на бугре и в густом подросте, на бугре у основания крупного дерева, в нише вывороченного комля дерева и под его стволом и т.д. В этих случаях паттерн обозначен по основному признаку среды, маскирующему и защищающему кабаргу. То есть, соответственно, в хвойном подросте, у дерева (менее 0.4 м от него), в нише корней комля дерева и т.д. Если кабарга ложилась далее 0.5 м от ближайшего дерева первого, второго древесных ярусов или ствола крупного подроста, зарослей тонкомера пихты, то выделялся другой паттерн – 4 на рис. 2.
Полная оценка защищенности кабарги от обнаружения и нападения пернатого хищника, такого как хохлатый орел с маневренным полетом в пологе леса (Елсуков, 2006, 2013; данные автора), затруднена. Случаи нападения и присутствия орла поблизости от лежащей кабарги были тщательно изучены. Эти наблюдения позволили выделить особенности мест расположения лежек, которые по-разному маскируют и защищают кабаргу от нападения пернатого хищника.
Способы статистической обработки. Метод тропления определенных особей представляет собой выборочный способ сбора данных. Поэтому для анализа статистических данных сведения о лежках классифицирована с учетом пола и возрастной группы кабарог: самцы, начиная с возраста одного года жизни и старше, самки, также начиная с этого возраста, молодые особи – сеголетки обоего пола.
Сравнение статистических распределений образцов расположения лежек кабарог разного пола и возраста, на разных участках проведено по χ2. При этом такое сравнение не характеризует отличий в долях или процентах каждого образца расположения лежек в местах обитаниях, в которых хохлатый орел был и не был замечен. Кроме того, при использовании троплений объем выборок для кабарог разного пола и возраста различался, как и объем для разных местообитаний. В связи с этим учтены рекомендации Зар (Zar, 1999) и Гланца (1999) о применении критерия z с поправкой Йеитса на непрерывность. Для определения различий в долях (процентах) образцов использована формула:
РЕЗУЛЬТАТЫ
Варианты расположения лежек. Кабарга устраивала лежки обычно на достаточно выровненной горизонтальной поверхности, выбирая такие участки и на крутых склонах. Нередко кабарга разгребает место для лежки. Зимой это позволяет ей удобно устроиться в снежной толще. Варианты расположения лежки объединены в семь основных паттернов, которые учитывают возможности маскировки кабарги в среде растительности и форм мезорельефа (рис. 2).
1] Группа паттернов а, в, с включает лежки на комлях упавших деревьев разных видов, в том числе тополе Максимовича (Populus maximowiczii) и ильме (род Ulmus), на лежащих и сломленных стволах деревьев диаметром больше 35 см. На комлях кабарги укладывались на высоте от ≈1 до 3 м. Лежки на сломленных стволах (на высоте от 1.2 до 1.7–2.5 м) наблюдали в снежный период, когда слой снега позволяет кабарге удобно устроиться на круглом стволе. С таких лежек существует неплохая возможность контролировать окрестности. Однако кабарга почти всегда была не защищена от нападения сверху. Устройство части дневных лежек имело в основе комфортную мотивацию, так как кабарга выбирала места, куда проникали лучи солнца, особенно во второй половине февраля и в марте. Самцы старше двух лет в основном выбирали лежки варианта в, но не а и с, как и некоторые самки.
2] а, в. Лежки на скалах, останцах скал и крупных камнях нередко встречались зимой и летом в местах с обычными выходами скал и крупных камней на склонах, а также в дубняках и приморских кедровниках на откосах у моря. В последнем случае нагромождения камней обычно неплохо маскируют кабаргу от “наземного” наблюдателя, но мало защищают от обнаружения пернатым хищником. В лесу участка Зимовейный два выхода скал-камней на поверхность нередко использовали самцы нескольких поколений, ложась рядом со скалой в зарослях пихтового подроста, на крутом склоне.
3]. Лежки на заснеженных или земляных буграх мезорельефа на высоте до 1.5–1.9 м над уровнем снега или земли среди леса, тонкомера пихты нередко в стороне (на расстоянии от 0.4–0.5 м и дальше) от стволов деревьев. Часть лежек данного образца была расположена на узких гребнях водоразделов, оканчивающихся крутым спуском. В спелом хвойном лесу без зарослей подроста кабарга на лежке заметна на склоне сверху, с высоты полета птицы.
4]. Лежки в лесу с выровненной поверхностью мезорельефа, в том числе в покопках (показано на рис. 2), на лежках других зверей или в лощине. Это обычный вариант лежек, устраиваемых на разном расстоянии от ближних деревьев, зарослей подроста.
5]. Лежки на расстоянии не дальше 0,5 м от стволов крупных деревьев, защищающих от нападения сверху, обычно, с одной стороны. Защищенность зависит от расположения лежки относительно ствола и соседних деревьев, наличия кустарника и подроста.
6]. Лежки в хвойном (обычно, пихтовом, еловом) подросте, тонкомере пихты с сомкнутостью крон до 1 и кустарнике: лещины (Corylus sp.), рододендрона (Rhododendron siсhotense), в зарослях багульника (Ledum palústre и др.), под густыми нижними ветвями крупных деревьев, среди бурелома. Такие лежки имеют обычно хорошую защищенность от нападения пернатого хищника сверху, сбоку.
7]. Лежки у комлей вывороченных стволов деревьев, в углублении, нише глубиной до 0.5–0.6 м между корней, под стволами или густыми кронами упавших хвойных деревьев, в их завалах. Присутствие кабарги на такой лежке можно было обнаружить только с очень близкого расстояния. Данные варианты наиболее защищены от нападений с воздуха.
Кабарги многократно использовали часть лежек, в том числе разные особи при смене их поколений. Так, при редких снегопадах на участке Зимовейный в декабре 1979 г. привыкшая к нам самка 13 раз ложилась дважды на прежних лежках из 18, на двух лежках лежала по 3 раза. На участке Нечетовский кабарги обычно многократно использовали лежки под комлями упавших деревьев. Под некоторыми из них кабаргу нередко встречали ежедневно, а пять таких замеченных нами мест четыре особи использовали в течение четырех лет. Зимой 2012–2013 г. сеголеток, обитающий на участке взрослого самца, нередко ложился днем на какую-либо из двух лежек у комлей упавших деревьев, которые в другое время использовали самец и самка.
Во всех случаях кабарга, так или иначе, маскирует свое положение на лежке, располагая ее в лесу с показателями сомкнутости крон хвойных деревьев первого и второго (пихтового) ярусов 0.7–1, среди зарослей подроста пихты, рододендрона. Не было встречено лежек среди редкостойного темнохвойного леса (с сомкнутостью крон ≤0.4) без развитого верхнего, среднего и нижнего хвойного полога. Также не было отмечено, чтобы звери ложились зимой на небольших полянках, в окнах древесного полога, не прикрытых сверху на уровне высокого подроста хвойными деревьями, рододендроном, багульником, днем освещенных прямыми лучами солнца. Если кабарги зимой собирали ночью обильный опад лишайника (роды Usnea, Evernia и др.), листья рододендрона в лиственничнике, то на дневные лежки обычно устраивались по соседству в сомкнутом темнохвойном лесу. У моря в дубняках и березняках, хорошо просматриваемых зимой, кабарга обычно ложится в зарослях рододендрона, у скал.
Летом в жаркую погоду лежки могут располагаться у небольших ключей, среди влажного травостоя. В июле 1975 г. наблюдали, как кабарга ложилась на влажной земле у горного склона, спускаясь каждый раз все ниже и ниже вслед за пересыхающим ключом. На замаскированной высокой травой лежке, у комлей деревьев, в каменной нише у ключа кабарги нередко затаивались. Так, в июле 1975 г. взрослая самка, заметив человека, опустилась на лежку в траве и, подпустив к себе на 2–3 м, ускакала на склон.
Расположение лежек на обследуемых участках. На участке Зимовейный кабарги наиболее часто ложились в лесу на разных возвышениях над поверхностью снега или почвы (табл. 1; образцы (1]+3])), затем у стволов деревьев (5]), в хвойном подросте и тонкомере (6]). Среди паттернов 1] и 3] явно преобладали лежки на буграх, невысоко на стволах упавших деревьев – 26.5% (3]), в сравнении с лежками на высоких комлях и сломанных деревьях – 6.3% (1]). На участке Нечетовский (табл. 2) преобладали лежки образца 6] – в подросте, тонкомере, затем на буграх (3]), реже на упавших стволах, (1] + 3]), наконец на выровненных участках (4]) и в нишах комлей, завалах леса (7]).
Таблица 1.
Соотношения паттернов расположения (процентов от общего N, в скобках – число встреч) лежек кабарги на участках исследований в бассейне р. Серебрянка с осени до весны 1974–1982 и 2004–2008 гг.
| Пол, группа возраста кабарги |
Места расположения лежек согласно выделенным образцам, рис. 3, список | Число лежек, N | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1], 3] на комлях, стволах деревьев, буграх на высоте от 0.4 до 1.2–2 м, в >0.4 м от стволов деревьев | 2] над скалами, крупными камнями в >1.5 м от деревьев | 4] в лесу с выровненной поверхностью в >0.5 м от деревьев | 5] у стволов деревьев, их комлей, пней, (<0.5 м) | 6] в подросте пихты и др., кустарнике, тонкомере пихты | 7] под стволами и кронами, в завалах упавших деревьев, в нишах у корней | ||||||||
| Самцы старше 1 года | 32.80 (245) | 1.47 (11) | 23.56 (176) | 26.10 (195) | 13.39 (99) | 2.68 (20) | 746 | ||||||
| Самки старше 1 года | 29.39 (139) | 0.85 (4) | 13.11 (62) | 24.95 (118) | 26.00 (123) | 5.70 (27) | 473 | ||||||
| Особи <1 года жизни | 24.0 (60) | 0 (0) | 10.80 (27) | 22.80 (57) | 35.20 (88) | 7.20 (18) | 250 | ||||||
| Всего | n | % | n | % | n | % | n | % | N | % | n | % | 1469 |
| 444 | 30.22 | 15 | 1.02 | 265 | 18.04 | 370 | 25.18 | 310 | 21.10 | 65 | 4.44 | 100% | |
Таблица 2.
Соотношения паттернов расположения (число встреч в скобках и процентов от общего N) лежек кабарги на участках исследований в бассейне р. Таежная с осени до весны в 2004–2015 гг.
| Пол, группа возраста кабарги | Места расположения лежек согласно выделенным образцам (рис. 3, список в тексте) | Число лежек, N | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1], 3] на комлях, стволах деревьев, пнях, буграх на высоте от 0.4 до 1.2–2 м, в >0.5 м от стволов деревьев | 2] на скалах, крупных камнях в >1–1.5 м от деревьев | 4] в лесу с выровненной поверхностью в >0.5 м от деревьев | 5] у стволов (<0.5 м), деревьев, комлей, пней | 6] в подросте пихты и др., кустарнике, тонкомере пихты | 7] под стволами, в кронах, завалах деревьев, в нишах у корней | ||||||||
| Самцы, старше 1 года | 11.68 (27) | 0 (0) | 17.75 (41) | 16.02 (37) | 41.13 (95) | 13.42 (31) | 231 | ||||||
| Самки старше 1 года | 20.80 (36) | 0 (0) | 12.14 (21) | 15.03 (26) | 39.31 (68) | 12.72 (22) | 173 | ||||||
| Особи <1 года жизни | 29.79 (14) | 0 (0) | 17.02 (8) | 4.26 (2) | 34.04 (16) | 14.89 (7) | 47 | ||||||
| Всего | n | % | n | % | n | % | n | % | n | % | n | % | 451 |
| 77 | 17.07 | 0 | 0 | 70 | 15.52 | 65 | 14.41 | 179 | 39.68 | 60 | 13.32 | 100% | |
Распределения процентов образцов лежек 1]–7] (табл. 1) на участках в бассейне рек Серебрянка и Таежная существенно различались для самцов: χ2 = 59.33, p < 0.001; самок: χ2 = 18.55, p = 0.0023; сеголетков: χ2 = 88,10, p < 0,001; здесь и далее при df = 5. В бассейне р. Серебрянка самцы выбирали, во многом, особые места для лежек в сравнении с самками (χ2 = 40.0, р < 0.001) и молодыми особями (χ2 = 52.64, р = < 0.001). На участке в бассейне р. Таежная (табл. 2) похожие различия выявлены для самцов и сеголетков: χ2 = 45.12, p < 0.001; для самок и сеголетков: χ2 = 32.43, p < 0.001. Однако выбор лежек самцами и самками на участке у р. Таежная имел большее сходство: χ2 = 6.78, p = 0.238. При этом следует учесть, что часть лежек неоднократно использовалась особями разного пола и возраста.
На участках Зимовейный и Поднебесный в 1974–1983 гг. 48.3% (709) из 1469 лежек образцов 1], 3] и 4] на буграх, стволах деревьев и на выровненной поверхности в лесу было расположено не ближе 0.5 м (в среднем не ближе 0.87 м; Me = 0.8, n = 241) от деревьев и их комлей, исключая редкий мелкий (диаметром до 1–2 см) лиственный подрост и кустарник, почти не маскирующий кабаргу. Из этого числа лежек 10% располагалось от 1 до 2.5–3 м и очень редко до 5 м от крупных деревьев, зарослей хвойного подроста. Большинство (59 из 71) таких лежек принадлежало взрослым (с 2–3 лет жизни) самцам.
Различие в процентах образцов лежек в бассейнах двух рек оказалось в основном существенным (p < 0.05) (табл. 3). Самцы и самки на Нечетовском участке гораздо реже ложились на упавших стволах, буграх, т.е. на возвышенных над поверхностью снега и почвы местах, чем самцы и самки на Зимовейном. Кабарги на Нечетовском явно предпочитали более защищенные лежки с лучшей маскировкой, самцы чаще ложились в зарослях хвойного подроста (образец 6]), самки – в подросте, у деревьев, в завалах деревьев (образцы 5], 6] и 7], (6] + 7])). Показатель z, характеризующий величину различия, для обобщенного образца (6] + 7]) лежек самок несколько меньше, чем для самцов. Небольшие различия выявлены для сеголетков обоего пола в процентах лежек на возвышенных местах (1]), в подросте и тонкомере пихты (6]), а также и для паттерна (6] + 7]). Сеголетки на Нечетовском чаще ложились у стволов, пней и комлей деревьев (5]).
Таблица 3.
Показатели статистического различия образцов расположения лежек кабарги на участках в бассейнах рек Серебрянка и Таежная по данным табл. 1 и 2
| Показатели | Сравниваемые диады процентов паттернов лежек | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| самцов | самок | молодых особей | |||||||||||||
| 1] | 5] | 6] | 7] | 6] + 7] | 1] | 5] | 6] | 7] | 6] + 7] | 1] | 5] | 6] | 7] | 6] + 7] | |
| $\hat {P}$ | 27.84 | 23.75 | 19.86 | 5.22 | 25.08 | 26.84 | 22.09 | 29.29 | 7.52 | 21.47 | 24.92 | 27.95 | 35.02 | 8.42 | 43.43 |
| z | 2.32 | 1.32 | 4.84 | 1.68 | 9.97 | 2.18 | 2.69 | 3.29 | 3.0 | 5.58 | 0.83 | 2.59 | 0.15 | 1.74 | 0.827 |
| р< | 0,02 | >0.05 | 0.001 | >0.05 | 0.001 | 0.05 | 0.01 | 0.001 | 0.005 | 0.001 | >0.05 | 0.01 | >0.05 | >0.05 | >0.05 |
Данные табл. 1 и 2, однако, не показывают одну существенную особенность. В 55% случаев для паттерна 7] самцы кабарги, в 72.7% самки и 71% молодые особи на Нечетовском участке устраивались под комлями упавших деревьев первого и второго ярусов, в нишах сводов корней. На “Зимовейном” таких случаев у всех кабарог не наблюдали. С другой стороны, на Нечетовском участке не было замечено, чтобы кабарги ложились на комлях и стволах упавших и сломанных деревьев на высоте более 0.5 м над поверхностью снега. Это относится и к сеголеткам, которые на Зимовейном в сравнении с Нечетовским нередко выбирали лежку на комле и упавшем стволе дерева на высоте 1–3 м.
На Зимовейном из 388 лежек, на которых кабарги находились в светлое время суток, 102 (26.34%) лежки соответствовали образцу 6] и 26 (6.70%) лежек – образцу 7]. Различия с долей ночных лежек (17.95% для образца 6] и 3.36% для 7] от 1159) существенны при$\quad\hat {P}$ = 20, z = 3.37, p < 0.01 для образца 6] и $\hat {P}$ = 5.88, z = 2.40, p < 0.02 для 7]. На участках по р. Таежной к образцам 6] и 7] относилось, соответственно, 49 (43.61%) и 23 (20.54%) из 112 дневных лежек. Различие с долей ночных лежек (38.35% для 6] и 10.91% для 7] от 339) значимо для 7]: $\hat {P}$ = 13.30, z = 2.61, p < 0.01. Однако оно менее существенно для образца 6]: $\hat {P}$ = 9.69; z = 0.99, p > 0.05.
Виды плотоядных, способных к нападению на кабаргу, и реагирование кабарги на хищных птиц. Харза и рысь – наиболее обычные хищники, добывающие кабаргу, особенно при повышенной их численности в 1970–1980 гг. В 2000 гг. мы отмечали падение численности харзы: в местах обитания кабарги на Зимовеном регистрировали 1–3 следа за зиму. Такая же ситуация наблюдалась и на Нечетовском участке, где только зимой 2012–2013 гг. было прослежено две охоты харз за кабаргой. Редко в 2000 гг. встречали и следы рысей, которые эпизодически заходили в горные хвойные леса стационаров. Лишь иногда рысь несколько суток обследовала горные склоны, например, в феврале 2012 г. трое суток на участке Нечетовский. В хвойных лесах в 1970–1980 гг. останки кабарги встречались в 51.3% экскрементов (n = 39) рыси. Но в 2000–2010 гг. при падении численности кабарги и зайца-беляка (Lepus timidus) рыси чаще добывали косулю, расселившуюся вдоль рек почти до хребта Сихотэ-Алинь. Из 7 жертв рыси, найденных в бассейне р. Серебрянка в 2004–2008 гг., 5 принадлежали косуле, тогда как в 1975–1981 гг. – 2 из 11 (χ2 = 5.95, p < 0.01).
Обычен на данных участках соболь. Известен случай добычи этим хищником молодой самки кабарги зимой на охотничьем участке по кл. Климкин в пределах стационара Нечетовский (сообщение А.Д. Сайко). В отличие от Зимовейного, где следы дальневосточного лесного кота были встречены всего два раза, на Нечетовском участке с 2004 г., особенно в 2010 гг. встречи этого вида были нередкими. В феврале и марте коты часто проходили по дороге вдоль границы заповедника, метили мочой комли снега у обочин. Следы котов встречали в высокоствольных кедровниках с дубом, лещиной (р. Corylus) на водоразделах. Здесь коты охотились за мышевидными грызунами, привлеченными опавшим желудем дуба. Волк, росомаха и лисица появлялись в окрестностях наших участков гораздо реже и не заходили в основные места обитания кабарги.
С 1975 г. в зимний период в центральной части заповедника показатель встреч орлана белохвоста не превышал 1–3 особи и белоплечего орлана – 0.3–0.7 особей на 100 км маршрута вдоль рек (всего 1320 км). Беркут встречался еще реже: одна птица на ≈200 км, и на Нечетовском участке зимой мы не отмечали этого хищника. Однако по сообщению охотников, в лесу ниже по течению реки и в бассейне соседней р. Кема известны редкие случаи добычи беркутом кабарги. Беркут и орланы, занятые в основном поиском падали, залетали зимой вдоль рек обычно не далее 30–60 км от моря. С 1975 г. в бассейне р. Серебрянка беркут, пролетающий над горным кедровником, был замечен только три раза. У побережья моря и в прилегающих дубняках, где показатель встреч орланов был 1.3–2 особи белохвостого и 0.5–1.2 белоплечего орланов на 10 км (на 235 км) в 1978 г. наблюдали, как два орлана белохвоста пытались напасть на кабаргу, стоящую на отстое прибрежных скал.
Весь год на участках исследований, в том числе по 1–7 встреч зимой регистрировали ястреба-тетеревятника (Accipiter gentilis) и более многочисленную длиннохвостую неясыть (Strix uralensis). Плотность населения неясыти, определенные по имитации брачного крика самцов и самок совы, весной на Зимовейном в 1970–1980 гг. составляла 5–7 гнездящихся пар на 10 км2, на Нечетовском в 2000 гг. – 2–4 пары. Эти птицы могут быть опасными для молодых особей, о нападении их на взрослых кабарог не известно. Среди добычи совы присутствовал заяц-беляк, однако мы наблюдали неудачные попытки охоты сов и на других сравнительно крупных животных, например на фокстерьера.
В бассейне р. Серебрянка хохлатого орла весь период исследований мы не регистрировали. Присутствие этого хищника в бассейне р. Таежная на промежутке 9 км вдоль реки в 2005–2015 гг. определено 12 раз, из них девять раз по свистящему громкому “клекоту”. Орел неоднократно был замечен охотниками и в соседнем бассейне р. Кема, несколько раз был отловлен в капканы, расставленные на пушных зверей. В сезоны 2012–2013 гг. и 2014–2015 гг. на ≈9.5 км2 участка Нечетовский орла мы встречали пять раз. Этот скрытный хищник только три раза был замечен над лесом. 7.03.2005 г. орел догнал и схватил на лету кедровку (Nucifraga caryocatactes) из пролетающей низко над лесом стаи. 20.03.2013 г. в 15 ч автор имитировал крик орла и, перекликаясь с ним, приблизился с расстояния 1.5 км на два десятка метров к птице, сидящей на дереве у гребня водораздела в ≈150 м от места, где в густом подросте пихты лежала одна из прослеживаемых нами кабарог. Орел, взлетев, с криком парил невысоко над кедровником долины ключа, быстро совершая неширокие круги над местом, где автор закончил перекличку с орлом. Птица по крику была замечена у этого места и на следующий день. 29.12.2014 г. хищник был отмечен по крику, затем визуально у гребня водораздела в кедровнике с дубом над комлем упавшего дерева, под которыми устроился на лежку самец кабарги, за которым мы следили троплением и по радиопеленгу. После взлета орел сразу улетел в полог леса. Также и в других случаях при приближении наблюдателя к кричащей птице, она не вылетала из полога высокого леса. В четырех из 11 случаев, когда орел издавал крик, данное место располагалось вблизи лежащей кабарги, один раз у места предыдущей добычи орлом длиннохвостой неясыти. В феврале 2012 г. кричащего орла заметили в лиственничнике нижней части склона, где в это время обитала самка кабарги, и в феврале 2013 г. были найдены останки кабарги, добытой этим хищником (случай описан в следующем разделе). В мае 2011 г. в 3.5 км выше по течению реки в небольшом распадке крик орла слышали у места, где затем были обнаружены останки кабарги. Среди других жертв этого хищника по следам лап, когтей и отпечаткам перьев на снегу отмечена длиннохвостая неясыть, два раза белка (Sciurus vulgaris). В апреле 2012 г. орел был замечен у останков косули, добытой зимой рысью.
Кабарги при криках орла затаивались и не покидали защищенные лежки. При появлении или пролете над лесом хищных птиц других видов привыкшие к нам кабарги выявляли повышенную настороженность. Так, днем 5.03.1981 г. взрослая самка собирала лишайник с поверхности снега. При появлении у нее за спиной и медленном пролете ястреба-тетеревятника на высоте около 20 м в долине над спелым кедровником с тополем Максимовича кабарга (еще до обнаружения птицы человеком) замерла на месте, как бы “напряглась”, следя за ястребом поворотом головы. После пролета хищника кабарга стояла еще около 10 с, а затем возобновила сбор пищи. В другом случае (14.03.1978 г.) пролет беркута на высоте 50–70 м над кедровником также вызвал настороженность самца кабарги, который, оставаясь неподвижным, следил за птицей поворотом головы.
Эпизод нападения хохлатого орла на кабаргу. Присутствие орла у останков кабарги отмечено на Нечетовском участке 8.05.2012 г. и 6.02.2013 г. В первом случае хохлатый орел по голосу был обнаружен в узком распадке вблизи днища долины реки, где были найдены шкура, кости и другие останки кабарги. Осталось неизвестным, была ли добыта кабарга орлом или другим хищником. Во втором эпизоде, произошедшем после небольшого снегопада 2.02.2013 г., хищничество хохлатого орла было установлено.
Орел напал на взрослую самку кабарги, которая несколько суток придерживалась окрестностей седловины водораздела. После очередного сбора пищи в небольшом пятне редкого лиственничника у подножия южного склона кабарга устроилась на лежку на расстоянии в 0.6 м от крупного кедра и 3–3.5 м от густого подроста пихты (1 на рис. 3а). Отпечатки маховых перьев в этом месте указывают на то, что на лежащую кабаргу орел напал сзади и с правой стороны, облетев ствол кедра. Далее до пункта 3 кабарга вместе с орлом ползла рывками, пыталась приподняться то на передние, то на задние ноги и оставляла широкую борозду в снегу. У пункта 2, где отпечатки крыльев орла выделяются особенно четко, кабарга лежала, но затем, пытаясь встать, поползла дальше. В метре от пункта 2 было обнаружено перо орла. Однако далее кабарга ползла уже без хищника за спиной: отпечатки перьев на снегу отсутствовали. Жертва подползла к сухому стволу небольшой упавшей пихты, где и была обнаружена 6.02.2013 г. в углублении снега диаметром до 1 м. Орел подлетал к этому месту, но полностью воспользоваться добычей (поклевы у бедра) ему не пришлось. Жертву нашел соболь, который устроил три норы рядом с жертвой (3 на рис. 3а). Присутствовали и следы еще одного соболя, подбежавшего со склона.
Рис. 3.
Схема места нападения хохлатого орла на кабаргу и повреждения, нанесенные орлом кабарге; а: 1 – лежка кабарги – место нападения орла (следы маховых перьев на снегу); 2 – лежка-волок на пути перемещения кабарги, следы крыльев; 3 – положение кабарги-жертвы в ямке у сломанной сухой пихты, норы соболя в снежной толще; б: 1 – шрам на надглазничном углублении с повреждением правого глаза и гематома; 2 – шрам ниже глазницы; 3 – длинный шрам-разрез когтем на шее с повреждением пищевода и рядом гематома; 4 – следы-порезы шкуры когтями одной лапы орла; 5, 6, 7 – гематомы и небольшие разрывы, порезы шкуры кабарги.
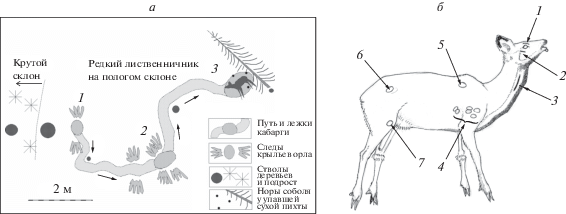
Повреждения, нанесенные орлом: порезы, проколы когтями, гематомы обнаружены с правой стороны тела кабарги (рис. 3б). Повреждения, приведшие к гибели животного, располагались на правой стороне головы и шеи (1–3). Их расположение указывает, что при нападении орел схватил кабаргу левой лапой за голову и верхнюю часть шеи, правой лапой – за область плеча (4). Длинный (9.5 см) порез-разрыв шкуры и пищевода 3 был нанесен задним когтем, вероятно, при резком повороте головы жертвы. Когти орла проникли через правое надглазничное углубление (1) черепа и глаз в глазницу. Жертва погибла не сразу, несколько раз переворачивалась и раскопала яму в подтаявшем снегу.
У жертвы хохлатый орел и падальщики: две сойки (Garrulus glandarius); гаички (Parus (Poecile) montanus), в основном соболь, объели мясо с предплечья и плеча, лопатки, с голени левой стороны тела (кабарга лежала на правом боку), повредили брюшную полость. Но внутренние органы и эмбрионы были целыми.
ОБСУЖДЕНИЕ
Поведение, обеспечивающее безопасность кабарги, включает ориентировочные реакции на густые, прикрытые сверху и во всем вертикальном пологе участки леса. Данная ориентация преобладает в разное время суток, при переходах и отдыхе (Зайцев, 1991). В результате кабарга обычно устраивает лежки в хвойном подросте, у деревьев, упавших их стволов (50.7–67.4% на обследованных участках). Кабарга явно оценивает возможности своей маскировки, затаиваясь на лежке, замирая при перемещении у стволов деревьев, на фоне пней, комлей упавших деревьев. Большое количество (от 17.1 до 30.2%) лежек располагалось на возвышенных местах (буграх, комлях упавших деревьев и др.), что свидетельствует также о значении контроля окрестностей. На скалах и камнях отмечено небольшое число лежек (до 1.02%; табл. 1, 2). Это связано с особенностью распределения кабарги в местах обитания Сихотэ-Алиня. Большие плотности ее населения не приурочены к участкам со скалами и их останцами (Матюшкин, 1974, 2005; Зайцев, 1991). Исключение составляют скалы и обрывы побережья моря, которые кабарга заселяет эпизодически. Тем не менее, вдали от моря положение лежек в местообитаниях кабарги в некоторых случаях выявило связь с редкими скалами.
Маскировка и защищенность кабарги на лежке от хищных птиц оценивалась по прикрытию сверху и с боков ветвями, стволами, сводами корней и др. В выборе лежки кабарги определены половозрастные особенности, заключающиеся в преимущественном выборе молодыми особями и самками более защищенных мест в лесу, чем взрослыми самцами (р < 0.001). Различия установлены также для самок и сеголетков (р < 0.001). Сравнение процентов паттернов расположения лежек (табл. 3) показало, что многие лежки на участке Нечетовский были более эффективно защищены от нападений хищной птицы, чем лежки на Зимовейном и Поднебесных ключах. Значительные различия установлены для самцов. Отличие для самок в показателе z было меньше (табл. 1–3). Сравнительно небольшие различия определены для сеголетков обоего пола, которые, однако, на участке Нечетовский чаще ложились у стволов, комлей упавших деревьев. Подобное сходство в составе паттернов лежек сеголетков, частично у самок, обусловлено, вероятно, эволюционно предопределенным стереотипом выбора. Прежде всего, это относится к сеголеткам, для которых особенно характерно затаивание при маскирующей окраске (Зайцев, 1991).
Особенно заметные различия заключаются в образцах устройства лежек под комлями и стволами упавших деревьев, в нишах между корней (рис. 2). На Зимовейном лежек в нишах между корнями мы не встречали вовсе, однако были обычны лежки на стволах сломанных деревьев, валежинах, комлях на высотах больше 0.5 м и до 3 м над снегом или землей. Показатели плотности населения кабарги на Зимовейном в 1974–2008 г. (11–24 особей на 10 км2) и Нечетовском в 2004–2015 г. (14–18 особей) были схожими. Сходство наблюдали в числе упавших деревьев, которые способны создать комфортные условия на лежках. Выбор лежек под стволами упавших деревьев на участке Нечетовский не был обусловлен только более низкими ночными температурами воздуха зимой. Под комли кабарги ложились и днем при плюсовых температурах в марте, апреле и позднее.
Лежки под комлями и стволами деревьев, среди подроста защищают затаившуюся кабаргу от нападения крупной птицы. В четырех случаях из 11 встреч хохлатый орел был замечен на присаде вблизи кабарог, лежащих в защищенном от нападения сверху месте. Но хищник не совершал нападение. Однако в случае описания 6.02.2013 г. кабарга лежала на расстоянии более 0.5 м от укрытия, что позволило орлу совершить успешное нападение. У птицы возникли явные затруднения при добыче взрослой самки, но она нанесла кабарге смертельные повреждения.
В случае наземного хищника (рысь, харза), следующего вдоль следа кабарги, комли деревьев, у которых лежит кабарга, могут составить препятствие для кабарги, способной выбежать лишь со стороны своего захода на лежку. Хищные звери, включая харзу, используют для поиска жертвы запах и визуальные признаки следа (Зайцев, 1991). Хищник, следующий за кабаргой по ее следу, не совершал нападения вплоть до обнаружения им убегающей жертвы. В результате, кабарга, находясь на лежке, особенно на бугре, стволе упавшего дерева начинала бегство еще до обнаружения ее хищником, особенно таким, как харза. При троплении и во время визуальных наблюдений, кабарги, собирающие пищу, нередко совершали быстрое бегство на короткие расстояния (вплоть до нескольких метров) в ответ на треск веток, падение комков снега, тревожные голоса птиц. Такое поведение согласуется с неожиданным нападением хищников, таких как соболь и, особенно, рысь, которая обычно подкрадывается к жертве (Зайцев, 1983; Олейников, Зайцев, 2014.). Комплекс поведения, обеспечивающего собственную безопасность, у кабарги соответствует в среде густого сомкнутого леса разным стратегиям поиска жертвы хищниками (Зайцев, 1991), в том числе и пернатыми.
Эти результаты согласуются с данными и предположениями о влиянии комплекса хищников на выработку компромиссных стратегий защиты от хищников (Mcintosh, Peckarsky 1999; Relyea 2003; Atwood et al., 2008; Thaker et al., 2011) и гипотезой эволюционного становления поведения жертв (Blumstein, 2006). Выбор кабаргой лежки определяется защитными, комфортными условиями, что отражается и в статистическом распределении паттернов расположения лежек. Не малая доля (53%) защищенных лежек в подросте и под упавшими деревьями позволяет полагать, что население кабарги на участке у р. Таежная оценивает риск нападения хищной птицы, как достаточно высокий.
При схожем составе хищников в бассейне рек Таежная и Серебрянка, исключая лесного кота у р. Таежная, который по категории веса (до 5.2 кг по Юдину, 2015) способен добыть взрослую кабаргу, присутствие хохлатого орла у р. Таежная составляет наиболее вероятную и единственно установленную причину различий в выборе кабаргой мест отдыха. Это тем более реально в аспекте различий оборонительного поведении кабарги на территориях с разным набором и обилием наземных хищников (Зайцев, 1983а; Зайцев, 1991). Устройство лежек на упавших деревьях, их комлях на участках у р. Серебрянка согласуется с почти полным отсутствием этого хищника (орел в бассейне реки встречен Елсуковым (2013) в 1983 г. и 1988 г.), редкими залетами зимой беркута. Тем не менее, кабарги и в этих условиях сохраняли повышенную осторожность по отношению к хищным птицам разных видов, следя за их пролетом над лесом.
Хохлатый орел в 1960 гг. относился к редким залетным видам Приморья и о-ва Сахалин (Иванов, Штегман, 1964; Горчаков, Нечаев, 1994, 1998). В настоящий период эта птица на юге Приморского края более обычна (Нечаев, Горчаков, 2001; Нечаев, 2005; Нечаев, Гамова, 2009; Нечаев, Харченко, 2012) и с 1967 г. регистрировалась в разные сезоны года в Сихотэ-Алинском заповеднике, где возможно ее гнездование (Елсуков, 1977, 2013). Об оседлом образе жизни и вероятном гнездовании свидетельствуют регулярные встречи нами орла в бассейне р. Таежная на протяжении более 10 лет. В 2011–2015 гг. орла встречали почти так же часто, как следы харзы и рыси. Елсуков (2013) встречал орла в окрестностях этого участка еще в 1975 г. Данный автор ссылается на сообщения охотников о трех нападениях орла на кабаргу, а также на белку, норку (Neovison vison), выдру (Lutra lutra) и домашнюю собаку (Canis lupus familiaris) и на случай добычи кабарги беркутом. Эпизод добычи кабарги, приведенный в данной статье, – подтверждение успешной добычи орлом этого копытного. Факты позволяют отнести хохлатого орла к не столь редким хищникам территории на р. Таежная на протяжении уже нескольких десятилетий.
Сразу после неудачного для харзы и рыси преследования, у кабарги увеличивались дистанция бегства, расстояния бегства от одной остановки до другой, менялась форма пути бегства, у привыкших к нам кабарог возрастала дистанция подпускания наблюдателя. Но уже через несколько дней восстанавливались прежние показатели поведения (Зайцев, 1991). Сохранялась устойчивая связь кабарги с темнохвойным лесом. Однако вселение “нового” хищника способно вызвать не только кратковременную и быструю смену показателей бдительности жертв, тактик использования ими фрагментов местообитаний, но и обусловить устойчивое изменение поведения, что показано, например, при реинтродукции гиеновых собак (Lycaon pictus) в местообитания трех видов африканских антилоп (Makin et al., 2018).
Таким образом, реально, что вселение хохлатого орла, регулярно встречаемого в бассейне р. Таежная, несколько десятилетий влияло на поведение потенциальных жертв, что вызвало устойчивые изменения в поведении кабарги. Это влияние отразилось в выборе лежек, защищающих кабаргу в пределах обычных ее местообитаний в темнохвойном лесу. При очень редких залетах беркута в места обитания кабарог и не частых встречах хохлатого орла, многие кабарги, наиболее вероятно, не подвергались нападению хищных птиц все это время. Можно полагать, что вселение хохлатого орла, как хищника с определенной стратегией охоты (выслеживание в пологе леса и подкарауливание), присутствие которого потенциальная жертва устанавливает по особым признакам, активизировало также определенную тактику поведения кабарги, которая уже имелась в эволюционном архетипе ее поведения. Кроме внешнего облика хищника, распознаваемого кабаргой (возможно, по размеру птицы), к таким признакам может относиться свистящий клекот, который орел издавал нередко вблизи лежащих, убитых кабарог и у других жертв. Подобный эффект сигналов разной природы (визуальных, химических и др.), исходящих от “нового” хищника, учитывается и другими авторами в идее эволюционного развития компромиссного поведения жертвы (Blumstein, 2006; Ferrari и др., 2007).
Некоторые перманентные свойства эволюционного архетипа обеспечения собственной безопасности у кабарги выявляются в выборе местообитания в густом лесу и защищенных лежек, особенно, молодыми особями и взрослыми самками в условиях отсутствия хохлатого орла и беркута. Однако влияние хохлатого орла, активного в светлое время суток, на участке у р. Таежная в сравнении с участками у р. Серебрянка, повлекло не только сдвиг частот паттернов выбора лежек у всех половозрастных групп кабарги, но и активизировало паттерны поведения, связанные с выбором лежек образцов 6] и 7] в наименее доступных для пернатых хищников местах. Тем не менее, подобные лежки кабарги использовали и в темное время суток, хотя процент защищенных от нападения хищной птицы лежек в светлое время суток был несколько больше, чем в ночное. Таким образом, сигналы, исходящие от хохлатого орла, активизировали паттерны выбора защищенной лежки, направленные не только на избегание нападения дневного хищника конкретного вида, но и других вероятных хищников, использующих нападение с полета.
Этот вывод согласуются с результатами и гипотезами о том, что сигналы, исходящие от одного хищника, вызывают реагирование жертвы на сигналы целого континуума видов хищников, имеющих некоторое сходство во внешнем облике или других особенностях (Griffin et al., 2001. Stankowich, Coss, 2007; Ferrari et al., 2008; etc.). Высокая бдительность к виду пролетающих хищных птиц и звукам (голосам, шорохам), исходящим от птиц разного размера в кронах деревьев (Зайцев, 1983, 1991), свидетельствует об эволюционно предопределенной оценке кабаргой риска нападения сверху, с полета птицы.
Ареалы горного хохлатого орла и других видов птиц рода Nisaëtus в Гималаях, Юго-восточной Азии (Степанян, 1990; Нечаев, Гамова, 2009; Grimmett et al., 2012) на значительном протяжении совмещаются с ареалами кабарги (восстановленный ареал по: Флеров, 1952) и мелкими видами семейства Tragulidae (роды Tragulus, Moschiola) (Wilson, Mittermeier, 2011). Как и эти копытные, хохлатые орлы рода Nisaëtus, используют полог леса в качестве основной арены жизни (Горчаков, Нечаев, 1994; Нечаев, Харченко, 2012; Галушин и др., 1991; Grimmett et al., 2012). Крупные дневные хищные птицы и совы обычно охотятся на трагулид в тропических лесах и в современную эпоху (Wilson, Mittermeier, 2011).
ЗАКЛЮЧЕНИЕ
Выявленные различия в выборе мест для лежек кабарог разного пола и возрастных групп в среде маскирующих объектов имеют эволюционные предпосылки. Наибольшее число защищенных от нападений сверху лежек учтено на участках в бассейне одной из двух обследованных бассейнов рек (р. Таежная). Однако более частые встречи крупных хищных птиц: беркута и орланов происходили в бассейне другой реки (р. Серебрянка). В результате исследования установлено, что причина различий в соотношении паттернов расположения лежек кабарги на данных участках заключается в обычном присутствии в бассейне р. Таежная хохлатого орла, который охотится, подкарауливая кабаргу на лежке или нападая с полета.
Хохлатый орел – относительно новый редкий вид для центрального Сихотэ-Алиня, встречи с которым происходят эпизодически, и только на определенных участках он встречался более часто. В многовидовой системе “хищник – жертва” появление “нового” хищника с особой тактикой охоты привело к возникновению и особых действий жертвы, обеспечивающих ее маскировку и защищенность от нападений этого хищника. В комплексе избегания встреч с хищниками разных видов и нападений этих хищников, использующих разные стратегии поиска жертвы, у потенциальной добычи сохраняются паттерны архетипа поведения, активизирующиеся при появлении “нового” хищника. В эволюции, вероятно, стратегии и тактики охоты хищных птиц существенным образом влияли на заселение современной кабаргой и трагулидами густых зарослей леса, ограничивающих для данных хищников доступность жертвы.
Список литературы
Арсеньев В.К., 1951. Сквозь тайгу. В горах Сихотэ-Алиня. По материалам экспедиций 1908–1910 гг. и 1927 г. Ред. Кумкес С.Н. М.: Географгиз. 404 с.
Бромлей Г.Ф., Кучеренко С.П., 1983. Копытные юга Дальнего Востока СССР. Ред. Насимович А.А. М.: Наука. 304 с.
Васильев Л.И., Флягина И.А., 2006. Растительный покров // Растительный и животный мир Сихотэ-Алинского заповедника. Отв. ред. Астафьев А.А. Владивосток: изд-во ОАО “Примполиграфкомбинат”. С. 122–219.
Воронов А.Г., 1973. Геоботаника. Ред. Н.А. Соколова. М.: Высшая школа. 384 с.
Галушин В.М., Дроздов Н.Н., Ильичев В.Д., Константинов В.М., Курочкин Е.Н. и др., 1991. Фауна мира: Птицы: Справочник. Ред. Ильичев В.Д. М.: Агропромиздат. 311 с.
Гланц C., 1999. Медико-биологическая статистика. Пер. с англ. Даниловой Ю.А. Ред. Бузикашвили Н.Е., Самойлова Д.В. М.: Практика. 459 с.
Горчаков Г.А., Нечаев В.А., 1994. Хохлатый орел, Spizaetus nipalensis (Hodgson, 1836), (Accipitridae, Aves) – новый гнездящийся вид фауны России // Бюллетень Московского общества испытателей природы. Отд. биол. Т. 99. Вып. 4. С. 15–17.
Горчаков Г.А., Нечаев В.А., 1998. Новые материалы о биологии хохлатого орла в Южном Приморье // Вопросы сохранения ресурсов малоизученных редких животных Севера. М.: ЦНИЛ охотничьего хоз-ва и заповедников. Ч. 1. С. 108–114.
Зайцев В.А., 1983. Оборонительное поведение кабарги (Moschus mоschiferus L.) в Среднем Сихотэ-Алине // Зоологический журнал. Т. 62. Вып. 11. С. 1718–1726.
Зайцев В.А., 1983а. Оборонительное поведение кабарги разных пространственных группировок // Поведение животных в сообществах. М.: Наука. С. 77–78.
Зайцев В.А., 1991. Кабарга Сихотэ-Алиня. Экология и поведение. Ред. Баскин Л.М., Филонов К.П. М.: Наука. 216 с.
Зайцев В.А., Зайцева В.К., Назаров А.А., 1988. Методические указания по учету кабарги. М.: Главное управление охотничьего хозяйства и заповедников. 15 с.
Зайцев В.А., Середкин И.В., Максимова Д.А., Пименова Е.А., Милаковский Б., 2013. Методы учета плотности населения и исследования распределения кабарги (Moschus moschiferus) в местообитаниях Сихотэ-Алиня / Научно-методические основы составления государственного кадастра животного мира республики Казахстан и сопредельных стран. Алматы: РПГ Институт зоологии. С. 73–80.
Елсуков С.В., 1977. О редких птицах северо-востока Приморья // Тез. докладов VII Всесоюзной орнитологической конференция. Киев: Наук. думка. С. 58–59.
Елсуков С.В., 2006. Птицы. Растительный и животный мир Сихотэ-Алинского заповедника. Отв. ред. Астафьев А.А. Владивосток: изд-во ОАО “Примполиграфкомбинат”. С. 272–296.
Елсуков С.В., 2013. Птицы северо-восточного Приморья. Неворобьиные. Ред. Нечаев В.А. Владивосток: Дальнаука. 536 с.
Иванов А.И., Штегман Ю.В., 1964. Краткий определитель птиц СССР. Отв. ред. Павловский Е.Н. М.-Л.: Наука. 528 с.
Капланов Л.Г., 1948. Тигр в Сихотэ-Алине // Тигр. Изюбрь. Лось. Материалы к познанию фауны и флоры СССР. Ред Огнев С.И., Гептнер В.Г. М.: Изд-во Московского общества испытателей природы. Нов. серия. Отдел. зоол. Вып. 14 (29). С. 18–49.
Матюшкин Е.Н., 1974. О взаимоотношениях харзы (Martes flavigula Bodd.) и кабарги (Moschus moschiferus L.) в Среднем Сихотэ-Алине и история формирования их биоценотической связи // Териология. Т. 2. Отв. ред. Воронцов Н.Н. Новосибирск: Наука, Сиб. отд. С. 227–252.
Матюшкин Е.Н., 2005. Избранные труды. Ред. Кулешова Л.В., Михайлов К.Г., Рожнов В.В. М.: Товарищество научных изданий КМК. 660 с.
Нечаев В.А., 2005. Хохлатый орел // Красная книга Приморского края. Животные. Отв. ред. В.А. Костенко. Владивосток: АВК “Апельсин”. С. 239–241.
Нечаев В.А., Гамова Т.В., 2009. Птицы Дальнего Востока России (аннотированный каталог). Отв. ред. Журавлев Ю.Н. Владивосток Дальнаука, 564 с.
Нечаев В.А., Горчаков Г.А., 2001. Хохлатый орел (Spizaetus nipalensis orientalis (Temminck et Schlegel, 1844)). Красная книга Российской Федерации (животные). Отв. ред. Данилов-Данильян В.И. М.: АСТ “Астраль”. С. 432–435.
Нечаев В.А., Харченко В.А., 2012. Современное распространение и особенности восточного хохлатого орла (Spizaetus nipalensis orientalis Temminck et Schlegel, 1844) в России // Вестник Красноярского гос. аграрного университета. № 5. С. 238–244.
Олейников А.Ю., Зайцев В.А., 2014. Охота соболя (Martes zibellina) на кабаргу (Moschus moschiferus) // Бюллетень Московского общества испытателей природы. Отд. Биол. Т. 119. Вып. 6. С. 20–28.
Серёдкин И.В., Зайцев В.А., Петруненко Ю.К., Максимова Д.А., Микелл Д.Г., 2017. Кабарга в питании тигра и медведей на Сихотэ-Алине // Экология. № 4. С. 299–303.
Степанян Л.С., 1990. Конспект орнитологической фауны СССР. Отв. ред. Соколов В.Е. М.: Наука. 728 с.
Туманов И.Л., Кожечкин В.В., 2012. Росомаха Палеарктики. Рец. Семечкин И.В., Виноградов В.В. СПб.: Издательский дом “Бранко”. 320 с.
Устинов С.К., 1978. Кабарга // Крупные хищники и копытные звери. М.: Лесная промышленность. Ред. Калецкий А.А., Руденко Т.А. С. 256–259.
Шеметова Н.С., 1975. Флора и растительность Сихотэ-Алинского государственного заповедника // Флора и растительность прибрежных районов юга Дальнего Востока. Труды Билого-почвенного института ДВНЦ АН СССР. Т. 24. (127). С. 5–85.
Флеров К.К., 1952. Кабарги и олени // Фауна СССР. Млекопитающие. Ред. Новиков Г.А. М.-Л. Изд-во АН СССР. Т. 1. Вып. 2. 256 с.
Юдин В.Г., 2015. Дальневосточный лесной кот. Отв. ред. Пикунов Д.Г. Владивосток: Дальнаука. 443 с.
Atwood T.C., Gese E.M., Kunkel K.E., 2008. Spatial partitioning of predation risk in a multiple predator–multipleprey system // Journal of Wildlife Management. V. 73. № 6. P. 876–884.
Blumstein D.T., 2006. The multipredator hypothesis and the evolutionary persistence of antipredator behavior // Ethology. V. 112. Is. 3. P. 209–217.
Griffin A.S., Evans C.S., Blumstein D.T., 2001. Learning specificity in acquired predator recognition // Animal behaviour. V. 62. P. 577–589
Grimmett R., Inskipp C., Inskipp T., Byers C., 2012. Birds of India, Pakistan, Nepal, Bangladesh, Bhutan, Sri Lanka, and the Maldives (Princeton field guides). (6 ed.). Princeton: Princeton University Press, Wiley. 528 p.
Ferrari M.C.O., Gonzalo A., Messier F., Chivers D.P, 2007. Generalization of learned predator recognition: an experimental test and framework for future studies // Proceedings of the Royal Society B. Biological Sciences. V. 274. P. 1853−1859.
Ferrari M.C.O., Messier F., Chivers D.P., 2008. Can prey exhibit threat-sensitive generalization of predator recognition? Extending the Predator Recognition Continuum Hypothesis // Proceedings of the Royal Society B. Biological Sciences. V. 275. P. 1811–1816.
Kerley L.L., Slaght J.C., 2013. First documented predation of sika deer (Cervus nippon) by golden eagle (Aquila chrysaëtos) in Russian Far East // Journal of Raptor Research. V. 47. № 3. P. 328–330.
Makin D.F., Chamaillé-Jammes S., Shrader A.M., 2018. Changes in feeding behavior and patch use by herbivores in response to the introduction of a new predator // Journal of Mammalogy. V. 99. Is. 2, 3. P. 341–350.
Maksimova D.A., Seryodkin I.V., ZaitsevV.A., 2015. Musk deer (Moschus moschiferus) population density based on pellet group count method in Sikhote-Alin // Achievements in the Life Sciences. № 9. P. 57–60.
Mcintosh A.R., Peckarsky B.L., 1999. Criteria determining behavioural responses to multiple predators by a stream mayfly // Oikos. № 85. Is. 3. P. 554–564.
Relyea R.A., 2003. How prey respond to combined predators: a review and an empirical test // Ecology. V. 84. Is. 7. P. 1827–1839.
Stankowich T., Coss R.G., 2007. The re-emergence of felid camouflage with the decay of predator recognition in deer under relaxed selection // Proceedings of the Royal Society B. Biological Sciences. V. 274. P. 175–182.
Thaker M., Vanak A.T., Owen C.R.,. Ogden M.B, Niemann S.M., Slotow R., 2011. Minimizing predation risk in a landscape of multiple predators: effects on the spatial distribution of African ungulates // Ecology. V. 92. Is. 2. P. 398–407.
Wilson D.E., Mittermeier R.A. (eds), 2011. Handbook of the Mammals of the World. V. 2. Hoofed Mammals. Barselona, Spain: Pub. by Lynx Edicions, Montseny 8. 886 p.
Zaitsev V.A., Seryodkin I.V., Maksimova D.A., Soutyrina S.V., 2015. Study of the musk deer population structure in Sikhote-Alin reserve //Achievements of Life Sciences. V. 9. Is. 2. P. 83–86.
Zar J.H., 1999. Biostatistical analysis (4. еd.). Ryu T., Snavtly S.L (eds). Upper Saddle River, New Jersey: Phentice Yall. 663 p. + Appendix, tables and Graphs
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал